

Population Dynamics of the Olive Fly, Bactrocera oleae (Diptera: Tephritidae), Are Influenced by Different Climates, Seasons, and Pest Management




Abstract
:1. Introduction
- (1)
- To monitor the changes in the OFF populations over space in the period June to October for three seasons, both within each season and among the seasons;
- (2)
- To analyze the changes in OFF populations in relation to previous populations for all years and other climatic variables using a linear mixed-effects model and considering spraying events.
2. Materials and Methods
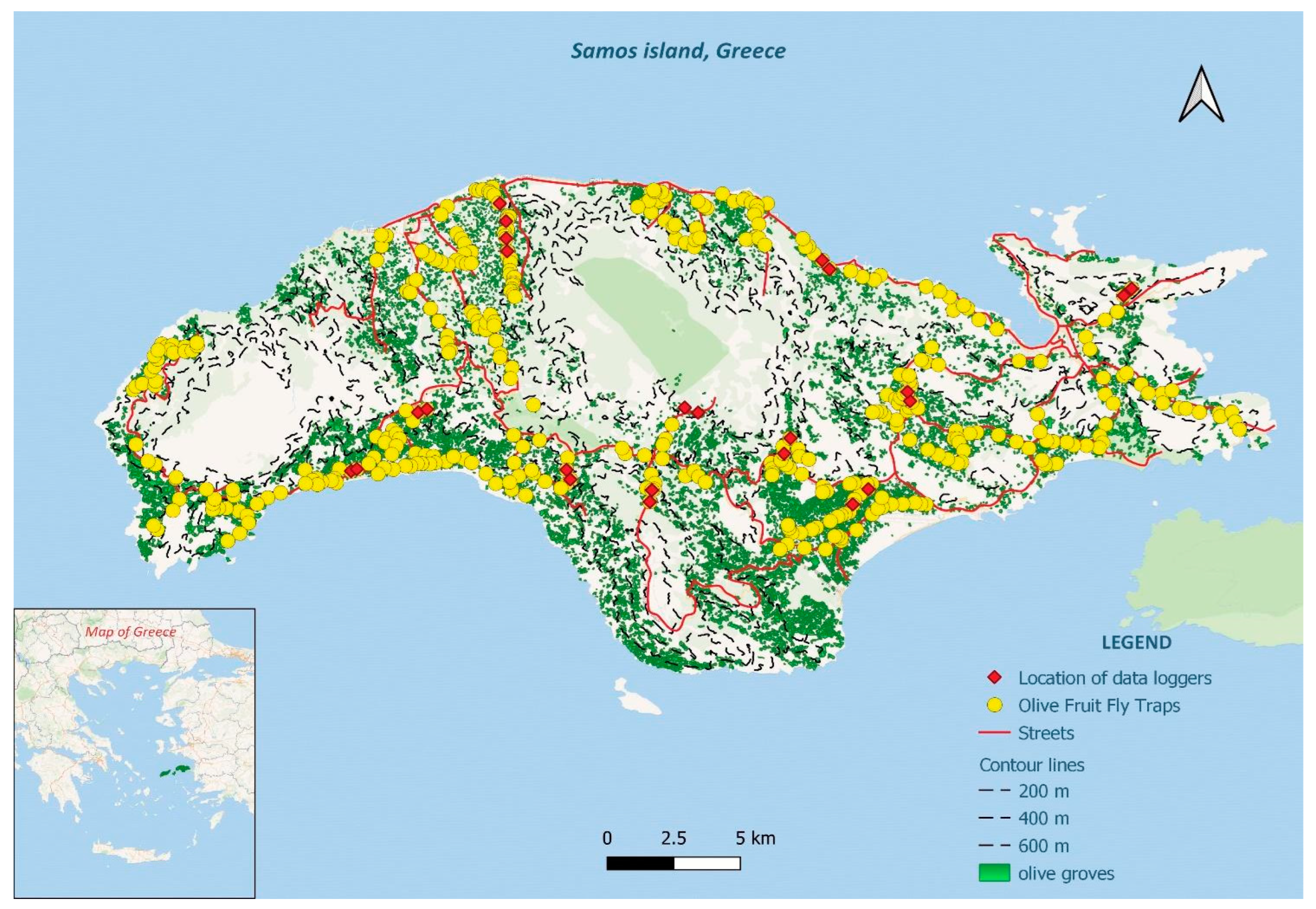
2.1. The Case Study Area: Samos Island
2.2. The Olive Fly Management Program on Samos
2.3. Statistical Analysis
2.4. Analysis
3. Results
3.1. Measured Weather Parameters
3.2. OFF Populations: Temporal and Spatial Differences
4. Discussion
4.1. Relevance of the Approach
4.2. Temporal Patterns of OFF Populations
4.3. Spraying
4.4. OFF Population Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daane, K.M.; Johnson, M.W. Olive Fruit Fly: Managing an Ancient Pest in Modern Times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef]
- Ordano, M.; Engelhard, I.; Rempoulakis, P.; Nemny-Lavy, E.; Blum, M.; Yasin, S.; Lensky, I.M.; Papadopoulos, N.T.; Nestel, D. Olive Fruit Fly (Bactrocera oleae) Population Dynamics in the Eastern Mediterranean: Influence of Exogenous Uncertainty on a Monophagous Frugivorous Insect. PLoS ONE 2015, 10, e0127798. [Google Scholar] [CrossRef]
- Neuenschwander, P.; Michelakis, S. Olive Fruit Drop Caused by Dacus oleae (Gmel.) (Dipt. Tephritidae). Z. Angew. Entomol. 1981, 91, 193–205. [Google Scholar] [CrossRef]
- Kapatos, E.T.; Fletcher, B.S. The Phenology of the Olive Fly, Dacus oleae (Gmel.) (Diptera, Tephritidae), in Corfu. Z. Angew. Entomol. 1984, 97, 360–370. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Cossu, Q.A. Effects of Climate Warming on Olive and Olive Fly (Bactrocera oleae (Gmelin)) in California and Italy. Clim. Chang. 2009, 95, 195–217. [Google Scholar] [CrossRef]
- Petacchi, R.; Marchi, S.; Federici, S.; Ragaglini, G. Large-Scale Simulation of Temperature-Dependent Phenology in Wintering Populations of Bactrocera oleae (Rossi). J. Appl. Entomol. 2015, 139, 496–509. [Google Scholar] [CrossRef]
- Tsiropoulos, G.J. Reproduction and Survival of the Adult Dacus oleae Feeding on Pollens and Honeydews1. Environ. Entomol. 1977, 6, 390–392. [Google Scholar] [CrossRef]
- Malheiro, R.; Casal, S.; Baptista, P.; Pereira, J.A. A Review of Bactrocera oleae (Rossi) Impact in Olive Products: From the Tree to the Table. Trends Food Sci. Technol. 2015, 44, 226–242. [Google Scholar] [CrossRef]
- Tzanakakis, M.E. Insects and Mites Feeding on Olive: Distribution, Importance, Habits, Seasonal Development, and Dormancy; BRILL: Leiden, The Netherlands, 2006; ISBN 978-90-474-1846-7. [Google Scholar]
- Malheiro, R.; Casal, S.; Baptista, P.; Pereira, J.A. Physico-Chemical Characteristics of Olive Leaves and Fruits and Their Relation with Bactrocera oleae (Rossi) Cultivar Oviposition Preference. Sci. Hortic. 2015, 194, 208–214. [Google Scholar] [CrossRef]
- Tzanakakis, M. Seasonal Development And Dormancy of Insects And Mites Feeding on Olive: A Review. Neth. J. Zool. 2003, 52, 87–224. [Google Scholar] [CrossRef]
- Burrack, H.J.; Connell, J.H.; Zalom, F.G. Comparison of Olive Fruit Fly ( Bactrocera oleae (Gmelin)) (Diptera: Tephritidae) Captures in Several Commercial Traps in California. Int. J. Pest Manag. 2008, 54, 227–234. [Google Scholar] [CrossRef]
- Kounatidis, I.; Papadopoulos, N.T.; Mavragani-Tsipidou, P.; Cohen, Y.; Tertivanidis, K.; Nomikou, M.; Nestel, D. Effect of Elevation on Spatio-Temporal Patterns of Olive Fly (Bactrocera oleae) Populations in Northern Greece. J. Appl. Entomol. 2008, 132, 722–733. [Google Scholar] [CrossRef]
- Baratella, V.; Pucci, C.; Paparatti, B.; Speranza, S. Response of Bactrocera oleae to Different Photoperiods and Temperatures Using a Novel Method for Continuous Laboratory Rearing. Biol. Control. 2017, 110, 79–88. [Google Scholar] [CrossRef]
- Genç, H.; Nation, J.L. Survival and Development of Bactrocera oleae Gmelin (Diptera:Tephritidae) Immature Stages at Four Temperatures in the Laboratory. Afr. J. Biotechnol. 2008, 7, 2495–2500. [Google Scholar]
- Tsitsipis, J.A. Effect of Constant Temperatures on Larval and Pupal Development of Olive Fruit Flies Reared on Artificial Diet. Env. Entomol. 1980, 9, 764–768. [Google Scholar] [CrossRef]
- Fletcher, B.S. The Biology of Dacine Fruit Flies. Annu. Rev. Entomol. 1987, 32, 115–144. [Google Scholar] [CrossRef]
- Tsitsipis, J.A. Effect of Constant Temperatures on The Eggs of The Olive Fruit Fly, Dacus oleae (Diptera, Tephritidae). 1977.
- Kapatos, E.T.; Fletcher, B.S. Mortality Factors and Life-Budgets for Immature Stages of the Olive Fly, Dacus oleae (Gmel.) (Diptera, Tephritidae), in Corfu. J. Appl. Entomol. 1986, 102, 326–342. [Google Scholar] [CrossRef]
- Margaritopoulos, J.T.; Skavdis, G.; Kalogiannis, N.; Nikou, D.; Morou, E.; Skouras, P.J.; Tsitsipis, J.A.; Vontas, J. Efficacy of the Pyrethroid Alpha-Cypermethrin against Bactrocera oleae Populations from Greece, and Improved Diagnostic for an IAChE Mutation. Pest Manag. Sci. 2008, 64, 900–908. [Google Scholar] [CrossRef]
- Kakani, E.G.; Zygouridis, N.E.; Tsoumani, K.T.; Seraphides, N.; Zalom, F.G.; Mathiopoulos, K.D. Spinosad Resistance Development in Wild Olive Fruit Fly Bactrocera oleae (Diptera: Tephritidae) Populations in California. Pest Manag. Sci. 2010, 66, 447–453. [Google Scholar] [CrossRef]
- Skouras, P.J.; Margaritopoulos, J.T.; Seraphides, N.A.; Ioannides, I.M.; Kakani, E.G.; Mathiopoulos, K.D.; Tsitsipis, J.A. Organophosphate Resistance in Olive Fruit Fly, Bactrocera oleae, Populations in Greece and Cyprus. Pest Manag. Sci. 2007, 63, 42–48. [Google Scholar] [CrossRef]
- Vontas, J.G.; Cosmidis, N.; Loukas, M.; Tsakas, S.; Hejazi, M.J.; Ayoutanti, A.; Hemingway, J. Altered Acetylcholinesterase Confers Organophosphate Resistance in the Olive Fruit Fly Bactrocera oleae. Pestic. Biochem. Physiol. 2001, 71, 124–132. [Google Scholar] [CrossRef]
- Kampouraki, A.; Stavrakaki, M.; Karataraki, A.; Katsikogiannis, G.; Pitika, E.; Varikou, K.; Vlachaki, A.; Chrysargyris, A.; Malandraki, E.; Sidiropoulos, N.; et al. Recent Evolution and Operational Impact of Insecticide Resistance in Olive Fruit Fly Bactrocera oleae Populations from Greece. J. Pest Sci. 2018, 91, 1429–1439. [Google Scholar] [CrossRef]
- Tscharntke, T.; Bommarco, R.; Clough, Y.; Crist, T.O.; Kleijn, D.; Rand, T.A.; Tylianakis, J.M.; van Nouhuys, S.; Vidal, S. Reprint of “Conservation Biological Control and Enemy Diversity on a Landscape Scale” [Biol. Control 43 (2007) 294–309]. Biol. Control. 2008, 45, 238–253. [Google Scholar] [CrossRef]
- Tscharntke, T.; Brandl, R. Plant-Insect Interactions in Fragmented Landscapes. Annu. Rev. Entomol. 2004, 49, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Nestel, D.; Carvalho, J.; Nemny-Lavy, E. The Spatial Dimension in the Ecology of Insect Pests and Its Relevance to Pest Management. In Insect Pest Management: Field and Protected Crops; Horowitz, A.R., Ishaaya, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 45–63. ISBN 978-3-662-07913-3. [Google Scholar]
- Pontikakos, C.M.; Tsiligiridis, T.A.; Drougka, M.E. Location-Aware System for Olive Fruit Fly Spray Control. Comput. Electron. Agric. 2010, 70, 355–368. [Google Scholar] [CrossRef]
- Pontikakos, C.M.; Tsiligiridis, T.A.; Yialouris, C.P.; Kontodimas, D.C. Pest Management Control of Olive Fruit Fly (Bactrocera oleae) Based on a Location-Aware Agro-Environmental System. Comput. Electron. Agric. 2012, 87, 39–50. [Google Scholar] [CrossRef]
- Ortega, M.; Pascual, S. Spatio-Temporal Analysis of the Relationship between Landscape Structure and the Olive Fruit Fly Bactrocera oleae (Diptera: Tephritidae): Effect of Landscape on B. oleae. Agric. For. Entomol. 2014, 16, 14–23. [Google Scholar] [CrossRef]
- Castrignanò, A.; Boccaccio, L.; Cohen, Y.; Nestel, D.; Kounatidis, I.; Papadopoulos, N.T.; De Benedetto, D.; Mavragani-Tsipidou, P. Spatio-Temporal Population Dynamics and Area-Wide Delineation of Bactrocera oleae Monitoring Zones Using Multi-Variate Geostatistics. Precis. Agric. 2012, 13, 421–441. [Google Scholar] [CrossRef]
- Kavroudakis, D.; Kizos, T.; Tscheulin, T.; Katsikogiannis, G.; Stavrianakis, G.; Tsalta, L. Spatial Analysis of Olive Fly on Samos Island. Int. J. Pest Manag. in press.
- Kizos, T.; Koulouri, M. Same Land Cover, Same Land Use at the Large Scale, Different Landscapes at the Small Scale: Landscape Change in Olive Plantations on Lesvos Island, Greece. Landsc. Res. 2010, 35, 449–467. [Google Scholar] [CrossRef]
- Blum, M.; Lensky, I.M.; Nestel, D. Estimation of Olive Grove Canopy Temperature from MODIS Thermal Imagery Is More Accurate than Interpolation from Meteorological Stations. Agric. For. Meteorol. 2013, 176, 90–93. [Google Scholar] [CrossRef]
- Blum, M.; Lensky, I.M.; Rempoulakis, P.; Nestel, D. Modeling Insect Population Fluctuations with Satellite Land Surface Temperature. Ecol. Model. 2015, 311, 39–47. [Google Scholar] [CrossRef]
- Michalakis, V.I.; Kopsachilis, V.; Katsikogiannis, G.; Vaitis, M.; Kizos, T. Integrated Geo-Spatial Information System for Olive Fruit Fly Management. In Proceedings of the 18th Panhellenic Entomological Congress, Athens, Greece, 15–18 October 2019. [Google Scholar]
- Bournakas, V. Management of Olive Fruit Fly with Bait Sprays Method in Integrated Pest Management; GAIA EΠΙΧΕΙΡΕΙΝ S.A.: Athens, Greece, 2017; ISBN 978-618-81642-6-0. [Google Scholar]
- Navrozidis, E.; Zartaloudis, Z.; Thomidis, T.; Karagiannidis, N.; Roubos, K.; Michailides, Z. Effect of Soil Plowing and Fertilization on the Susceptibility of Four Olive Cultivars to the Insect Bactrocera oleae and the Fungi Sphaeropsis dalmatica AndSpilocaea Oleagina. Phytoparasitica 2007, 35, 429–432. [Google Scholar] [CrossRef]
- Malheiro, R.; Casal, S.; Pinheiro, L.; Baptista, P.; Pereira, J.A. Olive Cultivar and Maturation Process on the Oviposition Preference of Bactrocera oleae (Rossi) (Diptera: Tephritidae). Bull. Entomol. Res. 2019, 109, 43–53. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.r-project.org/ (accessed on 13 December 2021).
- Gonçalves, M.F.; Torres, L.M. The Use of the Cumulative Degree-Days to Predict Olive Fly, Bactrocera oleae (Rossi), Activity in Traditional Olive Groves from the Northeast of Portugal. J. Pest Sci. 2011, 84, 187–197. [Google Scholar] [CrossRef]
- Yokoyama, V.Y. Olive Fruit Fly (Diptera: Tephritidae) in California: Longevity, Oviposition, and Development in Canning Olives in the Laboratory and Greenhouse. J. Econ. Entomol. 2012, 105, 186–195. [Google Scholar] [CrossRef]
- Kombargi, W.S.; Michelakis, S.E.; Petrakis, C.A. Effect of Olive Surface Waxes on Oviposition by Bactrocera oleae (Diptera: Tephritidae). J. Econ. Entomol. 1998, 91, 993–998. [Google Scholar] [CrossRef]
- Benhadi-Marín, J.; Santos, S.A.P.; Baptista, P.; Pereira, J.A. Distribution of Bactrocera oleae (Rossi, 1790) throughout the Iberian Peninsula Based on a Maximum Entropy Modelling Approach. Ann. Appl. Biol. 2020, 177, 112–120. [Google Scholar] [CrossRef]
- Gonçalves, F.; Torres, L. The Use of Trap Captures to Forecast Infestation by the Olive Fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), in Traditional Olive Groves in North-Eastern Portugal. Int. J. Pest Manag. 2013, 59, 279–286. [Google Scholar] [CrossRef]
- Burrack, H.J.; Bingham, R.; Price, R.; Connell, J.H.; Phillips, P.A.; Wunderlich, L.; Vossen, P.M.; O’Connell, N.V.; Ferguson, L.; Zalom, F.G. Understanding the Seasonal and Reproductive Biology of Olive Fruit Fly Is Critical to Its Management. Calif. Agric. 2011, 65, 14–20. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Birouraki, A. Residual Attractiveness of Various Bait Spray Solutions to Bactrocera oleae. Crop Prot. 2015, 68, 60–66. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Birouraki, A.; Gkilpathi, D.; Kapogia, E. Refreshing Bait Spots in an Olive Orchard for the Control of Bactrocera oleae (Diptera: Tephritidae). Crop Prot. 2017, 92, 153–159. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Marketaki, M.; Charalampous, A.; Anagnostopoulos, C.; Bempelou, E. Residual Degradation and Toxicity of Insecticides against Bactrocera oleae. Env. Sci. Pollut. Res. 2018, 25, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Eber, S.; Brandl, R. Ecological and Genetic Spatial Patterns of Urophora Cardui (Diptera: Tephritidae) as Evidence for Population Structure and Biogeographical Processes. J. Anim. Ecol. 1994, 63, 187–199. [Google Scholar] [CrossRef]
- Nestel, D.; Nemny-Lavy, E. Nutrient Balance in Medfly, Ceratitis Capitata, Larval Diets Affects the Ability of the Developing Insect to Incorporate Lipid and Protein Reserves. Entomol. Exp. Appl. 2007, 126, 53–60. [Google Scholar] [CrossRef]
- Shelly, T.; Epsky, N.; Jang, E.B.; Reyes-Flores, J.; Vargas, R. (Eds.) Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-017-9192-2. [Google Scholar]
- Rey, P.J.; Manzaneda, A.J.; Valera, F.; Alcántara, J.M.; Tarifa, R.; Isla, J.; Molina-Pardo, J.L.; Calvo, G.; Salido, T.; Gutiérrez, J.E.; et al. Landscape-Moderated Biodiversity Effects of Ground Herb Cover in Olive Groves: Implications for Regional Biodiversity Conservation. Agric. Ecosyst. Environ. 2019, 277, 61–73. [Google Scholar] [CrossRef]
- Delrio, G.; Deliperi, S.; Lentini, A. Experiments for the Control of Olive Fly Using a “Push-Pull” Method. IOBC/WPRS Bull. 2010, 59, 89–92. [Google Scholar]
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground Cover Management with Mixtures of Flowering Plants to Enhance Insect Pollinators and Natural Enemies of Pests in Olive Groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Broumas, T.; Haniotakis, G.; Liaropoulos, C.; Tomazou, T.; Ragoussis, N. The Efficacy of an Improved Form of the Mass-Trapping Method, Forthe Control of the Olive Fruit Fly, Bactrocera oleae (Gmelin) (Dipt., Tephritidae): Pilot-Scale Feasibility Studies. J. Appl. Entomol. 2002, 126, 217–223. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Birouraki, A.; Ioannou, A.; Kapogia, E. Improvement of Bait Sprays for the Control of Bactrocera oleae (Diptera: Tephritidae). Crop Prot. 2016, 81, 1–8. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Birouraki, A. Comparative Field Studies of Bactrocera oleae Baits in Olive Orchards in Crete. Crop Prot. 2014, 65, 238–243. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variable | Name | Definition | Data Availability |
|---|---|---|---|
| variable 1 | Spatial location and altitude of the traps | Geographical coordinates of trap locations | once |
| variable 2 | Hours with maximum temperature exceeding 32 °C | Hours per 10 days and month when maximum temperature exceeds 32 °C calculated by hourly measurements of temperature | 10 days/monthly |
| variable 3 | Hours with maximum temperature exceeding 35 °C | Hours per 10 days and month when maximum temperature exceeds 32 °C calculated by hourly measurements of temperature | 10 days/monthly |
| variable 4 | Hours with Relative Humidity below 60% | Hours per 10 days and month when RH is below 60% calculated by hourly measurements of temperature | 10 days/monthly |
| variable 5 | OFF population | Olive female fly populations as measured in each trap per 5 days, recalculated for 10 days | 10 days/monthly, seasonal |
| variable 6 | OFF population | Olive fly populations as measured in each trap per 5 days, recalculated for 10 days | 10 days/monthly, seasonal |
| variable 7 | % fertile infestations on the olive fruit/month | Percentage of fertile infestations of total infestations, measured in sampled olives per month for August, September, and October | monthly |
| variable 8 | Spraying incident | If a spray is performed, the area is considered as sprayed for the particular 10 day period | random |
| Average Hours with Temp > 32 °C | Tmean °C | RHmean% | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Month | 2017 | 2018 | 2019 | 2017 | 2018 | 2019 | 2017 | 2018 | 2019 |
| 6 | 24.9 | 7.7 | 30.1 | 25.8 | 24.9 | 27.2 | 52.5 | 61.4 | 55.8 |
| 7 | 52.3 | 34.5 | 30.4 | 28.2 | 27.7 | 27.0 | 46.5 | 56.2 | 50 |
| 8 | 36.2 | 33.2 | 47.3 | 27.7 | 27.7 | 28.3 | 52.4 | 58.3 | 49.9 |
| 9 | 13.2 | 11.9 | 4.7 | 24.3 | 24.7 | 23.5 | 51.8 | 58.1 | 60.2 |
| 10 | 0.7 | 0 | 0.2 | 18.6 | 19.1 | 20.7 | 62.1 | 72.7 | 70.9 |
| Altitude Zone | Year/Month | June | July | August | September | October |
|---|---|---|---|---|---|---|
| 0–200 m | 2017 | 23.7 | 38.8 *2 | 25.3 *3 | 53.7 *4 | 82.2 *7 |
| 2018 | 25.5 | 49.0 | 49.6 | 69.4 *5 | 100.6 *6 | |
| 2019 | 6.0 *1 | 13.5 | 53.4 | 83.6 | 59.7 | |
| 200–400 m | 2017 | 18.6 | 51.5 | 66.7 *3 | 70.6 *4 | 39.9 *7 |
| 2018 | 26.0 | 50.6 | 52.8 | 50.1 *5 | 51.7 *6 | |
| 2019 | 4.0 | 13.6 | 68.2 | 102.0 | 68.6 *8 | |
| >400 m | 2017 | 27.8 | 74.4 *2 | 33.8 *3 | 36.8 *4 | 28.4 *7 |
| 2018 | 21.6 | 42.4 | 49.0 | 51.8 | 41.2 *6 | |
| 2019 | 1.1 *1 | 27.9 | 35.9 | 54.3 | 45.2 *8 | |
| Total | 2017 | 22.9 | 43.2 | 33.8 | 56.0 | 71.0 |
| 2018 | 25.4 | 49.0 | 50.2 | 64.7 | 87.8 | |
| 2019 | 5.4 | 14.3 | 55.2 | 85.5 | 60.6 |
| 10-Day Periods for 3 Years | T1−T0 (%) | T2–T1 (%) | T3–T2 (%) | |||
|---|---|---|---|---|---|---|
| Unsprayed | Sprayed | Unsprayed | Sprayed | Unsprayed | Sprayed | |
| 1st | 69.8 | −26.0 | 4.1 | |||
| 2nd | −28.7 | 20.5 | −1.4 | 65.1 | −7.4 | 11.4 |
| 3rd | 5.7 | 2.6 | −27.4 | 19.7 | −14.7 | 29.3 |
| 4th | 0.5 | −34.8 | 17.1 | −34.5 | 36.6 | 90.1 |
| 5th | −8.4 | 98.7 | 40.1 | 39.1 | 14.1 | 40.6 |
| 6th | 42.5 | 9.5 | 22.4 | 32.1 | 14.0 | −24.4 |
| 7th | 52.8 | −17.3 | 12.5 | 8.5 | −10.5 | 18.5 |
| 8th | 21.9 | 2.3 | 11.4 | −16.7 | −0.4 | −4.2 |
| 9th | −3.7 | 18.0 | −3.4 | 17.1 | −15.2 | −28.1 |
| 10th | 2.1 | −9.1 | −28.1 | 6.6 | 46.6 | −0.5 |
| 11th | −3.4 | −41.3 | 22.6 | 37.8 | −16.4 | 3.2 |
| 12th | 15.7 | 51.0 | −9.0 | −15.6 | ||
| 13th | −11.8 | −10.2 | ||||
| Value | Std. Error | DF | t-Value | p-Value | |
|---|---|---|---|---|---|
| Intercept | 0.5037640 | 0.11655140 | 718 | 4.322248 | <0.001 |
| Altitude | 0.0032617 | 0.00037427 | 346 | 8.714936 | <0.001 |
| Spray | 0.3988375 | 0.06502091 | 718 | 6.133989 | <0.001 |
| 1st 10-days | 0.0282857 | 0.00176229 | 718 | 16.050507 | <0.001 |
| 2nd 10-days | 0.0131370 | 0.00153348 | 718 | 8.566800 | <0.001 |
| 3rd 10-days | 0.0176605 | 0.00211540 | 718 | 8.348552 | <0.001 |
| 1st: 2nd 10-days | −0.0000568 | 0.00000792 | 718 | −7.174855 | <0.001 |
| 1st: 3rd 10-days | −0.0002365 | 0.00003094 | 718 | −7.644837 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsikogiannis, G.; Kavroudakis, D.; Tscheulin, T.; Kizos, T. Population Dynamics of the Olive Fly, Bactrocera oleae (Diptera: Tephritidae), Are Influenced by Different Climates, Seasons, and Pest Management. Sustainability 2023, 15, 14466. https://doi.org/10.3390/su151914466
Katsikogiannis G, Kavroudakis D, Tscheulin T, Kizos T. Population Dynamics of the Olive Fly, Bactrocera oleae (Diptera: Tephritidae), Are Influenced by Different Climates, Seasons, and Pest Management. Sustainability. 2023; 15(19):14466. https://doi.org/10.3390/su151914466
Chicago/Turabian StyleKatsikogiannis, Georgios, Dimitris Kavroudakis, Thomas Tscheulin, and Thanasis Kizos. 2023. "Population Dynamics of the Olive Fly, Bactrocera oleae (Diptera: Tephritidae), Are Influenced by Different Climates, Seasons, and Pest Management" Sustainability 15, no. 19: 14466. https://doi.org/10.3390/su151914466
APA StyleKatsikogiannis, G., Kavroudakis, D., Tscheulin, T., & Kizos, T. (2023). Population Dynamics of the Olive Fly, Bactrocera oleae (Diptera: Tephritidae), Are Influenced by Different Climates, Seasons, and Pest Management. Sustainability, 15(19), 14466. https://doi.org/10.3390/su151914466