Abstract
Helicoverpa armigera (Hübner) is a key polyphagous pest with global spread that causes economic damage to agricultural crops. It is commonly managed by synthetic insecticides, which can cause negative impacts on the environment and human health, as well as toxicity to natural enemies. Hence, investigations into alternative pest management approaches such as the use of resistant host plant cultivars against H. armigera is important. In this study, the effects of various legume seeds based on meridic diets were evaluated using biology and population growth parameters of H. armigera under laboratory conditions. The results indicated that the shortest developmental time and pupal incubation periods occurred on white kidney bean Dehghan, while the longest periods occurred on broad bean Mahta. The intrinsic rate of population increase (r) was highest when H. armigera was reared on white kidney bean Dehghan and cowpea Arabi, and the lowest when reared on broad bean Mahta and common bean Khomein. Furthermore, the highest net reproductive rate (R0) occurred on white kidney bean Dehghan and the lowest on common bean Khomein. Cluster analysis revealed that white kidney bean Dehghan and cowpea Arabi were the most susceptible hosts, while common bean Khomein, and broad beans Feyz and Mahta were the poorest and perhaps least susceptible hosts for population growth of H. armigera. Our results suggest that the selective use of certain bean cultivars as potential sources of antibiosis against this pest may be useful as part of an integrated management program.
1. Introduction
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) is widely recognized as a destructive agricultural pest, known for its diverse host plant range and broad geographic distribution [1,2,3,4]. With over 200 species of plants being susceptible to attack, including economically significant crops such as tomato, cotton, maize, pigeon pea, chickpea, soybean, tobacco, and sorghum [5,6,7,8], H. armigera has garnered significant attention from the agricultural community.
Helicoverpa armigera life history characteristics facilitate adaptability to changing habitats and seasonal fluctuations [9,10,11]. These traits also explain the wide geographical distribution of the species in areas between 45° N and 45° S latitudes [10,12]. Female H. armigera can lay more than 4000 eggs in a single cycle, which can vary depending on host and environmental conditions [13]. The neonate larvae feed on reproductive and vegetative parts of the host plant such as flowers, buds, stems, and leaves, while older individuals prefer the reproductive parts of plants [14].
The nutritional value of host plants affects the development rate, survivorship, reproduction, and life table parameters of H. armigera, as well as its population dynamics [15,16,17,18]. Quantitative and qualitative nutritional changes in food sources can directly impact insect performance, altering developmental time, weight, and size [19,20,21]. Similarly, the quality of parental diets in Lepidoptera directly affects performance of future generations [22]. For example, plant protease inhibitors (PIs) disrupt the activity of digestive enzymes in insects, thereby decreasing the assimilation of foods and resulting in nutrient deficiencies, poor growth, and increased mortality in larvae [11,23]. Previous studies have revealed that certain plant seeds in particular produce antifeedant compounds and digestive enzyme inhibitors [24,25,26]. Furthermore, PI-producing plant cultivars appear to be promising candidates for pest management programs due to their tolerance traits [27,28].
Life tables provide a straightforward method for tracking herbivore population growth as influenced by host plant suitability [18], and provide key demographic parameters [29]. Demographic studies have a range of applications, including analysis of population stability and structure, estimating extinction probability, as well as predicting life history evolution, prevalence, and potential for invasiveness by pest species [30]. In the context of integrated pest management, life table analysis can help define cultivar susceptibility to pest species, cultivar selection, and crop breeding [31].
Despite the high level of natural mortality caused by abiotic factors, predation, disease, and plant defense, control of H. armigera is necessary using synthetic pesticides [32]. Over-dependence on insecticides for managing H. armigera has resulted in high levels of resistance to common organophosphate and pyrethroid insecticides used in field crops [33,34]. There is a need to explore alternative, cost-effective, and sustainable pest management strategies such as the use of host plant tolerance or resistance against H. armigera.
Jafari et al. (2023) evaluated the effects of various phytochemicals, including starches, proteins, flavonoids, and phenolics, in seeds from various legume varieties on the nutritional physiology and digestive function of H. armigera [8]. In the current report, we described a follow up investigation evaluating the effects of these legume varieties on the population growth parameters of H. armigera to establsish possible variations in their susceptability to this pest. The findings of this study may inform the selection of legume species with tolerance to H. armigera, which could be incorporated into integrated pest management strategies aimed at reducing the use of chemical insecticides.
2. Materials and Methods
2.1. Plant Seeds
Seeds of various legume species, including Broad Bean (Vicia faba L.) (cultivars Barekat, Mahta, Saraziri, Feyz, and Shadan), Bean (Phaseolus vulgaris L.) (white kidney bean Dehghan, common bean Khomein, red kidney bean Goli), and Cowpea (Vigna sinensis L.) (Mashhad and Arabi) were obtained from the Safiabad Agricultural and Natural Resources Research and Education Center, Dezful, Iran. The seeds were individually ground and stored at 4 °C for use in subsequent experiments. The artificial diet consisted of seed powder (250 g, each plant cultivar separately), wheat germ (30 g) as protein and carbohydrate sources, sorbic acid (1.1 g) as an antimicrobial agent, ascorbic acid (3.5 g) as a vitamin source, sunflower oil (5 mL) as a preservative, agar (14 g) as a moisturizer, methyl-p-hydroxy benzoate (2.2 g), formaldehyde 37% (2.5 g), and distilled water (650 mL). The artificial diets were prepared based on the method described by Shorey and Hale (1965) and Hemati et al. (2012), and were stored in the refrigerator for no less than two days prior to use [11,35].
2.2. Insect Rearing
Helicoverpa armigera larvae used in the experiment were collected from a tomato field in the Behbahan region (Khuzestan province) of southwestern Iran in early September 2020. The collected larvae were placed in plastic containers (diameter of 9 cm and depth of 1 cm) and fed various legume-based artificial diets and kept under controlled conditions (25 ± 1 °C, 60 ± 5% RH, and 16 h photoperiod). Adults that emerged were kept in plastic containers (15 cm diameter × 20 cm height) and provided wet cotton soaked in honey solution (10%). A second piece of cotton that was soaked in autoclaved water was provided in each container as an additional source of hydration. Adult H. armigera used in these experiments were derived from the third generation of laboratory-reared insects.
2.3. Experiments
Life table parameters of H. armigera were quantified using adult moths that emerged from larvae reared on various legume-based artificial diets. One hundred eggs laid by female moths within a 12 h period was used to quantify the survivorship and development of H. armigera on each of the tested legume seeds. The eggs were checked every 12 h, and the number of larvae that emerged was recorded. The neonate larvae were individually transferred into plastic plates (9 cm diameter and 1 cm depth) with a fine mesh net for aeration. Fresh artificial diet was provided as needed, and daily observations were made to record the mortality/survival of larvae as they molted to the next instar or until they emerged as adults. The head capsules or exuviae from molted larvae were used to identify larval instars. Fifth instar larvae were kept in small plastic containers (5 cm diameter, 8 cm depth) for pre-pupation and pupation, and daily records were taken for the duration of these stages and to record mortality. The gender of the pupae was determined and recorded.
After adult moths emerged, a pair of female and male moths were transferred to egg-laying containers (15 cm diameter and 20 cm height) with a fine mesh net for ventilation. A small cotton wick soaked in 10% honey solution was placed in the containers to provide a source of carbohydrates for the adult insects. The number of eggs deposited by each individual female was recorded daily, and the experiments were continued until the death of the last female in the cohort. The sex ratio of H. armigera on different host cultivars was determined by maintaining 250–300 eggs laid by females that emerged from larvae fed on each host cultivar until adult moths emerged.
The life table parameters of H. armigera on the legume cultivars were calculated according to Carey (2001), including the intrinsic rate of natural increase I, net reproductive rate (R0), gross reproductive rate (GRR), finite rate of increase (λ), mean generation time (T), and doubling time (DT) [36]. These estimates were based on Birch (1948) and Southwood and Henderson (2000) [37,38].
2.4. Data Analysis
The life table parameters of H. armigera reared on various legume cultivars were analyzed using the jackknife method [39]. The jackknife method is a technique that involves repeated recalculations of the estimator by leaving out each sample in turn [29]. The jackknife pseudo values for the host were tested for normality and then subjected to an analysis of variance (ANOVA) using the PROC GLM procedure at SAS Institute Inc. Based on the results, mean comparisons were conducted using Tukey’s test with a significance level of p < 0.01 (SAS version 9.4, Cary, NC, USA). Additionally, a dendrogram was generated for the various legumes based on the life table parameters of H. armigera on various legume cultivars using Ward’s method in SPSS 22.0 statistical software (Chicago, IL, USA).
3. Results and Discussion
3.1. Development of Immature Stages, Adult Longevity, and Lifespan
The mean durations of immature stages, adult longevity, and lifespan of H. armigera reared on diets based on legumes are given in Table 1. Legume cultivar had a significant main effect on the development of all immature stages (F = 93.79; df = 9, 590; p < 0.01); the longest (3.68 ± 0.031 days) and shortest (2.76 ± 0.033 days) durations of incubation occurred for broad bean Mahta and white kidney bean Dehghan, respectively. Larvae reared on the common bean Khomein and broad bean Mahta exhibited longer larval periods (F = 94.320; df = 9, 429; p < 0.01) (21.61 ± 0.218 and 21.5 ± 0.224 days, respectively) than those fed the other hosts, while the shortest occurred for white kidney bean Dehghan (16.160 ± 0.149 days). Pupal periods (F = 36.49; df = 9, 372; p < 0.01) and developmental time (F = 182.81; df = 9, 372; p < 0.01) were the longest for broad bean Mahta (16.48 ± 0.131and 44.82 ± 0.244 days, respectively). However, the shortest pupal periods were for white kidney bean Dehghan (14.31 ± 0.126) and cowpea Mashhad (14.38 ± 0.132 days). The adult longevity (from adult emergence to death) and entire life span (from egg to death of adult) of H. armigera (female and male) are shown in Table 1. Both of these parameters were significantly affected by legume cultivar incorporated into the diet (F = 30.28; df = 9, 201; p < 0.01). The longest female longevity was observed for cowpea Arabi and white kidney bean Dehghan (23.60 ± 0.507 and 23.32 ± 0.519 days, respectively), while the shortest occurred for broad bean Mahta (18.11 ± 0.361 days). Females had the longest total life span (F = 11.41; df = 9, 200; p < 0.01) when they were reared on broad bean Shadan (64.28 ± 0.551 days) and the shortest for cowpea Mashhad and red kidney bean Goli (58.18 ± 0.558 and 58.35 ± 0.787 days, respectively). The longevity of males (F = 20.67; df = 9, 162; p < 0.01) was the longest (22.3 ± 0.529 days) and shortest (15.63 ± 0.328 days) for cowpea Mashhad and common bean Khomein, respectively. The male lifespan (F = 13.69; df = 8, 162; p < 0.01) was the longest when larvae were fed broad bean Feyz (64.17 ± 0.591 days) and shortest when larvae were reared on white kidney bean Dehghan (56.6 ± 0.559 days).

Table 1.
Mean (±SE) duration of developmental stages and adult longevity of Helicoverpa armigera on various legume cultivars.
Plant traits associated with tolerance or resistance can negatively impact the development and growth of H. armigera and its demographic characteristics [40,41]. Our results indicate that H. armigera was able to complete its life cycle, survive, and reproduce on meridic diets based on all of the legume varieties tested. However, the emergence period of H. armigera varied significantly depending on the legume seeds, which affected various other biological parameters. In this study, the broad bean Mahta appeared to exhibit the greatest tolerance against H. armigera, which significantly extended the duration of the immature period. Given that immature stages (particularly larvae) of H. armigera are most susceptible to natural enemies and pesticides [42], extending the development time of larvae fed broad bean Mahta should increase their exposure to these causes of mortality [43].
Previous studies have reported that the developmental time of H. armigera can be significantly affected by host plant quality [5,31,44]. For instance, Naseri et al. (2009) reported that the immature period of H. armigera ranged between 34.21 to 42.71 days for various soybean varieties [44], while Karimi et al. (2012) found that the developmental period ranged between 35.76 to 44.82 days on various canola cultivars [45]. These findings are somewhat different from the values obtained in the present study. These differences may be attributed to genetic variations among the populations evaluated, or differences in rearing conditions and the host species or varieties evaluated.
Fallahnejad-Mojarrad et al. (2017) investigated the biological parameters of H. armigera on semi-artificial diets based on leguminose varieties and found that the pre-adult period ranged from 22.71 to 28.94 days for cowpea and canola, respectively [46]. Similarly, Fathipour et al. (2020) reported the immature period of H. armigera for five known host species, including chickpea, soybean, corn, navy bean, and cowpea, and these ranged from 34.98 to 42.95 days. Faster development and higher reproductive output of insects are good indicators of host plant suitability and quality as a food resource [41]. However, these data must be linked to other parameters to verify host suitability [5].
The present study was consistent with previous investigations in that the pre-adult period of H. armigera was affected by host plant cultivar and varied among 10 various legumes. Significant variations in starch, protein, flavonoid, and total phenolic content were quantified between seeds of various legume species and there were significant correlations between the digestive enzyme activity of H. armigera and the occurrence of protease inhibitors in these legume cultivars [8]. In this case, phytochemical metabolites were considered as likely factors influencing the quantity and quality of the host plants consumed by the larvae, as reflected in the life history of the pest.
3.2. Survival Rate
The highest and lowest survival rates of larval stages were observed on cowpea Arabi and common bean Khomein, respectively (Figure 1). The overall survival rate recorded during the immature stage was higher for cowpea Arabi and common bean Khomein than the other cultivars examined. Survival rates of individuals that developed into adults from the initial cohort stage were estimated at 0.31, 0.21, 0.34, 0.27, 0.24, 0.50, 0.17, 0.31, 0.54, and 0.39 for the broad bean Barekat, Mahta, Saraziri, Feyz and Shadan, white kidney bean Dehghan, common bean Khomein, red kidney bean Goli, cowpea Arabi, and cowpea Mashhad cultivars, respectively.
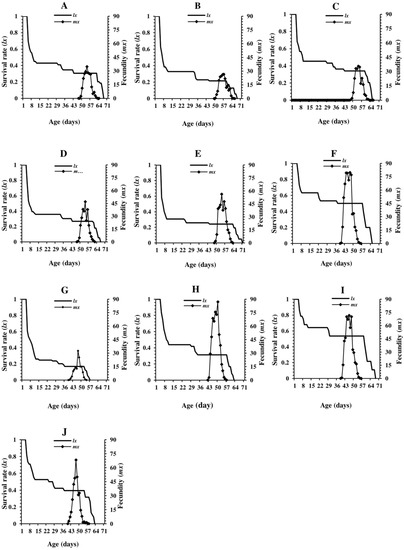
Figure 1.
Age-specific survival rate (lx) and fecundity (mx) of Helicoverpa armigera on various legume cultivars. (A) Broad bean Barekat, (B) Broad bean Mahta, (C) Broad bean Saraziri, (D) Broad bean Feyz, (E) Broad bean Shadan, (F) White kidney bean Dehghan, (G) Common bean Khomein, (H) Red kidney bean Goli, (I) Cowpea Arabi, and (J) Cowpea Mashhad.
Oviposition by females was initiated 40–50 days after rearing began, depending on the cultivar, which corresponded to approximately 3–4 days after adult eclosion. The age-specific fecundity (mx) of H. armigera adults emerging from diets composed of various legume cultivars ranged between 27–82 females/female/day and occurred 7–10 days after eclosion. The maximum age females reached when reared on the various cultivars ranged between 62–70 days, depending on the bean cultivar (Figure 1).
3.3. Oviposition Period and Fecundity
The adult pre-oviposition period (APOP), total pre-oviposition period (TPOP), oviposition period, and fecundity of H. armigera adults emerging from the larvae reared on artificial diets based on seeds of different legume cultivars are given in Table 2. Legume cultivar significantly affected the APOP (F = 16.77; df = 9, 140; p < 0.01), which was the longest for broad bean Mahta (4.60 ± 0.363 days) and the shortest for cowpea Mashhad (2.40 ± 0.131 days). The longest TPOP (F = 82.59; df = 9, 140; p < 0.01) was for broad bean Mahta (49.1 ± 0.47 days) and the shortest was for white kidney bean Dehghan and cowpea Arabi (37.6 ± 0.277 and 39.23 ± 0.358 days, respectively). The oviposition period (F = 4.81; df = 9, 140; p < 0.01) was the shortest on broad bean Mahta (10.27 ± 0.581 days).
Table 2.
Mean ± SE duration of pre-oviposition and oviposition periods and fecundity of Helicoverpa armigera on various legume cultivars.
There was a significant difference in both the daily and total number of eggs laid per individual fed H. armigera reared on diets containing the various legume cultivars. Both the daily (F = 34.95; df = 9, 140; p < 0.01) and total (F = 55.30; df = 9, 140; p < 0.01) fecundity rates of H. armigera were lowest when the larvae reared on common bean Khomein (49 and 585 eggs, respectively). The highest daily fecundity was for red kidney bean Goli (106 eggs), and the total fecundity was the highest for white kidney bean Dehghan (1292 eggs).
Fecundity of adult Lepidapera is often related to larval diets [31,47]. Our results with H. armigera similarly indicated an effect of larval diet on the subsequent fecundity of adult females, which may be related to differences in the protein content of the diets required for egg production. The highest fecundity occurred fed a diet containing white kidney bean Dehghan, which is a highly proteinaceous resource [8]. In general, the fecundity of H. armigera appeared to be greater for diets characterized by higher amounts of primary metabolites, including proteins and starch [8].
3.4. Population Growth Parameters
The population parameters calculated using the age-stage, female-based life tables for H. armigera on different legume cultivars are listed in Table 3. The highest and lowest net reproductive rate (R0) and gross reproductive rate (GRR) of H. armigera occurred for white kidney bean Dehghan (310 and 6210 females) and common bean Khomein (26 and 155 females), respectively (Table 3). Furthermore, the intrinsic rate of natural increase (r) was highest for white kidney bean Dehghan (0.124) and cowpea Arabi (0.123), and the lowest for broad bean Mahta (0.066). In addition, both the mean generation time (T) and doubling time (DT) were the shortest on white kidney bean Dehghan (46 and 5.59 days) and cowpea Arabi (46 and 5.65 days), and the longest on the broad bean Mahta (56.5 and 10.46 days).
Table 3.
Mean (±SE) population growth parameters of Helicoverpa armigera on various legume cultivars.
In the present study, the intrinsic rate of increase (r) values for H. armigera ranged from 0.069 to 0.123 per day on the various legume seed diets, which was lower than that reported in previous studies (0.126 and 0.299 per day) [5,31,45,46]. Differences between our results and previous studies may be attributed to differences in the experimental conditions or host varieties used for rearing H. armigera.
The highest rates of intrinsic population increase for H. armigera were observed for white kidney bean Dehghan and cowpea Arabi seeds, suggesting that these cultivars are suitable hosts of this pest. Jafari et al. (2023) indicated that white kidney bean Dehghan and cowpea Arabi seeds were characterized by the lowest amounts of phytochemical metabolites, including total phenolic and flavonoid contents, among the various legume varieties examined [8]. It is possible that the greater suitability of these two cultivars for H. armigera development is related to their lower phytochemical defense.
3.5. Cluster Analysis
Different legume cultivars were grouped within clusters according to the demographic parameters of H. armigera reared on diets based on the 10 legume cultivars (Figure 2). Cluster A included subclusters A1 (broad beans Mahta and Feyz, and common bean Khomein) as the most resistant cultivars and A2 (broad bean Barekat, Saraziri, and Shadan and cowpea Mashhad) as relatively unsuitable hosts. In addition, cluster B consisted of subclusters B1 (white kidney bean Dehghan and cowpea Arabi) grouped as the most susceptible cultivars and B2 (red kidney bean Goli) as a relatively suitable host for H. armigera.
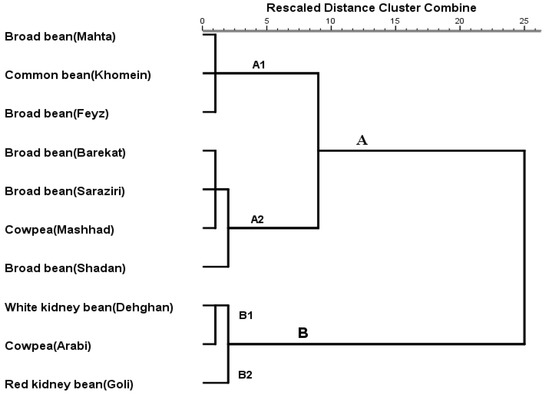
Figure 2.
Dendrogram of different host plants according to life table parameters of Helicoverpa armigera on various legume cultivars (Ward’s method).
The cluster analysis indicates groups of cultivars based on their similarities in suitability as food resources to H. armigera and may reflect similarity in physiological or plant defense characteristics [21,48]. In particular, those cultivars in subcluster A1, including broad beans Mahta and Feyz, and common bean Khomein, appear most tolerant to H. armigera feeding, and may be a useful as selections to diminish damage by this pest.
4. Conclusions
Our results indicate that the host plant component of the meridic diets evaluated was a significant factor affecting key biological growth parameters of the H. armigera. Specifically, insects reared on white kidney bean (Dehghan) and cowpea (Arabi) displayed the shortest mean generation time, and the highest finite rate of increase, and intrinsic rate of increase. Additionally, our findings confirmed that the growth and development of H. armigera was poorest for broad beans (Mahta and Feyz) and common bean (Khomein), suggesting that these cultivars were the most resistant among those evaluated. These findings suggest that Mahta and Feyz broad beans could be useful selections for cultivation in areas where damage by H. armigera is typically high; however, this hypothesis will require further testing with intact plants under field conditions. In addition, future investigations to identify the plant compound(s) from the bean seeds investigated here that underly delayed growth and development of H. armigera could yield useful feeding or digestibility inhibitors. The identification of these inhibitory compounds could open opportunities to generate transgenic pest-resistant plants as a sustainable pest management strategy. Quantifying phytochemical differences between the cultivars investigated here is a logical next step. For example, protease inhibitors are known to reduce population growth parameters in H. armigera and should be explored in future research.
Author Contributions
Conceptualization, S.A.H. and B.H.; methodology, S.A.H. and H.J.; software, S.A.H. and H.J.; validation, S.A.H. and B.H.; formal analysis, S.A.H. and H.J.; investigation, S.A.H., H.J. and L.L.S.; resources, S.A.H., B.H. and L.L.S.; data curation, S.A.H., B.H., H.J. and L.L.S.; writing—original draft preparation, H.J. and S.A.H.; writing—review and editing, L.L.S.; visualization, S.A.H., H.J. and L.L.S.; supervision, S.A.H. and B.H.; project administration, S.A.H. and B.H.; funding acquisition, S.A.H. and B.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Shahid Chamran University of Ahvaz, Ahvaz, Iran (Grant No. SCU.AP99.39134).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated in this study are available from the corresponding author upon reasonable request.
Acknowledgments
This research was financially supported by Shahid Chamran University of Ahvaz, Ahvaz, Iran, which is greatly appreciated.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Zalucki, M.P.; Furlong, M.J. Forecasting Helicoverpa populations in Australia: A comparison of regression-based models and a bioclimatic based modelling approach. Insect Sci. 2005, 12, 45–56. [Google Scholar] [CrossRef]
- Silva, I.F.; Baldin, E.L.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Morando, R.; Paula-Moraes, S.V. Biotic potential and life table of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) from three Brazilian regions. Neotrop. Entomol. 2018, 47, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Fathipour, Y.; Babaei, M.; Bagheri, A.; Talebi, A.A.; Yazdanpanah, S. Demographic parameters of Helicoverpa armigera on ten corn hybrids-mediated artificial diets reveals striking differences. J. Crop Prot. 2021, 10, 363–374. [Google Scholar]
- Atashi, N.; Shishehbor, P.; Seraj, A.A.; Rasekh, A.; Hemmati, S.A.; Riddick, E.W. Effects of Helicoverpa armigera Egg Age on Development, Reproduction, and Life Table Parameters of Trichogramma euproctidis. Insects 2021, 12, 569. [Google Scholar] [CrossRef]
- Liu, Z.; Li, D.; Gong, P.; Wu, K. Life table studies of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different host plants. Environ. Entomol. 2004, 33, 1570–1576. [Google Scholar] [CrossRef]
- Talekar, N.S.; Opena, R.T.; Hanson, P. Helicoverpa armigera management: A review of AVRDC’s research on host plant resistance in tomato. Crop Prot. 2006, 25, 461–467. [Google Scholar] [CrossRef]
- Hemati, S.A.; Naseri, B.; Nouri Ganbalani, G.; Rafiee Dastjerdi, H.; Golizadeh, A. Effect of different host plants on nutritional indices of the pod borer, Helicoverpa armigera. J. Insect Sci. 2012, 12, 55. [Google Scholar] [CrossRef]
- Jafari, H.; Hemmati, S.A.; Habibpour, B. Evaluation of artificial diets based on different legume seeds on the nutritional physiology and digestive function of Helicoverpa armigera (Hübner). Bull. Entomol. Res. 2023, 113, 133–143. [Google Scholar] [CrossRef]
- Karim, S. Management of Helicoverpa armigera: A review and prospectus for Pakistan. Pak. J. Biol. Sci. 2000, 3, 1213–1222. [Google Scholar] [CrossRef]
- Fitt, G.P. The ecology of Heliothis species in relation to agroecosystems. Annu. Rev. Entomol. 1989, 34, 17–53. [Google Scholar] [CrossRef]
- Hemati, S.A.; Naseri, B.; Ganbalani, G.N.; Dastjerdi, H.R.; Golizadeh, A. Digestive proteolytic and amylolytic activities and feeding responses of Helicoverpa armigera (Lepidoptera: Noctuidae) on different host plants. J. Econ. Entomol. 2012, 105, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Mironidis, G.K.; Savopoulou-Soultani, M. Development, survivorship, and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) under constant and alternating temperatures. Environ. Entomol. 2014, 37, 16–28. [Google Scholar] [CrossRef]
- Hardwick, D.F. The corn earworm complex. Mem. Ent. Soc. Can. 1965, 97, 5–247. [Google Scholar] [CrossRef]
- Garcia, F.M. Analysis of the Spatio–temporal Distribution of Helicoverpa armigera Hb. In a Tomato Field using a Stochastic Approach. Biosyst. Eng. 2006, 93, 253–259. [Google Scholar] [CrossRef]
- Yongming, R.; Kunjun, W. Performances of the cotton boll worm, Helicoverpa armigera on different food plants. Kun chong xue bao. Acta Entomol. Sin. 2001, 44, 205–212. [Google Scholar]
- Kim, D.S.; Lee, J.H. Egg and larval survivorship of Carposina sasakii (Lepidoptera: Carposinidae) in apple and peach and their effects on adult population dynamics in orchards. Environ. Entomol. 2002, 31, 686–692. [Google Scholar] [CrossRef]
- Da Silva, I.F.; Baldin, E.L.L.; Specht, A.; Roque-Specht, V.F.; Morando, R.; Malaquias, J.V.; Paula-Moraes, S.V. Role of nutritional composition in the development and survival of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on artificial diet and natural hosts. Bull. Entomol. Res. 2021, 111, 257–269. [Google Scholar] [CrossRef]
- Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Life Table Parameters and Digestive Enzyme Activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on Selected Legume Cultivars. Insects 2022, 13, 661. [Google Scholar] [CrossRef]
- Yamasaki, A.; Fujisaki, K. Larval feeding preference and performance of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different flower parts of cosmos. Appl. Entomol. Zool. 2010, 45, 627–633. [Google Scholar] [CrossRef]
- Babamir-Satehi, A.; Habibpour, B.; Aghdam, H.R.; Hemmati, S.A. Interaction between feeding efficiency and digestive physiology of the pink stem borer, Sesamia cretica Lederer (Lepidoptera: Noctuidae), and biochemical compounds of different sugarcane cultivars. Arthropod Plant Interact. 2022, 16, 309–316. [Google Scholar] [CrossRef]
- Zamani Fard, S.; Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Growth, consumption and digestive enzyme activities of Spodoptera littoralis (Boisd) on various mung bean cultivars reveal potential tolerance traits. J. Appl. Entomol. 2022, 146, 1145–1154. [Google Scholar] [CrossRef]
- Woestmann, L.; Saastamoinen, M. The importance of trans-generational effects in Lepidoptera. Curr. Zool. 2016, 62, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.; Dunse, K.; Fox, J.; Evans, S.; Anderson, M. Biotechnological approaches for the control of insect pests in crop plants. In Pesticides-Advances in Chemical and Botanical Pesticides; IntechOpen: London, UK, 2012; pp. 269–308. [Google Scholar]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Grossi-de-Sá, M.F. Plant α-amylase inhibitors and their interaction with insect α-amylases: Structure, function and potential for crop protection. Eur. J. Biochem. 2002, 269, 397–412. [Google Scholar] [CrossRef]
- Gomes, C.E.; Barbosa, A.E.; Macedo, L.L.; Pitanga, J.C.; Moura, F.T.; Oliveira, A.S.; Moura, R.M.; Queiroz, A.F.; Macedo, F.P.; Andrade, L.B.; et al. Effect of trypsin inhibitor from Crotalaria pallida seeds on Callosobruchus maculatus (cowpea weevil) and Ceratitis capitata (fruit fly). Plant Physiol. Biochem. 2005, 43, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.P.; Casado-Filho, E.L.; Corrêa, A.S.; Farias, L.R.; Bloch, C.; Grossi de Sá, M.F.; Mendes, P.A.; Quirino, B.F.; Noronha, E.F.; Franco, O.L. Identification of an α-amylase inhibitor from Pterodon pubescens with ability to inhibit cowpea weevil digestive enzymes. J. Agric. Food Chem. 2007, 55, 4382–4387. [Google Scholar] [CrossRef]
- Gatehouse, A.M.; Gatehouse, J.A. Identifying proteins with insecticidal activity: Use of encoding genes to produce insect-resistant transgenic crops. Pestic. Sci. 1998, 52, 165–175. [Google Scholar] [CrossRef]
- Mehrabadi, M.; Franco, O.L.; Bandani, A.R. Plant proteinaceous alpha-amylase and proteinase inhibitors and their use in insect pest control. In New Perspectives in Plant Protection; IntechOpen: Rijeka, Croatia, 2012; pp. 229–246. [Google Scholar]
- Maia, A.D.H.; Luiz, A.J.; Campanhola, C. Statistical inference on associated fertility life table parameters using jackknife technique: Computational aspects. J. Econ. Entomol. 2000, 93, 511–518. [Google Scholar] [CrossRef]
- Vargas, R.I.; Walsh, W.A.; Kanehisa, D.; Jang, E.B.; Armstrong, J.W. Demography of four Hawaiian fruit flies (Diptera: Tephritidae) reared at five constant temperatures. Ann. Entomol. Soc. Am. 1997, 90, 162–168. [Google Scholar] [CrossRef]
- Razmjou, J.; Naseri, B.; Hemati, S.A. Comparative performance of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on various host plants. J. Pest Sci. 2014, 87, 29–37. [Google Scholar] [CrossRef]
- Fitt, G.P. Cotton pest management. Part 3. An Australian perspective. Annu. Rev. Entomol. 1994, 39, 543–562. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I.; Attique, M.R. Pyrethroid resistance of Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. Bull. Entomol. Res. 1997, 87, 343–347. [Google Scholar] [CrossRef]
- Kranthi, K.R.; Jadhav, D.R.; Kranthi, S.; Wanjari, R.R.; Ali, S.S.; Russell, D.A. Insecticide resistance in five major insect pests of cotton in India. Crop Prot. 2002, 21, 449–460. [Google Scholar] [CrossRef]
- Shorey, H.H.; Hale, R.L. Mass-rearing of the larvae of nine noctuid species on a simple artificial medium. J. Econ. Entomol. 1965, 58, 522–524. [Google Scholar] [CrossRef]
- Carey, J.R. Insect biodemography. Annu. Rev. Entomol. 2001, 46, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Birch, L. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Henderson, P.A. Absolute population estimates by sampling a unit of soil or litter habitat: Extraction techniques. In Ecological Methods, 3rd ed.; Blackwell Science: Malden, MA, USA, 2000; pp. 213–241. [Google Scholar]
- Meyer, J.S.; Ingersoll, C.G.; McDonald, L.L.; Boyce, M.S. Estimating uncertainty in population growth rates: Jackknife vs. bootstrap techniques. Ecology 1986, 67, 1156–1166. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.V.; Noldus, L.P.J.J. Whitefly-plant relationships: Behavioural and ecological aspects. In Whiteflies: Their Bionomics, Pest Status and Management; Intercept: Andover, UK, 1990; Volume 47, p. 49. [Google Scholar]
- Fathipour, Y.; Baghery, F.; Bagheri, A.; Naseri, B. Development, reproduction and life table parameters of Helicoverpa armigera (Lepidoptera: Noctuidae) on five main host plants. J. Crop Prot. 2020, 9, 551–561. [Google Scholar]
- Alami, S.; Naseri, B.; Golizadeh, A.; Razmjou, J. Age-stage, two-sex life table of the tomato looper, Chrysodeixis chalcites (Lepidoptera: Noctuidae), on different bean varieties. Arthropod-Plant Inter. 2014, 8, 475–484. [Google Scholar] [CrossRef]
- Du, L.; Ge, F.; Zhu, S.; Parajulee, M.N. Effect of cotton variety on development and reproduction of Aphis gossypii (Homoptera: Aphididae) and its predator Propylaea japonica (Coleoptera: Coccinellidae). J. Econ. Entomol. 2004, 97, 1278–1283. [Google Scholar] [CrossRef]
- Naseri, B.; Fathipour, Y.; Moharramipour, S.; Hosseininaveh, V. Life table parameters of the cotton bollworm, Helicoverpa armigera (Lep: Noctuidae) on different soybean cultivars. J. Entomol. Soc. Iran 2009, 29, 25–40. [Google Scholar]
- Karimi, S.; Fathipour, Y.; Talebi, A.A.; Naseri, B. Evaluation of canola cultivars for resistance to Helicoverpa armigera (Lepidoptera: Noctuidae) using demographic parameters. J. Econ. Entomol. 2012, 105, 2172–2179. [Google Scholar] [CrossRef] [PubMed]
- Fallahnejad-Mojarrad, N.; Goldasteh, S.; Rafiei-Karahroodi, Z.; Vafaei Shoushtari, R. Response of the Cotton Bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae) to Different Semi-Artificial Diets. J. Agric. Sci. Technol. 2017, 19, 1303–1318. [Google Scholar]
- Kumral, N.A.; Kovanci, B.; Akbudak, B. Life tables of the olive leaf moth, Palpita unionalis (Hübner) (Lepidoptera: Pyralidae), on different host plants. J. Biol. Environ. Sci. 2007, 1, 105–110. [Google Scholar]
- Shishehbor, P.; Hemmati, S.A. Investigation of secondary metabolites in bean cultivars and their impact on the nutritional performance of Spodoptera littoralis (Lep.: Noctuidae). Bull. Entomol. Res. 2022, 112, 378–388. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).