Abstract
To satisfy the increasing demand for the protein supply of the growing global population, cultivated meat is being garnered significant attention that can meet consumer needs with a reduced impact on the planet nowadays. Due to being a nascent industry, there are still many hurdles to overcome such as cost reduction. This study aims to investigate the effect of microbiota-derived postbiotics to reduce the need for growth factors (GFs) or fetal bovine serum (FBS), as the main cost drivers of this technology. For this purpose, postbiotic Biftek-1 (B-1) was extracted from Enterococcus hirae, an isolate from cow milk microbiota, and bovine satellite cells (bSCs) were isolated from muscle biopsies, then identified by flow cytometry and immunofluorescence assay. The effects of different doses of GFs and B-1 on cell viability and myogenic profile of bSCs were evaluated. According to the results, low concentrations of GFs (40 ng/mL IGF1 and 20 ng/mL FGF2) combined with our growth medium supplement B-1 significantly increased bSCs proliferation, similar to FBS, but did not lead to any significant changes on their myogenic profiles. To the best of our knowledge, this study is the first attempt to use the postbiotics on cultivated meat technology.
1. Introduction
Cultivated meat, also known as cultured meat, is a breakthrough technology for producing genuine meat without slaughtering animals. It has emerged as a potential tool for global meat production with positive attributes such as promoting sustainability and animal welfare by reducing carbon emission rates associated with traditional animal husbandry [1]. Production of cultured meat using animal muscle cells grown in bioreactors is a promising technology for sustainable meat industry. However, cultivating the cells outside of the host requires a suitable cell culture medium that contains different factors and components needed for cell growth. In addition to basal medium, specific factors such as fetal bovine serum (FBS) or recombinant growth factors (GFs) have to be used to permit the long-term maintenance and proliferation of cells [2]. Therefore, scaling up this process to produce tons of meat at a reasonable cost is going to be difficult. Although more than 80 start-ups were declared with a focus on cultured meat productions in 2021, no industrial-scale production has been shown yet [3]. An optimistic projection for the cost of cultivated meat is 63 USD/kg [4], a considerably higher price than that of conventional meat, which averages at 14.26 USD/kg as of September 2023 [5]. The main reason behind this high market value for cultivated meat is the cost of FBS, which is estimated to be 55–95% of the final price [2]. In addition to financial drawbacks, production of FBS also raises scientific and ethical concerns. Since FBS is an animal product, it poses risks of containing microorganisms such as bacteria or viruses, or potentially harmful metabolites such as endotoxins. Additionally, its batch-to-batch variation is another important issue to deal with in both life sciences and the cultivated meat industry. Moreover, during the production process of FBS, not only the mother but also the fetus is slaughtered which leads to ethical problems [6]. Thus, efforts to lower the costs and ethical impacts of FBS are needed.
In recent years, recombinant GFs such as basic fibroblast growth factor 2 (FGF2), transforming growth factor β (TGF- β), and insulin-like growth factor 1 (IGF1) are used as serum-free alternatives to FBS [7,8]. However, they are still the main cost drivers of basal media, estimated to contribute more than 95% of the total cost [9]. Today, a milligram of FGF2, an essential component of the serum-free growth medium for muscle stem cell maintenance, is priced at EUR 750–7300 according to its production method, purity, and origin. Considering the volume of growth medium and GFs required to produce 1 kg of cultivated meat (50 L) to achieve a cell yield that is comparable to that of FBS, new scenarios to lower the cost of GFs are still needed to make culture meat comparable with conventional meat [10,11]. Recently, unique bio-production approaches such as chicken bioreactors to produce GFs in egg whites, TGF-β in silkworm cocoons, and FGF2 in transgenic fruit flies have been applied by different companies and research groups [12]. Synthetic production of GFs using Escherichia coli-based open cell-free synthesis has been also developed as another promising approach [13]. However, these systems are still more costly for culture meat companies. Therefore, novel strategies for manufacturing of culture meat at a reduced cost by replacing GFs with less expensive ones have still gained attention. Nowadays, researchers are looking to replace growth factors with plant-based alternatives [14] or with bioactive compounds such as cyanobacteria [15] and algae extracts [8].
Similar to these implementation approaches with a different resource, the current study was designed to investigate whether or not metabolic byproducts of microbiota-derived commensal microorganisms called postbiotics would reduce or eliminate the need for GFs for the bovine satellite cell (bSC) proliferation. Postbiotics, also known as metabolites or mediators, refer to “soluble factors secreted by living bacteria or released by bacterial lysis” including short-chain fatty acids (SCFAs), exopolysaccharides, functional proteins and enzymes, cell wall and cell surface proteins, etc. Although the main mechanism of action by which postbiotics exert specific effects has not been fully clarified, available scientific evidence points out that postbiotics can modulate the host metabolism and signaling pathways such as regulation of skeletal muscle metabolism and mass [16,17,18]. Due to the significant roles of postbiotics, the main goal of this study is to evaluate the microbiota derived-postbiotics to increase the effectiveness of GFs against FBS. To the best of our knowledge, this study is the first attempt to use the postbiotics on cultivated meat technology.
2. Materials and Methods
2.1. Bacterial Strain, Culture Medium and Growth Conditions
The strain Enterococcus hirae EIR/CM-1 (NCBI GenBank Accession Number: OR144426) (EIR; the code of our culture collection, CM; cow milk, 1; the first isolate from the sample), isolated from lactating Holstein cow milk and identified by 16S ribosomal-RNA subunit gene sequencing, was kindly obtained from the Pharmabiotic Technologies Research Laboratory, Department of Biology, Faculty of Science, Ankara University (Turkey). For isolation of the strain, 1 mL cow milk samples were serially ten-fold diluted, and pour-plated on De Man, Rogosa, and Sharpe agar (MRS, Merck, Darmstadt, Germany) plates. After the incubation period at 37 °C for 48 h, bacterial colonies were randomly selected, and pure colonies were maintained in MRS broth supplemented with 50% glycerol at −80 °C for long-term storage [19]. The sequence of 16S rRNA gene region (1.492 bp) searched on BLAST against GenBank Bacteria and Archaea 16S ribosomal RNA sequences database most closely matched with Enterococcus hirae with 99% similarity and registered in NCBI with the accession number of OR144426. The experimental protocol for the isolation of the bacterial strain from cow milk microbiota was approved by the Ankara University (Turkey) Institutional Animal Care and Use Committee (protocol number 2017-7-60).
2.2. Preparation of the Microbiota-Derived Postbiotics
The active culture of E. hirae EIR/CM-1 adjusted to 109 CFU/mL was inoculated into MRS broth media and incubated at 37 °C for 24 h (late log phase) under aerobic conditions. Following the incubation protocol, postbiotics secreted into the culture medium were obtained using centrifugation (Benchtop Microfuge 20R, Beckman Coulter, München, Germany) at 4000× g, 4 °C for 20 min. The cell-free supernatant including postbiotics was then collected by filtration through a 0.22 μm cellulose acetate membrane (Sartorius, Goettingen, Germany), and freeze-dried (freezing conditions of −20 °C, a vacuum pressure of 0.120 mB, and a condenser temperature of −58 °C; Martin Christ Gefriertrocknungsanlagen GmbH, Harz, Germany). The powdered form of postbiotics was suspended in sterile dH2O at a final concentration of 100 mg/mL as growth medium supplement B-1 and stored at −20 °C until cell culture studies were conducted [20].
2.3. Isolation of Bovine Muscle Satellite Cells (bSCs)
Bovine muscle satellite cells (bSCs) were isolated from fresh muscle biopsies by enzymatic digestion protocol adapted from Skrivergaard et al. (2021) and Ding et al. (2018) [21,22]. Briefly, muscle biopsies were obtained from young male Brown Swiss cattle (aged 20–24 months), washed twice with 70% ethanol to remove any potential contaminants, and then stored in DMEM High Glucose (HG, Thermo Fisher Scientific, Waltham, MA, USA, #10313021) containing 10% FBS (Thermo Fisher Scientific, Waltham, MA, USA, #10500-064), 1% L-Glutamine (Biological Industries, Cromwell, CT, USA, #03-020-1B), and 1% penicillin-streptomycin solution (Biological Industries, USA, #03-031-1B) at 4 °C for 24 h. Muscle tissue was finely minced with a small sterile scissor and incubated in digestion medium containing DMEM HG, 1% Pen-Strep solution, 2.5 mM CaCl2, 108 U/mL Collagenase (Gibco, #17101-015), and 4.8 mg/mL Dispase II (Sigma Aldrich, St. Louis, MO, USA, #D4693) at 37 °C for 30 min on a shaker (1 g of tissue/10 mL). Following the enzymatic digestion, the cells were transferred to Gentle MACS C Tubes (Miltenyi Biotech, Bergisch Gladbach, Germany, #130-093-237) and homogenized by using the Gentle MACS dissociator. The enzyme activity was terminated by adding 10 mL neutralization culture medium (DMEM HG containing 20% FBS, 1% L-Glutamine, and 1% penicillin-streptomycin solution). The suspension was passed through a series of strainers (100 µm, 70 µm, 40 µm) and centrifuged at 443× g for 5 min at room temperature (RT). The resulting pellet was seeded in T25 flask coated with 0.5% Matrigel® Matrix Basement Membrane (Corning, Corning, NY, USA) and incubated under standard conditions (at 37 °C, in a 5% CO2 atmosphere). The experimental protocol for the isolation of the bSCs from cattle biopsies was approved by the Selcuk University (Konya, Turkey) Institutional Animal Care and Use Committee (protocol number 2021-92).
2.4. Identification of the bSCs by Flow Cytometry and Immunofluorescence Assay
The cells were detached from the culture flasks by trypsinization (0.25%) and centrifuged at 200× g for 5 min at RT. The pellets were suspended in 5% BSA (Sigma Aldrich, St. Louis, MO, USA) and 0.05% sodium azide (Sigma Aldrich, USA) in 1X PBS. The cell suspensions were incubated with the following antibodies for 40 min at 4 °C: PE Anti-human CD29 (Biolegend, San Diego, CA, USA, #303004), PE-Cyanine 7 anti-human CD56 (Tonbo Bioscience, Cologne, Germany, #60-0564-T025), FITC mouse anti-sheep CD31 (BioRad, Hercules, CA, USA, #MCA1097F), and FITC mouse anti-sheep CD45 (BioRad, #MCA2220F). Upon the completion of incubation, cells were washed with 1X PBS, centrifuged, resuspended in 200 μL 1X PBS, and analyzed using the ACEA Novocyte flow cytometer (Agilent Technologies, Santa Clara, CA, USA) [22].
For the immunofluorescense assay described by Ding et al. (2018) [22], cells were fixed with absolute methanol, blocked in 1X PBS-T containing 5% BSA, and then stained with the following antibodies at 4 °C overnight: Anti-Desmin (Sigma Aldrich #D1033) or Anti-Pax-3/7 (Santa Cruz, Santa Cruz, CA, USA #365843). Then, the cells were incubated with Alexa Fluor 488-labeled secondary antibody for 1 h at RT. DAPI (Thermo, #P36931) was used to stain nuclei for counter staining. The images were acquired with a ZEISS Primovert Microscope with 40X magnification.
2.5. Evaluation of the Effects of GFs and Their Combinations with B-1
The bSCs were maintained under standard conditions (at 37 °C, in a 5% CO2 atmosphere) in DMEM HG medium with 10% FBS, 1% penicillin-streptomycin, and 1% L-glutamate (treatment medium). The cells were harvested and passaged when they reached 80% confluence. The cells at passage two were detached with 0.25% Trypsin/EDTA (Sarstedt, Nümbrecht, Germany),USA), counted (Bio-Rad, TC20 Automated Cell Counter), and seeded onto 96-well plates coated with 0.5% Matrigel at 10,000 cells/well density. The bSCs were incubated in the following media formulations: HAM’s F12 with 10% FBS for FBS group, only HAM’s F12 for No FBS group, and HAM’s F12 with different concentrations of GFs, separately, as GF groups. BGF groups (combinations of selected GFs with supplement B-1) were incubated in 0.5 and 1 mg/mL B-1, 40 ng/mL IGF1 (Zhuhai Pareto Pharmaceutical Science Co. Ltd., Zhuhai, China), and 20 ng/mL FGF2 (Zhuhai Pareto Pharmaceutical Science Co. Ltd.). At 72 h after the initiation of the treatments, 10% MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide, Sigma Aldrich, USA) was added into the wells. The plates were incubated at 37 °C for 4 h. The media were discarded, and formazan was dissolved in DMSO (Dimethyl sulfoxide, Sigma Aldrich, USA). The OD values were measured at 570 nm.
2.6. Quantitative Real-Time PCR (qRT-PCR)
For the treatment procedure for bSCs, 3 and 6 days were used due to the observation of significant differences between No FBS and 10% FBS groups, and the prevention of cell overcrowding, based on our preliminary studies. Following the treatment protocol as given in Section 2.5, total RNA was isolated from bSCs (at day 3 and 6 of the treatment procedure) using the PureZOL RNA Isolation Reagent (BioRad, #7326890), according to the manufacturer’s protocol. The cDNAs were synthesized with the iScript cDNA Synthesis Kit (BioRad, #1708891), following the manufacturer’s guidelines. Reaction ingredients [0.5 μL of cDNA, 5 μL of SsoAdvanced Universal SYBR Green Supermix (BioRad, #1725270), and 10 μM from each primer pair listed in Table 1] were mixed for qRT-PCR analysis and run in LightCycler® 480 (Roche, Basel, Switzerland) [15,16]. The gene expression levels relative to the housekeeping gene (i.e., Rps15a) were calculated using the 2−ΔCt formula.

Table 1.
The primer list used in this study.
2.7. Statistical Analysis
All assays were performed with three independent experiments (biological replicates) and each measurement was carried out in triplicate (technical replicates). The differences between groups were analyzed by one-way ANOVA, and two factor analysis was conducted by two-way ANOVA using GraphPad Prism Version 9 (San Diego, CA, USA). In both cases, the analyses were followed by post-hoc Bonferroni. All results were presented as a mean ± standard deviation and p < 0.05 was used to indicate a significant difference.
3. Results
3.1. Confirmation of Satellite Cell Identity
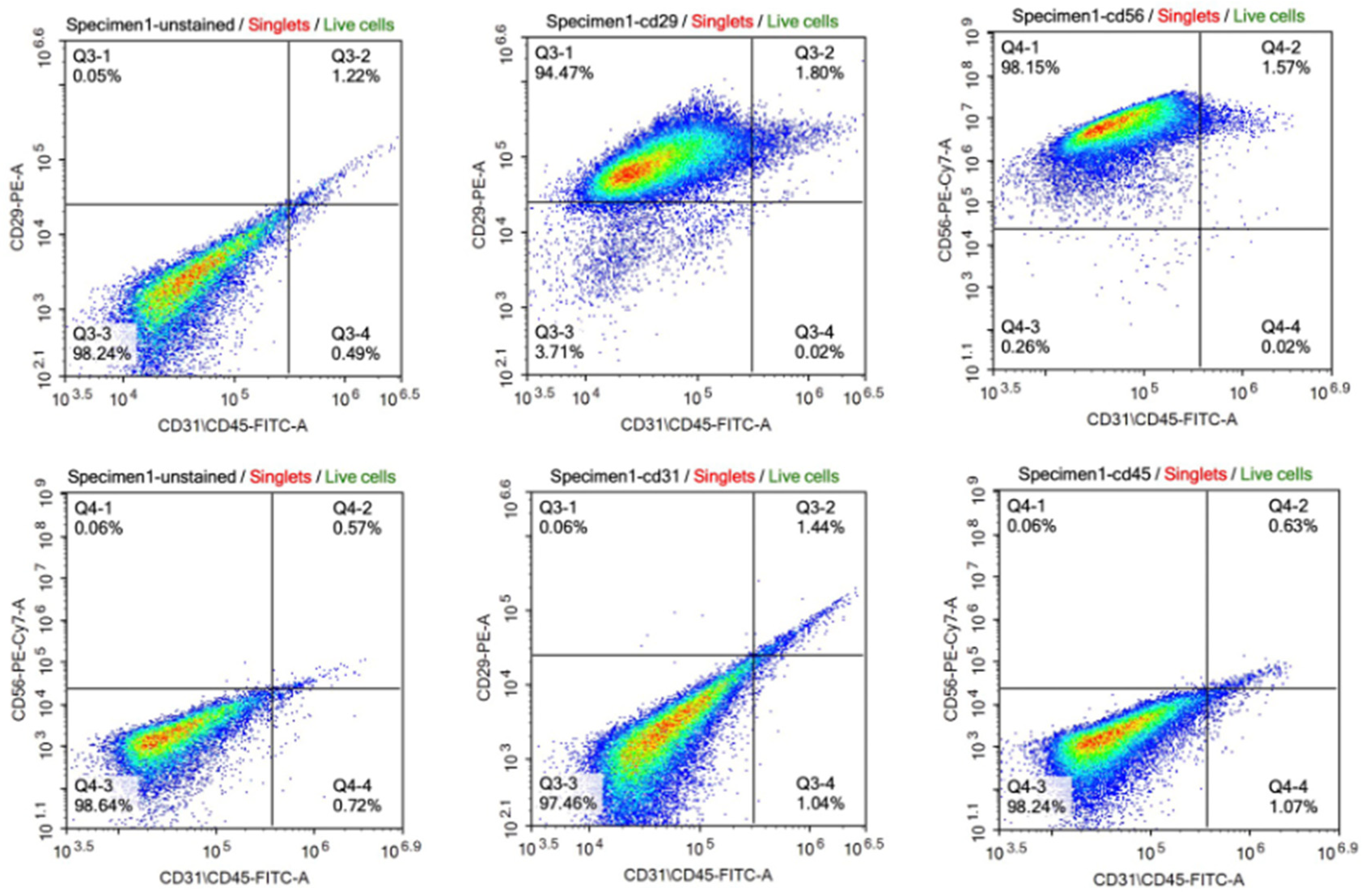
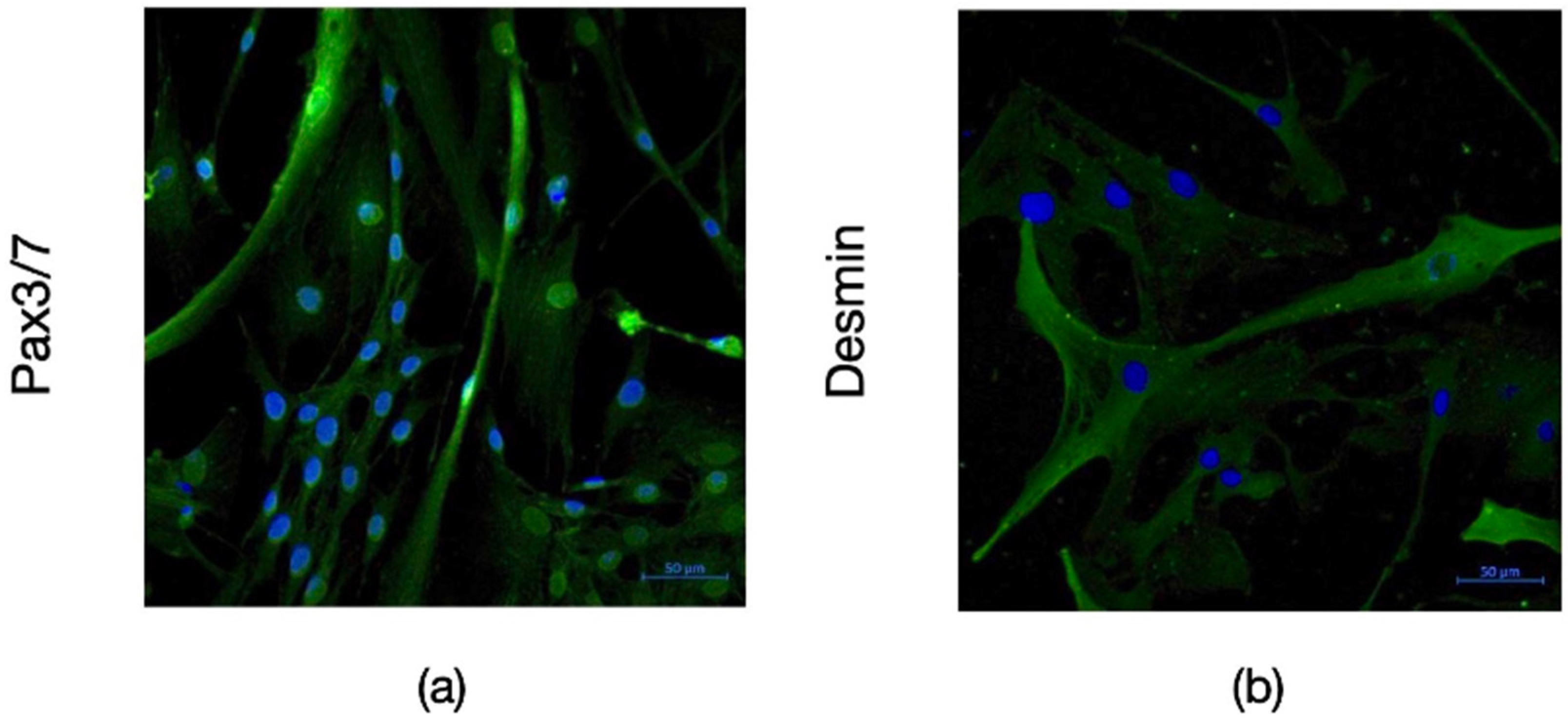
The satellite identity of the cells isolated from bovine muscle biopsy was characterized by flow cytometry and immunofluorescence staining. According to the results of flow cytometry analysis, the isolated cells expressed CD56 and CD29 which are the main surface markers of satellite and myoblast cells, respectively. However, they did not express the CD45 and CD31 markers used as negative controls (Figure 1). The data obtained from immunohistochemical staining also supported these results with the expression of Pax3/7 as satellite cell marker and Desmin as myoblast marker on isolated cells (Figure 2).
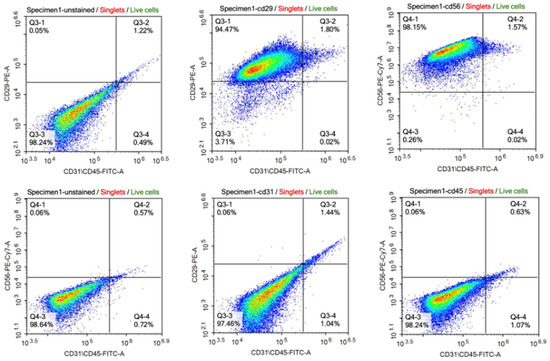
Figure 1.
Characterization of the cell population by flow cytometry. The cells express CD29 (94.47% + 1.80) and CD56 (98.15% + 1.57) as surface markers of satellite cells and myoblasts, respectively (upper panel). The cells express very low CD31 (1.44% + 0.06) and CD45 (0.63% + 0.06) as surface markers of epithelial cell and hematopoietic lineage, respectively (lower panel). [CD29: Integrin beta 1; CD56: Neural cell adhesion molecule (NCAM); CD31: Platelet endothelial cell adhesion molecule (PECAM-1); CD45: Protein tyrosine phosphatase receptor type (PTPRC); FITC-A: Fluorescein isothiocyanate; PE: Phycoerythrin; PE-Cy7: PE-Cyanine 7].
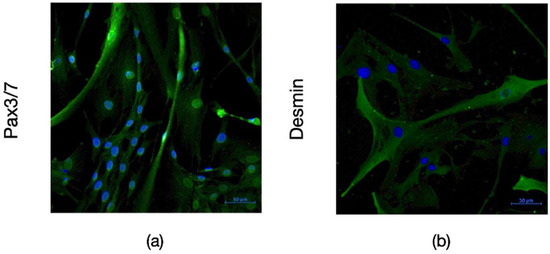
Figure 2.
Immunofluoresence imaging of bSCs with Pax3/7 ((a), green) and Desmin ((b), green) antibodies acquired with ZEISS Primovert Microscope with 40X magnification. Nuclei were stained by DAPI (blue). [bSCs: Bovine satellite cells; Pax3/7: Paired box gene 3/7; DAPI: 4′,6-diamidino-2-phenylindole].
3.2. The Effects of GFs on bSC Proliferation
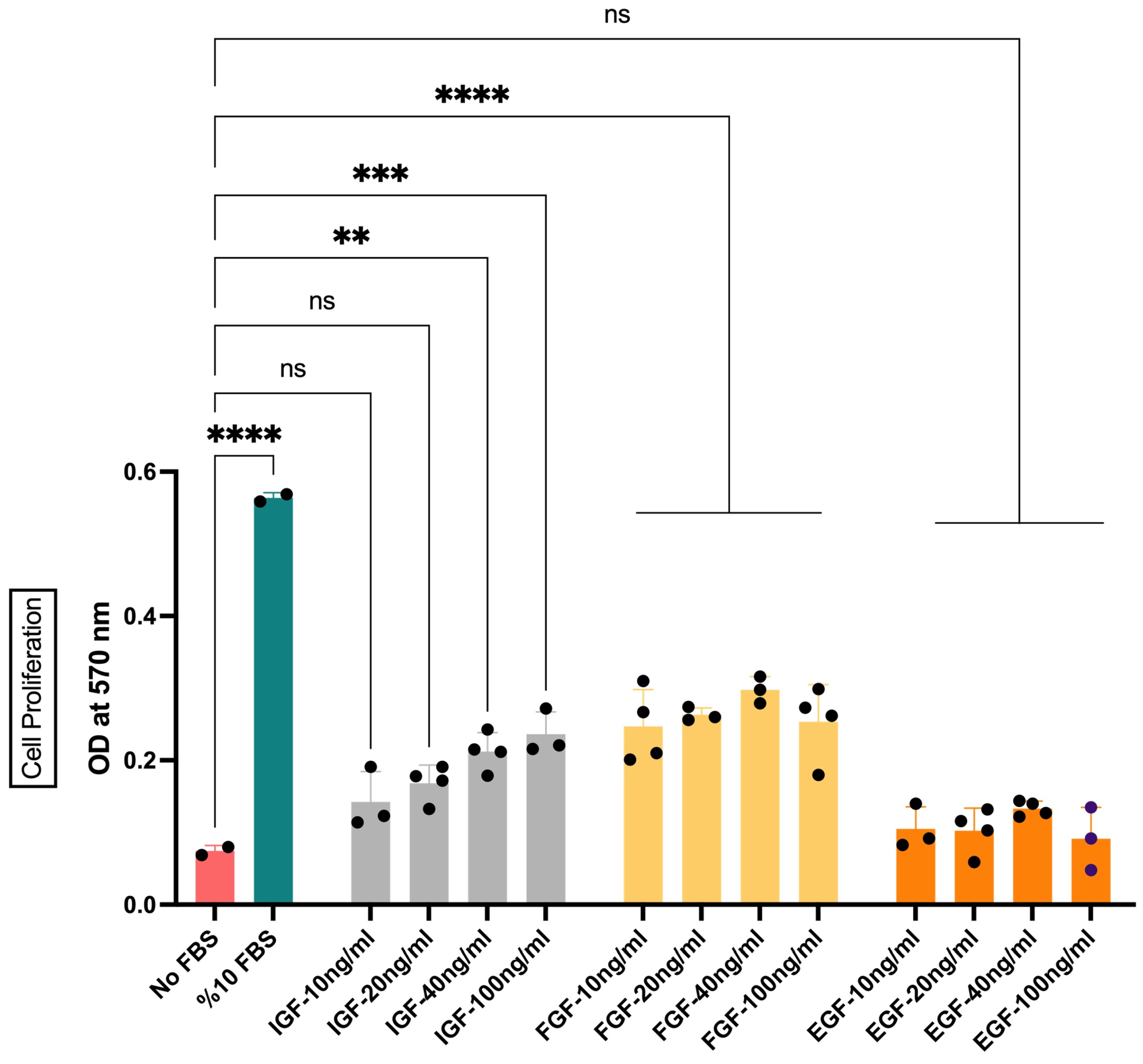
To eliminate the need of FBS for cell proliferation and survival, GFs are accepted as an alternative approach. Many of the well-characterized GFs have been applied in this context. Amongst these, the effects of IGF1, FGF2, and EGF on the viability of bSCs were evaluated. Different concentrations of the GFs were initially applied to determine the minimum effective dose for each GF (Figure 3). Our data showed an ascending pattern of OD values with increasing IGF1 concentrations. However, only 40 ng/mL and 100 ng/mL concentrations resulted in statistically significant differences from the No FBS group. Therefore, 40 ng/mL was chosen as the minimum effective dose for IGF1 (Figure 3). A more stable pattern for FGF2 was observed. Lower concentrations (i.e., 10 ng/mL and 20 ng/mL) of FGF2 were able to induce significant differences in cell proliferation, while higher concentrations did not result in further increase. The concentration of 20 ng/mL was chosen for further analysis due to its less variation in response (Figure 3). The treatment with different concentrations of EGF, on the other hand, did not yield any significant results (Figure 3). These results suggested that EGF alone was insufficient for initiating proliferative response of bSCs and was not used in further groups.
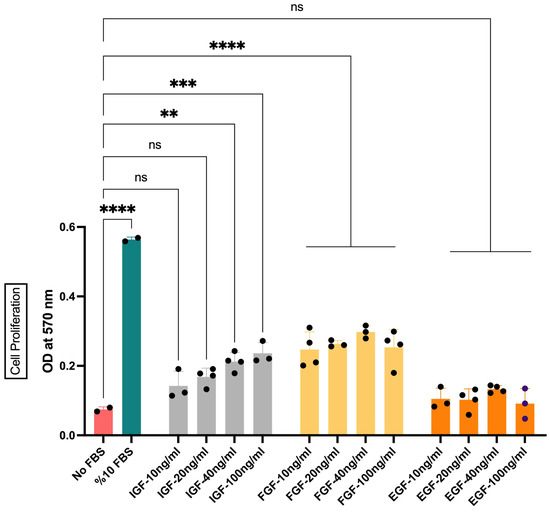
Figure 3.
The effect of IGF1, FGF2, and EGF on the proliferation of bSCs assessed with MTT assay. Mean OD values at 570 nm are given for each group. **: p < 0.01, ***: p < 0.001, ****: p < 0.0001, ns: not significant. The error bars represent ± SEM. [IGF: insulin-like growth factor 1; FGF: fibroblast growth factor 2; EGF: epidermal growth factor, OD: optical density].
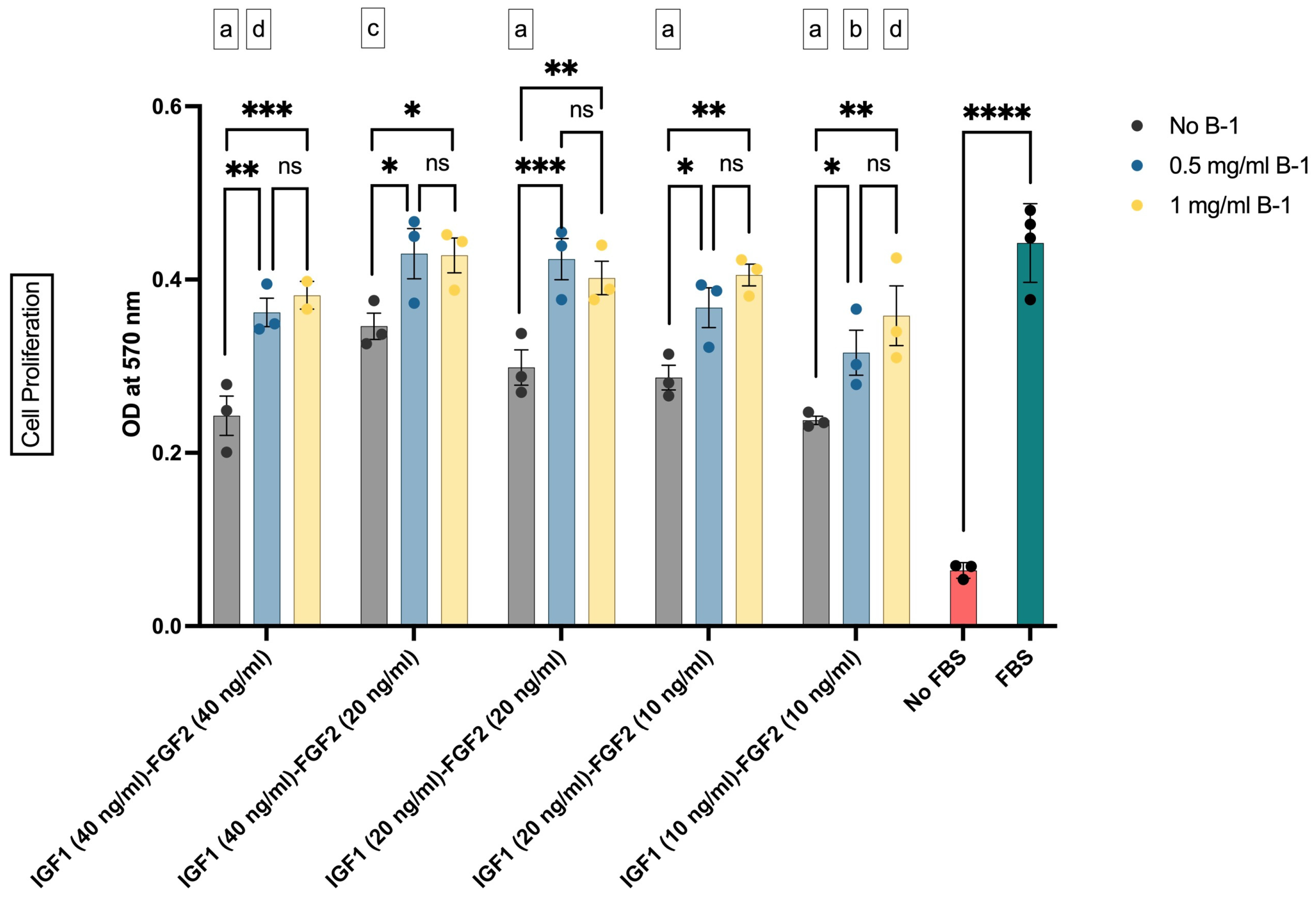
3.3. The Effects of Supplement B-1 and GF Combinations on bSC Proliferation
In the second step of our study, it was investigated whether or not our in-house-produced postbiotics as supplement B-1 enhanced the proliferative effects of GF combinations. The effective GF concentrations and their combinations were combined with two different concentrations of B-1, and their impact on the cell proliferation was tested. In this setup, 0.5 mg/mL or 1 mg/mL of B-1 was added to 40 ng/mL, 20 ng/mL, and 10 ng/mL concentrations of GF combinations (Figure 4). Our results showed that addition of B-1 into the culture medium augmented the proliferative effect of GFs (Figure 4). Both concentrations of B-1 resulted in similar OD values, with only numerical increases observed in 1 mg/mL groups. These results indicated that B-1 increased the cell proliferation, regardless of the GF concentrations.
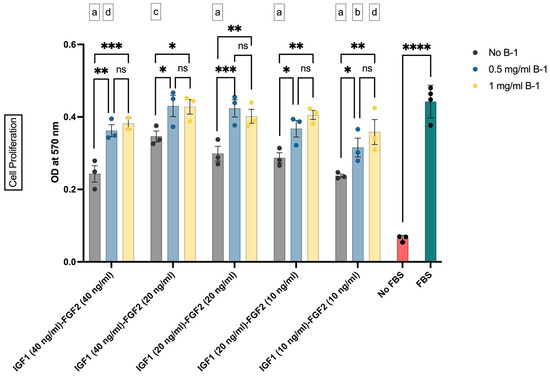
Figure 4.
The effect of IGF1, FGF2, and supplement B-1 on the proliferation of bSCs assessed with MTT assay. Mean OD values at 570 nm are given for each group. *: p < 0.05; **: p < 0.01; ***: p < 0.001; ****: p < 0.0001; ns: not significant. The comparisons of the group mean with that of FBS group are given as letter codes (a–d). a: p < 0.0001; b: p < 0.001; c: p < 0.01; d: p < 0.05. The error bars represent ± SEM. [IGF1: insulin-like growth factor 1; FGF2: fibroblast growth factor 2; B-1: postbiotic supplement Biftek-1; OD: optical density].
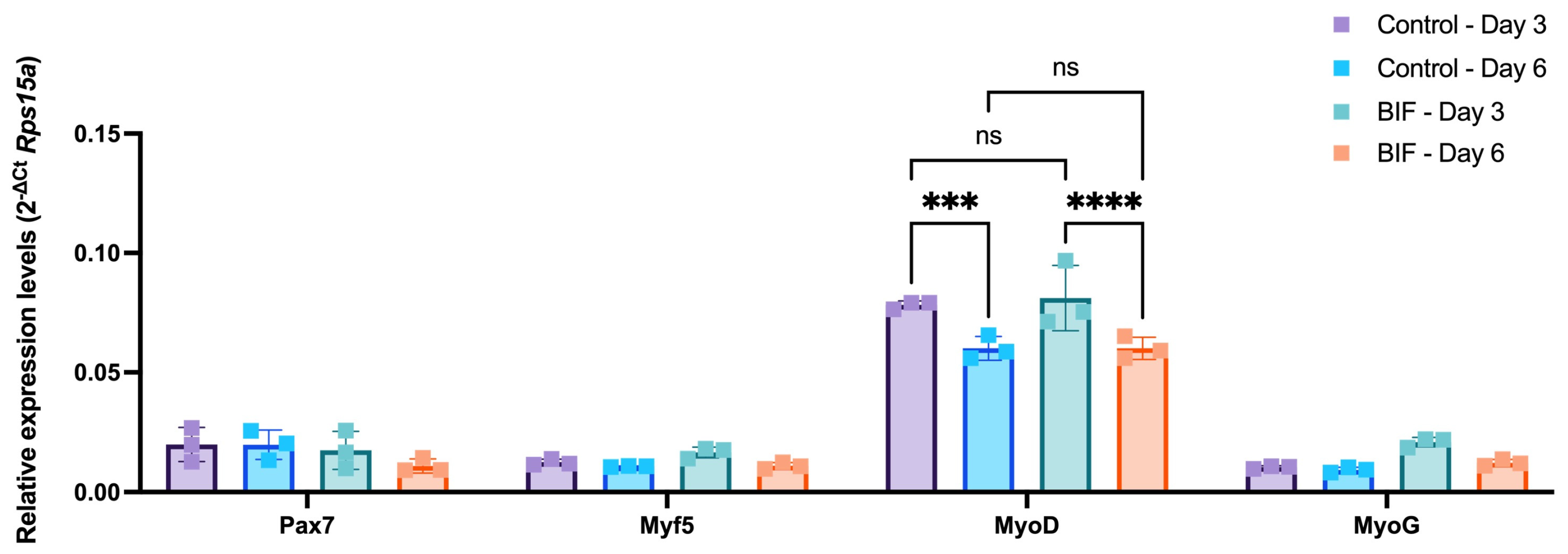
3.4. Myogenic mRNA Expression Profiles of bSCs treated with Supplement B-1 and GF Combinations
Another critical point to consider while using serum-free growth media in the cultivated meat applications is retaining the stemness of the cells. The starting material, the muscle biopsy, is often limited. Hence, methods that maximize the cell yield by preserving the initial muscle stem cell pool for longer periods are required. Therefore, to investigate whether the selected B-1 and GF combinations (i.e., 0.5 mg/mL B-1, 40 ng/mL IGF1 and 20 ng/mL FGF2; referred to as BIF hereafter) caused any changes in the myogenic profile of bSCs, the expression levels of genes that have been established as markers of different myogenic progression levels were quantified (Figure 5). For this purpose, Pax7, Myf5, MyoD, and MyoG were selected as key markers for the evaluation of the different stages of differentiation. The bSCs were treated with either BIF or 10% FBS for 3 and 6 days to capture the time-dependent changes. Our data showed that BIF treatment did not lead to any significant changes in the expression levels of the genes of interest. However, the duration of the treatment affected the MyoD gene expression, since a downregulation at day 6 in both groups was observed. Overall, our results indicated that BIF did not drive bSCs to differentiation, which could serve as a growth medium supplement alternative to FBS in cultivated meat applications.
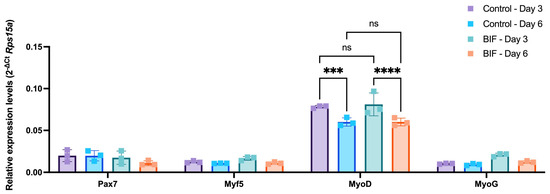
Figure 5.
Time-course changes in the expression levels of myogenic genes in response to BIF or 10% FBS-containing growth medium (Control) following 3 and 6 days of incubation. Mean delta Ct values as normalized to rps15a are given for each group. Neither the treatment type nor the duration altered the expression levels of Pax7, Myf5, and MyoG. MyoD expression levels were not affected by the treatment type; however, the treatment duration had a significant effect on MyoD. In both treatment groups, MyoD was downregulated from day 3 to day 6. ***: p < 0.001, ****: p < 0.0001, ns: not significant. The error bars represent ± SEM. [BIF: B-1 (0.5 mg/mL) + IGF1 (40 ng/mL) + FGF2 (20 ng/mL); Pax7: paired box gene 7; Myf5: myogenic factor 5; MyoD: myoblast determination protein; MyoG: Myogenin].
4. Discussion
The global population is expected to surpass 10 billion by 2050 [25]. The Food and Agriculture Organization (FAO) has forecast that meat production will not be sufficient to fulfill the demand of this growing population, due to the resources and arable land limitations [26]. To satisfy the increasing demand for food by the growing human population, cultured meat is presented as a good and environmentally friendly alternative for consumers. For generating 1000 kg of cultured meat, 1014 cells are needed, with a final cell density of around 4 × 107 cells/mL in the bioreactor with a batch volume of 20,000 L. The main limitation for production of cultured meat at this scale is the cost of media and supplements. It has been reported that FBS and GFs are the main cost drivers. Moreover, expensive proteins such as albumin, transferrin, and insulin, which are used at significantly higher concentrations, are also the bigger cost contributors [27]. Therefore, replacement of them by functional alternatives seems the only solution for this technology.
To lower the costs and environmental impacts of the production of GFs and other recombinant proteins, animal-free protein hydrolysates obtained from animals, plants, and insects with their promoting effects on cell growth have gained attention as a low-cost alternative [28]. Moreover, bioactive compounds derived from cyanobacteria and algae are considered as cheap alternatives to GFs and FBS in cultured meat technology [29,30,31]. In our study, postbiotics derived from a bacterium of cow milk microbiota have been used to formulate low-cost and animal-free cell culture supplements. For this purpose, bSCs were isolated from bovine muscle and their profiles observed on flow cytometry and immunofluorescent staining were found consistent with satellite cells and myoblasts similar to Ding et al. [22].
The initial strategy of this study was to define the minimum effective concentrations of GFs for the proliferation of bSCs. Our results suggested that very low concentrations of GFs might be able to induce changes in the proliferation rate of bSCs. The lack of a significant difference between the highest and lowest concentrations applied (10 ng/mL and 100 ng/mL, respectively), particularly for FGF2 and EGF, might be related to receptor desensitization, which is a well-established phenomenon [32,33,34]. Furthermore, none of the GFs were able to replicate the OD values obtained from the FBS group. This observation is consistent with the literature, since most serum-free growth medium alternatives include more than one GF as well as other metabolites in the recipe for successful outcomes in the cultivated meat applications [10]. Another interesting observation obtained was the ineffectiveness of EGF on the proliferation of bSCs, whereas IGF1 and FGF2 resulted in dose-dependent changes. Although all three GFs have been implicated to promote muscle cell proliferation [35,36], they might have different roles in satellite cell biology with respect to the receptors they activate and the signaling cascades activated by them. Indeed, muscle cells cultured in growth medium containing only EGF have been reported to have lower proliferation rates than those cultured in media containing FGF2 or EGF + FGF2 [27].
The second strategy of this study was to investigate the synergistic effects of supplement B-1 with GFs on the proliferation of bSCs. Our results showed that bSCs grown in serum-free conditions with low concentrations of GFs have shown enhanced proliferation when B-1 was used as a supplement. Similar to our approach, algae extract obtained from Chlorella vulgaris [31] and cyanobacteria extracts derived from Spirulina [30] have been used as a partial FBS and GFs replacement in mesenchymal stem cells and cell cultures, respectively. In another study conducted by Ghosh et al. (2023), bioactive compound was extracted from the filamentous nitrogen-fixing cyanobacteria Anabaena sp. PCC 7120 by the ultrasonication method and used as a growth supplement to cultivate muscle cells. Their results indicated that water-soluble compounds had a positive role in muscle cell proliferation [15]. Although several compounds have been clarified in these extracts such as phenolic compounds and proteins, it is still unknown which compounds or molecules are responsible for stimulation of the cell growth [37]. Similar to this observation, the observed effect of B-1 supplement can be related to the diverse interactions of different compounds. According to the available current evidence, the interaction between postbiotics and host cells can be through potential cell receptors. Proteins, namely p40 and p75, were clarified from Lactobacillus rhamnosus GG postbiotics which induced the proliferation, migration, and survival when tested on both murine and human cell lines via cell receptors [38,39]. However, since postbiotics can contain hundreds or even thousands of metabolites including SCFAs, proteins, phenolic compounds, and their conjugates, it is not possible to give a completely clear formulation.
Since satellite cells can lose their stemness if they are not cultured in optimum conditions, monitoring the expression levels of myogenic markers in response to serum-free growth medium formulations is an integral part of the process. For this purpose, the expression levels of myogenic genes in bSCs incubated in control (10% FBS) and BIF-containing medium were determined. Our results indicated that the medium compounds did not affect the expression levels of Pax7, Myf5, MyoG, and MyoD as well-established markers of myogenic progression [23]. Notably, a decrease in the expression levels of MyoD in both groups from day 3 to 6 was observed. Studies conducted on Jeju Black pig muscle cells showed that the presence of bFGF affected the rate of proliferation and, together with EGF, also the level of expression of the MyoD gene [40]. Due to being a key regulator of myogenesis, downregulation of MyoD may indicate depletion of the stem cell pool. Nevertheless, since this downregulation was observed in both groups and to the same extent, the underlying mechanism is possibly related to time spent in culture rather than the medium content.
5. Conclusions
Nowadays, postbiotics with their unique metabolic profiles and functions are considered as key mediators for various cellular processes and metabolic pathways [41]. Although the use of postbiotics is common in various scientific fields as well as in pharmaceutical products, they were transferred to a new technology/field within the scope of this study, for the first time. According to our results, B-1 derived from Enterococcus hirae, an isolate of cow milk microbiota, has a potential role in muscle cell proliferation for cultivated meat technology. Further studies will be conducted to transfer the in vitro test results into proper cell production in the bioreactor.
6. Patents
Akcali, K.C., Erikci, E. and Kiran, F., Biftek Inc., 2022. Microbiota-derived postbiotics: Alternative supplement to fetal bovine serum for cultured meat. U.S. Patent Application 17/486,944.
Author Contributions
Conceptualization, K.C.A. and F.K.; methodology, D.C.-B., K.C.A. and F.K.; validation, D.C.-B. and E.E.; formal analysis, D.C.-B. and K.G.; investigation, D.C.-B., K.G. and I.A.; writing—original draft preparation, D.C.-B. and F.K.; writing—review and editing, E.E. and K.C.A.; visualization, D.C.-B. and K.G.; supervision, K.C.A. and F.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of ANKARA UNIVERSITY (2017-7-60, 10 July 2017) and SELCUK UNIVERSITY (2021-92, 1 July 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
Authors Dilan Celebi-Birand, Kardelen Genc, Ilknur Agun were employed by the company Biftek Inc. Authors Erdem Erikci, Kamil Can Akcali and Fadime Kiran own stocks in Company Akerbio Biotechnology and also were the inventors of patent application.
References
- Kumar, P.; Sharma, N.; Sharma, S.; Mehta, N.; Verma, A.K.; Chemmalar, S.; Sazili, A.Q. In-vitro meat: A promising solution for susustainability of meat sector. J. Anim. Sci. Technol. 2021, 63, 693–724. [Google Scholar] [CrossRef] [PubMed]
- Swartz, E. Cell Culture Media and Growth Factor Trends in the Cultivated Meat Industry; The Good Food Institute (GFI): Washington, DC, USA, 2021. [Google Scholar]
- Byrne, B. State of the Industry Report|Cultivated Meat, Cell Agriculture Greece; The Good Food Institute (GFI): Washington, DC, USA, 2021. [Google Scholar]
- Garrison, G.L.; Biermacher, J.T.; Brorsen, B.W. How much will large-scale production of cell-cultured meat cost. J. Agric. Food Res. 2022, 10, 100358. [Google Scholar] [CrossRef]
- Beef-Prices by Country around the World, September 2023. Available online: https://www.globalproductprices.com/rankings/beef_price/ (accessed on 2 October 2023).
- Ong, K.J.; Johnston, J.; Datar, I.; Sewalt, V.; Holmes, D.; Shatkin, J.A. Food safety considerations and research priorities for the cultured meat and seafood industry. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5421–5448. [Google Scholar] [CrossRef] [PubMed]
- Chelladurai, S.K.; Christyraj, S.J.D.; Rajagopalan, K.; Yesudhason, B.V.; Venkatachalam, S.; Mohan, M.; Vasantha, N.C.; Christyraj, J.R.S. Alternative to FBS in animal cell culture—An overview and future perspective. Heliyon 2021, 7, e07686. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Haraguchi, Y.; Takahashi, H.; Kawashima, I.; Shimizu, T. Development of serum-free and grain-derived-nutrient-free medium using microalga-derived nutrients and mammalian cell-secreted growth factors for sustainable cultured meat production. Sci. Rep. 2023, 13, 498. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, S.Y.; Yun, S.H.; Jeong, J.W.; Kim, J.H.; Kim, H.W.; Choi, J.S.; Kim, G.D.; Joo, S.T.; Choi, I.; et al. Review of the current research on Fetal Bovine Serum and the development of cultured meat. Food Sci. Anim. Resour. 2022, 42, 775–799. [Google Scholar] [CrossRef] [PubMed]
- Stout, A.J.; Mirliani, A.B.; Rittenberg, M.L.; Shub, M.; White, E.C.; Yuen, J.S.K.; Kaplan, D.L. Simple and effective serum-free medium for sustained expansion of bovine satellite cells for cell cultured meat. Commun. Biol. 2022, 5, 466–479. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of skeletal muscle stem cells by fibroblast growth factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef]
- Reducing Growth Factor Costs for Cultured Meat Production, October 2021. Available online: https://www.cellgs.com/blog/reducing-growth-factor-costs-for-cultured-meat-production.html (accessed on 13 November 2023).
- Smolskaya, S.; Logashina, Y.A.; Andreev, Y.A. Escherichia coli extract-based cell-free expression system as an alternative for difficult-to-obtain protein biosynthesis. Int. J. Mol. Sci. 2020, 21, 928. [Google Scholar] [CrossRef]
- Swartz, E.; Ravi, A.; Reeber, A.; Levink, J.; Huang, T.; Smith, B. Anticipated Growth Factor Costs and Volumes; The Good Food Institute (GFI): Washington, DC, USA, 2021. [Google Scholar]
- Ghosh, J.; Haraguchi, Y.; Asahi, T.; Nakao, Y.; Shimizu, T. Muscle cell proliferation using water-soluble extract from nitrogen-fixing cyanobacteria Anabaena sp. PCC 7120 for sustainable cultured meat production. Biochem. Biophys. Res. Commun. 2023, 682, 316–324. [Google Scholar] [CrossRef]
- Zolkiewicz, J.; Marzec, A.; Ruszczynski, M.; Feleszko, W. Postbiotics—A step beyond pre- and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Giron, M.; Thomas, M.; Dardevet, D.; Chassard, C.; Savary-Auzeloux, I. Gut microbes and muscle function: Can probiotics make our muscles stronger? J. Cachexia Sarcopenia Muscle 2022, 13, 1460–1476. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Xing, D. The current and future perspectives of postbiotics. Probiotics Antimicrob. Proteins 2023, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sevin, S.; Karaca, B.; Haliscelik, O.; Kibar, H.; OmerOglou, E.; Kiran, F. Postbiotics secreted by Lactobacillus sakei EIR/CM-1 isolated from cow milk microbiota, display antibacterial and antibiofilm activity against ruminant mastitis-causing pathogens. Ital. J. Anim. Sci. 2021, 20, 1302–1316. [Google Scholar] [CrossRef]
- Akcali, K.C.; Erikci, E.; Kiran, F. Microbiota-Derived Postbiotics: Alternative Supplement to Fetal Bovine Serum for Cultured Meat. U.S. Patent 17/486,944, 31 February 2022. [Google Scholar]
- Skrivergaard, S.; Rasmussen, M.K.; Therkildsen, M.; Young, J.F. Bovine satellite cells isolated after 2 and 5 days of tissue storage maintain the proliferative and myogenic capacity needed for cultured meat production. Int. J. Mol. Sci. 2021, 22, 8376. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Swennen, G.N.M.; Messmer, T.; Gagliardi, M.; Molin, G.M.D.; Li, C.; Zhou, G.; Post, M.J. Maintaining bovine satellite cells stemness through p38 pathway. Sci. Rep. 2018, 8, 10808. [Google Scholar] [CrossRef] [PubMed]
- Coles, C.A.; Wadeson, J.; Leyton, C.P.; Siddell, J.P.; Greenwood, P.L.; White, J.D.; McDonagh, M.B. Proliferation rates of bovine primary muscle cells relate to liveweight and carcase weight in cattle. PLoS ONE 2015, 10, e0124468. [Google Scholar] [CrossRef]
- Wang, G.H.; Liang, C.C.; Li, B.Z.; Du, X.Z.; Zhang, W.Z.; Cheng, G.; Zan, L.S. Screening and validation of reference genes for qRT-PCR of bovine skeletal muscle-derived satellite cells. Sci. Rep. 2022, 12, 5653. [Google Scholar] [CrossRef]
- UN. UN [United Nations] World Population Prospects 2019; UN: New York, NY, USA, 2019. [Google Scholar]
- Food and Agriculture Organization: How to Feed the World in 2050. Available online: https://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 3 October 2023).
- Specht, L. An Analysis of Culture Medium Costs and Production Volumes for Cultivated Meat; The Good Food Institute (GFI): Washington, DC, USA, 2020. [Google Scholar]
- Obaidi, I.; Mota, L.M.; Quigley, A.; Butler, M. The role of protein hydrolysates in prolonging viability and enhancing antibody production of CHO cells. Appl. Microbiol. Biotechnol. 2021, 105, 3115–3129. [Google Scholar] [CrossRef]
- Jeong, Y.; Choi, W.Y.; Park, A.; Lee, Y.J.; Lee, Y.; Park, G.H.; Lee, S.-J.; Lee, W.-K.; Ryu, Y.-K.; Kang, D.-H. Marine cyanobacterium Spirulina maxima as an alternate to the animal cell culture medium supplement. Sci. Rep. 2021, 11, 4906. [Google Scholar] [CrossRef]
- Tuomisto, H.L.; Teixeira de Mattos, M.J. Environmental impacts of cultured meat production. Environ. Sci. Technol. 2011, 45, 6117–6123. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.Y.; Chua, M.L.; Zhang, C.; Hong, S.; Kumar, Y.; Gokhale, R.; Ee, P.L.R. Chlorella vulgaris extract as a serum replacement that enhances mammalian cell growth and protein expression. Front. Bioeng. Biotechnol. 2020, 8, 564667. [Google Scholar] [CrossRef] [PubMed]
- Countaway, J.L.; Nairn, A.C.; Davis, R.J. Mechanism of desensitization of the epidermal growth factor receptor protein-tyrosine kinase. J. Biol. Chem. 1992, 267, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Higa-Nakamine, S.; Noguchi, N.; Maeda, N.; Kondo, Y.; Toku, S.; Kukita, I.; Sugahara, K. Desensitization by different strategies of epidermal growth factor receptor and ErbB4. J. Pharmacol. Sci. 2014, 124, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.J.; Mittal, B.; Walsh, F.S.; Doherty, P. FGF inhibits neurite outgrowth over monolayers of astrocytes and fibroblasts expressing transfected cell adhesion molecules. J. Cell Sci. 1995, 108, 3523–3530. [Google Scholar] [CrossRef] [PubMed]
- Charge, S.B.; Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, O.M.; Vega-Soto, E.E.; Nguyen, M.H.; Cederna, P.S.; Larkin, L.M. Impact of human epidermal growth factor on tissue-engineered skeletal muscle structure and function. Tissue Eng. Part A 2021, 27, 1151–1159. [Google Scholar] [CrossRef]
- Hubalek, S.; Post, M.J.; Moutsatsou, P. Towards resource-efficient and cost-efficient cultured meat. Curr. Opin. Food Sci. 2022, 47, 100885. [Google Scholar] [CrossRef]
- Yan, F.; Liu, L.; Dempsey, P.J.; Tsai, Y.H.; Raines, E.W.; Wilson, C.L.; Cao, H.; Cao, Z.; Liu, L.; Polk, D.B. A Lactobacillus rhamnosus GG-derived soluble protein, p40, stimulates ligand release from intestinal epithelial cells to transactivate epidermal growth factor receptor. J. Biol. Chem. 2013, 288, 30742–30751. [Google Scholar] [CrossRef]
- Kang, S.J.; Jun, J.S.; Hong, K.W. Transcriptome analysis reveals immunomodulatory effect of spore-displayed p75 on human intestinal epithelial Caco-2 Cells. Int. J. Mol. Sci. 2022, 23, 14519. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Song, K.D.; Kim, S.J.; Kim, D.C.; Lee, S.C.; Son, Y.J.; Choi, H.W.; Shim, K. Growth factors improve the proliferation of Jeju black pig muscle cells by regulating myogenic differentiation 1 and growth-related genes. Anim. Biosci. 2021, 34, 1392–1402. [Google Scholar] [CrossRef]
- Ozma, M.A.; Abbasi, A.; Akrami, S.; Lahouty, M.; Shahbazi, N.; Ganbarov, K.; Pagliano, P.; Sabahi, S.; Köse, Ş.; Yousefi, M.; et al. Postbiotics as the key mediators of the gut microbiota-host interactions. Infez. Med. 2022, 30, 180–193. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).