1. Introduction
Fishing activities and, in particular, bottom trawling directly and indirectly affect fish assemblages and cause a number of undesirable consequences on fish populations such as reduced biomass, truncated size structures, and disrupted food webs, with a large part of world stocks falling in the range between fully exploited to depleted [
1,
2,
3,
4,
5,
6]. Fisheries management aiming at sustainable yields through approaches based on catch quota and control of fishing effort have proved generally effective when applied to monospecific fisheries [
7,
8] and even then, stock collapses have occurred with dramatic consequences [
9]. Multispecies fisheries, which dominate in warm-temperate and tropical regions, are more complex to manage due to the high number of species involved, the variety of their ecological and biological traits, and the complex interactions with habitats and environmental parameters. An ecosystem-based approach addressing the fish assemblage as a whole as well as the habitat has been often invoked, especially in areas characterized by conflicting human activities—including but not limited to fishing—that would benefit by a spatial approach [
10,
11]. Models as well as empirical observation suggest that multispecies fisheries may benefit from partial or total protection in terms of increased biomass [
12,
13,
14,
15], better structured size distributions with a higher abundance of larger fish [
16] (‘filling-in effect’ in Baskett & Barnett, [
17]), increased resilience to overfishing for overexploited low-mobility species and for underexploited highly mobile species [
18], increased and less variable catches in surrounding areas [
19,
20], increased yield reliability under different management scenarios [
21], better economic performance of outside fisheries at determinate management conditions [
22], and export of adult biomass [
13,
23,
24].
Marine protected areas (MPAs)
sensu lato are widespread tools for the conservation and enhancement of living marine resources. Several types of MPAs exist, ranging from no-take reserves to partially protected areas (PPAs) that allow some nondestructive fishing activities [
24,
25], each of them is widely applied with different levels of success [
26]. PPAs and multiple-use MPAs have been advocated as effective large-scope management tools, sometimes even more effective than fully protected MPAs in that they address more widely the different ecosystem services while considering also human activities and sustainable resource use [
27]. Sciberras et al. [
28] observed that while PPAs (such as no-trawl areas) may produce less strong effects than no-take areas, they still have the advantage of offering an effective management option where the institution of fully protected areas is not socio-economically viable. PPAs based on the ban of towed fishing gears have been created in tropical (southeastern Asia: [
29]), warm-temperate (Mediterranean: Pipitone et al. [
30]), and temperate (North Sea: Pastoors et al. [
31]) areas, where they have generally yielded very encouraging results in terms of fish stock enhancement. Furthermore, coastal small-scale fisheries, which represent a widespread and important socio-economic sector in most temperate and tropical areas and are generally considered a sustainable activity [
32], may vastly benefit from their inclusion in MPAs’ and PPAs’ management plans [
33,
34,
35].
PPAs have been realized in the Mediterranean in the form of spatial fishery restrictions of various sizes and with different objectives [
30]. The Gulf of Castellammare (NW Sicily) makes up a case study as a no-trawl area where a year-round trawl ban has been in place since 1990 while artisanal and recreational fishing are permitted [
36]. The main effects of trawl exclusion on demersal fish and invertebrates in the Gulf after the first few years have been investigated, including those on fish biomass [
36] and size spectra [
37], the food web [
38,
39,
40,
41], fish growth and condition factor [
42,
43,
44], stock dynamics of commercial species [
45], and benthic communities [
46,
47]. However, as regards the effects on fish biomass, published data are spatially and temporally limited, and those on fish size have been scarcely addressed to date.
The general objective of the present study was to evaluate the effects of the trawl ban on the shelf demersal assemblage in the Gulf of Castellammare (GCAST) no-trawl area. To attain this objective, we used fisheries-independent surveys and compared (1) the biomass of the total demersal assemblage and of selected species before and after 1990 (i.e., the starting year of the ban) in the GCAST and in two areas open to trawlers, namely, the Gulfs of Termini Imerese (GTERM) and Sant’Agata (GSANT); (2) the proportion of different commercial categories of fish in GCAST before and after 1990, with the purpose of inferring the potentialities of the artisanal fishery inside the no-trawl area and to inform future management measures; and (3) the size structure of selected species in GCAST and in the two trawled gulfs after 1990.
The following hypotheses based on the effects expected from the trawl ban were tested:
Hypothesis (H1). The demersal biomass in the GCAST after 1990 is higher than in the GCAST before 1990 and than in the GTERM and the GSANT after 1990, and is similar in the three gulfs before 1990, and in the GTERM and the GSANT before vs. after 1990;
Hypothesis (H2). The proportion of highly commercial demersal species in the GCAST is higher after 1990;
Hypothesis (H3). The size structures of demersal species in the GCAST after 1990 are different and with larger median size and an increased proportion of larger individuals than in the GTERM and the GSANT.
4. Discussion
Spatial measures such as the trawl ban in the GCAST represent an application of a multispecies spatial approach that matches the principles of ecosystem-based fisheries management. Spatially explicit management has the potential to preserve ecosystem resilience and allow sustainable fishing and protection of habitats, community structure, and ecosystem functions [
64,
65]. Although fisheries-induced impacts may have evolutionary components that are hardly reversible, leading in some cases to the lack of recovery after protection [
66], most of the partial or absent effectiveness observed in some cases has been explained mainly with unsuccessful habitat protection, lack of compliance, mismanagement issues, inappropriate planning, unexpected fish stock redistribution, modifications of fishing pressure, and impact from recreational fishing or fisheries-unrelated human activities [
67,
68,
69,
70,
71,
72,
73].
Overall, the use of fishery reserves (
sensu Auster & Shackell [
74]) and, more generally, of partially protected areas as management tools has been enthusiastically fostered and the evidence of their potential in an ecosystem-based approach has been steadily growing [
75,
76,
77,
78], although a high variability in their effectiveness—see examples above—should be acknowledged. The Mediterranean Sea, an area historically exploited by coastal and offshore fisheries [
79], provides several examples of the successful use of trawl bans and other area-based management tools [
30], some of which date back to more than one century ago such as the trawl ban imposed in 1896 in northern Sicily, leading to an increase of catches and an enhancement of the economic condition of a vast number of artisanal fishermen [
75].
Our study has provided evidence that a year-round trawl ban imposed on the soft bottoms of the continental shelf in a warm-temperate region such as the Mediterranean may lead to a remarkable enhancement of the demersal fish assemblage. The effect is clearer for biomass but less defined for the size structure of the affected populations.
4.1. Biomass
The total biomass of the demersal fish assemblage in the GCAST underwent an increase after the 1990 trawl ban, confirming our first hypothesis. An increase in finfish and shellfish biomass is the most commonly observed effect of fishing exclusions in coastal and offshore ecosystems [
12]. The increase of the total biomass observed in the GCAST should be considered a clear sign of ecosystem recovery after a multidecadal intense fishing impact on the area [
80].
The similarity between the before-1990 biomass value in the GCAST and the before- and after-1990 values in the two trawled gulfs shows that in the absence of the trawl ban, the three areas shared a similar amount of trawlable resources. The postban biomass increase in the GCAST had already been observed as early as 1994 [
36], although supported by a much smaller dataset and a weaker statistical design than in the present work. Similar evidence of biomass increase was recorded in a few other trawl exclusion areas in the Mediterranean [
52], the Atlantic Ocean [
24,
81,
82,
83,
84], and the Pacific Ocean [
73], even though only a few studies [
69,
85] have considered the effect of protection on the whole trawlable assemblage rather than on a few target species.
Considering the ten target species, they reflected, to a large extent, the same general trend of increased biomass observed for the total trawlable assemblage in the GCAST after the ban. Red mullet in particular has undergone a dramatic increase [
36,
45] and has possibly contributed to the recovery of two of its main predators, white grouper
Epinephelus aeneus and anglerfish [
86]. The biomass of red mullet, pandora, annular seabream, and axillary seabream increased also in a no-trawl area in the NW Aegean Sea [
52]. Likewise, positive effects of a trawl ban on the recruitment of hake, along with some evidence of spillover to adjacent fishing grounds, were recorded from another small no-trawl area located off the Catalan coast [
87].
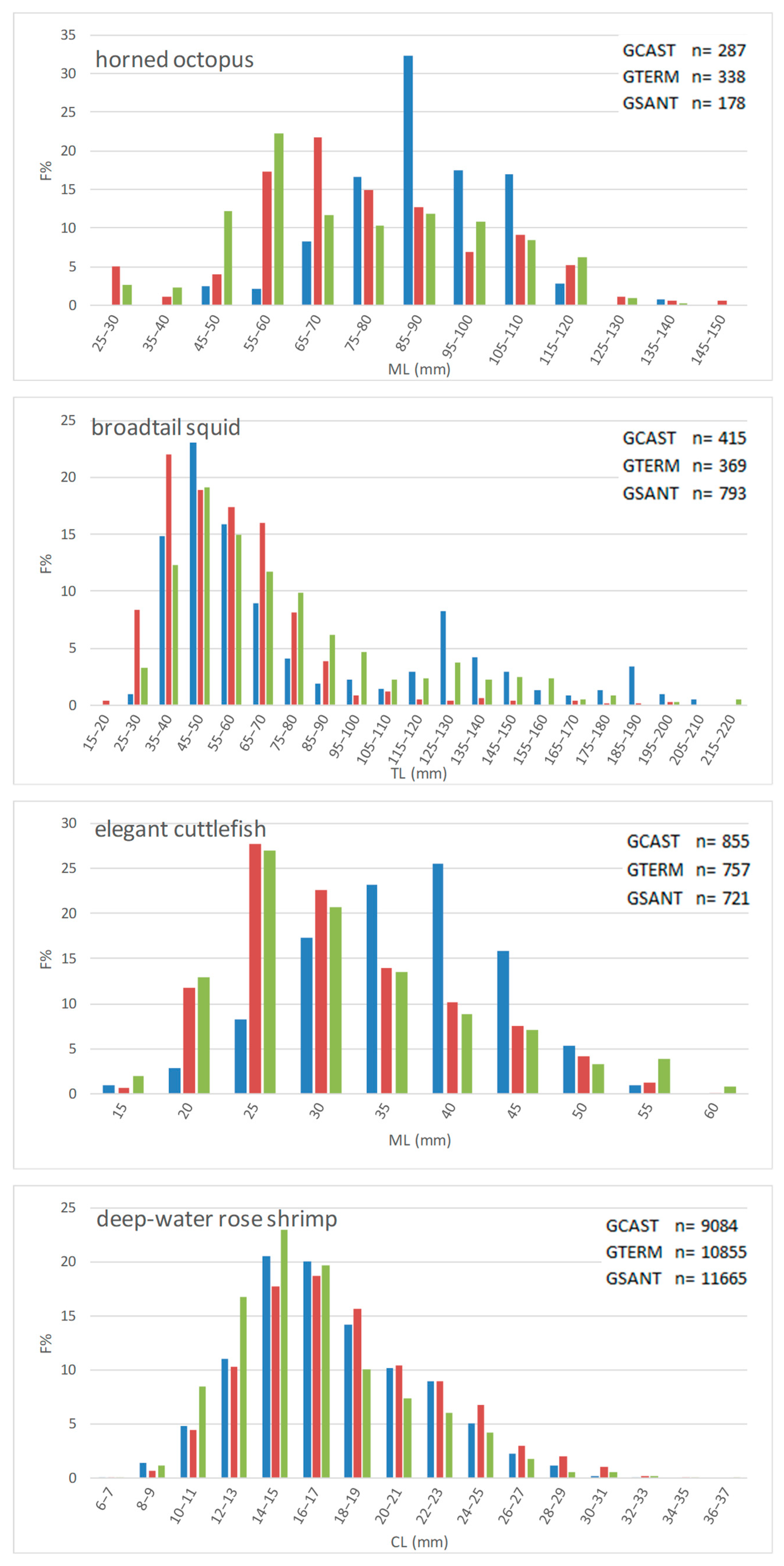
The general postban biomass increase observed in the GCAST did not involve all target species though. The biomass of horned octopus and deep-water rose shrimp was similar in the after-1990 GCAST and in some of the other Time/Locality combinations; while that of common octopus was higher after 1990 but only in the GCAST. Cephalopods are short-lived, fast growing, rapid-turnover species, and as such, they are expected to suffer from intense fishing less than slower growing, longer living species as most demersal finfish are [
88,
89]. This might explain the observed biomass values of the first two species but not the increased biomass of common octopus and common cuttlefish in the after-1990 GCAST. Environmental factors have been suggested to impact cephalopods even more than anthropogenic factors such as fishing [
89,
90], so unknown, complex interactions between co-occurring natural and human impacts, as well as possible predation release due to the after-1990 increase of fish biomass in the GCAST might have driven the observed trends.
Deep-water rose shrimp is another fast growing, short-lived species and a prey for a number of fishes [
48,
91]. Its short life span associated with low biomass of predators could justify the high biomass values recorded in the trawled areas, while the increase of potential predators such as anglerfish and hake and the intense fishing pressure exerted on the shelf break outside the trawl ban area, where most of the population lives (pers. observ.) could have hampered a larger increase in the GCAST, where, nonetheless, protection triggered a biomass increase after the ban.
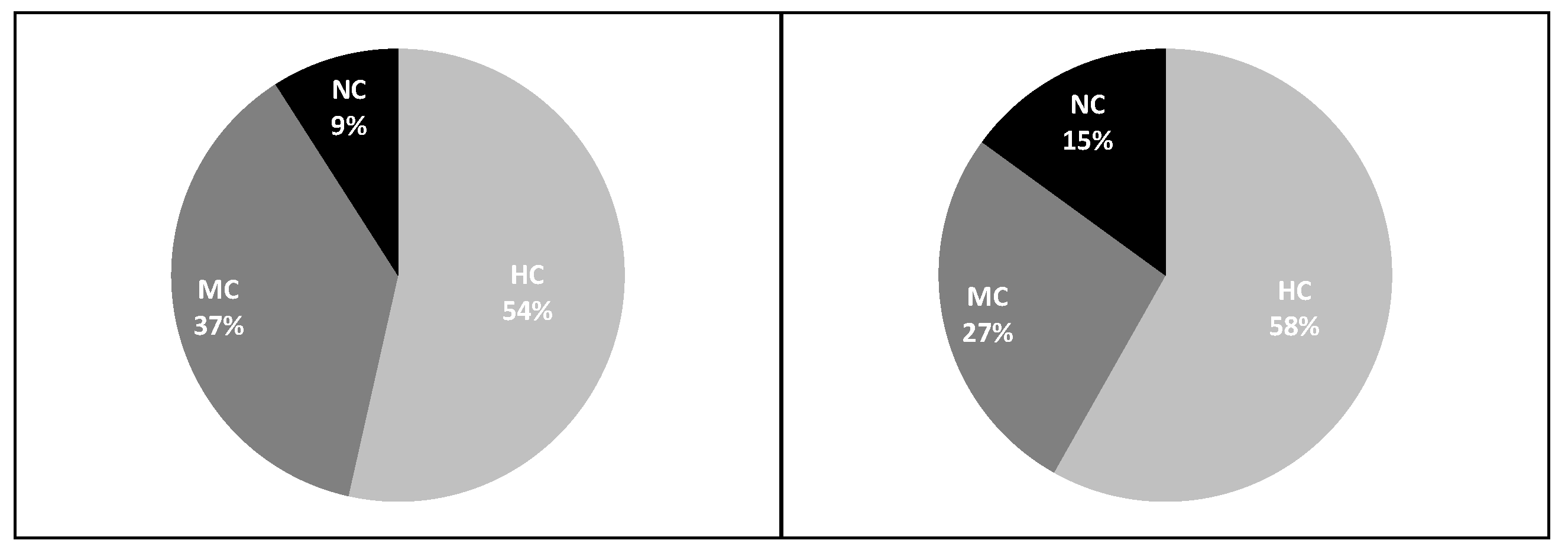
The biomass of highly commercial species (i.e., generally large-sized, long-living demersal finfish) is supposed to increase as a result of protection, while that of noncommercial species is expected to remain unchanged or even to increase in exploited assemblages, due to the selective pressure exerted on larger, more valuable species [
2,
92]. Our second hypothesis was based on this postulate and was largely confirmed by our findings. The percentage contribution of HC and NC species to the total survey catch in the GCAST has slightly increased after the ban, while the MC species have remarkably decreased. Larger biomass of noncommercial species was also reported from a no-trawl area in the NW Aegean when compared to areas with medium and high level of fishing pressure [
52]. These results might be due to the long-term protection allowed to soft-bottom shelf communities in the GCAST and the NW Aegean: in the absence of trawling impact, the benthic and nekto-benthic fish assemblages have recovered, so also the smaller, noncommercial fishes living in those habitats could rebuild and thrive.
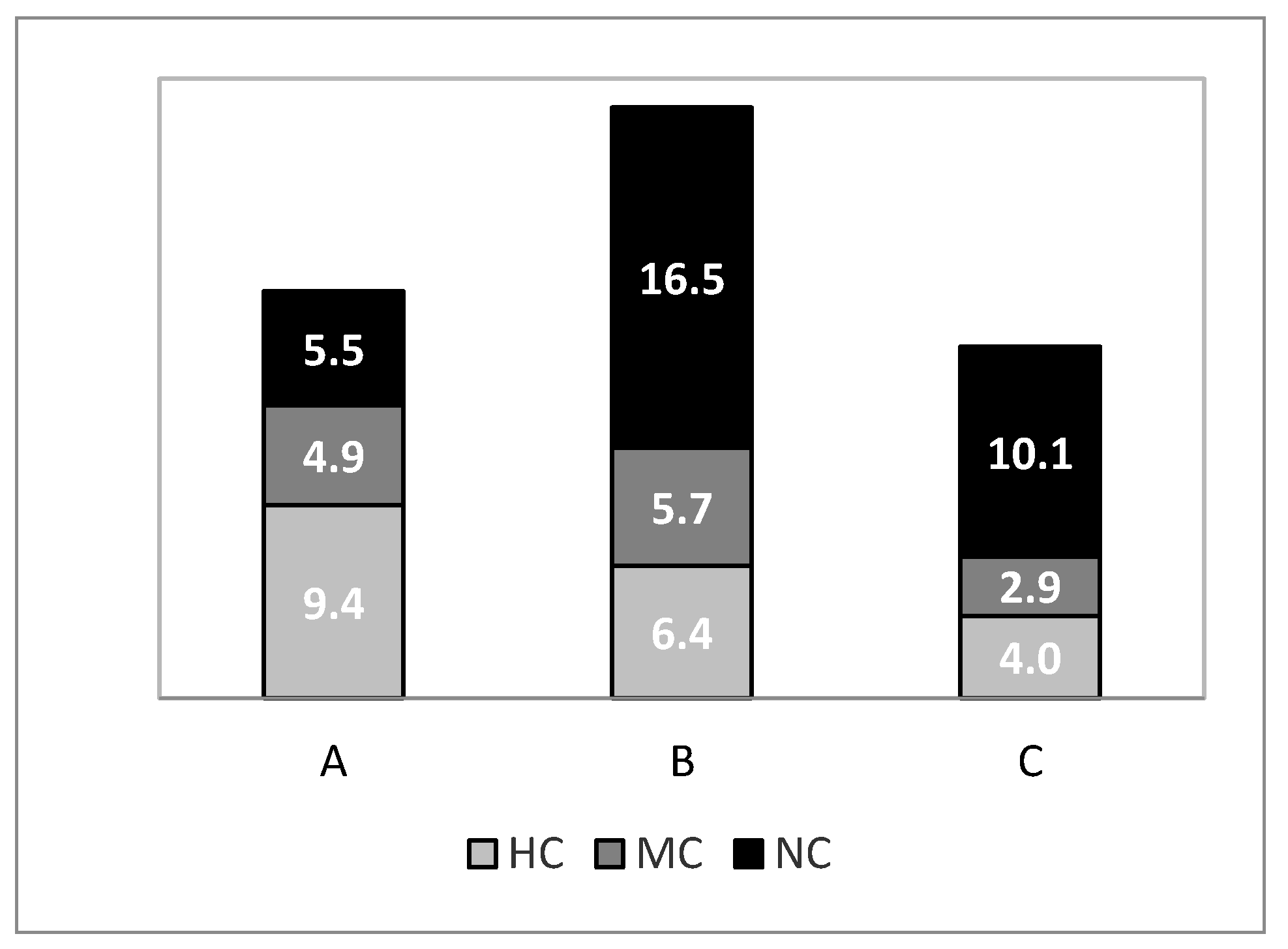
The increased and larger biomass of HC species, coupled with a lower biomass of NC species at lower depths (stratum A) offers potential economic advantages to artisanal fishermen in the GCAST, who operate largely within the -100 m isobath (pers. observ.). Trawl bans in coastal areas may provide an increase of income to artisanal fishermen [
29] [
34] because of fish biomass increase and among-fleets conflict reduction [
80], and an opportunity to expand the fishing areas in the absence of competing trawlers [
35], especially when coupled with economic incentives meant to keep a low trawling intensity also on adjacent grounds [
93].
4.2. Size
Fish populations are expected to contain more abundant older and larger fish and have a wider, less contracted size distribution when subject to full or partial protection [
17,
94]. A reduction of fishing pressure can also lead to the survival and increased abundance of large females, which are able to produce more and better eggs and to contribute to higher recruitment [
95], and provide the conditions for a recovery from overexploitation, as suggested by Fiorentino et al. [
45] for red mullet in the Gulf of Castellammare.
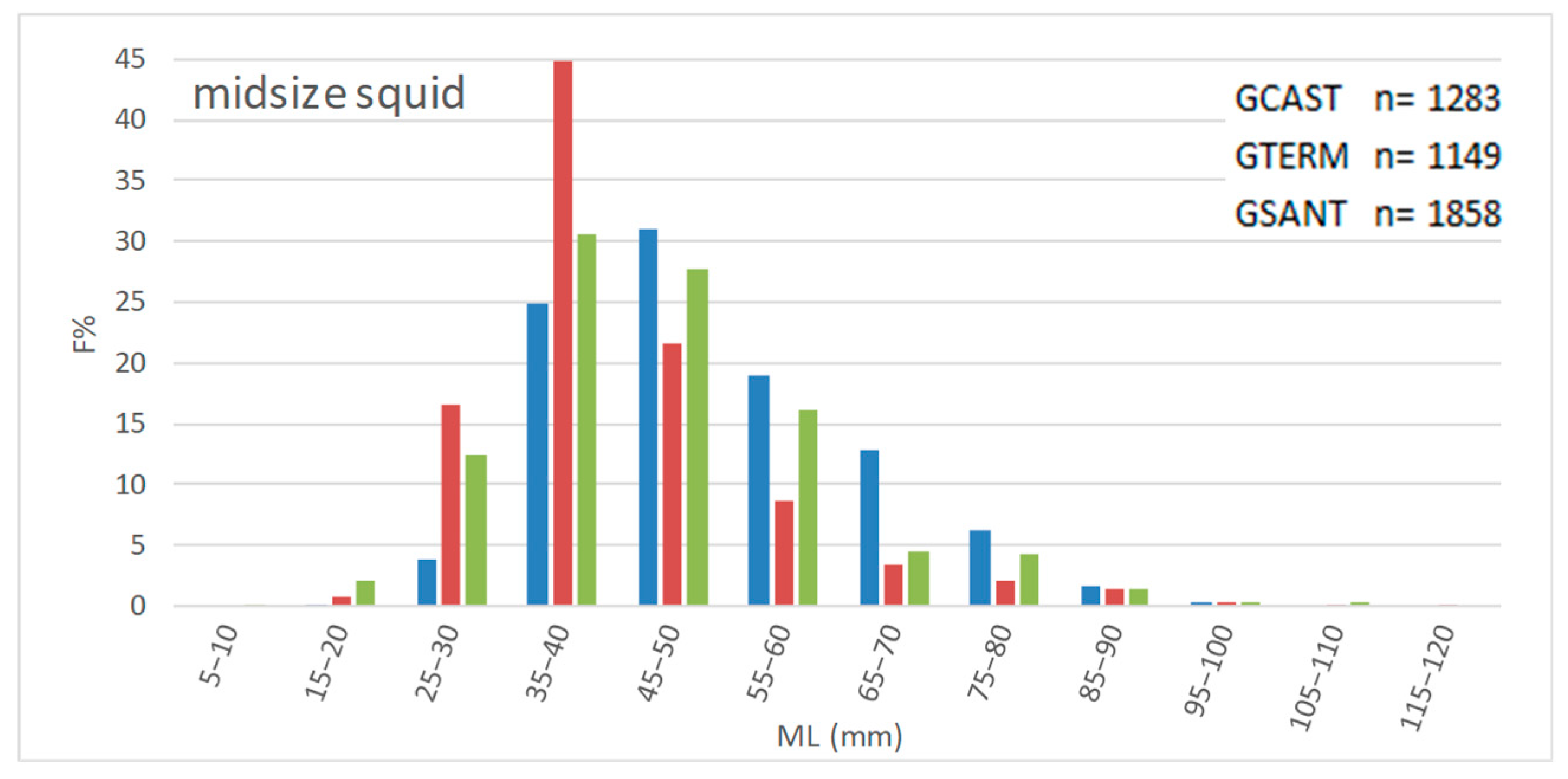
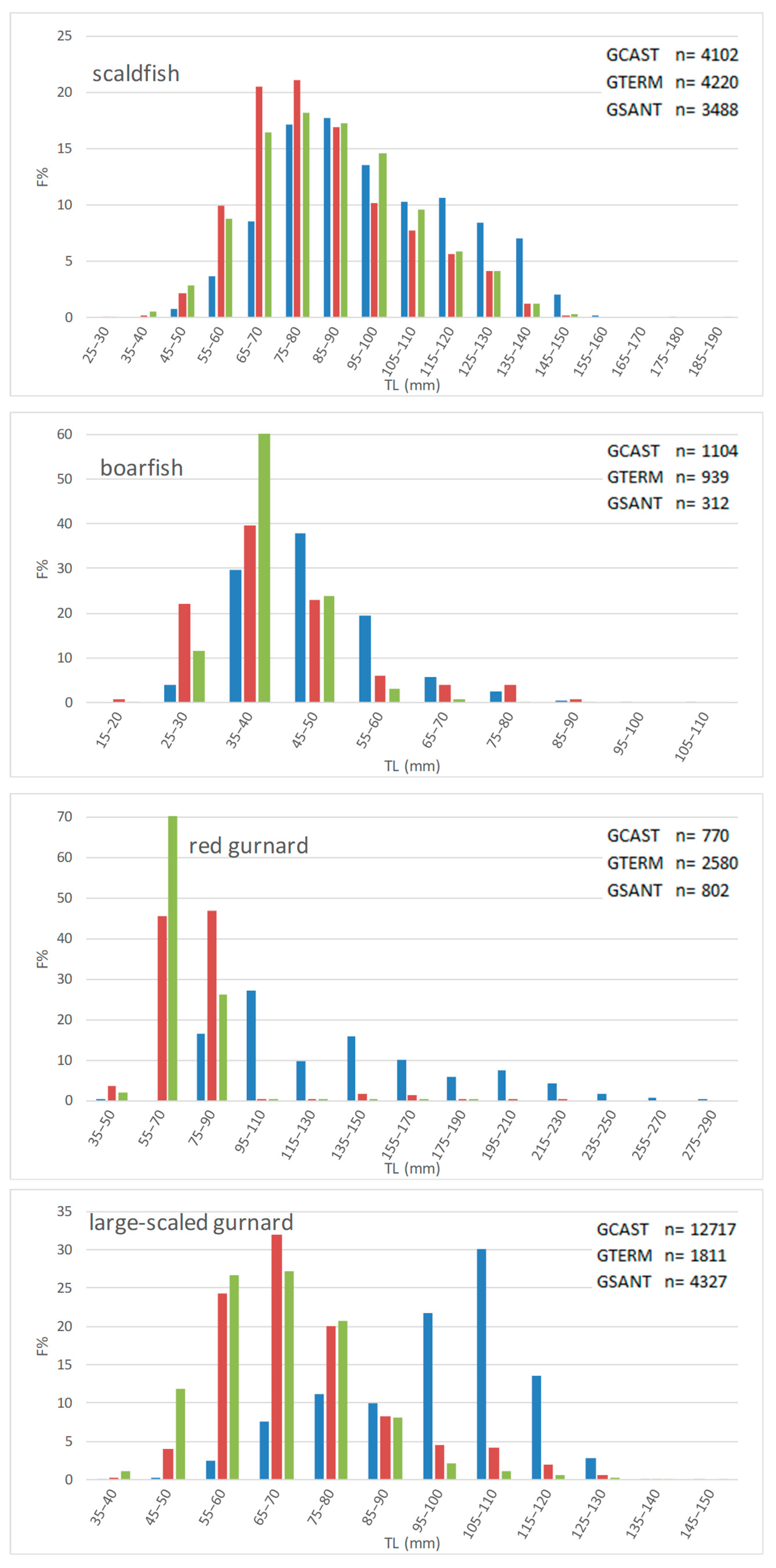
To assess the effect of the trawl ban on the size structure of demersal fish populations, we compared LFDs and some size-based indices in the GCAST and in two trawled areas comparing length data collected 15 years after the start of the ban. Overall, our third hypothesis has been partially confirmed by data.
LFDs were different in all possible pairs of gulfs, showing differences for each species not only between the GCAST and each of the GTERM and the GSANT but also between the two trawled gulfs—except for elegant cuttlefish in GTERM vs. GSANT. The size-based indices displayed different trends in the three gulfs although values were generally higher in the no-trawl area. L
m was higher in the GCAST for all species except broadtail squid and deep-water rose shrimp, whereas L
95 and L
2/3 were higher in the GCAST for seven and nine species out of fourteen, respectively, indicating a presence of large individuals of some species in the trawled gulfs. As regards the last two indices, the observed situation is not easy to interpret but it does not seem to be directly related to bio-ecological traits, since species with different life span and different habitat had higher values in the trawled gulfs. It is not unlikely that not only fishing pressure, but possibly also unknown site-specific variables or density-dependent effects may have affected the demographic structure of the fished populations. A complex interaction between resource availability and density-dependence [
81,
96] may have played a role in the growth performance of some species in our study areas, as well as the pressure exerted by artisanal fishermen targeting a small number of benthic and bentho-pelagic species—especially red mullet, pandora and cephalopods—using size-selective fishing gear [
35,
97].


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}