Abstract
Plants of the Ambrosia genus are invasive and cause many ecological problems, including the oppression of the growth of agricultural crops and native plants, land depletion, and the production of strong allergens. The use of weeds as a sustainable feedstock for biogas production, either methane or hydrogen, is a promising way to fulfill the energy needs of the current generation, eliminate the depletion of non-renewable carbon resources, and preserve the ecosystem degradation caused by invasive species impacts. A diversified microbial community was used as inoculum and Ambrosia artemisiifolia L. biomass as a substrate for anaerobic degradation and methane production. In this regard, the development of biotechnological approaches to ragweed degradation will promote the integration of new renewable energy systems. Herein, we have shown the high effectiveness of combining the processes of anaerobic degradation of plant biomass for methane production and detoxification of meal-containing model sewage by a diversified microbial community. Thus, the maximum methane yield was 56.0 L kg−1 TS. The presence of 500 mg L−1 Cu(II) slightly inhibited methane synthesis, and the methane yield was 38.4 L kg−1 TS. In contrast to a diversified microbial community, the natural microbiome of ragweed almost did not synthesize methane and did not degrade plant biomass (Kd = 2.3). Methanogens effectively immobilized Cr(IV), Cu(II), and Fe(III) during ragweed fermentation at initial concentrations of 100–200 mg L−1. The obtained results showed the high effectiveness of applying a diversified microbial community in a sewage treatment plant for the degradation of a noxious plant, Ambrosia artemisiifolia L.
1. Introduction
Noxious weeds are widespread in the countries of Europe [1], America, and Asia and pose a significant danger to the environment [2,3]. The toxic metabolites of invasive plants have a greater impact on the plants and microorganisms that live in those biogeocenoses. Many of them, for example, the Ambrosia genus, also pose a danger to human health, causing allergic reactions as well as skin and respiratory tract irritation. In addition, invasive weeds can rapidly colonize agricultural and urban lands, inhibit the growth of valuable agricultural crops (allelopathic effect), and reduce the harvest [4,5,6]. Thus, the problem of spreading these plants entails serious economic and social consequences. The plant Ambrosia artemisiifolia (family Asteraceae) is an especially noxious weed that can cause significant human health damage [7]. This weed is a monoecious, wind-pollinated, annual herb native to North America whose height varies from 10 cm to 2.5 m, depending on environmental conditions [8]. Ragweed is the most common alien species found in Europe, producing a large amount of highly allergenic pollen [9]. Before the appearance of control methods for Ambrosia-induced allergies in Europe, 13.5 million people were suffering from them, costing the economy approximately 7.4 billion EUR per year [10]. The benefit of biological control of ragweed using the leaf beetle Ophraella communa has also been proven [10]. However, this still does not help to solve the global problem of the spread of ragweed in the world [11]. Suppression of the development of ragweed is caused only by strong herbicides [12], after which the land becomes unsuitable for use for many years. This also conferred resistance to several herbicides targeting photosystem II, protoporphyrinogen oxidase, 5-enolpyruvylshikimate-3-phosphate synthase, and/or acetolactate synthase (ALS) on Ambrosia artemisiifolia L. [12]. Therefore, mechanical removal is one of the most common methods of ragweed control [13]. Most European cities spend significant budget funds on the mechanical removal and disposal of invasive weeds. These methods partially allow for the avoidance of their harmful effects on the environment and human health. However, after removal, the plant biomass is typically either buried in landfills or subjected to mechanical and chemical processing.
We consider ragweed biomass a valuable energy resource [13]. The feedstocks generated from weed biomass are sustainable sources for the production of biologically active substances (such as flavonoids, phenolic compounds, vitamins, etc.) [14] or for biogas production (methane or hydrogen). This weed can be used as a source of organic substances, in particular carbohydrates, which will ensure high efficiency in biogas production. The concentration of cellulose, lignin, and hemicellulose in the ragweed biomass ranges from 25.3% to 30.9%, 24.2% to 28.3%, and 13.1% to 17.7%, respectively [13]. Due to the high content of these compounds, ragweed is extremely valuable for the microbial synthesis of biofuels (H2 and CH4 gases). Lignocellulosic biomass is the earth’s most abundant renewable feedstock that can be degraded by different microorganisms. Methane fermentation of organic biomass is performed in the syntrophic association between fermenting bacteria and methanogenic archaea [15,16]. Methane degradation of cellulose is carried out by several microorganisms with cellulolytic activity (bacterial genera of Bacillus (B. licheniformis, B. cereus, B. subtilis, Pseudomonas, and Serratia) [17]. Cellulose is degraded to monomers, simpler carbohydrates (glucose), which are a substrate for other microorganisms that synthesize hydrogen (Clostridium sp.), methane (Methanosarcina, Methanobacterium), and carbon dioxide (CO2). Thus, microbial conversion of organic biomass (biopolymers) is a multistage process that takes place with the participation of microorganisms in a syntrophic association: degradation of cellulose (hydrolysis), acidogenesis, acetogenesis, and methanogenesis [18]. The diversified microbial community of sludge from wastewater treatment plants contains all the necessary physiological groups of microorganisms (aerobes, facultative anaerobes, and strict anaerobes) in syntrophic association and is adapted to the gradual degradation of plant biomass [19]. Given the wide diversity of this community, it is capable of effective detoxification of toxic metals, in particular copper. Copper is a widespread pollutant that accumulates both in the environment and in the sewage of industrial enterprises [20]. The combination of the processes of degradation of noxious weeds, synthesis of methane, and degradation of biomass is a new and promising scientific approach that simultaneously solves three environmental problems: disposal of invasive plants, lack of energy fuel, and metal pollution. Despite the significant interest of scientists and the huge potential for beneficial use of invasive weeds in combination with their utilization, the scope of their application is usually limited to the mechanical treatment of the territories.
This work aimed to confirm experimentally the capability of ragweed degradation, methane production, and simultaneous detoxification of sewage from toxic compounds of heavy metals (copper, chromium, and iron) by a diversified microbial community. The objective was to evaluate the biotechnological potential of ragweed as a new renewable energy source.
2. Materials and Methods
2.1. Ragweed Samples Preparation for the Methane Production
Ragweed (Ambrosia artemisiifolia L.) dried biomass was used as a feedstock for CH4 production and heavy metal immobilization. Ragweed biomass was collected in September 2022 at Sofiivska Borshchahivka village (Kyiv region, Ukraine) during active flowering (Figure 1).

Figure 1.
The population of Ambrosia artemisiifolia L. in the Sofiivska Borshchahivka village, Kyiv region, Ukraine: (a,b) July 2022; (c,d) September 2022.
The weed was taken from the soil and dried at 30 °C until it reached a constant weight. The whole plant (the root and aerial part) was used for research. The plant biomass was crushed to a size of 0.5–0.8 cm before the anaerobic degradation (Figure 1d).
2.2. Extraction of Ambrosia artemisiifolia L. Bioactive Compounds
A porcelain mortar was used for grinding. Extraction was performed after complete grinding in a mortar. The extracts were prepared in a Soxhlet extractor (100 mL) using 7.2 g of air-dried plant material (the whole plant) in 70% ethanol (250 mL) for 12 h within 50 full cycles. The dry matter content of each extract was determined in five repetitions by the oven-drying method at 105 °C to a completely dry weight [21].
2.3. Measurement of Phenols, Flavonoids, and Total Carbohydrate Concentration and Antioxidant Activity of the Extract of the Whole Plant of Ambrosia artemisiifolia L.
The ragweed biomass contains several organic substances, in particular phenols and carbohydrates, which determine the efficiency of anaerobic fermentation and methane synthesis. Total phenols were measured by the Folin-Ciocalteu reagent and presented as mg of GA/g of extract in terms of GAE (gallic acid equivalent). The standard curve equation is y = 0.9757x − 0.0037, r2 = 0.998) [22]. The total flavonoid content was calculated by a modified aluminium chloride colorimetric assay and presented in RUE (rutin equivalent) as mg of RUE/g of the extract [23]. The standard curve equation is y = 0.7791x − 0.0115, r2 = 0.997). The antioxidant activity of the extracts (AOA) was resolved by the DPPH method with the 2,2-diphenyl-1-picrylhydrazyl reagent (DPPH) [24]. The parameter of antioxidant activity was presented in percentages (%). The calculation of carbohydrates in the ragweed extracts was performed via the phenol-sulfuric acid method [22,25] (the standard curve equation: y = 1.5954x − 0.0194, r2 = 0.992). The content of the presented substances was calculated as mg g−1 of totally dry extract.
2.4. Inoculum Obtaining
Sludge from the sewage treatment plant in Kyiv was used as an inoculum and source of a diversified microbial community (DMC). It contained a large number of different physiological groups of microorganisms with cellulolytic activity as well as methane-producing bacteria [26,27]. The samples were taken in Kyiv (Ukraine) at a typical wastewater treatment plant (Bortnychi Aeration Station, Kyiv region, Ukraine). The native microbiome of the Ambrosia artemisiifolia L. weed was used as the first variant of inoculum. The sludge from the methane tank, which contained a diverse microbial community, was used as a second variant of inoculum. Substrate and inoculum were introduced into anaerobic jars at a concentration of 4% of the total reaction volume. The jars were closed hermetically and placed in a thermostat at a temperature of 30 °C for 75 days. The presence of methanogens was checked by measuring the concentration of CH4 in the gas phase [28] by gas chromatography [29]. The data were presented as mean values with standard deviations (n = 3).
2.5. Determination of the Main Cultural Parameters of Ambrosia artemisiifolia L. Microbial Degradation
To determine the patterns of Ambrosia degradation by a diversified microbial community (in syntrophic association), we performed five variants of the experiment: The first was the control variant (degradation of the ragweed by the native microbiome of Ambrosia artemisiifolia L. without using additional inoculum). The second variant was experimental (degradation of Ambrosia artemisiifolia L. by a diversified microbial community). The diversified microbial community was also used as an inoculum in the third, fourth, and fifth variants. Copper(II), chromium(VI), and iron(III) were inserted into different anaerobic jars in the active phase of degradation (42 days) at a concentration of 500 mg L−1. The active phase of fermentation was observed on the 42nd day of degradation, during which a significant amount of anaerobic biomass was present along with a low redox potential and active methane synthesis. These conditions were favorable for the microbial immobilization of metals.
Several metabolic parameters were measured, including cumulative gas production (mL), gas phase concentration (% of CH4 and CO2), dissolved organic carbon concentration (DOC, mg L−1), pH, and redox potential (Eh, mV). The degradation was performed in anaerobic jars with a volume of 500 mL. The jar was filled with 20 g of air-dried and crushed Ambrosia artemisiifolia L. weed and 400 mL of tap water. The duration of the degradation process was 75 days at 30 °C.
The H2, CH4, and CO2 concentrations were measured via the gas chromatography method [30]. The efficiency of methane synthesis was expressed in liters per kilogram of total solids (L kg−1 TS). The EZODO MP-103 universal ionomer was used to measure the Eh and pH of the medium [31,32].
The concentration of soluble organic compounds was determined via the permanganate method and presented as the medium’s dissolved organic carbon content [33]. Carbon concentration was determined according to standard calibration curves: y = 0.12x − 12.178, r2 = 0.994.
2.6. Determination of the Dynamics of Copper Immobilization during the Degradation of Ambrosia artemisiifolia L.
The capability of a diversified microbial community (DMC) to immobilize metal ions during ragweed degradation was investigated. The stock solutions were 40,000 mg L−1 Cu(II), Cr(VI), and Fe(III) prepared by dissolving CuSO4∙5H2O, K2CrO4 or FeSO4·5H2O in distilled water. The Fe(II) solution was treated with hydrogen peroxide for complete iron oxidation. Sodium citrate was used as a chelator to stabilize Cu(II) and Fe(III) in a soluble state in the culture fluid. The Cu(II) concentration was determined via the colorimetric method with 4-(2-pyridylazo)resorcinol (PAR). The methods are based on the property of PAR to form red-colored complexes with cations of bivalent heavy metals, including Cu2+ [34]. The concentration of Cr(VI) in the cultural fluid was measured via the colorimetric method with DFC (1,5-diphenylcarbazide) [35]. A colorimetric method using qualitative reactions with o-phenanthroline and KSCN was used to determine the concentrations of Fe(II) and Fe(III) in the medium, respectively [36]. These reactions are based on the formation of red-colored complexes of KSCN with Fe(III) and o-phenanthroline with Fe(II). On the 42nd day of degradation, metal solutions containing Cu(II), Cr(VI), and Fe(III) were added to the different sealed glass jars to the final concentrations of 500 mg L−1. The experiment was carried out in triplicate (n = 3). Standard deviation (SD) and average values (x) were calculated.
3. Results
3.1. Bioactive Compound Content in the Extracts of Ambrosia artemisiifolia L.
The concentration of widespread exometabolites of ragweed was studied in this work. Both soluble organic compounds (sugars, flavonoids, phenols, etc.) and insoluble ones (cellulose, lignin-cellulose, etc.) will determine the efficiency of organic biomass degradation and methane synthesis [37]. Herein, we focused on determining the concentration of soluble organic compounds (which are highly available for microorganisms) in the prepared ragweed extract. According to literature data, the concentration of cellulose in ragweed ranges from approximately 20 to 30% [13].
Whole-plant extracts (aerial and root parts) were prepared using 70% aqueous ethanol to determine the concentrations of carbohydrates, phenols, flavonoids, and antioxidant activity. We observed that ragweed extracts contained carbohydrates, phenols, and flavonoids. The total concentration of carbohydrates in the ragweed was 0.7 mg mL−1 of extract or 303.1 mg g−1 of dry matter. The flavonoid concentration was 0.5 mg mL−1 or 186.3 mg g−1 of extract. The total concentration of the phenols amounted to 0.7 mg of GAE mL−1 or 275.7 mg of GAE g−1 of extract. In terms of the dry mass of ragweed, the concentration of phenolic compounds, flavonoids, and carbohydrates (sugars) was quite high and amounted to 21.5 ±3.5, 14.5 ± 2.3, and 23.7 ± 1.5 mg g−1 of plant, respectively.
The concentrations of the investigated bioactive compounds in the extract (mg mL−1, mg g−1 of extract) and ragweed biomass (mg g−1 of the plant) are presented in Table 1.

Table 1.
Total carbohydrates, phenol, and flavonoid concentrations in the extract and biomass of Ambrosia artemisiifolia L.
In addition, the level of antioxidant activity of the ragweed extracts was also very high and amounted to 86.3%. In this way, we ensured that our plants contained a sufficient amount of organic compounds, in particular carbohydrates. The high content of carbohydrates indicates the capability of anaerobic fermentation of the ragweed substrate with high efficiency. Therefore, we hypothesized that ragweed would be efficiently degraded by a diversified microbial community with methane production.
3.2. Immobilization of Copper during the Degradation of Ragweed Biomass
Solutions of copper, chromium, and iron were added into the anaerobic jars (bioreactor) with tap water and Ambrosia artemisiifolia L. dried biomass at the 42nd hour of cultivation to the final concentrations of 100, 200, 500, and 1000 mg L−1 in the active phase of microbial growth. It was necessary to determine the level of resistance of the microbial community and the concentration at which maximum heavy metal immobilization will occur. Immobilization occurred in the anaerobic jars due to the active growth of a diversified microbial community (inoculum). The active phase of degradation was dominated by methanogenic microorganisms that consumed the hydrolysis products of ragweed (acetate, glucose, and hydrogen) and synthesized methane (CH4) and carbon dioxide (CO2). Methanogenic microorganisms actively immobilized copper compounds in the active phase of growth at concentrations of 100, 200, and 500 mg L−1 (Figure 2).
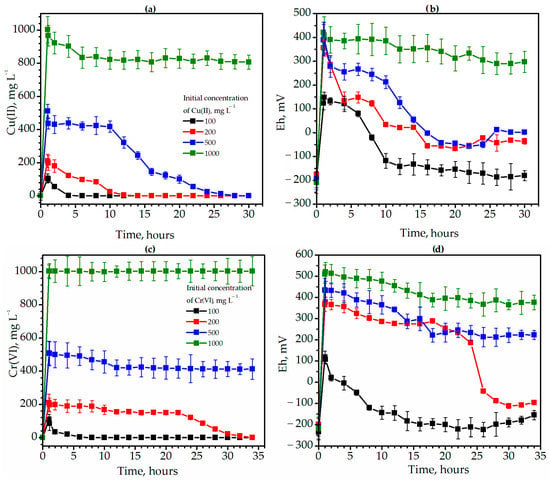
Figure 2.
The dynamics of Cu(II) (a) and Cr(VI) (c) immobilization and Eh (b,d) during degradation of Ambrosia artemisiifolia L. weed in the presence of 100 (black lines), 200 (red lines), 500 (blue lines), and 1000 (green lines) mg L−1 Cu(II) (a,b) and Cr(VI) (c,d).
The efficiency of immobilization in the variants with 100, 200, and 500 mg L−1 Cu(II) was 100%. As expected, the fastest immobilization occurred in the presence of 100 mg L−1 Cu(II) (Figure 2b, black lines). Thus, the Cu(II) concentration decreased twice, from 105.4 to 55.3 within two hours and from 105.4 to 2.2 mg L−1 within 4 h. The duration of complete immobilization was 6 h. Cu(II) compounds also precipitated gradually at the other concentrations. However, the immobilization durations were 12 and 30 h at 200 mg L−1 and 500 mg L−1, respectively. The concentration of 1000 mg L−1 Cu(II) of copper was found to be detrimental to the microbial community. The efficiency of immobilization was only 19.5% in the variant with 1000 mg L−1 Cu(II) concentration. The Cu(II) concentration decreased only from 1003 mg L−1 to 807 mg L−1. After the introduction of 1000 mg of Cu(II), the redox potential increased sharply from −210 mV to +421 mV, after which the microbial community could not adapt to the action of this extreme factor (Figure 2a).
Chromium turned out to be more toxic than copper. Thus, effective immobilization occurred only in the presence of 100 and 200 mg L−1 Cr(VI) (Figure 2c). The duration of complete immobilization was only 8 h for 100 mg L−1 Cr(VI) and 34 h for 200 mg L−1. The Cr(VI) concentration decreased from 211 to 155.1 mg L−1 in 12 h and to 22.2 mg L−1 in 30 h (Figure 2c). In variants with 500 mg L−1 of Cr(VI), only partial immobilization of chromium occurred with an efficiency of 19.0%. Thus, the concentration of Cr(VI) decreased from 509 to 412.5 mg L−1 during the 26 h of growth of the methanogenic microbial community and remained stable thereafter. The introduction of 1000 mg L−1 Cr(VI) led to a complete cessation of microbial growth.
The insertion of chromium also led to a significant increase in the redox potential of the medium. The Eh increased from −234 mV to +118 mV and from −218 to +510 mV in the variants with 100 and 1000 mg L−1 Cr(VI), respectively (Figure 2d).
The methanogenic microbial community immobilized iron with the lowest efficiency compared to copper and chromium immobilization. Methanogens reduced Fe(III) to Fe(II), which then precipitated as insoluble compounds of divalent and trivalent iron (Figure 3).
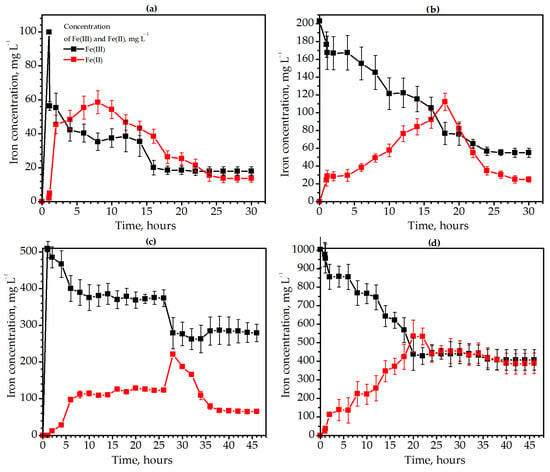
Figure 3.
The dynamics of the iron (Fe(II) and Fe(III)) immobilization by the methanogenic microbial community during degradation of Ambrosia artemisiifolia L. weed at the initial concentrations of 100 (a), 200 (b), 500 (c), and 1000 (d) mg L−1 Fe(III).
Thus, the immobilization efficiency was 66.4% in the variant with 100 mg L−1 Fe(III). The concentration of Fe(III) decreased from 99.8 to 56.4 mg L−1 and the concentration of Fe(II) increased from 0 to 45.5 mg L−1 during the 2 h after metal insertion. However, soluble compounds of Fe(II) were gradually immobilized, and the concentration of Fe(III) and Fe(II) in the medium was 18 and 15.5 mg L−1 after 24 h of iron insertion, respectively. In the variant with 200 mg of Fe(III), the immobilization began only 18 h after the insertion of iron. Initially, iron was reduced from Fe(III) to Fe(II). Thus, at 18 h, the solution contained 76.7 mg L−1 of Fe(III) and 112.4 mg L−1 Fe(II). After that, iron began to precipitate, and at 28 h, 55 mg L−1 Fe(III) and 25.1 mg L−1 Fe(II) remained in the solution (Figure 3b). The efficiency of immobilization was 60.5%. Similar patterns of iron transformation were observed in the variants with 500 and 1000 mg L−1 Fe(III). The effectiveness of immobilization was 38.3% (46 h) and 20.5 (40 h), respectively. In contrast to copper and chromium, iron did not cause significant changes in the redox potential after insertion into a metabolically active medium. The redox potential remained at the optimal level for methanogens (from −150 to −220 mV) in the presence of Fe(III) in the amount of 100–1000 mg L−1.
3.3. The Dynamics of the Metabolic Parameters of the Microbial Community during the Degradation of Ragweed Biomass
The main metabolic parameters of ragweed degradation in different experimental variants were studied. The first variant was the control, where only the natural ragweed microbiome was used as the inoculum. In the second and third variants, degradation occurred via a diversified microbial community (the source, the sewage treatment plant in Kyiv), but the solutions of heavy metals were added to the bioreactor to the final concentration of 500 mg L−1. The patterns of the influence of heavy metals (Cu(II), Cr(VI), and Fe(III)) on the metabolism of the diversified microbial community were similar. In this case, the graphs show a variant of the effect of copper on the main metabolic parameters of microbial growth. The dynamics of pH, Eh, DOC (dissolved organic carbon), as well as ammonium ions (NH4+), were studied (Figure 4). In control conditions with the natural ragweed microbiome, the pH decreased sharply and fluctuated between 5.2 and 5.8 starting from the 7th day of the degradation (Figure 4a).
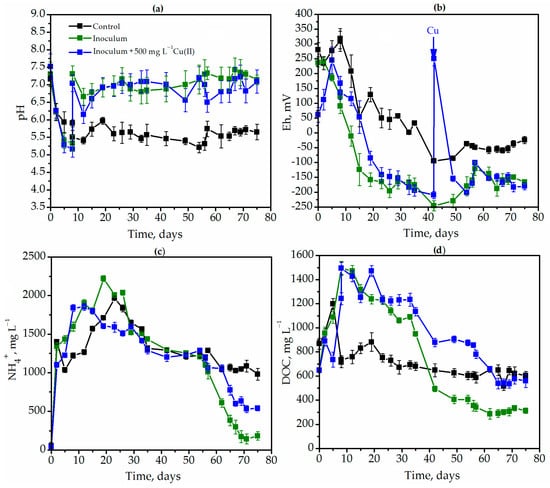
Figure 4.
The dynamics of pH (a), redox-potential (b), concentrations of ammonium ions (c) and dissolved organic carbon (d) during Ambrosia artemisiifolia L. degradation in different experimental conditions: the black lines represent the degradation in control conditions by the native microbiome; the green lines represent degradation by DMC; and the blue lines represent degradation by DMC in the presence of 500 mg L−1 Cu(II).
Acidification could occur as a result of the partial hydrolysis of plants or the digestion of the available sugars, which results in the formation of organic acids. In the absence of a diversified microbial community, microorganisms capable of consuming the organic acids as a substrate were absent (in contrast to experimental variants with DMC inoculum). In the variant with DMC inoculum, a significant increase in the pH index to neutral values was observed from the 7th to the 10th day, which indicates a high metabolic activity of microorganisms and the accumulation of alkaline ions in the solution. Autoregulation of the pH index to more optimal values indicates the presence of various physiological groups of microorganisms in the bioreactor.
A similar phenomenon also occurred with the pH index. The microbial community decreased the redox potential to a level more optimal for the growth of strict anaerobes and methane synthesis compared to the control (Figure 4b). In the control treatment, the Eh decreased only to −75 mV (Figure 4, black lines), which is not low enough for the development of strict anaerobic methanogens. However, in the variant with DMC, Eh decreased to −250 mV and remained at a low and optimal level for methanogens till the end of the degradation (Figure 4b, green lines). Adaptation of the DMC to cultivation conditions was also manifested in the variant under the copper influence. Before the copper insertion, the redox potential dropped to −200 mV. The copper presence led to a sharp increase in the Eh from −200 to +250 mV and a slowdown in the growth of anaerobic microorganisms (Figure 4b, blue lines). However, the Eh again decreased to more optimally negative values within 15 h, and by day 50, it was −150 mV (Figure 4b, blue lines). Intensive hydrolysis of plant biomass was confirmed by an increase in the concentration of ammonium ions and DOC in the culture liquid. The initial concentration of ammonium ions in all experimental variants ranged from 42.9 to 54.2 mg L−1 and increased to 1101.6–1399.3 mg L−1 within two days (Figure 4c). Ammonium ions accumulated in the medium for 26 days. For example, in the variant with the inoculum on the 19th day, it was 2220.8 mg L−1. However, on the 75th day, the concentration had already decreased from 2220.8 mg L−1 to 183.5 mg L−1. Fluctuations in the concentration of organic compounds were also observed during the degradation process. In the control variant, the concentration increased from 867.8 to 1197.2 mg L−1 on the 5th day and slightly decreased to 603.0 mg L−1 (Figure 4d) on the last day of degradation. In the experimental variant with the DMC inoculum, the concentration of DOC increased to 1493.4 mg L−1 and remained at this level for up to 8 days (Figure 4d, green lines). However, the diversified microbial community actively grew and synthesized biogas. On the 75th day, the DOC concentration decreased to 312 mg L−1 as shown in Figure 4c. Copper inhibited the transformation of the substrate compared to the experimental variant under metal-free degradation by DMC. However, after the complete immobilization of copper, substrate consumption resumed. On the 75th day of degradation, the concentration of DOC decreased from 1485.5 mg L−1 (8 days) to 559.8 mg L−1 (Figure 4c).
3.4. Methane Production and the Efficiency of Ragweed Degradation
Intensive degradation of ragweed biomass was also confirmed by significant methane synthesis in experimental variants where the inoculum was DMC from a sewage treatment plant (Figure 5a). Active synthesis of methane began on the 23rd day, and on the 35th day its concentration was 48.8%; on the 69th day, it was 64.8%. In the control variant without inoculum, methane was synthesized only at a residual concentration that did not exceed 15.3% (Figure 5a). Cumulative methane production in the control was only 130 mL in the variant with DMC inoculum and under the influence of copper, it was 1120 mL and 735 mL. Thus, copper inhibited methane production by 1.5 times (Figure 5c). Similar patterns of changes in the concentration and carbon dioxide synthesis were also observed in both the control and the experimental variants (Figure 5b,d). The cumulative production of CO2 in the control was 170 mL, in the variant with DMC inoculum, 1350 mL, and in the variant with copper, 1134 mL (Figure 5d).
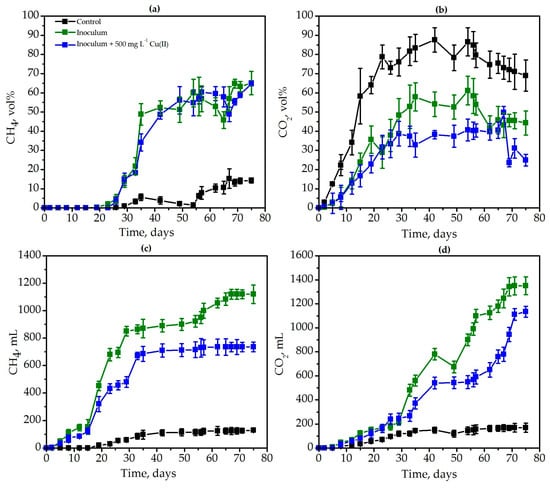
Figure 5.
The dynamics of the CH4 (a) and CO2 (b) concentration as well as methane (c) and carbon dioxide (d) production during the degradation of Ambrosia artemisiifolia L. weed: the degradation in control conditions with the native microbiome (black lines), the degradation by inoculum (DMC, green lines), as well as under the influence of 500 mg L−1 Cu(II) (blue lines).
The methane and carbon dioxide yields during the degradation of ragweed biomass were calculated. This also confirmed the high efficiency of ragweed degradation by a diversified microbial community (Table 2).
Table 2.
The effectiveness of ragweed degradation in the presence of heavy metals.
Thus, the effective degradation of ragweed by a diversified microbial community with a high yield of CH4 was demonstrated. It was determined that the ragweed biomass was actively degraded in the absence of additional co-substrates in the variants with DMC inoculum. Intensive degradation of the ragweed could be ensured by the presence of syntrophic relationships in the microbial community, in which the exometabolites of some microorganisms (hydrolytics and cellulolytics) are substrates for others (methanogens). Copper inhibited the development of the microbial community; however, it was completely immobilized by microorganisms at a concentration of 500 mg L−1. After copper immobilization, the metabolic activity of DMC was restored. Iron did not suppress methane synthesis or the growth of microorganisms.
4. Discussion
The study of beneficial properties of microorganisms and plants is currently of great interest to both science and industry [38,39]. Microorganisms and their exometabolites have found diverse applications in various industrial fields. In pharmacology. They are utilized for synthesizing biologically active compounds. In the agroindustry, they are used to suppress the growth of phytopathogenic bacteria and fungi and preserve crops [40,41]. Moreover, they are employed in environmental protection biotechnologies to degrade toxic organic [42] and inorganic waste [43]. The use of microorganisms for the biological purification of soil and wastewater from toxic metals is one of the most in-demand fields [44]. This work showed the possibility of using a diversified microbial community with large species diversity and broad substrate specificity [45]. The application of such a microbial community is very promising in industrial conditions [46] for biogas synthesis or degradation of multicomponent food waste [47]. It is commonly known that various species or physiological groups of microorganisms differ in their metabolism and sensitivity to extreme factors [48]. Therefore, relying on a single microbial strain for the fermentation of various organic wastes can result in low efficiency [49]. To improve such processes, it is recommended to spread the use of expensive co-substrates, which are economically costly and unprofitable [50]. The development of methods for controlling and utilizing invasive weeds has been actively underway for the last decade, with special attention paid to aquatic invasive plants, such as water lettuce [51,52], as well as goldenrod, nettle, wormwood, etc. [53]. Several studies of biological and chemical control with the use of herbicides have already been conducted for noxious ragweed [12].
However, the attention of scientists is focused on the study of allergens produced by these weeds, the selection of invasive weed inhibitors, etc. [6]. A review of the literature shows that the most widespread method of ragweed control is its mechanical removal from the land and burial, or physical and chemical processing [54]. Our study suggests that the mechanical extraction of ragweed followed by its use as a feedstock for biomethane production and the degradation of toxic metals is a promising biological method for controlling and utilizing invasive weeds. Some experimental data have already been obtained on the possibility of using invasive plants as substrates for obtaining bioethanol [55], biomethane [56], organic acids, and other useful products. However, ragweed remains unexplored in this aspect. Recently, there has been only one study describing the high energy potential of ragweed, based on an analysis of its cellulose and lignin content [13]. Due to the presence of toxicants in the pollen and toxic exometabolites, it is believed that this plant has an allelopathic effect and can be toxic to most microorganisms [57]. However, the selection and adaptation of the microbial community for the degradation of these plants is the key to effective utilization and obtaining useful products. This is confirmed by the example of our research. Under experimental conditions, the yield of methane was 56 L kg−1 TS. The experimental or industrial production of biogas from ragweed has not been reported yet. However, in other studies, slightly more biogas was obtained during the fermentation of another invasive weed with the presence of an additional co-substrate such as food waste, cow manure, etc.). For example, the optimization of anaerobic co-digestion of Solidago canadensis L. biomass and cattle slurry was conducted, and the maximum methane production of 143.7 L kg−1 of volatile solids (VS) was obtained.
In addition, we showed that ragweed biomass contains some biologically active substances, phenolic compounds, and soluble carbohydrates. Studies showing the possibility of extracting pharmacological compounds from invasive weeds have already been published [58]. Thus, the scientific approach is versatile and does not pose an ecological danger to the environment, as, for example, in the case of the use of herbicides and the adaptation of ragweed to them. Further research in this direction will contribute to the accumulation of data on the disposal of harmful ragweed and the obtaining of useful products from it.
5. Conclusions
The obtained results demonstrate the high effectiveness of using a diversified microbial community in sewage treatment plants for the degradation of Ambrosia artemisiifolia L., a noxious plant. Microorganisms in the syntrophic association have shown promise for the utilization of ragweed and the synthesis of biogas, as well as the complete immobilization of toxic copper compounds. Ragweed biomass was effectively degraded with methane production, which indicates the capability of its use as a sustainable feedstock for renewable energy carriers. The development of a full-scale treatment plant to determine the patterns of ragweed degradation will be the next step in our study.
Author Contributions
Conceptualization, O.H. and O.T.; methodology, R.M.; software, V.H.; validation, G.G., A.T. and S.K.; formal analysis, I.B.; investigation, O.H., O.S., A.T. and S.K.; resources, R.M.; data curation, R.M.; writing—original draft preparation, O.H.; writing—review and editing, R.M.; visualization, V.H.; supervision, R.M; project administration, O.T.; funding acquisition, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by the Scientific Grant Agency of the Ministry of Education, Science, Research and Sport of the Slovak Republic and the Slovak Academy of Sciences, project No. 1/0882/21.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
O.H. is grateful to the International Visegrad Fund for support within the Scholarship Program (ID 52211282). V.H. is grateful to the National Scholarship Programme of the Slovak Republic for the support of mobility of students, Ph.D. students, university teachers, researchers, and artists (SAIA) for scholarship (ID 39277).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Leru, P.M.; Anton, V.F.; Eftimie, A.M.; Stefanut, S. Biologic Pollution Due to Ambrosia (Ragweed) Pollen in Urban Environment of Bucharest. Int. J. Environ. Res. Public Health 2022, 19, 10613. [Google Scholar] [CrossRef]
- Mangold, J.M.; Frame-Martin, S.A.; Raile, E.D. Noxious weed views and behaviors in Montana after 25 years of public education. Invasive Plant Sci. Manag. 2021, 14, 262–269. [Google Scholar] [CrossRef]
- Sivicek, V.A.; Johnson, S.A.; Jarvis, J.L.; Dreslik, M.J. Noxious and Selected Invasive Plant Populations along Illinois Toll Highway Rights-of-Way. Illinois Natural History Survey, 2019. Available online: https://www.ideals.illinois.edu/items/114183 (accessed on 15 December 2022).
- Han, C.; Shao, H.; Zhou, S.; Mei, Y.; Cheng, Z.; Huang, L.; Lv, G. Chemical composition and phytotoxicity of essential oil from invasive plant, Ambrosia artemisiifolia L. Ecotoxicol. Environ. Saf. 2021, 211, 111879. [Google Scholar] [CrossRef] [PubMed]
- Šućur, J.; Konstantinović, B.; Crnković, M.; Bursić, V.; Samardžić, N.; Malenčić, D.; Prvulović, D.; Popov, M.; Vuković, G. Chemical Composition of Ambrosia trifida L. and Its Allelopathic Influence on Crops. Plants 2021, 10, 2222. [Google Scholar] [CrossRef]
- Bonea, D.; Bonciu, E.; Niculescu, M.; Olaru, A.L. The allelopathic, cytotoxic and genotoxic effect of Ambrosia artemisiifolia on the germination and root meristems of Zea mays. Caryologia 2018, 71, 24–28. [Google Scholar] [CrossRef]
- Agnew, M.; Banic, I.; Lake, I.R.; Goodess, C.; Grossi, C.M.; Jones, N.R.; Plavec, D.; Epstein, M.; Turkalj, M. Modifiable Risk Factors for Common Ragweed (Ambrosia artemisiifolia) Allergy and Disease in Children: A Case-Control Study. Int. J. Environ. Res. Public Health 2018, 15, 1339. [Google Scholar] [CrossRef]
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Chauvel, B.; Dullinger, S.; Fumanal, B.; Guisan, A.; et al. Biological Flora of the British Isles: Ambrosia artemisiifolia. J. Ecol. 2015, 103, 1069–1098. [Google Scholar] [CrossRef]
- Gerber, E.; Schaffner, U.; Gassmann, A.; Hinz, H.L.; Seier, M.; Müller-Schärer, H. Prospects for biological control of Ambrosia artemisiifolia in Europe: Learning from the past. Weed Res. 2011, 51, 559–573. [Google Scholar] [CrossRef]
- Schaffner, U.; Steinbach, S.; Sun, Y.; Skjøth, C.A.; de Weger, L.A.; Lommen, S.T.; Augustinus, B.A.; Bonini, M.; Karrer, G.; Šikoparija, B.; et al. Biological weed control to relieve millions from Ambrosia allergies in Europe. Nat. Commun. 2020, 11, 1745. [Google Scholar] [CrossRef]
- Makra, L.; Matyasovszky, I.; Hufnagel, L.; Tusnady, G. The History of Ragweed in the World. Appl. Ecol. Environ. Res. 2015, 13, 489–512. [Google Scholar] [CrossRef]
- Loubet, I.; Caddoux, L.; Fontaine, S.; Michel, S.; Pernin, F.; Barrès, B.; Le Corre, V.; Délye, C. A high diversity of mechanisms endows ALS-inhibiting herbicide resistance in the invasive common ragweed (Ambrosia artemisiifolia L.). Sci. Rep. 2021, 11, 19904. [Google Scholar] [CrossRef] [PubMed]
- Voća, N.; Bilandžija, N.; Peter, A.; Krička, T. Ragweed (Ambrosia artemisiifolia L.) Biomass as a Source for Energy Production. Technol. Acta-Sci. J. Chem. Technol. 2020, 13, 17–22. [Google Scholar] [CrossRef]
- Janaćković, P.; Rajčević, N.; Gavrilović, M.; Novaković, J.; Radulović, M.; Miletić, M.; Janakiev, T.; Dimkić, I.; Marin, P.D. Essential Oil Composition of Ambrosia artemisiifolia and Its Antibacterial Activity against Phytopathogens. Biol. Life Sci. Forum 2022, 15, 22. [Google Scholar] [CrossRef]
- Hattori, S. Syntrophic Acetate-Oxidizing Microbes in Methanogenic Environments. Microbes Environ. 2008, 23, 118–127. [Google Scholar] [CrossRef]
- Worm, P.; Müller, N.; Plugge, C.M.; Stams, A.J.M.; Schink, B. Syntrophy in Methanogenic Degradation. In (Endo) Symbiotic Methanogenic Archaea; Springer: Berlin/Heidelberg, Germany, 2010; pp. 143–173. [Google Scholar] [CrossRef]
- Thapa, S.; Mishra, J.; Arora, N.; Mishra, P.; Li, H.; O′Hair, J.; Bhatti, S.; Zhou, S. Microbial cellulolytic enzymes: Diversity and biotechnology with reference to lignocellulosic biomass degradation. Rev. Environ. Sci. Bio/Technol. 2020, 19, 621–648. [Google Scholar] [CrossRef]
- Patel, S.K.; Das, D.; Kim, S.C.; Cho, B.-K.; Kalia, V.C.; Lee, J.-K. Integrating strategies for sustainable conversion of waste biomass into dark-fermentative hydrogen and value-added products. Renew. Sustain. Energy Rev. 2021, 150, 111491. [Google Scholar] [CrossRef]
- Ziemiński, K. Methane fermentation process as anaerobic digestion of biomass: Transformations, stages and microorganisms. Afr. J. Biotechnol. 2012, 11, 4127–4139. [Google Scholar] [CrossRef]
- Agoro, M.A.; Adeniji, A.O.; Adefisoye, M.A.; Okoh, O.O. Heavy Metals in Wastewater and Sewage Sludge from Selected Municipal Treatment Plants in Eastern Cape Province, South Africa. Water 2020, 12, 2746. [Google Scholar] [CrossRef]
- Fridh, L.; Volpé, S.; Eliasson, L. An accurate and fast method for moisture content determination. Int. J. For. Eng. 2014, 25, 222–228. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Acree, T.E.; Decker, E.A.; Penner, M.H.; Reid, D.S.; Schwartz, S.J.; Shoemaker, C.F.; Smith, D.M.; Sporns, P. Handbook of Food Analytical Chemistry, Volume 1: Water, Proteins, Enzymes, Lipids, and Carbohydrates; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 0-471-70909-3. Available online: https://www.wiley.com/en-us/Handbook+of+Food+Analytical+Chemistry,+Volume+1:+Water,+Proteins,+Enzymes,+Lipids,+and+Carbohydrates-p-9780471663782 (accessed on 12 November 2022).
- Pękal, A.; Pyrzynska, K. Evaluation of Aluminium Complexation Reaction for Flavonoid Content Assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Semenov, V.L.; Iarosh, A.M. A method of determining the antioxidative activity of biological matter. Ukr. Biokhimicheskii Zhurnal (1978) 1985, 57, 50–52. Available online: https://pubmed.ncbi.nlm.nih.gov/4024299/ (accessed on 13 November 2022).
- Liu, D.; Wong, P.; Dutka, B. Determination of carbohydrate in lake sediment by a modified phenol-sulfuric acid method. Water Res. 1973, 7, 741–746. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, Y.; Zhao, Y.; Zhu, X. New Sludge Pretreatment Method to Improve Methane Production in Waste Activated Sludge Digestion. Environ. Sci. Technol. 2010, 44, 4802–4808. [Google Scholar] [CrossRef]
- Harada, H.; Uemura, S.; Momonoi, K. Interaction between sulfate-reducing bacteria and methane-producing bacteria in UASB reactors fed with low strength wastes containing different levels of sulfate. Water Res. 1994, 28, 355–367. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Y.; Li, Y.; Cheng, Y.; Zhu, W. The enrichment of anaerobic fungi and methanogens showed higher lignocellulose degrading and methane producing ability than that of bacteria and methanogens. World J. Microbiol. Biotechnol. 2020, 36, 125. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ji, Q.; Fu, X.; Yu, X.; Ye, Z.; Zhang, M.; Sun, C.; Qiu, Y. Low-Cost Detection of Methane Gas in Rice Cultivation by Gas Chromatography-Flame Ionization Detector Based on Manual Injection and Split Pattern. Molecules 2022, 27, 3968. [Google Scholar] [CrossRef]
- Acree, W.E. Basic Gas Chromatography (McNair, Harold M.; Miller, James M.). J. Chem. Educ. 1998, 75, 1094. [Google Scholar] [CrossRef]
- Tashyrev, O.; Prekrasna, I. Express Method for Redox Potential and Ph Measuring in Microbial Cultures. Int. J. Bioautomation 2014, 18, 217–230. Available online: http://www.biomed.bas.bg/bioautomation/2014/vol_18.3/files/18.3_05.pdf (accessed on 15 December 2022).
- Zehnder, A.J.B.; Wuhrmann, K. Titanium (III) Citrate as a Nontoxic Oxidation-Reduction Buffering System for the Culture of Obligate Anaerobes Vertebrate Central Nervous System: Same Neurons Mediate Both Electrical and Chemical Inhibitions. Science 1975, 194, 1165–1166. [Google Scholar] [CrossRef]
- Suslova, O.; Govorukha, V.; Brovarskaya, O.; Matveeva, N.; Tashyreva, H.; Tashyrev, O. Method for Determining Organic Compound Concentration in Biological Systems by Permanganate Redox Titration. Int. J. Bioautomation 2014, 18, 45–52. Available online: https://biomed.bas.bg/bioautomation/2014/vol_18.1/files/18.1_05.pdf (accessed on 21 November 2022).
- Prekrasna, I.P.; Tashyrev, O.B. Copper resistant strain Candida tropicalis RomCu5 interaction with soluble and insoluble copper compounds. Biotechnol. Acta 2015, 8, 93–102. [Google Scholar] [CrossRef]
- Lace, A.; Ryan, D.; Bowkett, M.; Cleary, J. Chromium Monitoring in Water by Colorimetry Using Optimised 1,5-Diphenylcarbazide Method. Int. J. Environ. Res. Public Health 2019, 16, 1803. [Google Scholar] [CrossRef] [PubMed]
- Govorukha, V.M.; Havrylyuk, O.A.; Tashyrev, O.B. Regularities of quantitative distribution for Fe(III)-reducing bacteria in natural ecosystems. Biotechnol. Acta 2015, 8, 123–128. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, L.; Jonker, A.; Munidasa, S.; Pacheco, D. A Review: Plant Carbohydrate Types—The Potential Impact on Ruminant Methane Emissions. Front. Vet. Sci. 2022, 9, 880115. [Google Scholar] [CrossRef]
- Moliszewska, E.B.; Nabrdalik, M. Application and Biological Impact of Endophytic Bacteria as IAA Producers. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Elsevier: Amsterdam, The Netherlands, 2020; pp. 77–87. [Google Scholar] [CrossRef]
- Havryliuk, A.; Hovorukha, V.M.; Sachko, A.V.; Gladka, G.V.; Bida, I.O.; Tashyrev, O.B. Bioremoval of hazardous cobalt, nickel, chromium, copper and cadmium compounds from contaminated soil by Nicotiana tabacum plants and associated microbiome. Biosyst. Divers. 2021, 29, 88–93. [Google Scholar] [CrossRef]
- Nabrdalik, M.; Moliszewska, E.; Wierzba, S. Importance of Endophytic Strains Pantoea agglomerans in the Biological Control of Rhizoctonia solani. Ecol. Chem. Eng. S 2018, 25, 331–342. [Google Scholar] [CrossRef]
- Chołuj, D.; Moliszewska, E.B. The influence of Aphanomyces cochlioides on selected physiological processes in sugar beet leaves and yield parameters. Eur. J. Plant Pathol. 2012, 132, 59–70. [Google Scholar] [CrossRef]
- Tashyrev, O.; Hovorukha, V.; Havryliuk, O.; Sioma, I.; Gladka, G.; Kalinichenko, O.; Włodarczyk, P.; Suszanowicz, D.; Zhuk, H.; Ivanov, Y. Spatial Succession for Degradation of Solid Multicomponent Food Waste and Purification of Toxic Leachate with the Obtaining of Biohydrogen and Biomethane. Energies 2022, 15, 911. [Google Scholar] [CrossRef]
- Hovorukha, V.M.; Tashyrev, O.B.; Matvieieva, N.A.; Tashyreva, H.O.; Havryliuk, O.A.; Bielikova, O.I.; Sioma, I.B. Integrated Approach for Development of Environmental Biotechnologies for Treatment of Solid Organic Waste and Obtaining of Biohydrogen and Lignocellulosic Substrate. Environ. Res. Eng. Manag. 2019, 74, 31–42. [Google Scholar] [CrossRef]
- Havryliuk, O.; Hovorukha, V.; Patrauchan, M.; Youssef, N.H.; Tashyrev, O. Draft whole genome sequence for four highly copper resistant soil isolates Pseudomonas lactis strain UKR1, Pseudomonas panacis strain UKR2, and Pseudomonas veronii strains UKR3 and UKR4. Curr. Res. Microb. Sci. 2020, 1, 44–52. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Z.; Liu, X.; Zhang, X.-H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef]
- Maukonen, J.; Saarela, M. Microbial communities in industrial environment. Curr. Opin. Microbiol. 2009, 12, 238–243. [Google Scholar] [CrossRef]
- Sun, L.; Liu, T.; Müller, B.; Schnürer, A. The Microbial Community Structure in Industrial Biogas Plants Influences the Degradation Rate of Straw and Cellulose in Batch Tests. Biotechnol. Biofuels 2016, 9, 128. [Google Scholar] [CrossRef] [PubMed]
- Comeau, Y. Microbial Metabolism. In Biological Wastewater Treatment: Principles, Modelling and Design; IWA Publishing: London, UK, 2008; pp. 9–32. [Google Scholar]
- Lee, D.-J.; Show, K.-Y.; Su, A. Dark fermentation on biohydrogen production: Pure culture. Bioresour. Technol. 2011, 102, 8393–8402. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chui, C.; Zhang, S.; Liu, Q.; Li, B.; Shi, J.; Liu, L. Hydrogen Production by the Thermophilic Dry Anaerobic Co-Fermentation of Food Waste Utilizing Garden Waste or Kitchen Waste as Co-Substrate. Sustainability 2022, 14, 7367. [Google Scholar] [CrossRef]
- Koley, A.; Mukhopadhyay, P.; Show, B.K.; Ghosh, A.; Balachandran, S.; Santiniketan, V.-B. Biogas Production Potentiality of Water Hyacinth, Pistia and Duckweed: A Comparative Analysis. In Proceedings of the National Symposium: “Recent Trends in Sustainable Technology-Techno-Commercial Developments”, Kolkata, India, 9–10 September 2022. [Google Scholar]
- Havryliuk, O.; Hovorukha, V.; Savitsky, O.; Trilis, V.; Kalinichenko, A.; Dołhańczuk-Śródka, A.; Janecki, D.; Tashyrev, O. Anaerobic Degradation of Environmentally Hazardous Aquatic Plant Pistia stratiotes and Soluble Cu(II) Detoxification by Methanogenic Granular Microbial Preparation. Energies 2021, 14, 3849. [Google Scholar] [CrossRef]
- Dubrovskis, V.; Plume, I.; Straume, I. Suitability of Common Nettle (Urticadioica) and Canadian Goldenrod (Solidago canadensis) for Methane Production. Agron. Res. 2018, 16, 4. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Park, J.; Seong, S.; Oh, J.-W. Effects of Mechanical Drying on the Removal of Pollen Allergens. Int. Arch. Allergy Immunol. 2020, 181, 675–679. [Google Scholar] [CrossRef]
- Aggarwal, N.K.; Kumar, N.; Mittal, M. Potential of Weed Biomass for Bioethanol Production. In Bioethanol Production; Springer: Berlin/Heidelberg, Germany, 2022; pp. 65–71. [Google Scholar] [CrossRef]
- Nong, H.T.T.; Unpaprom, Y.; Chaichompoo, C.; Ramaraj, R. Biomethane potential of invasive aquatic weed water primrose. Glob. J. Sci. Eng. 2021, 5, 1–5. [Google Scholar] [CrossRef]
- Vidotto, F.; Tesio, F.; Ferrero, A. Allelopathic effects of Ambrosia artemisiifolia L. in the invasive process. Crop. Prot. 2013, 54, 161–167. [Google Scholar] [CrossRef]
- Lalthanpuii, P.; Lalchhandama, K. Imperata Cylindrica: A Noxious Weed of Pharmacological Potentials; Atlantis Press: Amsterdam, The Netherlands, 2018; pp. 173–177. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).