
Predicting and Mapping Dominant Height of Oriental Beech Stands Using Environmental Variables in Sinop, Northern Turkey

Abstract
:
1. Introduction
2. Materials and Methods
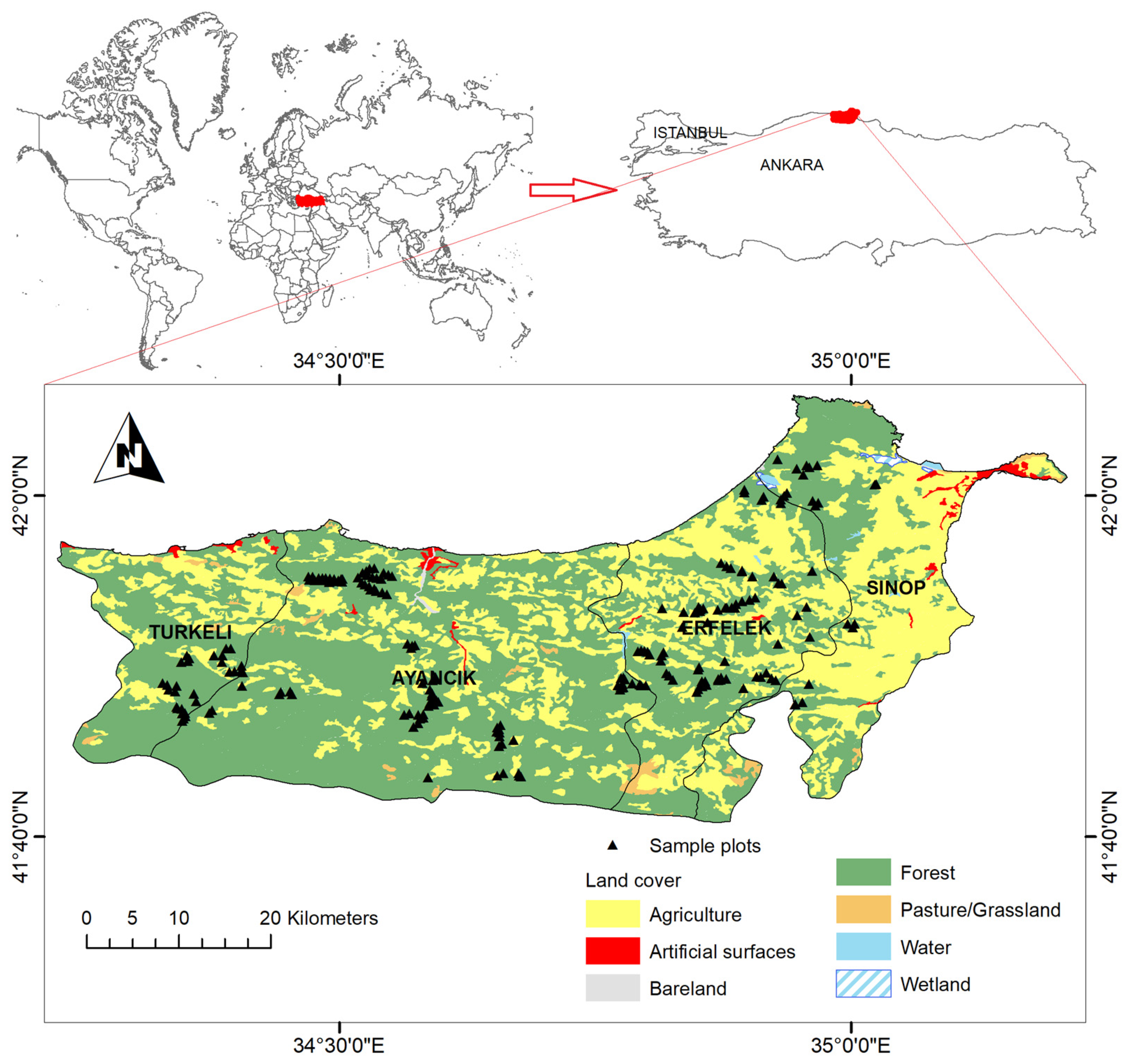
2.1. Site Description
2.2. Data Collection and Analyses
2.2.1. Field Data Collection
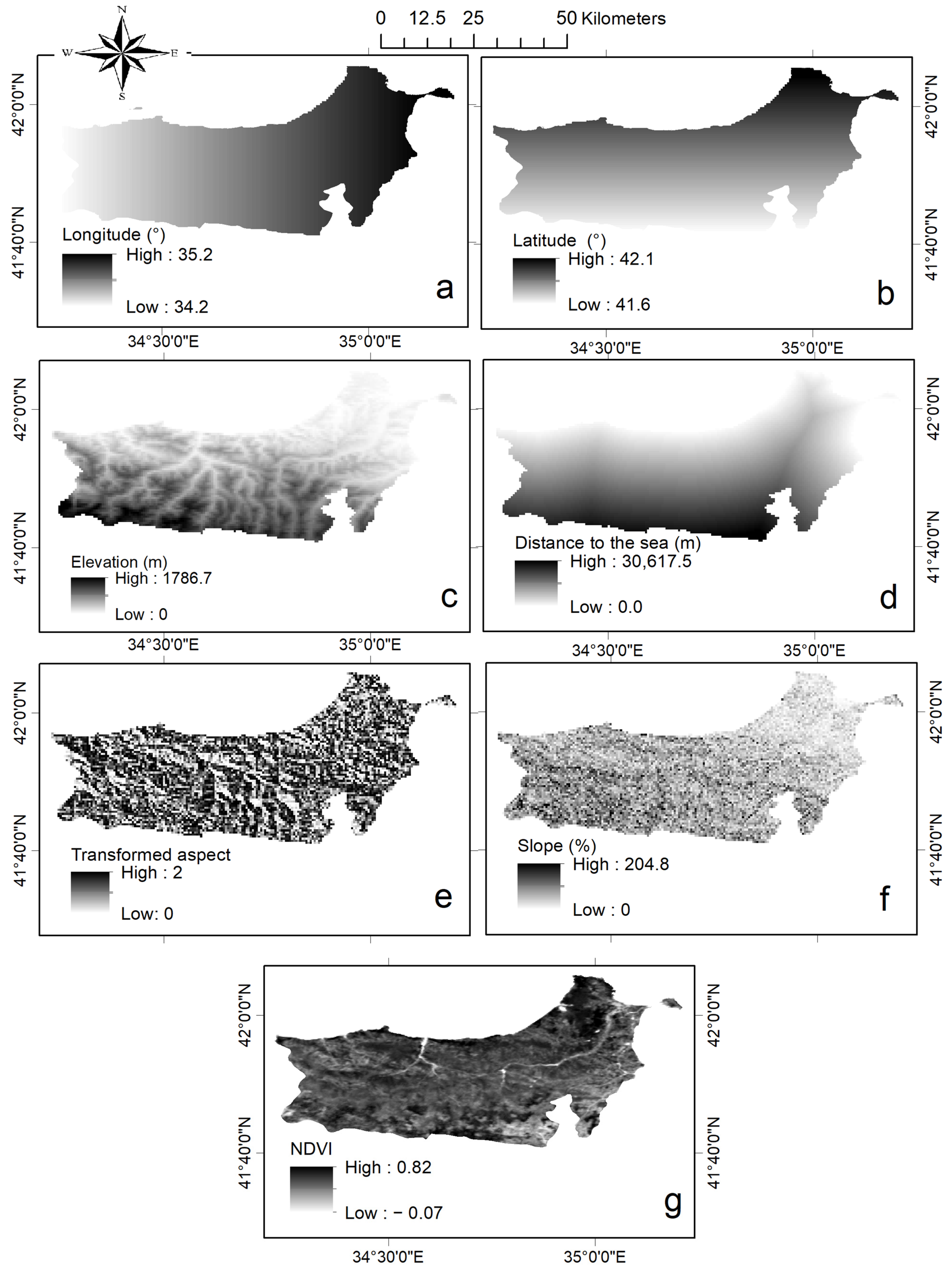
2.2.2. Spatial Data Extraction
2.2.3. Analysis and Mapping
3. Results and Discussion
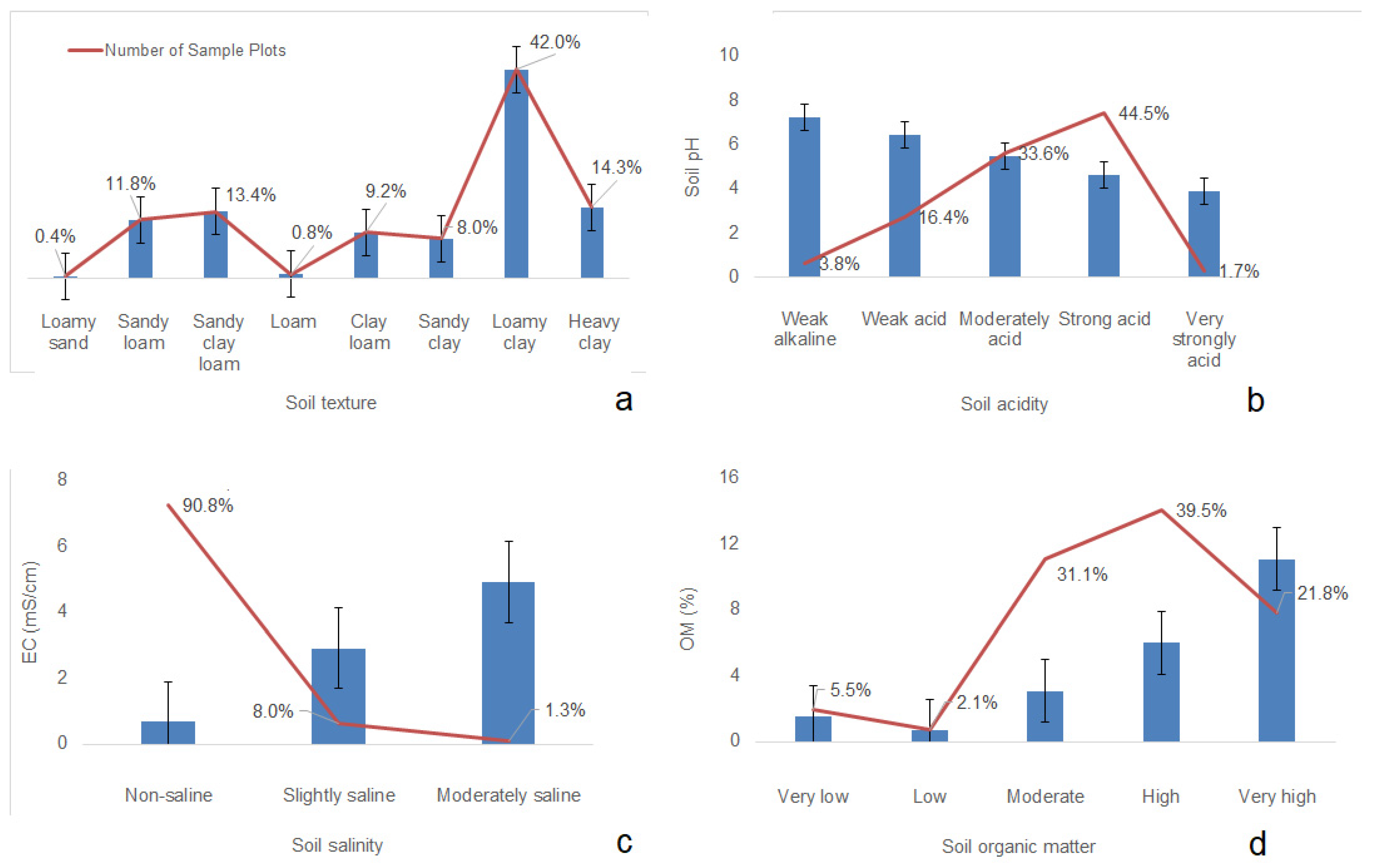
3.1. Field-Based Data
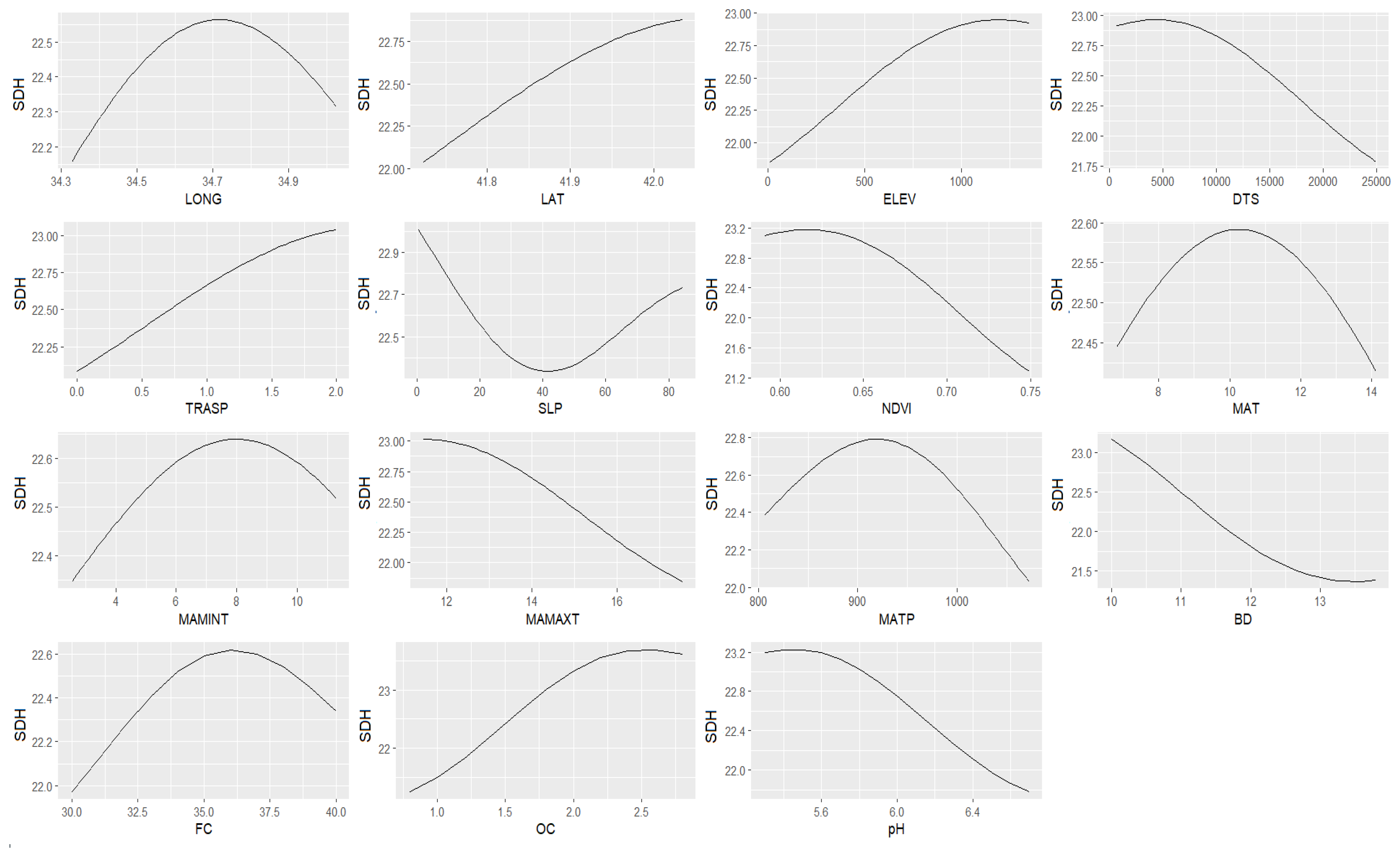
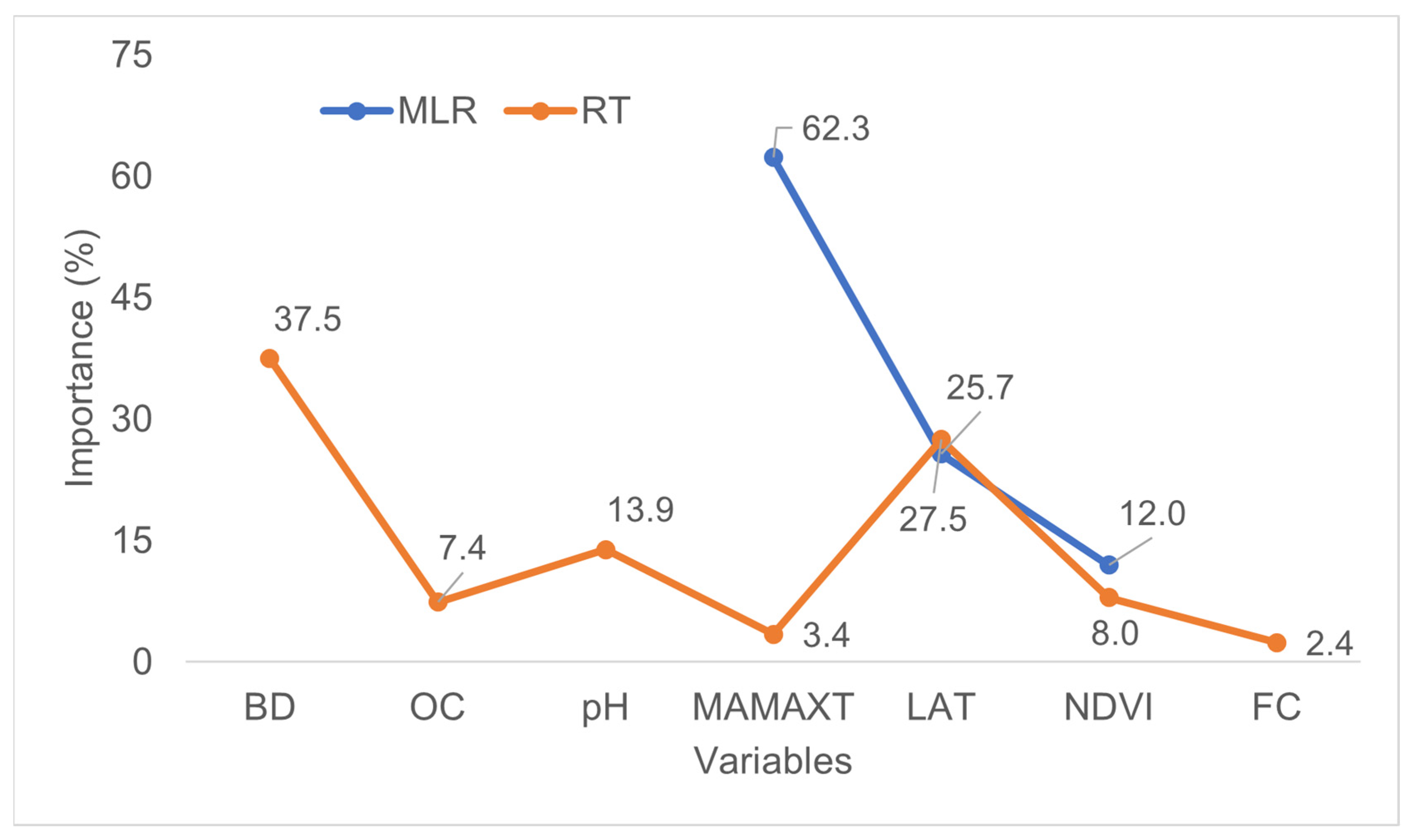
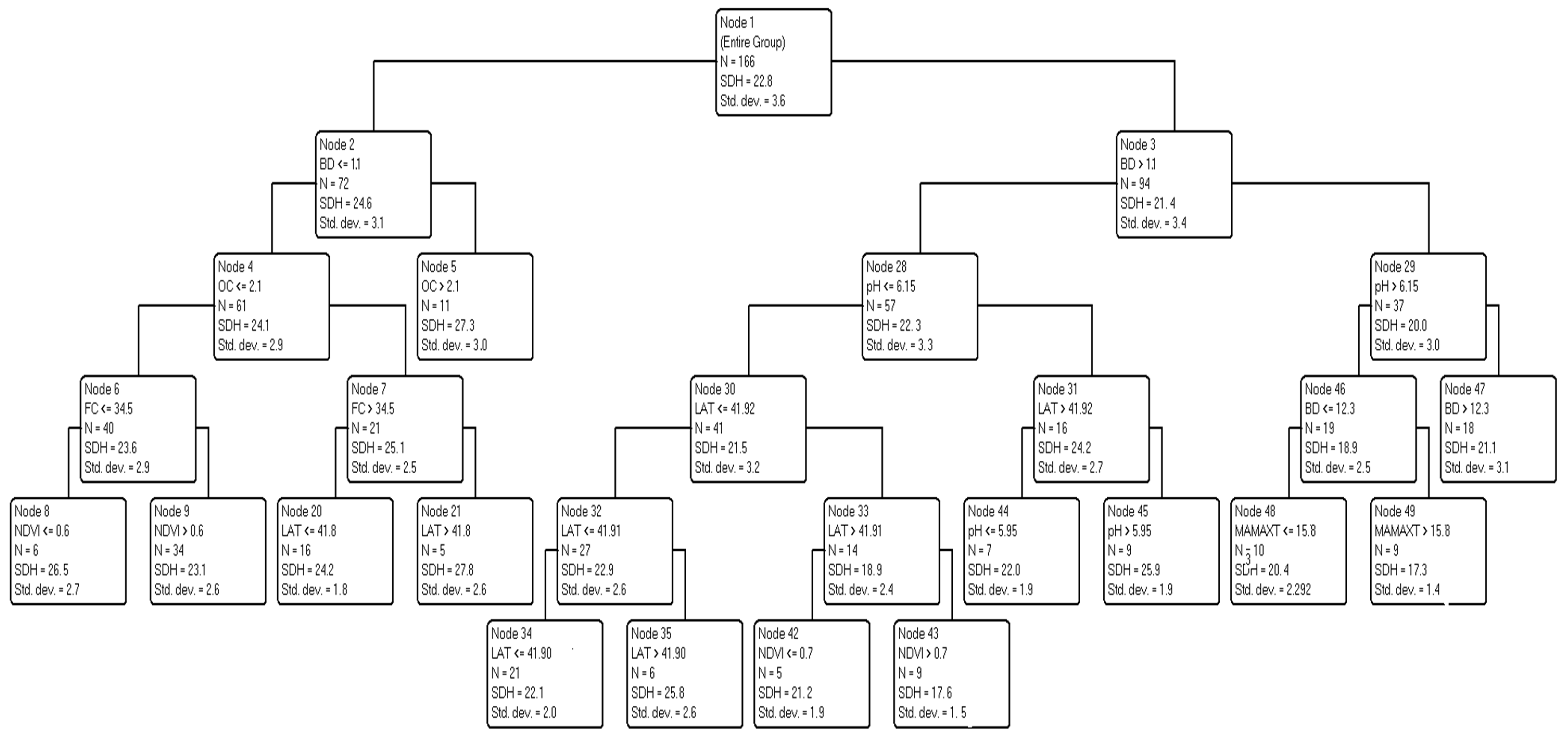
3.2. Relationships between the Stand Dominant Height (SDH) and Spatial Data
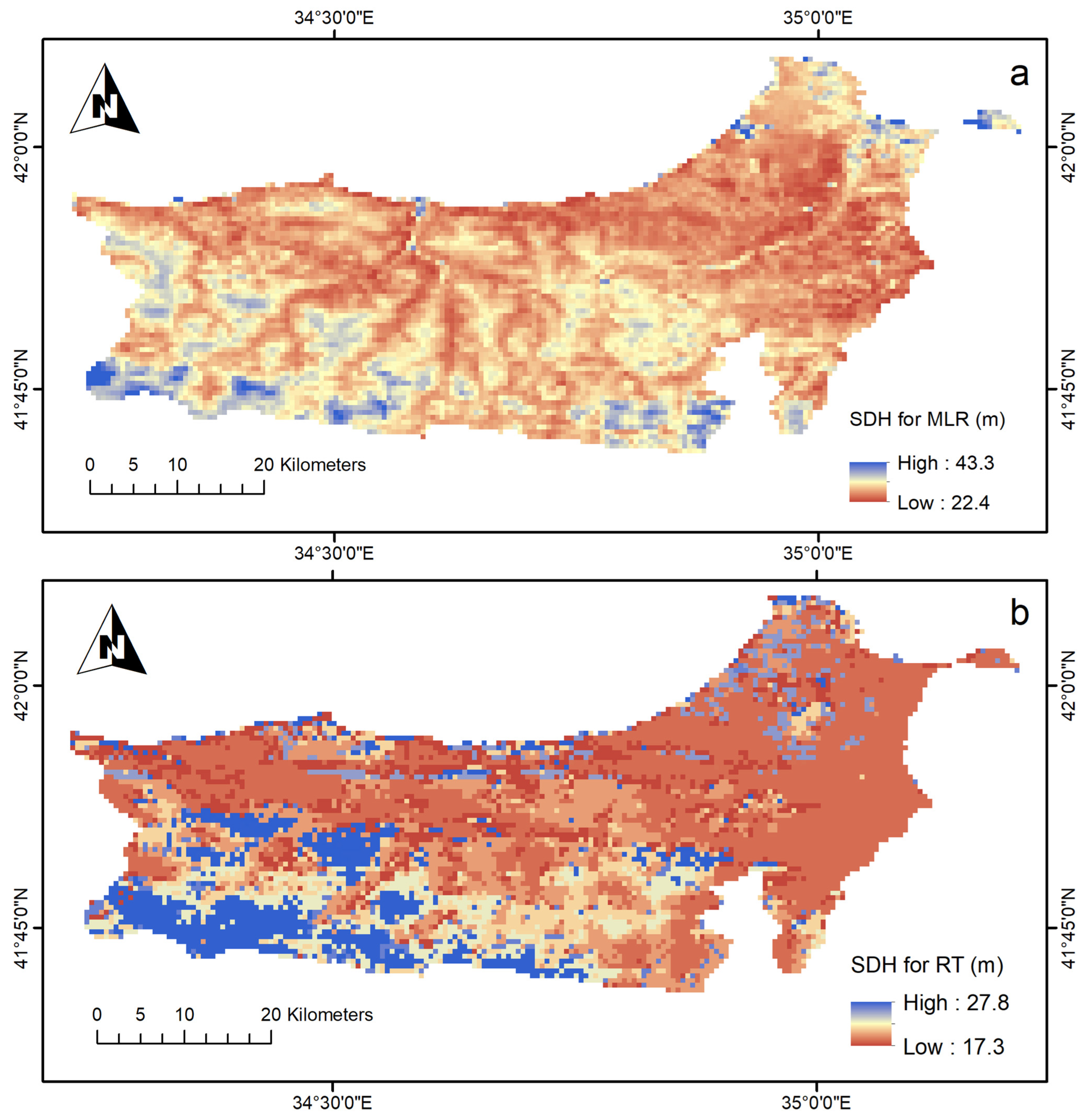
3.3. Predicting and Mapping SDH
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13. [Google Scholar] [CrossRef]
- García, O.; Batho, A. Top Height Estimation in Lodgepole Pine Sample Plots. West. J. Appl. For. 2005, 20, 64–68. [Google Scholar] [CrossRef]
- Noordermeer, L.; Gobakken, T.; Næsset, E.; Bollandsås, O.M. Predicting and mapping site index in operational forest inventories using bitemporal airborne laser scanner data. For. Ecol. Manag. 2020, 457, 117768. [Google Scholar] [CrossRef]
- Yener, I.; Altun, L. Predicting Site Index for Oriental Spruce (Picea orientalis L. (Link)) Using Ecological Factors in the Eastern Black Sea, Turkey. Fresenius Environ. Bull. 2018, 27, 3107–3116. [Google Scholar]
- Mohammadi, J.; Shataee, S.; Babanezhad, M. Estimation of forest stand volume, tree density and biodiversity using Landsat ETM plus Data, comparison of linear and regression tree analyses. Procedia Environ. Sci. 2011, 7, 299–304. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O.; Bergeron, Y.; Fenton, N.; Cheng, L.Z.; Anyomi, K. Effects of topography and thickness of organic layer on productivity of black spruce boreal forests of the Canadian Clay Belt region. For. Ecol. Manag. 2014, 330, 144–157. [Google Scholar] [CrossRef]
- Alavi, S.J.; Ahmadi, K.; Dormann, C.; Serra-Diaz, J.M.; Nouri, Z. Assessing the dominant height of oriental beech (Fagus orientalis L.) in relation to edaphic and physiographic variables in the Hyrcanian Forests of Iran. Biotechnol. Agron. Soc. Environ. 2020, 24, 262–273. [Google Scholar] [CrossRef]
- Littke, K.M.; Harrison, R.B.; Zabowski, D. Determining the Effects of Biogeoclimatic Properties on Different Site Index Systems of Douglas-fir in the Coastal Pacific Northwest. For. Sci. 2016, 62, 503–512. [Google Scholar] [CrossRef]
- Ozkan, K. Modelling ecological data using classification and regression tree technique (CART). Turk. J. For. 2012, 13, 1–4. [Google Scholar]
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer Classification and Regression Tree Techniques: Bagging and Random Forests for Ecological Prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- Yuan, Z.Q.; Ali, A.; Wang, S.P.; Gazol, A.; Freckleton, R.; Wang, X.G.; Lin, F.; Ye, J.; Zhou, L.; Hao, Z.Q.; et al. Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests. Sci. Total Environ. 2018, 630, 422–431. [Google Scholar]
- Gulsoy, S.; Cinar, T. The Relationships between Environmental Factors and Site Index of Anatolian Black Pine (Pinus Nigra Arn. Subsp. Pallasiana (Lamb.) Holmboe) Stands in Demirci (Manisa) District, Turkey. Appl. Ecol. Environ. Res. 2019, 17, 1235–1246. [Google Scholar] [CrossRef]
- Seltmann, C.T.; Wernicke, J.; Petzold, R.; Baumann, M.; Munder, K.; Martens, S. The relative importance of environmental drivers and their interactions on the growth of Norway spruce depends on soil unit classes: A case study from Saxony and Thuringia, Germany. For. Ecol. Manag. 2021, 480, 118671. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Feng, Z.K.; Wang, S.; Ji, W.X. Disentangling the Factors that Contribute to the Growth of Betula spp. and Cunninghami lanceolata in China Based on Machine Learning Algorithms. Sustainability 2022, 14, 8346. [Google Scholar] [CrossRef]
- Velioglu, E.; Guener, S.T.; Karakurt, H.; Tastan, Y.; Yavuz, Z.; Tugrul, D. Relationships between site index and ecological variables of trembling poplar forests (Populus tremula L.) in Turkiye. Environ. Monit. Assess. 2023, 195, 308. [Google Scholar]
- Aertsen, W.; Kint, V.; van Orshoven, J.; Ozkan, K.; Muys, B. Comparison and ranking of different modelling techniques for prediction of site index in Mediterranean mountain forests. Ecol. Model. 2010, 221, 1119–1130. [Google Scholar] [CrossRef]
- Orlandini, S.; Bindi, M.; Howden, M. Plant biometeorology and adaptation. In Biometeorology for Adaptation to Climate Variability and Change; Springer: Berlin/Heidelberg, Germany, 2009; pp. 107–129. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. J. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar]
- Stage, A.R.; Salas, C. Interactions of elevation, aspect, and slope in models of forest species composition and productivity. For. Sci. 2007, 53, 486–492. [Google Scholar]
- Becknell, J.M.; Powers, J.S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 2014, 44, 604–613. [Google Scholar] [CrossRef]
- Brito, V.V.; Rubilar, R.A.; Cook, R.L.; Campoe, O.C.; Carter, D.R.; Mardones, O. Evaluating remote sensing indices as potential productivity and stand quality indicators for Pinus radiata plantations. Sci. For. 2021, 49, e3316. [Google Scholar] [CrossRef]
- Gazol, A.; Rozas, V.; Arribas, S.C.; Ponce, R.A.; Rodriguez-Puerta, F.; Gomez, C.; Olano, J.M. Stand characteristics modulate secondary growth responses to drought and gross primary production in Pinus halepensis afforestation. Eur. J. For. Res. 2023, 142, 353–366. [Google Scholar]
- Zushi, K. Regional estimation of Japanese cedar (Cryptomeria japonica D. Don) productivity by use of digital terrain analysis. J. For. Res. 2007, 12, 289–297. [Google Scholar] [CrossRef]
- NASA/METI/AIST, Japan Spacesystems and U.S./Japan ASTER Science Team. ASTER Global Digital Elevation Model [Data Set]. 2019. Available online: https://lpdaac.usgs.gov/products/astgtmv003/ (accessed on 9 September 2022).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Hengl, T.; Wheeler, I. Soil Organic Carbon Content in x 5 g/kg at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution (Version v02) [Data Set]; Zenodo: Geneva, Switzerland, 2018. [Google Scholar]
- Hengl, T. Clay Content in % (kg/kg) at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution (Version v02) [Data Set]; Zenodo: Geneva, Switzerland, 2018. [Google Scholar]
- Hengl, T. Soil Bulk Density (Fine Earth) 10 x kg/m-Cubic at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution (Version v02) [Data Set]; Zenodo: Geneva, Switzerland, 2018. [Google Scholar]
- Xu, C.; Du, X.; Jian, H.; Dong, Y.; Qin, W.; Mu, H.; Yan, Z.; Zhu, J.; Fan, X. Analyzing large-scale Data Cubes with user-defined algorithms: A cloud-native approach. Int. J. Appl. Earth Obs. Geoinf. 2022, 109, 102784. [Google Scholar] [CrossRef]
- OGM. Ormanlarımızda Yayılış Gösteren Asli Ağaç Türleri; Orman Genel Müdürlüğü: Ankara, Turkey, 2009; p. 48. [Google Scholar]
- OGM, Orman Varlığımız. TC Çevre Ve Orman Bakanlığı; Orman Genel Müdürlüğü Yayınları: Ankara, Turkey, 2020. [Google Scholar]
- Guvendi, E. Saf Doğu Kayını (Fagus orientalis Lipsky.) Ormanlarının Ekolojik Tabanlı İdare Sürelerinin Belirlenmesi (Sinop-Ayancık-Türkeli Örneği). Ph.D. Thesis, Karadeniz Technical University, Trabzon, Türkiye, 2013. [Google Scholar]
- MTA. Geological Map of Turkey; General Directorate of Mineral Research and Exploration: Ankara, Turkey, 2019. [Google Scholar]
- Hengl, T.; Nauman, T. Predicted USDA SOIL Great Groups at 250 m (Probabilities), v01; Zenodo: Geneva, Switzerland, 2018. [Google Scholar]
- Özdemir, N. One of the effective natural disaster in Sinop: The landslide. Dicle Univ. J. Ziya Gökalp Fac. Educ. 2005, 5, 67–106. [Google Scholar]
- Cover Corine Land, European Union, Copernicus Land Monitoring Service. 2018. Available online: https://sdi.eea.europa.eu/catalogue/copernicus/api/records/71c95a07-e296-44fc-b22b-415f42acfdf0?language=all (accessed on 9 September 2022).
- Yıldız, S. Sinop ve Çevresinin Bitki Örtüsü. Master’s Thesis, Istanbul University, Istanbul, Türkiye, 2000. (In Turkish). [Google Scholar]
- Pennock, D.; Yates, T.; Braidek, J. Soil sampling designs. Soil Sampl. Methods Anal. 2008, 2, 25–37. [Google Scholar]
- van Laar, A.; Akça, A. Forest Mensuration; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Schoeneberger, P.J. Field Book for Describing and Sampling Soils; National Soil Survey Center, Natural Resources Conservation Service, U.S. Department of Agriculture: Lincoln, NE, USA, 2002.
- Cepel, N. Forest Ecology; Istanbul University, Faculty of Forestry: İstanbul, Turkey, 1978. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Kalra, Y.P.; Maynard, D.G. Methods Manual for Forest Soil and Plant Analysis; Forestry Canada, Northwest Region, Northern Forestry Centre: Edmonton, AB, Canada, 1991; p. 116. [Google Scholar]
- Walkley, A. A critical examination of a rapid method for determining organic carbon in soils-Effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 1947, 63, 251–264. [Google Scholar] [CrossRef]
- Bolton, D.K.; White, J.C.; Wulder, M.A.; Coops, N.C.; Hermosilla, T.; Yuan, X. Updating stand-level forest inventories using airborne laser scanning and Landsat time series data. Int. J. Appl. Earth Obs. Geoinf. 2018, 66, 174–183. [Google Scholar] [CrossRef]
- ESRI. ArcMap 10.2, 10.2; ESRI: Redlands, CA, USA, 2014. [Google Scholar]
- Kelso, N.V.; Patterson, T. Natural earth vector. Cartogr. Perspect. 2009, 64, 45–50. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Yener, İ. Development of high-resolution annual climate surfaces for Turkey using ANUSPLIN and comparison with other methods. Atmósfera 2022, 37, 425–444. [Google Scholar] [CrossRef]
- Greenwell, B.M. pdp: An R package for constructing partial dependence plots. R J. 2017, 9, 421. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Sherrod, P.H. DTREG Predictive Modeling Software. 2020. Available online: http://www.dtreg.com (accessed on 9 September 2022).
- Tomlin, C.D. Geographic Information Systems and Cartographic Modeling; Prentice Hall: Englewood Cliffs, NJ, USA, 1990; Volume 249. [Google Scholar]
- Tomlin, C.D. Map algebra: One perspective. Landsc. Urban Plan. 1994, 30, 3–12. [Google Scholar] [CrossRef]
- Yıldız, O.; Bircan, Ş. Batı Karadeniz kıyı bölgesi’nde yetişen Kayın (Fagus orientalis Lipsky) odununun fiziksel ve mekanik özelliklerinin yetişme ortamı değişkenleriyle ilişkisi. Anadolu Orman Araştırmaları Derg. 2022, 8, 61–72. [Google Scholar] [CrossRef]
- Göl, C.; Çelik, N.; Çakır, M.; Gül, E. Türkmen Dağı (Evkondu Tepe) doğu kayını (Fagus orientalis Lipsky.) ormanlarının bazı yetişme ortamı özellikleri. Süleyman Demirel Üniversitesi Orman Fakültesi Derg. Seri A 2008, 1, 48–60. [Google Scholar]
- Özel, H.; Ertekin, M. Ecological conditions in the natural group regeneration areas of oriental beech (Fagus orientalis Lipsky.) in Bartın and Devrek district. Bartın Orman Fakültesi Derg. 2010, 12, 47–64. [Google Scholar]
- Yılmaz, M. Effects of soil and environmental factors on the site productivity of pure Oriental beech forests in Akkuş Region of Turkey. Eurasian J. For. Sci. 2019, 7, 107–120. [Google Scholar]
- Yilmaz, M.; Usta, A.; Ozturk, I. Relationships between site productivity and environmental variables of Oriental beech forests in northeastern Turkey. Austrian J. For. Sci. 2022, 139, 239–262. [Google Scholar]
- Bolat, I.; Kara, O.; Tunay, M. Seasonal Changes of Microbial Biomass Carbon, Nitrogen, and Phosphorus in Soil Under an Oriental Beech Stand. Forestist 2022, 72, 259–265. [Google Scholar] [CrossRef]
- Azaryan, M.; Vajari, K.A.; Amanzadeh, B. Soil properties and fine root morphological traits in relation to soil particle-size fractions in a broad-leaved beech (Fagus orientalis Lipsky) forest. Acta Oecologica 2022, 117, 103852. [Google Scholar]
- Akbas, M.; Tufekcioglu, A. Contribution of the Root Component to Soil Respiration in Oriental Beech Stands in Artvin, Turkey. For. Sci. 2022, 68, 399–409. [Google Scholar]
- Amolikondori, A.; Vajari, K.A.; Feizian, M.; Di Iorio, A. Influences of forest gaps on soil physico-chemical and biological properties in an oriental beech (Fagus orientalis L.) stand of Hyrcanian forest, north of Iran. Iforest-Biogeosci. For. 2020, 13, 124–129. [Google Scholar] [CrossRef]
- Guner, S.T. Relatıonshıps between Sıte Index and Ecologıcal Varıables of Orıental Beech Forest in the Marmara Regıon of Turkey. Fresenius Environ. Bull. 2021, 30, 6920–6927. [Google Scholar]
- Mohammadi, M.F.; Jalali, S.G.; Kooch, Y.; Said-Pullicino, D. The effect of landform on soil microbial activity and biomass in a Hyrcanian oriental beech stand. Catena 2017, 149, 309–317. [Google Scholar] [CrossRef]
- Kooch, Y.; Parsapour, M.K.; Egli, M.; Moghimian, N. Forest floor and soil properties in different development stages of Oriental beech forests. Appl. Soil Ecol. 2021, 161, 103823. [Google Scholar]
- Bakhshandeh-Navroud, B.; Vajari, K.A.; Pilehvar, B.; Kooch, Y. The interactions between tree-herb layer diversity and soil properties in the oriental beech (Fagus orientalis Lipsky) stands in Hyrcanian forest. Environ. Monit. Assess. 2018, 190, 425. [Google Scholar] [CrossRef]
- Bailey, R.G. Ecosystem Geography: From Ecoregions to Sites; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Spurr, S.H.; Barnes, B.V. Forest Ecology; Wiley: Hoboken, NJ, USA, 1991. [Google Scholar]
- Atalay, İ. Türkiye’nin Ekolojik Bölgeleri (Ecoregions of Turkey); TC Orman Bakanlığı Yayını: Ankara, Turkey, 2002. [Google Scholar]
- Afif-Khouri, E.; Alvarez-Alvarez, P.; Fernandez-Lopez, M.J.; Oliveira-Prendes, J.A.; Camara-Obregon, A. Influence of climate, edaphic factors and tree nutrition on site index of chestnut coppice stands in north-west Spain. Forestry 2011, 84, 385–396. [Google Scholar] [CrossRef]
- Güner, Ş.T.; Comez, A.; Ozkan, K.; Karatas, R.; Celik, N. Modelling the productivity of Anatolian black pine plantations in Turkey. Istanb. Univ. Fac. For. 2016, 66, 159–172. [Google Scholar]
- Klinka, K.; Wang, Q.; Carter, R.E.; Chen, H.Y.H. Height growth-elevation relationships in subalpine forests of interior British Columbia. For. Chron. 1996, 72, 193–198. [Google Scholar] [CrossRef]
- Jiao, K.W.; Gao, J.B.; Liu, Z.H. Precipitation Drives the NDVI Distribution on the Tibetan Plateau While High Warming Rates May Intensify Its Ecological Droughts. Remote Sens. 2021, 13, 1305. [Google Scholar] [CrossRef]
- Shi, L.; Fan, H.J.; Yang, L.Y.; Jiang, Y.T.; Sun, Z.H.; Zhang, Y.L. NDVI-based spatial and temporal vegetation trends and their response to precipitation and temperature changes in the Mu Us Desert from 2000 to 2019. Water Sci. Technol. 2023, 88, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Bollandsas, O.M.; Orka, H.O.; Dalponte, M.; Gobakken, T.; Naesset, E. Modelling Site Index in Forest Stands Using Airborne Hyperspectral Imagery and Bi-Temporal Laser Scanner Data. Remote Sens. 2019, 11, 1020. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P.; Kettle, W.D. Relations between NDVI and tree productivity in the central Great Plains. Int. J. Remote Sens. 2004, 25, 3127–3138. [Google Scholar] [CrossRef]
- Erasmi, S.; Klinge, M.; Dulamsuren, C.; Schneider, F.; Hauck, M. Modelling the productivity of Siberian larch forests from Landsat NDVI time series in fragmented forest stands of the Mongolian forest-steppe. Environ. Monit. Assess. 2021, 193, 200. [Google Scholar] [PubMed]
- Main-Knorn, M.; Moisen, G.G.; Healey, S.P.; Keeton, W.S.; Freeman, E.A.; Hostert, P. Evaluating the Remote Sensing and Inventory-Based Estimation of Biomass in the Western Carpathians. Remote Sens. 2011, 3, 1427–1446. [Google Scholar] [CrossRef]
- Fang, K.Y.; Frank, D.; Zhao, Y.; Zhou, F.F.; Seppa, H. Moisture stress of a hydrological year on tree growth in the Tibetan Plateau and surroundings. Environ. Res. Lett. 2015, 10, 034010. [Google Scholar] [CrossRef]
- Hynes, A.; Hamann, A. Moisture deficits limit growth of white spruce in the west-central boreal forest of North America. For. Ecol. Manag. 2020, 461, 117944. [Google Scholar] [CrossRef]
- Menendez-Miguelez, M.; Alvarez-Alvarez, P.; Majada, J.; Canga, E. Effects of soil nutrients and environmental factors on site productivity in Castanea sativa Mill. coppice stands in NW Spain. New For. 2015, 46, 217–233. [Google Scholar]
- Zhou, Y.; Lei, Z.; Zhou, F.; Han, Y.; Yu, D.; Zhang, Y. Impact of climate factors on height growth of Pinus sylvestris var. mongolica. PLoS ONE 2019, 14, e0213509. [Google Scholar]
- Wilde, S.A. Forest soils, their properties and relation to silviculture. In Forest Soils, Their Properties and Relation to Silviculture; S. A. Wilde. Ronald Press: New York, NY, USA, 1958. [Google Scholar]
- Fitzpatrick, E.A. An Introduction to Soil Science; Longman Singapore Publishers Pte Limited: Singapore, 1995. [Google Scholar]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils; Pearson: London, UK, 2016. [Google Scholar]
- Aertsen, W.; Kint, V.; De Vos, B.; Deckers, J.; Van Orshoven, J.; Muys, B. Predicting forest site productivity in temperate lowland from forest floor, soil and litterfall characteristics using boosted regression trees. Plant Soil 2012, 354, 157–172. [Google Scholar]
- Subedi, S.; Fox, T.R. Predicting Loblolly Pine Site Index from Soil Properties Using Partial Least-Squares Regression. For. Sci. 2016, 62, 449–456. [Google Scholar] [CrossRef]
- Aydın, M.; Kılıç, Ş. Toprak Bilimi; Nobel Yayınları: Çankaya, Turkey, 2020. [Google Scholar]
- Binkley, D.; Fisher, R. Ecology and Management of Forest Soils; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Irmak, A. Orman Ekolojisi; Taş Matbaası: Istanbul, Turkey, 1970; p. 367. [Google Scholar]
- Berberoglu, S.; Evrendilek, F.; Ozkan, C.; Donmez, C. Modeling forest productivity using envisat MERIS data. Sensors 2007, 7, 2115–2127. [Google Scholar] [CrossRef] [PubMed]
- Gulsoy, S.; Suel, H.; Celik, H.; Ozdemir, S.; Ozkan, K. Modeling Site Productivity of Anatolian Black Pine Stands in Response to Site Factors in Buldan District, Turkey. Pak. J. Bot. 2014, 46, 213–220. [Google Scholar]
- Li, A.M.; Huang, C.Q.; Sun, G.Q.; Shi, H.; Toney, C.; Zhu, Z.L.; Rollins, M.G.; Goward, S.N.; Masek, J.G. Modeling the height of young forests regenerating from recent disturbances in Mississippi using Landsat and ICESat data. Remote Sens. Environ. 2011, 115, 1837–1849. [Google Scholar]
- Yue, C.F.; Kahle, H.P.; von Wilpert, K.; Kohnle, U. A dynamic environment-sensitive site index model for the prediction of site productivity potential under climate change. Ecol. Model. 2016, 337, 48–62. [Google Scholar] [CrossRef]
- Park, H.J.; Park, S.I.; Kim, H.S.; Lee, E.S.; Kim, H.J.; Lee, S.H. Selection of geographical factors using the random forest analysis method for developing the site index equation of Pinus densiflora stands in Republic of Korea. For. Sci. Technol. 2019, 15, 19–23. [Google Scholar]
- Dos Reis, A.A.; Franklin, S.E.; Acerbi, F.W.; Ferraz, A.C.; de Mello, J.M. Classification of Eucalyptus plantation Site Index (SI) and Mean Annual Increment (MAI) prediction using DEM-based geomorphometric and climatic variables in Brazil. Geocarto Int. 2022, 37, 1256–1273. [Google Scholar] [CrossRef]
- Scolforo, H.F.; McTague, J.P.; Burkhart, H.; Roise, J.; Alvares, C.A.; Stape, J.L. Site index estimation for clonal eucalypt plantations in Brazil: A modeling approach refined by environmental variables. For. Ecol. Manag. 2020, 466, 118079. [Google Scholar] [CrossRef]
- Li, X.Y.; Duan, A.G.; Zhang, J.G. Site index for Chinese fir plantations varies with climatic and soil factors in southern China. J. For. Res. 2022, 33, 1765–1780. [Google Scholar]
- McKenney, D.W.; Pedlar, J.H. Spatial models of site index based on climate and soil properties for two boreal tree species in Ontario, Canada. For. Ecol. Manag. 2003, 175, 497–507. [Google Scholar] [CrossRef]
- Jiang, H.Q.; Radtke, P.J.; Weiskittel, A.R.; Coulston, J.W.; Guertin, P.J. Climate- and soil-based models of site productivity in eastern US tree species. Can. J. For. Res. 2015, 45, 325–342. [Google Scholar]
- Restrepo, H.I.; Orrego, S.A.; Salazar-Uribe, J.C.; Bullock, B.P.; Montes, C.R. Using biophysical variables and stand density to estimate growth and yield of Pinus patula in Antioquia, Colombia. Open J. For. 2019, 9, 195–213. [Google Scholar]
- Swenson, J.J.; Waring, R.H.; Fan, W.; Coops, N. Predicting site index with a physiologically based growth model across Oregon, USA. Can. J. For. Res. 2005, 35, 1697–1707. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Abbr. | Minimum | Maximum | Mean | SE | SD | |
|---|---|---|---|---|---|---|---|
| Digitally extracted variables | Longitude (°) | LONG | 34.3 | 35.0 | 34.7 | <0.1 | 0.2 |
| Latitude (°) | LAT | 41.7 | 42.0 | 41.9 | <0.1 | 0.1 | |
| Elevation (m) | ELEV | 12.0 | 1352.0 | 630.9 | 21.8 | 324.6 | |
| Distance to the sea (m) | DTS | 692.5 | 24,935.8 | 12,279.0 | 385.3 | 5741.2 | |
| Transformed aspect | TRASP | 0.0 | 2.0 | 1.1 | <0.1 | 0.7 | |
| Slope (%) | SLP | 5.0 | 84.4 | 31.8 | 1.1 | 16.8 | |
| Vegetation index | NDVI | 0.6 | 0.7 | 0.7 | <0.1 | <0.1 | |
| Mean temperature (°) | MAT | 6.8 | 14.1 | 10.6 | 0.1 | 1.8 | |
| Minimum mean temperature (°) | MINMT | 2.5 | 11.3 | 7.0 | 0.1 | 2.2 | |
| Maximum mean temperature (°) | MAXMT | 11.5 | 17.5 | 14.8 | 0.1 | 1.5 | |
| Total precipitation (mm) | TP | 806.4 | 1072.9 | 925.4 | 3.6 | 54.3 | |
| Soil bulk density (kg/m3) | BD | 10.0 | 13.8 | 11.5 | 0.1 | 0.9 | |
| Soil field capacity (%) | FC | 30.0 | 40.0 | 33.5 | 0.1 | 2.0 | |
| Soil organic carbon (%) | OC | 0.8 | 2.8 | 1.6 | <0.1 | 0.4 | |
| Soil pH | pH | 5.3 | 6.7 | 6.0 | <0.1 | 0.3 | |
| Observed variables | Stand dominant height (m) | SDH | 14.6 | 33.4 | 22.8 | 0.2 | 3.7 |
| Stand age (year) | SA | 29.0 | 148.0 | 73.3 | 1.4 | 21.2 | |
| Soil depth (cm) | SDEPTH | 40.0 | 145.0 | 105.2 | 1.2 | 18.2 | |
| Sand content (%) | SAND | 21.0 | 90.0 | 51.3 | 0.9 | 14.3 | |
| Silt content (%) | SILT | 3.0 | 39.0 | 18.5 | 0.4 | 6.0 | |
| Clay content (%) | CLAY | 5.0 | 60.0 | 30.2 | 0.8 | 12.0 | |
| Available water capacity (%) | AWC | 1.9 | 23.3 | 12.3 | 0.2 | 3.3 | |
| Soil pH | pH | 3.8 | 7.4 | 5.3 | 0.1 | 0.8 | |
| Soil electrical conductivity (mS/cm) | ECe | 0.1 | 6.48 | 0.9 | 0.1 | 0.9 | |
| Soil organic carbon (%) | OC | 0.2 | 10.5 | 3.4 | 0.1 | 2.0 | |
| Soil lime content (%) | LIME | 0.0 | 17.4 | 1.9 | 0.1 | 2.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yener, I.; Guvendi, E. Predicting and Mapping Dominant Height of Oriental Beech Stands Using Environmental Variables in Sinop, Northern Turkey. Sustainability 2023, 15, 14580. https://doi.org/10.3390/su151914580
Yener I, Guvendi E. Predicting and Mapping Dominant Height of Oriental Beech Stands Using Environmental Variables in Sinop, Northern Turkey. Sustainability. 2023; 15(19):14580. https://doi.org/10.3390/su151914580
Chicago/Turabian StyleYener, Ismet, and Engin Guvendi. 2023. "Predicting and Mapping Dominant Height of Oriental Beech Stands Using Environmental Variables in Sinop, Northern Turkey" Sustainability 15, no. 19: 14580. https://doi.org/10.3390/su151914580