


Freeze–Thaw Cycles Have More of an Effect on Greenhouse Gas Fluxes than Soil Water Content on the Eastern Edge of the Qinghai–Tibet Plateau

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
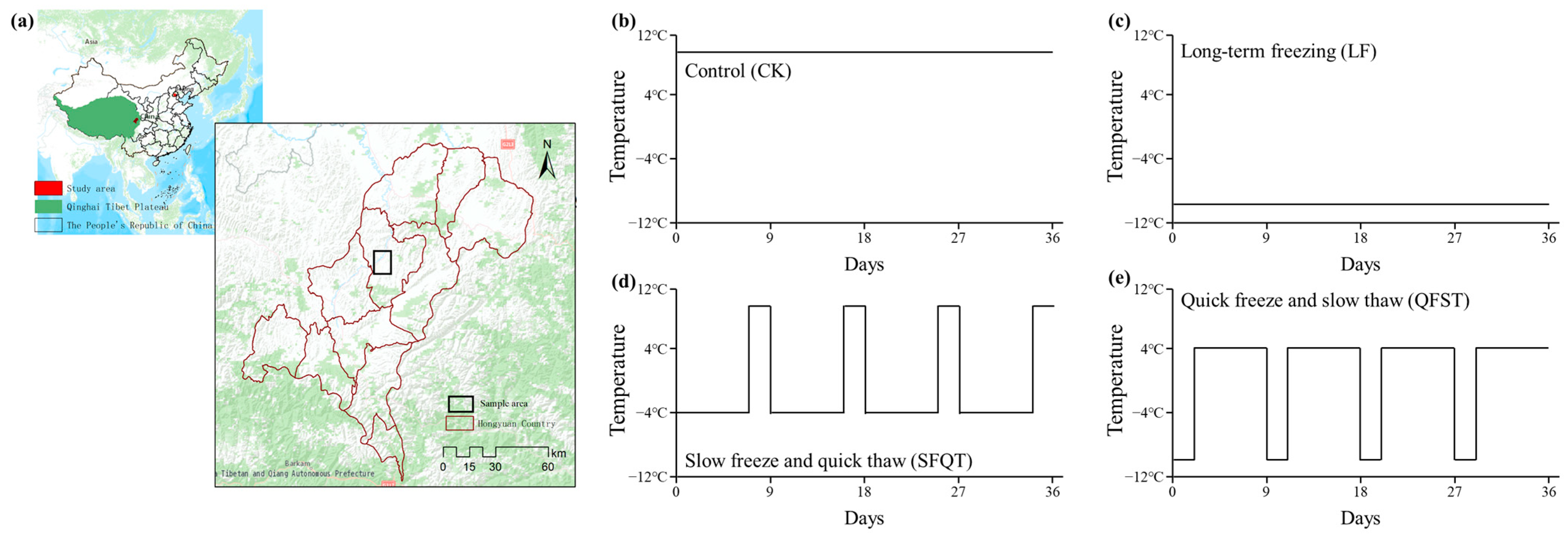
2.1. Site Description and Soil Collection
2.2. Microcosm Experiment
2.3. Soil Properties, Enzyme Activities, and Soil Microbial Biomass
2.4. Analysis of GHG Fluxes
2.5. Statistical Analyses
3. Results
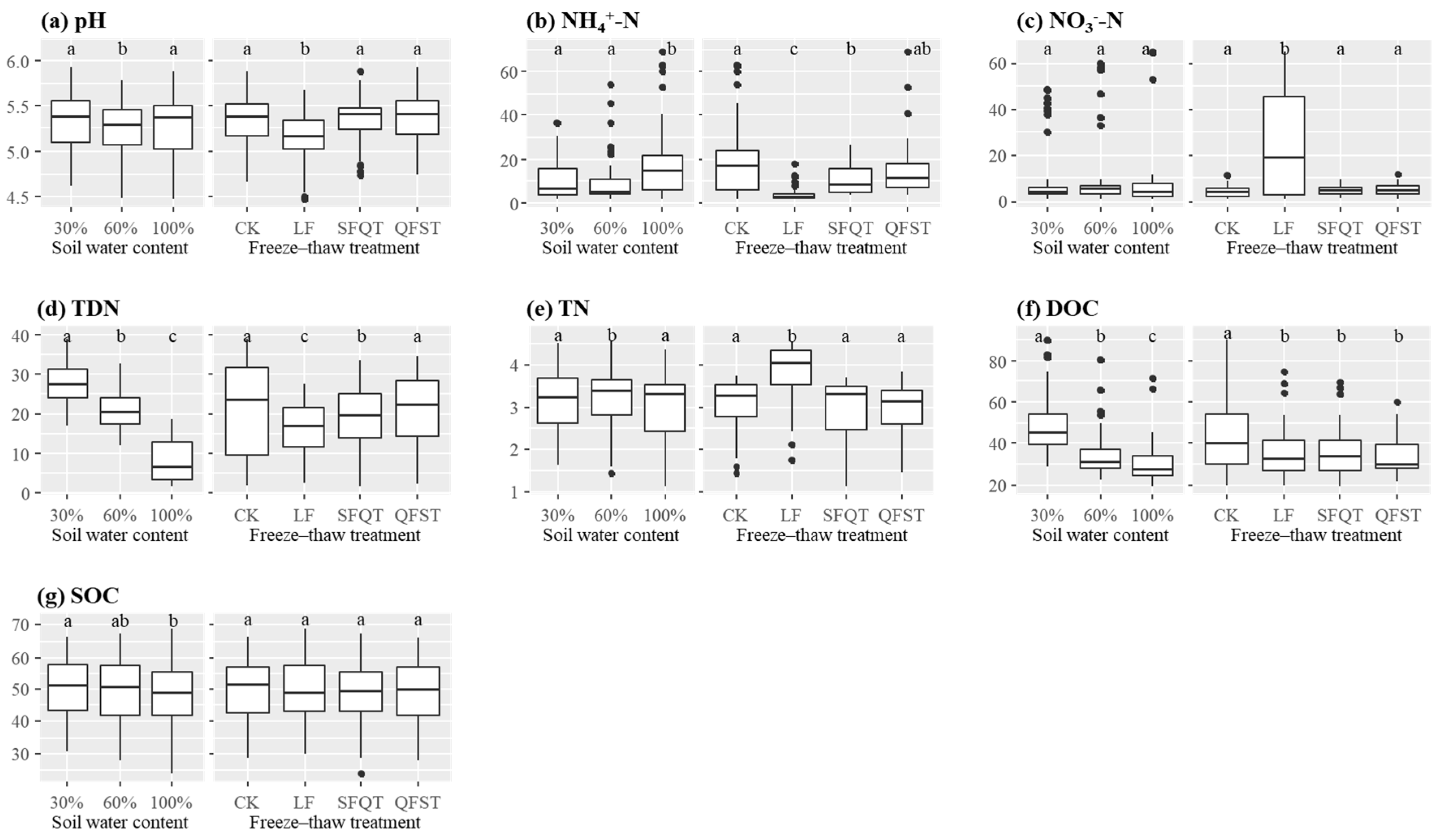
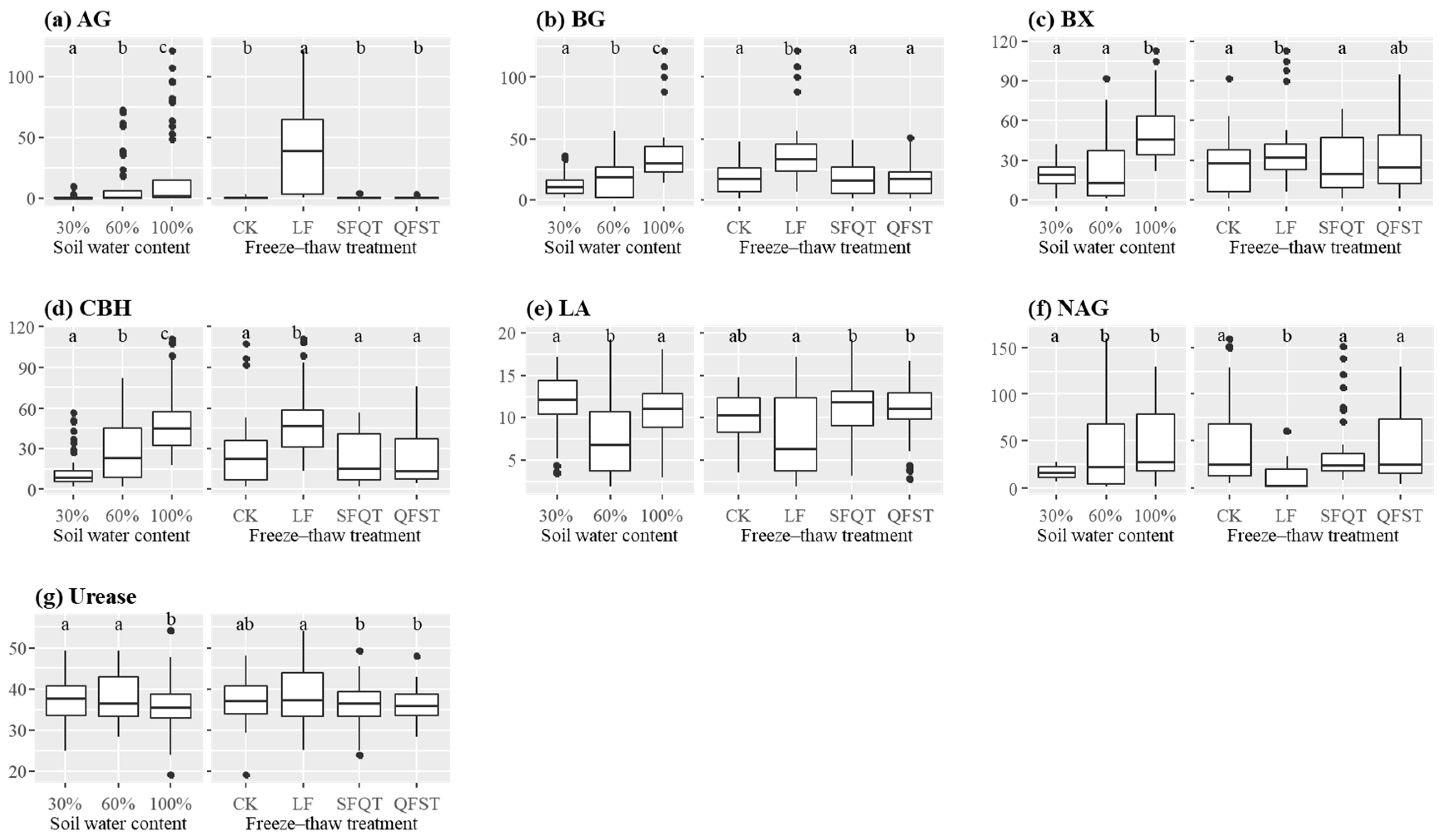
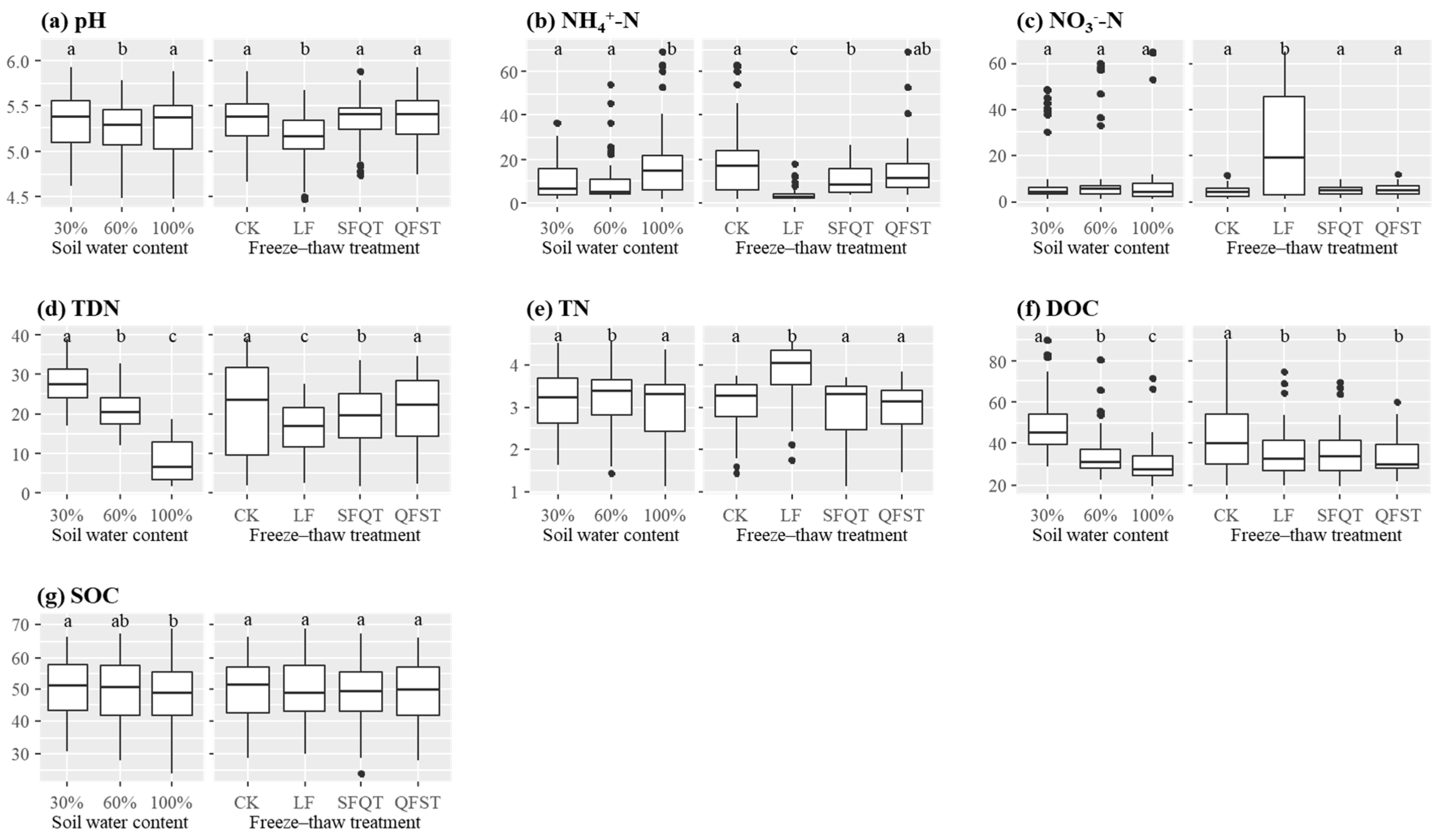
3.1. Soil Properties and Enzyme Activities
3.2. Soil Microbial Biomass and Soil GHG Fluxes
4. Discussion
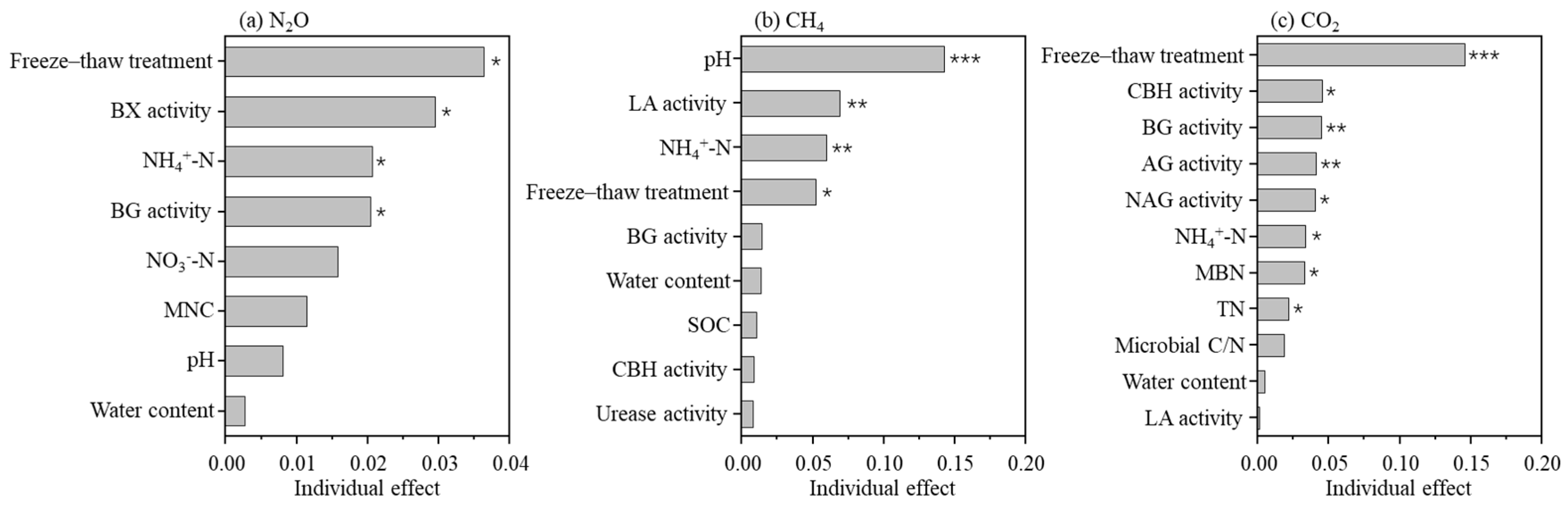
4.1. Freeze–Thaw Treatments Increase N2O Flux
4.2. Freeze–Thaw Treatments Reduce CH4 Flux
4.3. Freeze–Thaw Treatments Have Different Impacts on CO2 Flux
4.4. The Changes in Soil Nutrients, Enzyme Activities, and Soil Microbes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| QTP | Qinghai-Tibetan Plateau |
| GHG | greenhouse gas |
| N2O | nitrous oxide |
| CH4 | methane |
| CO2 | carbon dioxide |
| WC | water content |
| FTT | freeze–thaw treatment |
| CK | control |
| LF | long-term freezing |
| SFQT | slow freeze and quick thaw |
| QFST | quick freeze and slow thaw |
| TDN | total dissolved nitrogen |
| TN | total nitrogen |
| DOC | dissolved organic carbon |
| SOC | soil organic carbon |
| AG | αG1,4-glucosidase |
| BG | βG1,4-glucosidase |
| BX | βX1,4-xylosidase |
| CBH | cellobiohydrolase |
| LA | leucine aminopeptidase |
| NAG | βAG,4-N-acetylglucosaminidase |
| MBC | microbial biomass carbon |
| MBN | microbial biomass nitrogen |
| OLS | ordinary least squares |
References
- He, J.; Dong, S.; Shang, Z.; Sundqvist, M.K.; Wu, G.; Yang, Y. Above-belowground interactions in alpine ecosystems on the roof of the world. Plant Soil 2021, 458, 1–6. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, X.; Yu, H.; Liu, Q.; Xu, L. Land Use-Driven Changes in Ecosystem Service Values and Simulation of Future Scenarios: A Case Study of the Qinghai–Tibet Plateau. Sustainability 2021, 13, 4079. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Y.; Wu, J.; Li, S.; Zhang, B.; Zu, J.; Zhang, H.; Ding, M.; Paudel, B. Increasing sensitivity of alpine grasslands to climate variability along an elevational gradient on the Qinghai–Tibet Plateau. Sci. Total Environ. 2019, 678, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Zhang, Z.; Tian, J.; Kong, R.; Chen, X. Increasing Negative Impacts of Climatic Change and Anthropogenic Activities on Vegetation Variation on the Qinghai–Tibet Plateau during 1982–2019. Remote Sens. 2022, 14, 4735. [Google Scholar] [CrossRef]
- Chen, J.; Yan, F.; Lu, Q. Spatiotemporal Variation of Vegetation on the Qinghai–Tibet Plateau and the Influence of Climatic Factors and Human Activities on Vegetation Trend (2000–2019). Remote Sens. 2020, 12, 3150. [Google Scholar] [CrossRef]
- Duan, A.; Wu, G.; Zhang, Q.; Liu, Y. New proofs of the recent climate warming over the Tibetan Plateau as a result of the increasing greenhouse gases emissions. Chin. Sci. Bull. 2006, 51, 1396–1400. [Google Scholar] [CrossRef]
- Panchasara, H.; Samrat, N.H.; Islam, N. Greenhouse Gas Emissions Trends and Mitigation Measures in Australian Agriculture Sector—A Review. Agriculture 2021, 11, 85. [Google Scholar] [CrossRef]
- Jing, X.; Sanders, N.J.; Shi, Y.; Chu, H.; Classen, A.T.; Zhao, K.; Chen, L.; Shi, Y.; Jiang, Y.; He, J.S. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nat. Commun. 2015, 6, 8159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Noah, I.; Friedman, S.P. Review and evaluation of root respiration and of natural and agricultural processes of soil aeration. Vadose Zone J. 2018, 17, 1–47. [Google Scholar] [CrossRef] [Green Version]
- Schulz-Zunkel, C.; Krüger, F. Trace Metal Dynamics in Floodplain Soils of the River Elbe: A Review. J. Environ. Qual. 2009, 38, 1349–1362. [Google Scholar] [CrossRef]
- Ye, C.; Guo, Z.; Cai, C.; Wang, J.; Deng, J. Effect of water content, bulk density, and aggregate size on mechanical characteristics of Aquults soil blocks and aggregates from subtropical China. J. Soils Sediments 2017, 17, 210–219. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.; Zechmeister-Boltenstern, S. Greenhouse gas emissions from European soils under different land use: Effects of soil moisture and temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, K.; Liu, F.; Che, Z. Effects of Rainfall on the Characteristics of Soil Greenhouse Gas Emissions in the Wetland of Qinghai Lake. Atmos 2022, 13, 129. [Google Scholar] [CrossRef]
- Darenova, E.; Cater, M.; Pavelka, M. Different harvest intensity and soil CO2 efflux in sessile oak coppice forests. Iforest-Biogeosci. For. 2016, 9, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Q.; Qiao, Y.; Du, K.; Li, Z.; Tian, C.; Zhu, N.; Peifang, L.; Yue, Z.; Cheng, H.; et al. Trade-offs between high yields and soil CO2 emissions in semi-humid maize cropland in northern China. Soil Tillage Res. 2022, 221, 105412. [Google Scholar] [CrossRef]
- Grogan, P.; Michelsen, A.; Ambus, P.; Jonasson, S. Freeze–thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms. Soil Biol. Biochem. 2004, 36, 641–654. [Google Scholar] [CrossRef]
- Grogan, P.; Jonasson, S. Controls on annual nitrogen cycling in the understory of a subarctic birch forest. Ecology 2003, 84, 202–218. [Google Scholar] [CrossRef]
- Lipson, D.A.; Schmidt, S.K.; Monson, R.K. Links between microbial population dynamics and nitrogen availability in an alpine ecosystem. Ecology 1999, 80, 1623–1631. [Google Scholar] [CrossRef]
- Risk, N.; Snider, D.; Wagner-Riddle, C. Mechanisms leading to enhanced soil nitrous oxide fluxes induced by freeze–thaw cycles. Can. J. Soil Sci. 2013, 93, 401–414. [Google Scholar] [CrossRef]
- Xu, X. Effect of freeze–thaw disturbance on soil C and N dynamics and GHG fluxes of East Asia forests: Review and future perspectives. Soil Sci. Plant Nutr. 2022, 68, 15–26. [Google Scholar] [CrossRef]
- Wu, X.; Wang, F.; Li, T.; Fu, B.; Lv, Y.; Liu, G. Nitrogen additions increase N2O emissions but reduce soil respiration and CH4 uptake during freeze–thaw cycles in an alpine meadow. Geoderma 2020, 363, 114157. [Google Scholar] [CrossRef]
- Chen, Z.; Ge, S.; Zhang, Z.; Du, Y.; Yao, B.; Xie, H.; Liu, P.; Zhang, Y.; Wang, W.; Zhou, H. Soil Moisture but Not Warming Dominates Nitrous Oxide Emissions during Freeze–Thaw Cycles in a Qinghai–Tibetan Plateau Alpine Meadow with Discontinuous Permafrost. Front. Ecol. Evol. 2021, 9, 676027. [Google Scholar] [CrossRef]
- Bai, J.; Hou, P.; Jin, D.; Zhai, J.; Ma, Y.; Zhao, J. Habitat Suitability Assessment of Black-Necked Crane (Grus nigricollis) in the Zoige Grassland Wetland Ecological Function Zone on the Eastern Tibetan Plateau. Diversity 2022, 14, 579. [Google Scholar] [CrossRef]
- Saiya-Cork, K.; Sinsabaugh, R.; Zak, D. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Cuello, J.P.; Hwang, H.Y.; Gutierrez, J.; Kim, S.Y.; Kim, P.J. Impact of plastic film mulching on increasing greenhouse gas emissions in temperate upland soil during maize cultivation. Appl. Soil Ecol. 2015, 91, 48–57. [Google Scholar] [CrossRef]
- Bauer, D.F. Constructing Confidence Sets Using Rank Statistics. J. Am. Stat. Assoc. 1972, 67, 687–690. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J.; Rohlf, J.F. Biometry, 3rd ed.; Macmillan: New York, NY, USA, 1995. [Google Scholar]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Libby, M.D.; VanderZaag, A.C.; Gregorich, E.G.; Wagner-Riddle, C. An improved laboratory method shows that freezing intensity increases N2O emissions. Can. J. Soil Sci. 2020, 100, 136–149. [Google Scholar] [CrossRef]
- Yang, W.H.; Hall, S.J.; McNicol, G. Global gases. In Principles and Applications of Soil Microbiology, 3rd ed.; Gentry, T.J., Fuhrmann, J.J., Zuberer, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 557–579. [Google Scholar] [CrossRef]
- Horák, J.; Kotuš, T.; Toková, L.; Aydın, E.; Igaz, D.; Šimanský, V. A Sustainable Approach for Improving Soil Properties and Reducing N2O Emissions Is Possible through Initial and Repeated Biochar Application. Agronomy 2021, 11, 582. [Google Scholar] [CrossRef]
- Wick, B.; Veldkamp, E.; De Mello, W.; Keller, M.; Crill, P. Nitrous oxide fluxes and nitrogen cycling along a pasture chronosequence in Central Amazonia, Brazil. Biogeosciences 2005, 2, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.M.; Bork, E.W.; Chang, S.X.; Carlyle, C.N.; Ma, Z.; Döbert, T.F.; Kaliaskar, D.; Boyce, M.S. Adaptive multi-paddock grazing lowers soil greenhouse gas emission potential by altering extracellular enzyme activity. Agronomy 2020, 10, 1781. [Google Scholar] [CrossRef]
- Ma, Q.; Li, J.; Aamer, M.; Huang, G. Effect of chinese milk vetch (Astragalus sinicus L.) and rice straw incorporated in paddy soil on greenhouse gas emission and soil properties. Agronomy 2020, 10, 717. [Google Scholar] [CrossRef]
- Waldrop, M.; Firestone, M. Altered utilization patterns of young and old soil C by microorganisms caused by temperature shifts and N additions. Biogeochemistry 2004, 67, 235–248. [Google Scholar] [CrossRef]
- Zalman, C.; Keller, J.; Tfaily, M.; Kolton, M.; Pfeifer-Meister, L.; Wilson, R.; Lin, X.; Chanton, J.; Kostka, J.; Gill, A.; et al. Small differences in ombrotrophy control regional-scale variation in methane cycling among Sphagnum-dominated peatlands. Biogeochemistry 2018, 139, 155–177. [Google Scholar] [CrossRef]
- Weslien, P.; Kasimir Klemedtsson, Å.; Börjesson, G.; Klemedtsson, L. Strong pH influence on N2O and CH4 fluxes from forested organic soils. Eur. J. Soil Sci. 2009, 60, 311–320. [Google Scholar] [CrossRef]
- Wormald, R.M.; Rout, S.P.; Mayes, W.; Gomes, H.; Humphreys, P.N. Hydrogenotrophic methanogenesis under alkaline conditions. Front. Microbiol. 2020, 11, 614227. [Google Scholar] [CrossRef]
- Wilson, J.S.; Baldwin, D.S.; Rees, G.N.; Wilson, B.P. The effects of short-term inundation on carbon dynamics, microbial community structure and microbial activity in floodplain soil. River Res. Appl. 2011, 27, 213–225. [Google Scholar] [CrossRef]
- Willis, W.; Carlson, C.; Alessi, J.; Haas, H. Depth of freezing and spring run-off as related to fall soil-moisture level. Can. J. Soil Sci. 1961, 41, 115–123. [Google Scholar] [CrossRef]
- Sharma, S.; Szele, Z.; Schilling, R.; Munch, J.C.; Schloter, M. Influence of freeze-thaw stress on the structure and function of microbial communities and denitrifying populations in soil. Appl. Environ. Microbiol. 2006, 72, 2148–2154. [Google Scholar] [CrossRef] [Green Version]
- Seneesrisakul, K.; Sutabutr, T.; Chavadej, S. The Effect of Temperature on the Methanogenic Activity in Relation to Micronutrient Availability. Energies 2018, 11, 1057. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Song, C.; Hou, A.; Song, Y.; Zhu, X.; Cagle, G.A. Shifts in soil bacterial and archaeal communities during freeze-thaw cycles in a seasonal frozen marsh, Northeast China. Sci. Total Environ. 2018, 625, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Montzka, S.A.; Dlugokencky, E.J.; Butler, J.H. Non-CO2 greenhouse gases and climate change. Nature 2011, 476, 43–50. [Google Scholar] [CrossRef]
- Azizi-Rad, M.; Guggenberger, G.; Ma, Y.; Sierra, C.A. Sensitivity of soil respiration rate with respect to temperature, moisture and oxygen under freezing and thawing. Soil Biol. Biochem. 2022, 165, 108488. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, N.; Lu, Y.; Han, X.; Yang, Z. Effects of Supplementary Irrigation on Soil Respiration of Millet Farmland in a Semi-Arid Region in China. Atmos 2022, 13, 1584. [Google Scholar] [CrossRef]
- Elberling, B. Seasonal trends of soil CO2 dynamics in a soil subject to freezing. J. Hydrol. 2003, 276, 159–175. [Google Scholar] [CrossRef]
- Lv, J.; Liu, X.; Liu, H.; Wang, X.; Li, K.; Tian, C.; Christie, P. Greenhouse gas intensity and net annual global warming potential of cotton cropping systems in an extremely arid region. Nutr. Cycl. Agroecosyst. 2014, 98, 15–26. [Google Scholar] [CrossRef]
- Nelson, D.L. Carbohydrates and Glycobiology. In Lehninger Principles of Biochemistry, 8th ed.; W.H. Freeman: New York, NY, USA, 2021; p. 956. [Google Scholar]
- Miller, M.; Palojärvi, A.; Rangger, A.; Reeslev, M.; Kjøller, A. The use of fluorogenic substrates to measure fungal presence and activity in soil. Appl. Environ. Microbiol. 1998, 64, 613–617. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; Liu, R.; Du, Z.; Zhou, L.; Li, S.; Liu, H.; Shao, J.; Wang, J.; Nie, Y.; et al. Soil fungi and fine root biomass mediate drought-induced reductions in soil respiration. Funct. Ecol. 2020, 34, 2634–2643. [Google Scholar] [CrossRef]
- Gao, D.; Liu, Z.; Bai, E. Effects of in situ freeze-thaw cycles on winter soil respiration in mid-temperate plantation forests. Sci. Total Environ. 2021, 793, 148567. [Google Scholar] [CrossRef]
- Niboyet, A.; Bardoux, G.; Barot, S.; Bloor, J.M. Elevated CO2 mediates the short-term drought recovery of ecosystem function in low-diversity grassland systems. Plant Soil 2017, 420, 289–302. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, J.; Li, X.; Liu, C.; Lin, W.; Zheng, W.; Chen, Y.; Yang, Y. Nitrogen addition accelerates the nitrogen cycle in a young subtropical Cunninghamia lanceolata (Lamb.) plantation. Ann. For. Sci. 2019, 76, 31. [Google Scholar] [CrossRef] [Green Version]
- Gmach, M.R.; Cherubin, M.R.; Kaiser, K.; Cerri, C.E.P. Processes that influence dissolved organic matter in the soil: A review. Sci. Agric. 2019, 77, e20180164. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, S.; Huo, L.; Sun, G.; Lu, X.; Jiang, M.; Yu, X. Wetland saturation with introduced Fe (III) reduces total carbon emissions and promotes the sequestration of DOC. Geoderma 2018, 325, 141–151. [Google Scholar] [CrossRef]
- Song, Y.; Zou, Y.; Wang, G.; Yu, X. Altered soil carbon and nitrogen cycles due to the freeze-thaw effect: A meta-analysis. Soil Biol. Biochem. 2017, 109, 35–49. [Google Scholar] [CrossRef]
- Wu, Q.; Yue, K.; Ma, Y.; Hedenec, P.; Cai, Y.; Chen, J.; Zhang, H.; Shao, J.; Chang, S.X.; Li, Y. Contrasting effects of altered precipitation regimes on soil nitrogen cycling at the global scale. Glob. Change Biol. 2022, 28, 6679–6695. [Google Scholar] [CrossRef]
- Halmø, G.; Eimhjellen, K. Low temperature removal of nitrate by bacterial denitrification. Water Res. 1981, 15, 989–998. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L. A theoretical model of litter decay and microbial interaction. Ecol. Monogr. 2006, 76, 151–174. [Google Scholar] [CrossRef]
- Zhao, X.-Q.; Zhou, X.-M. Ecological basis of alpine meadow ecosystem management in Tibet: Haibei alpine meadow ecosystem research station. Ambio 1999, 28, 642–647. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, C.; Wang, Y.; Xu, Z.; Han, H.; Li, L.; Wan, S. Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland. Sci. Total Environ. 2013, 444, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Soil Enzyme Activity in a Mediterranean Forest after Six Years of Drought. Soil Sci. Soc. Am. J. 2010, 74, 838–851. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, P.; Buchmann, T.; Cesarz, S.; Eisenhauer, N.; Roscher, C. Fertilization, soil and plant community characteristics determine soil microbial activity in managed temperate grasslands. Plant Soil 2017, 419, 189–199. [Google Scholar] [CrossRef]
- Bhanwaria, R.; Singh, B.; Musarella, C.M. Effect of organic manure and moisture regimes on soil physiochemical properties, microbial biomass Cmic:Nmic:Pmic turnover and yield of mustard grains in arid climate. Plants 2022, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Tiziani, R.; Miras-Moreno, B.; Malacrinò, A.; Vescio, R.; Lucini, L.; Mimmo, T.; Cesco, S.; Sorgonà, A. Drought, heat, and their combination impact the root exudation patterns and rhizosphere microbiome in maize roots. Environ. Exp. Bot. 2022, 203, 105071. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WC | FTT | pH | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | TDN (mg kg−1) | TN (g kg−1) | DOC (mg kg−1) | SOC (g kg−1) |
|---|---|---|---|---|---|---|---|---|
| 30% | CK | 4.83 ± 0.09 ab | 19.1 ± 2.9 ab | 4.1 ± 0.4 abcd | 32.2 ± 1.6 a | 3.1 ± 0.2 ab | 57.5 ± 5.0 a | 50.4 ± 3.4 ab |
| LF | 4.62 ± 0.09 ab | 2.6 ± 0.2 c | 32.0 ± 5.2 e | 22.7 ± 0.8 bc | 3.8 ± 0.2 c | 48.3 ± 3.9 b | 50.1 ± 2.9 a | |
| SFQT | 5.11 ± 0.08 a | 7.7 ± 1.2 d | 4.5 ± 0.6 abcd | 26.8 ± 1.0 d | 2.8 ± 0.2 ad | 48.4 ± 3.7 b | 49.2 ± 2.9 abc | |
| QFST | 4.90 ± 0.11 a | 8.9 ± 1.2 de | 3.6 ± 0.6 ab | 29.1 ± 0.8 ad | 3.0 ± 0.2 ad | 42.8 ± 2.9 bc | 49.5 ± 3.1 ab | |
| 60% | CK | 4.71 ± 0.10 b | 18.7 ± 5.1 abe | 5.2 ± 0.6 cd | 26.0 ± 1.3 bd | 3.0 ± 0.2 abd | 39.5 ± 5.2 cd | 49.3 ± 3.5 abc |
| LF | 4.58 ± 0.09 c | 2.7 ± 0.2 c | 25.1 ± 7.5 ace | 16.5 ± 0.8 e | 3.9 ± 0.2 c | 34.4 ± 2.5 d | 49.7 ± 2.7 ab | |
| SFQT | 5.10 ± 0.08 ab | 9.7 ± 2.1 ade | 4.8 ± 0.4 abcd | 19.8 ± 1.1 f | 3.3 ± 0.1 be | 32.1 ± 2.1 de | 48.1 ± 3.4 abc | |
| QFST | 4.81 ± 0.08 ab | 9.2 ± 1.7 de | 5.4 ± 0.6 cd | 22.1 ± 1.0 cf | 3.1 ± 0.1 abd | 29.4 ± 1.5 ef | 48.0 ± 3.8 abc | |
| 100% | CK | 5.16 ± 0.10 a | 22.3 ± 7.1 abde | 3.3 ± 1.0 cbd | 7.1 ± 1.6 g | 3.0 ± 0.2 ad | 36.8 ± 4.7 cde | 47.8 ± 2.3 bc |
| LF | 4.57 ± 0.10 c | 7.0 ± 1.5 de | 18.4 ± 7.4 abcde | 8.2 ± 1.1 g | 3.6 ± 0.3 e | 26.5 ± 1.9 f | 48.2 ± 3.2 abc | |
| SFQT | 4.78 ± 0.10 a | 15.7 ± 2.2 ab | 4.9 ± 0.6 cd | 8.9 ± 1.7 g | 2.9 ± 0.2 ad | 28.1 ± 2.0 ef | 45.7 ± 3.1 c | |
| QFST | 4.89 ± 0.08 a | 28.7 ± 4.9 b | 6.2 ± 0.9 ac | 8.8 ± 1.7 g | 2.8 ± 0.2 d | 29.7 ± 1.8 ef | 48.0 ± 3.0 abc | |
| Effect of WC | 0.214 | 0.003 | 0.641 | <0.001 | 0.490 | <0.001 | 0.388 | |
| Effect of FTT | 0.001 | <0.001 | 0.036 | 0.024 | <0.001 | 0.099 | 0.852 | |
| Effect of WC × FTT | 0.799 | 0.144 | 0.029 | 0.447 | 0.998 | 0.810 | 1.000 | |
| WC | FTT | AG | BG | BX | CBH | LA | NAG | Urease |
|---|---|---|---|---|---|---|---|---|
| 30% | CK | 0.6 ± 0.1 a | 11.6 ± 2.0 ab | 15.3 ± 4.1 abcd | 6.1 ± 0.8 a | 10.2 ± 1.0 abc | 14.8 ± 1.6 a | 37.2 ± 1.0 abcde |
| LF | 3.3 ± 0.7 b | 21. 6 ± 2.7 cd | 23.4 ± 1.9 abcd | 32.4 ± 4.5 bc | 14.0 ± 0.6 d | 19.1 ± 1.9 ab | 38.6 ± 1.7 ab | |
| SFQT | 0.7 ± 0.1 ac | 9.7 ± 1.8 ab | 18.0 ± 1.7 ab | 8.1 ± 0.7 d | 12.2 ± 1.0 ade | 15.2 ± 1.5 ab | 36.5 ± 1.5 acde | |
| QFST | 0.8 ± 0.1 acd | 9.0 ± 1.4 ab | 17.9 ± 2.0 abcd | 8.6 ± 1.3 ade | 11.6 ± 1.0 abd | 16.6 ± 1.6 ab | 37.8 ± 1.5 abcd | |
| 60% | CK | 1.0 ± 0.1 acd | 14.2 ± 4.0 ac | 27.1 ± 8.7 acef | 24.3 ± 4.2 b | 8.7 ± 0.8 efh | 73.0 ± 16.2 c | 40.8 ± 1.8 b |
| LF | 45.2 ± 5.6 e | 35.6 ± 4.8 ef | 26.8 ± 4.2 cdeg | 45.5 ± 5.5 cfg | 3.5 ± 0.3 h | 1.6 ± 0.3 d | 38.0 ± 1.8 abcd | |
| SFQT | 1.3 ± 0.2 df | 12.5 ± 3.7 ab | 16.9 ± 7.6 bd | 21.5 ± 5.9 bdeh | 9.5 ± 1.5 abcefg | 54.8 ± 14.0 ce | 37.0 ± 1.8 acde | |
| QFST | 1.1 ± 0.2 cd | 10.7 ± 3.0 b | 21.1 ± 7.9 bdg | 21.2 ± 5.9 beh | 8.8 ± 1.1 bcfg | 34.2 ± 10.2 bf | 35.3 ± 1.3 cde | |
| 100% | CK | 2.1 ± 0.2 bf | 27.1 ± 2.4 de | 36.6 ± 3.1 efg | 50.9 ± 8.8 cfg | 11.3 ± 0.7 abef | 49.8 ± 13.0 e | 33.4 ± 1.5 e |
| LF | 79.1 ± 6.6 g | 58.7 ± 10.2 f | 61.4 ± 8.8 h | 67.8 ± 8.0 f | 7.3 ± 1.0 cg | 11.4 ± 5.5 ad | 39.0 ± 2.5 abc | |
| SFQT | 2.1 ± 0.3 bf | 29.7 ± 3.4 e | 47.3 ± 4.8 fh | 38.6 ± 3.8 gh | 11.4 ± 1.0 abef | 48.8 ± 10.2 cef | 35.0 ± 1.6 de | |
| QFST | 2.0 ± 0.2 bf | 31.6 ± 3.5 e | 54.6 ± 5.3 h | 43.5 ± 6.1 gh | 12.1 ± 0.5 ab | 73.3 ± 11.3 ce | 35.3 ± 0.8 cde | |
| Effect of WC | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.002 | 0.186 | |
| Effect of FTT | <0.001 | <0.001 | 0.249 | <0.001 | 0.046 | <0.001 | 0.447 | |
| Effect of WC × FTT | 0.861 | 0.712 | 0.698 | 0.717 | 0.002 | <0.001 | 0.296 | |
| WC | FTT | MBC | MBN | Microbial C/N | N2O | CH4 | CO2 |
|---|---|---|---|---|---|---|---|
| 30% | CK | 490.1 ± 31.2 a | 25.4 ± 2.5 ab | 21.6 ± 2.6 abc | 0.81 ± 0.08 abcd | 0.90 ± 0.05 abc | 0.77 ± 0.07 abc |
| LF | 603.0 ± 27.3 b | 27.4 ± 3.1 a | 24.7 ± 2.4 abc | 24.03 ± 10.12 efg | 0.81 ± 0.02 abdef | 0.55 ± 0.01 de | |
| SFQT | 565.8 ± 21.7 b | 17.8 ± 2.4 cd | 40.5 ± 6.8 d | 1.60 ± 0.55 abe | 0.81 ± 0.02 def | 1.29 ± 0.08 f | |
| QFST | 600.7 ± 18.0 b | 29.6 ± 3.0 a | 22.7 ± 2.3 ab | 2.71 ± 0.77 cdfh | 0.82 ± 0.02 ade | 0.67 ± 0.04 a | |
| 60% | CK | 382.6 ± 29.1 cde | 15 ± 2.9.0 bcde | 34.3 ± 5.1 ad | 0.71 ± 0.15 ac | 0.84 ± 0.01 abcdf | 1.06 ± 0.11 fgh |
| LF | 458.6 ± 13.9 acd | 20.6 ± 3.0 abcde | 27.9 ± 3.9 abd | 14.16 ± 7.01 bdefhgi | 0.81 ± 0.02 de | 0.55 ± 0.01 d | |
| SFQT | 438.4 ± 17.5 acd | 18.4 ± 1.1 ce | 24.5 ± 1.4 a | 9.18 ± 3.04 gi | 0.79 ± 0.03 def | 1.34 ± 0.17 fg | |
| QFST | 442.4 ± 28.3 ac | 22.7 ± 2.1 abe | 21.2 ± 2.2 abc | 6.74 ± 2.31 fghi | 0.79 ± 0.02 de | 0.92 ± 0.06 bh | |
| 100% | CK | 366.6 ± 38.9 def | 23.0 ± 2.8 abce | 17.7 ± 1.8 ce | 5.63 ± 3.25 abcdhi | 0.86 ± 0.01 bc | 0.88 ± 0.10 abch |
| LF | 359.5 ± 17.4 ef | 27.5 ± 3.1 a | 14.8 ± 1.6 e | 1.46 ± 0.38 abcd | 0.80 ± 0.01 e | 0.53 ± 0.01 d | |
| SFQT | 341.1 ± 20.8 ef | 17.8 ± 1.1 cde | 19.5 ± 1.1 bc | 2.22 ± 0.84 abcdefh | 0.86 ± 0.03 c | 0.87 ± 0.13 abcegh | |
| QFST | 311.2 ± 27.9 f | 15.2 ± 1.6 d | 21.3 ± 1.7 bc | 0.81 ± 0.11 ac | 0.85 ± 0.01 bcf | 0.78 ± 0.03 c | |
| Effect of WC | <0.001 | 0.009 | <0.001 | 0.036 | 0.106 | 0.035 | |
| Effect of FTT | 0.271 | 0.044 | 0.103 | 0.028 | 0.010 | <0.001 | |
| Effect of WC × FTT | 0.487 | 0.007 | 0.051 | 0.009 | 0.857 | 0.074 | |
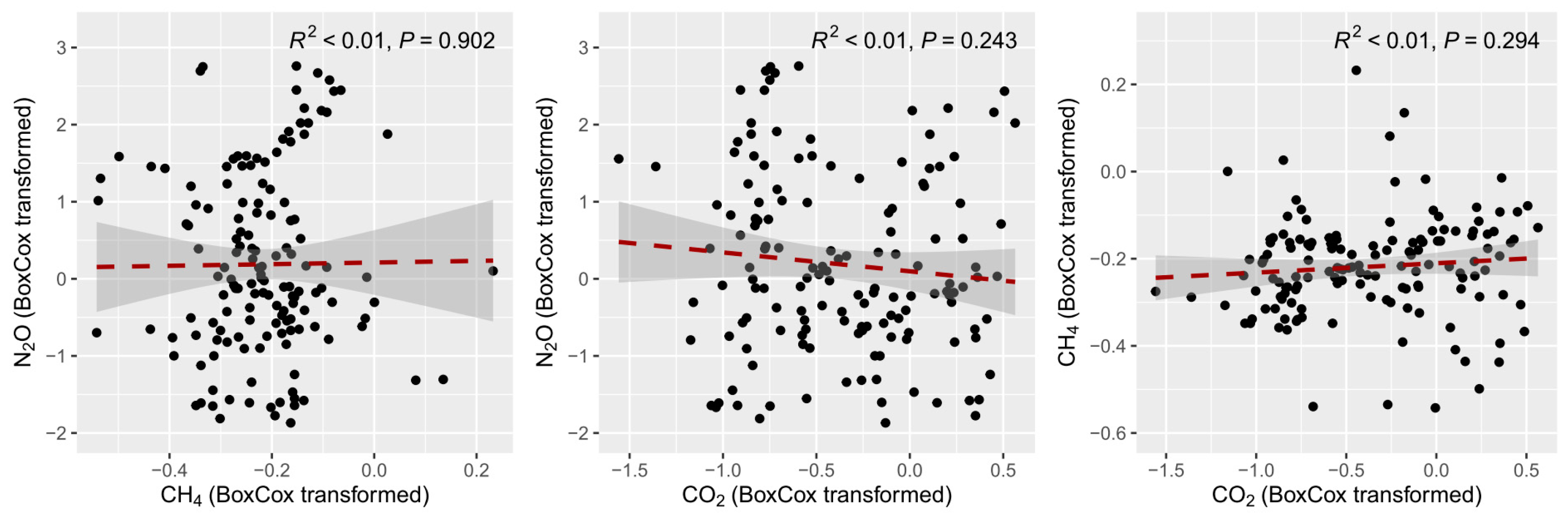
| Variables | N2O | CH4 | CO2 | |||
|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | |
| pH | −0.173 | 0.039 | −0.423 | <0.001 | −0.012 | 0.891 |
| NH4+-N | −0.211 | 0.011 | 0.278 | 0.001 | 0.372 | <0.001 |
| NO3−-N | 0.205 | 0.014 | 0.076 | 0.367 | −0.142 | 0.089 |
| TDN | 0.095 | 0.258 | 0.020 | 0.814 | 0.106 | 0.204 |
| TN | 0.051 | 0.543 | −0.146 | 0.080 | −0.311 | <0.001 |
| DOC | 0.061 | 0.469 | 0.082 | 0.328 | 0.103 | 0.221 |
| SOC | 0.075 | 0.370 | 0.201 | 0.016 | 0.012 | 0.886 |
| AG | −0.019 | 0.824 | −0.032 | 0.705 | −0.425 | <0.001 |
| BG | −0.216 | 0.009 | −0.207 | 0.013 | −0.422 | <0.001 |
| BX | −0.265 | 0.001 | −0.016 | 0.847 | −0.114 | 0.172 |
| CBH | −0.132 | 0.114 | −0.180 | 0.031 | −0.396 | <0.001 |
| LA | 0.056 | 0.504 | 0.281 | 0.001 | 0.180 | 0.031 |
| NAG | −0.009 | 0.917 | 0.111 | 0.187 | 0.379 | <0.001 |
| Urease | 0.068 | 0.421 | 0.191 | 0.022 | 0.032 | 0.703 |
| MBC | 0.213 | 0.010 | −0.051 | 0.547 | -0.096 | 0.253 |
| MBN | 0.017 | 0.836 | −0.078 | 0.356 | −0.304 | <0.001 |
| Microbial C/N | 0.121 | 0.147 | 0.031 | 0.716 | 0.257 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Qin, M.; Yang, X.; Bai, W.; Yao, Y.; Wang, J. Freeze–Thaw Cycles Have More of an Effect on Greenhouse Gas Fluxes than Soil Water Content on the Eastern Edge of the Qinghai–Tibet Plateau. Sustainability 2023, 15, 928. https://doi.org/10.3390/su15020928
Zhao S, Qin M, Yang X, Bai W, Yao Y, Wang J. Freeze–Thaw Cycles Have More of an Effect on Greenhouse Gas Fluxes than Soil Water Content on the Eastern Edge of the Qinghai–Tibet Plateau. Sustainability. 2023; 15(2):928. https://doi.org/10.3390/su15020928
Chicago/Turabian StyleZhao, Shanshan, Mingsen Qin, Xia Yang, Wenke Bai, Yunfeng Yao, and Junqiang Wang. 2023. "Freeze–Thaw Cycles Have More of an Effect on Greenhouse Gas Fluxes than Soil Water Content on the Eastern Edge of the Qinghai–Tibet Plateau" Sustainability 15, no. 2: 928. https://doi.org/10.3390/su15020928
APA StyleZhao, S., Qin, M., Yang, X., Bai, W., Yao, Y., & Wang, J. (2023). Freeze–Thaw Cycles Have More of an Effect on Greenhouse Gas Fluxes than Soil Water Content on the Eastern Edge of the Qinghai–Tibet Plateau. Sustainability, 15(2), 928. https://doi.org/10.3390/su15020928