
Biodiversity Characteristics and Carbon Sequestration Potential of Successional Woody Plants versus Tree Plantation under Different Reclamation Treatments on Hard-Coal Mine Heaps––A Case Study from Upper Silesia


 ,
,  ,
,  , , , ,
, , , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Analysis
2.3. Biomass Study
2.4. Plant Diversity
2.5. Statistical Analysis
3. Results
3.1. Vegetation Biomass
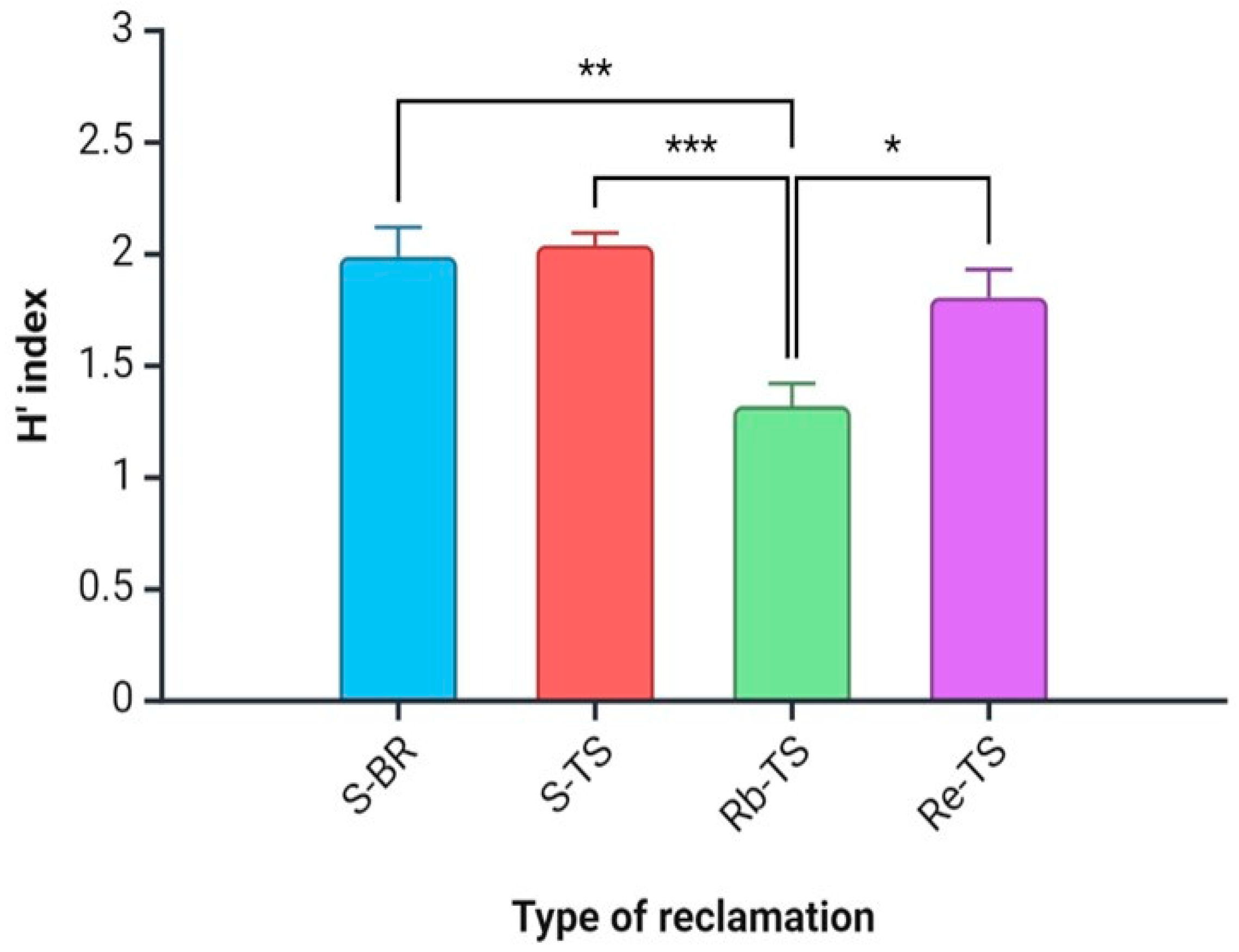
3.2. Plant Biodiversity
3.3. Carbon Storage in Ecosystems
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skousen, J.; Zipper, C.E. Post-mining policies and practices in the Eastern USA coal region. Int. J. Coal Sci. Technol. 2014, 1, 135–151. [Google Scholar] [CrossRef]
- Bell, F.G.; Donnelly, L.J. Mining and Its Impact on the Environment, 1st ed.; CRC Press: London, UK, 2006. [Google Scholar]
- Pietrzykowski, M.; Daniels, W.L. Estimation of carbon sequestration by pine (Pinus sylvestris L.) ecosystems developed on reforested post-mining sites in Poland on differing mine soil substrates. Ecol. Eng. 2014, 73, 209–218. [Google Scholar] [CrossRef]
- Burghardt, W.; Niggemeyer, M.; Braunersreuther, M. Ways of soil development on stony substrate from hard coal mining spoil. Soil Sci. Ann. 2020, 71, 382–394. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Błońska, A.; Kompała-Bąba, A.; Woźniak, G. Effects of Calamagrostis epigejos, Chamaenerion palustre and Tussilago farfara on nutrient availability and microbial activity in the surface layer of spoil heaps after hard coal mining. Ecol. Eng. 2015, 83, 328–337. [Google Scholar] [CrossRef]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Soil Reclamation of Abandoned Mine Land by Revegetation: A Review. Int. J. Soil Sediment Water 2010, 3, 13. [Google Scholar]
- Kumari, S.; Maiti, S.K. Reclamation of coalmine spoils with topsoil, grass, and legume: A case study from India. Environ. Earth Sci. 2019, 78, 429. [Google Scholar] [CrossRef]
- Kumari, S.; Maiti, S.K. Nitrogen recovery in reclaimed mine soil under different amendment practices in tandem with legume and non-legume revegetation: A review. Soil Use Manag. 2022, 38, 1113–1145. [Google Scholar] [CrossRef]
- Ghose, M.K. Management of topsoil for geo-environmental reclamation of coal mining areas. Environ. Geol. 2001, 40, 1405–1410. [Google Scholar]
- Prach, K.; Hobbs, R.J. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Baasch, A.; Tischew, S.; Bruelheide, H. Twelve years of succession on sandy substrates in a post-mining landscape: A Markov chain analysis. Ecol. Appl. 2010, 20, 1136–1147. [Google Scholar] [CrossRef]
- Prach, K.; Walker, L.R. Four opportunities for studies of ecological succession. Trends Ecol. Evol. 2011, 26, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Peng, S.; Lines, L.R.; Zhu, G.; Hu, Z.; Cui, F. Understanding the capability of an ecosystem nature-restoration in coal mined area. Sci. Rep. 2019, 9, 19690. [Google Scholar] [CrossRef]
- Vachova, P.; Vach, M.; Skalicky, M.; Walmsley, A.; Berka, M.; Kraus, K.; Hnilickova, H.; Vinduskova, O.; Mudrak, O. Reclaimed Mine Sites: Forests and Plant Diversity. Diversity 2022, 14, 13. [Google Scholar] [CrossRef]
- Prach, K.; Tolvanen, A. How can we restore biodiversity and ecosystem services in mining and industrial sites? Environ. Sci. Pollut. Res. 2016, 23, 13587–13590. [Google Scholar] [CrossRef]
- Woźniak, G.; Chmura, D.; Dyderski, M.K.; Błońska, A.; Jagodziński, A.M. How different is the forest on post-coal mine heap regarded as novel ecosystem? For. Ecol. Manag. 2022, 515, 120205. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Seabloom, E.W.; Borer, E.T.; Adler, P.B.; Harpole, W.S.; Hautier, Y.; Hillebrand, H.; Lind, E.M.; Pärtel, M.; et al. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 2016, 529, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Kompała-Bąba, A.; Sierka, E.; Bierza, W.; Błońska, A.; Woźniak, G. Eco-physiological responses of Calamagrostis epigejos L. (Roth) and Solidago gigantea Aition to complex environmental stresses in coal-mine spoil heaps. Land Degrad. Dev. 2021, 32, 5427–5442. [Google Scholar] [CrossRef]
- Sierka, E.; Radosz, Ł.; Ryś, K.; Woźniak, G. Ecosystem Services and Post-industrial Areas. In Green Scenarios: Mining Industry Responses to Environmental Challenges of the Anthropocene Epoch, 1st ed.; Dyczko, A., Jagodziński, A., Woźniak, G., Eds.; Taylor & Francis Group: London, UK, 2022; pp. 265–274. [Google Scholar]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Misebo, A.M.; Pietrzykowski, M.; Woś, B. Soil carbon sequestration in novel ecosystems at post-mine sites—A new insight into the determination of key factors in the restoration of terrestrial ecosystems. Forests 2022, 13, 63. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, J.; Hu, P.; Zhang, W.; Xiao, L.; Ye, Y.; Xiao, D.; Zhao, J.; Xiao, J.; Wang, K. Bedrock outcrops weakly promote rather than inhibit soil carbon sequestration after vegetation restoration. Sci. Total Environ. 2023, 858, 159470. [Google Scholar] [CrossRef]
- Dong, L.; Fan, J.; Li, J.; Zhang, Y.; Liu, Y.; Wu, J.; Li, A.; Shangguan, Z.; Deng, L. Forests have a higher soil C sequestration benefit due to lower C mineralization efficiency: Evidence from the central loess plateau case. Agric. Ecosyst. Environ. 2022, 339, 108144. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D.; Rich, R.; Pastore, M.A.; Worm, K. Synergistic effects of four climate change drivers on terrestrial carbon cycling. Nat. Geosci. 2020, 13, 787–793. [Google Scholar] [CrossRef]
- Espigares, T.; Moreno-de las Heras, M.; Nicolau, J.M. Performance of Vegetation in Reclaimed Slopes Affected by Soil Erosion. Restor. Ecol. 2011, 19, 35–44. [Google Scholar] [CrossRef]
- Mudrák, O.; Frouz, J.; Velichová, V. Understory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng. 2010, 36, 783–790. [Google Scholar] [CrossRef]
- Šebelíková, L.; Řehounková, K.; Prach, K. Spontaneous revegetation vs. forestry reclamation in post-mining sand pits. Environ. Sci. Pollut. Res. 2016, 23, 13598–13605. [Google Scholar] [CrossRef] [PubMed]
- Pietrzykowski, M.; Krzaklewski, W. Soil organic matter, C and N accumulation during natural succession and reclamation in an opencast sand quarry (southern Poland). Arch. Agron. Soil Sci. 2007, 53, 473–483. [Google Scholar] [CrossRef]
- Frouz, J.; Pižl, V.; Cienciala, E.; Kalčík, J. Carbon storage in post-mining forest soil, the role of tree biomass and soil bioturbation. Biogeochemistry 2009, 94, 111–121. [Google Scholar] [CrossRef]
- Abakumov, E.V.; Cajthaml, T.; Brus, J.; Frouz, J. Humus accumulation, humification, and humic acid composition in soils of two post-mining chronosequences after coal mining. J. Soils Sediments 2013, 13, 491–500. [Google Scholar] [CrossRef]
- Skousen, J.; Johnson, C.; Garbutt, K. Natural revegetation of 15 abandoned mine land sites in West Virginia. J. Environ. Qual. 1994, 23, 1224–1230. [Google Scholar] [CrossRef]
- Vacek, Z.; Cukor, J.; Vacek, S.; Podrázský, V.; Linda, R.; Kovařík, J. Forest biodiversity and production potential of post-mining landscape: Opting for afforestation or leaving it to spontaneous development? Cent. Eur. For. J. 2018, 64, 116–126. [Google Scholar] [CrossRef]
- Frouz, J.; Dvorščík, P.; Vávrová, A.; Doušová, O.; Kadochová, Š.; Matějíček, L. Development of canopy cover and woody vegetation biomass on reclaimed and unreclaimed post-mining sites. Ecol. Eng. 2015, 84, 233–239. [Google Scholar] [CrossRef]
- Nave, L.E.; Swanston, C.W.; Mishra, U.; Nadelhoffer, K.J. Afforestation Effects on Soil Carbon Storage in the United States: A Synthesis. Soil Sci. Soc. Am. J. 2013, 77, 1035–1047. [Google Scholar] [CrossRef]
- Misebo, A.M.; Szostak, M.; Sierka, E.; Pietrzykowski, M.; Woś, B. The interactive effect of reclamation scenario and vegetation types on physical parameters of soils developed on carboniferous mine spoil heap. Land Degrad. Dev. 2023, 34, 3593–3605. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and stem volume equations for tree species in Europe. Silva Fenn. 2005, 63, 1–63. [Google Scholar] [CrossRef]
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.J.; Thivolle-Cazat, A.; Bronisz, K.; et al. Comparison of carbon estimation methods for European forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Ochał, W.; Socha, J.; Grabczyński, S. Accuracy of empirical formulas for determining aboveground biomass of black alder (Alnus glutinosa (L.) Gaertn.). Sylwan 2014, 158, 431–442. [Google Scholar]
- Annighöfer, P.; Ameztegui, A.; Ammer, C.; Balandier, P.; Bartsch, N.; Bolte, A.; Coll, L.; Collet, C.; Ewald, J.; Frischbier, N.; et al. Species-specific and generic biomass equations for seedlings and saplings of European tree species. Eur. J. For. Res. 2016, 135, 313–329. [Google Scholar] [CrossRef]
- Bronisz, K.; Strub, M.; Cieszewski, C.; Bijak, S.; Bronisz, A.; Tomusiak, R.; Wojtan, R.; Zasada, M. Empirical equations for estimating aboveground biomass of Betula pendula growing on former farmland in central Poland. Silva Fenn. 2016, 50, 1559. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of common and grey alders growing on abandoned farmland and some practical implications. Biomass Bioenergy 2000, 18, 147–159. [Google Scholar] [CrossRef]
- Zasada, M.; Bronisz, K.; Bijak, S.; Wojtan, R.; Tomusiak, R.; Dudek, A.; Michalak, K. Empirical formulae for determination of the dry biomass of aboveground parts of the tree. Sylwan 2008, 152, 27–39. [Google Scholar]
- Blujdea, V.N.B.; Pilli, R.; Dutca, I.; Ciuvat, L.; Abrudan, I.V. Allometric biomass equations for young broadleaved trees in plantations in Romania. For. Ecol. Manag. 2012, 264, 172–184. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of European aspen growing on abandoned farmland and some practical implications. Biomass Bioenergy 1999, 17, 471–480. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Allometric equations for estimating compartment biomass and stem volume in mature hybrid poplars: General or site-specific? Forests 2017, 8, 309. [Google Scholar] [CrossRef]
- Annighöfer, P.; Mölder, I.; Zerbe, S.; Kawaletz, H.; Terwei, A.; Ammer, C. Biomass functions for the two alien tree species Prunus serotina Ehrh. and Robinia pseudoacacia L. in floodplain forests of Northern Italy. Eur. J. For. Res. 2012, 131, 1619–1635. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System); Version 13; TIBCO Software Inc.: Palo Alto, CA, USA, 2017. [Google Scholar]
- Fettweis, U.; Bens, O.; Hüttl, R.F. Accumulation and properties of soil organic carbon at reclaimed sites in the Lusatian lignite mining district afforested with Pinus sp. Geoderma 2005, 129, 81–91. [Google Scholar] [CrossRef]
- Frouz, J. Effects of soil development time and litter quality on soil carbon sequestration: Assessing soil carbon saturation with a field transplant experiment along a post-mining chronosequence. Land Degrad. Dev. 2016, 28, 664–672. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Characterization of organic matter and carbon cycling in rehabilitated lignite-rich mine soils. Water Air Soil Pollut. 2003, 3, 153–166. [Google Scholar] [CrossRef]
- Chabbi, A.; Sebilo, M.; Rumpel, C.; Schaaf, W.; Mariotti, A. Origin of nitrogen in reforested lignite-rich mine soils revealed by stable isotope analysis. Environ. Sci. Technol. 2008, 42, 2787–2792. [Google Scholar] [CrossRef]
- Amichev, B.Y.; Burger, J.A.; Rodrigue, J.A. Carbon sequestration by forests and soils on mined land in the Midwestern and Appalachian coalfields of the U.S. For. Ecol. Manag. 2008, 256, 1949–1959. [Google Scholar] [CrossRef]
- Akala, V.A.; Lal, R. Soil organic carbon pools and sequestration rates in reclaimed minesoils in Ohio. J. Environ. Qual. 2001, 30, 2098–2104. [Google Scholar] [CrossRef] [PubMed]
- Frouz, J.; Krištůfek, V.; Livečková, M.; van Loo, D.; Jacobs, P.; Van Hoorebeke, L. Microbial properties of soil aggregates created by earthworms and other factors: Spherical and prismatic soil aggregates from unreclaimed post-mining sites. Folia Microbiol. 2011, 56, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Bartuska, M.; Frouz, J. Carbon accumulation and changes in soil chemistry in reclaimed open-cast coal mining heaps near Sokolov using repeated measurement of chronosequence sites. Eur. J. Soil Sci. 2015, 66, 104–111. [Google Scholar] [CrossRef]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cecillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P.; et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Woś, B.; Chodak, M.; Józefowska, A.; Pietrzykowski, M. Influence of tree species on carbon, nitrogen, and phosphorus stocks and stoichiometry under different soil regeneration scenarios on reclaimed and afforested mine and post-fire forest sites. Geoderma 2022, 415, 115782. [Google Scholar] [CrossRef]
- Dilly, O.; Munch, J.-C. Microbial biomass content, basal respiration and enzyme activities during the course of decomposition of leaf litter in a black alder (Alnus glutinosa (L.) Gaertn.) forest. Soil Biol. Biochem. 1996, 28, 1073–1081. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Gruba, P.; Socha, J. Exploring the effects of dominant forest tree species, soil texture, altitude, and pHH2O on soil carbon stocks using generalized additive models. For. Ecol. Manag. 2019, 447, 105–114. [Google Scholar] [CrossRef]
- De Vos, B.; Cools, N.; Ilvesniemi, H.; Vesterdal, L.; Vanguelova, E.; Carnicelli, S. Benchmark values for forest soil carbon stocks in Europe: Results from a large scale forest soil survey. Geoderma 2015, 251–252, 33–46. [Google Scholar]
- Vindušková, O.; Frouz, J. Soil carbon accumulation after open-cast coal and oil shale mining in Northern Hemisphere: A quantitative review. Environ. Earth Sci. 2013, 69, 1685–1698. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Krzaklewski, W. Reclamation to Forest in Sand-Filing Mining Exemplified on Szczakowa Sand Mine Cast; Publishing House of the University of Agriculture: Krakow, Poland, 2009. [Google Scholar]
- Vacek, Z.; Linda, R.; Cukor, J.; Vacek, S.; Šimůnek, V.; Gallo, J.; Vančura, K. Scots pine (Pinus sylvestris L.), the suitable pioneer species for afforestation of reclamation sites? For. Ecol. Manag. 2021, 485, 118951. [Google Scholar] [CrossRef]
- Groninger, J.; Skousen, J.; Angel, P.; Barton, C.; Burger, J.; Zipper, C. Mine reclamation practices to enhance forest development through natural succession. In The Forestry Reclamation Approach: Guide to Successful Reforestation of Mined Lands; Adams, M.B., Ed.; General Technical Report NRS-169; Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017; Chapter 8; pp. 1–7. [Google Scholar]
- Huston, M.A.; Marland, G. Carbon management and biodiversity. J. Environ. Manag. 2003, 67, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, N.; Wright, W.; Loyn, R.H.; Nally, R.M. Ecologically complex carbon’—Linking biodiversity values, carbon storage and habitat structure in some austral temperate forests. Glob. Ecol. Biogeogr. 2011, 20, 260–271. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variant | Woodland from Succession on Bare Carboniferous Rock | Woodland from Succession in Site after Topsoil Application | Woodland with Robinia pseudoacacia from Planting in Site after Topsoil Application | Woodland with a Mixture of Different Tree Species on Site after Topsoil Application | |
|---|---|---|---|---|---|
| S-BR | S-TS | Rb-TS | Re-TS | ||
| Dominant tree species | Betula pendula, Populus tremula, Pinus sylvestris | Populus tremula | Robinia pseudoacacia | Betula pendula, Alnus glutinosa | |
| Admixture | Populus nigra, Populus hybrid ‘Androscoggin’ | Populus nigra, Padus serotina, Betula pendula | –– | Salix alba, Robinia pseudoacacia | |
| Basic soil parameter | Soil layer | Value (mean ± SE) | |||
| Sand (2–0.05 mm) [%] | 0–10 cm | 67 ± 3 | 41 ± 1 | 55 ± 7 | 47 ± 8 |
| 10–30 cm | 42 ± 4 | 43 ± 10 | 44 ± 5 | 43 ± 5 | |
| Silt (0.05–0.002 mm) [%] | 0–10 cm | 26 ± 2 | 48 ± 2 | 38 ± 7 | 44 ± 7 |
| 10–30 cm | 44 ± 4 | 47 ± 8 | 48 ± 5 | 47 ± 5 | |
| Clay (<0.002 mm) [%] | 0–10 cm | 8 ± 1 | 11 ± 1 | 7 ± 1 | 9 ± 1 |
| 10–30 cm | 15 ± 0 | 10 ± 2 | 9 ± 1 | 9 ± 0 | |
| pH | 0–10 cm | 5.3 ± 0.2 | 6.9 ± 0.3 | 7.2 ± 0.1 | 6.8 ± 0.1 |
| 10–30 cm | 5.6 ± 0.3 | 7.0 ± 0.4 | 7.6 ± 0.1 | 7.6 ± 0.1 | |
| BD 1 [g cm−3] | 0–10 cm | 1.20 ± 0.06 | 1.59 ± 0.06 | 1.40 ± 0.04 | 1.22 ± 0.10 |
| 10–30 cm | 1.17 ± 0.07 | 1.51 ± 0.06 | 1.38 ± 0.05 | 1.71 ± 0.11 | |
| Species Source | Biomass Fraction 1 | Form of Equation | Parameter | ||||
|---|---|---|---|---|---|---|---|
| b0 | b1 | b2 | b3 | b4 | |||
| Robinia pseudoacacia [40] | ST | 3.6628 | 2.3732 | 1.0733 *2 | –– | –– | |
| B | 3.481 | 2.0551 | 1.1387 * | –– | –– | ||
| FL | 4.3429 | 1.1018 | 1.0814 * | –– | –– | ||
| AB | 4.2568 | 2.2698 | 1.0913 * | –– | –– | ||
| Betula pendula [41] | ST | 0.02606 | 1.70530 | 1.16391 | –– | –– | |
| B | 0.12882 | 0.03762 | −0.04669 | –– | –– | ||
| FL | 0.06611 | 0.01182 | −0.01331 | –– | –– | ||
| AB | –– | –– | –– | –– | –– | ||
| Alnus glutinosa [42] | ST | 0.00119 | 2.17247 | –– | –– | –– | |
| B | 0.0000006 | 3.28106 | –– | –– | –– | ||
| FL | 0.00239 | 1.32553 | –– | –– | –– | ||
| AB | 0.00079 | 2.28546 | –– | –– | –– | ||
| Pinus sylvestris [43] | ST | 0.00041 | 1.627725 | 1.390374 | 0.000192 | 2.11719 | |
| B | 0.0000038 | 3.653659 | −1.6008 | –– | –– | ||
| FL | 0.000212 | 2.30978 | −0.58099 | –– | –– | ||
| AB | –– | –– | –– | –– | –– | ||
| Salix alba [44] | ST | 2.8992 | 2.6728 | 1.2119 * | –– | –– | |
| B | 2.6787 | 2.6347 | 1.3421 * | –– | –– | ||
| FL | 0.3882 | 3.3431 | 1.8357 * | –– | –– | ||
| AB | –– | –– | –– | –– | –– | ||
| Populus tremula [45] | ST | 0.000065 | 2.603533 | –– | –– | –– | |
| B | 0.000515 | 1.873298 | –– | –– | –– | ||
| FL | 0.000847 | 1.481578 | –– | –– | –– | ||
| AB | 0.000146 | 2.603533 | –– | –– | –– | ||
| Populus nigra [40] | ST | 0.000065 | 2.603533 | –– | –– | –– | |
| B | 0.000515 | 1.873298 | –– | –– | –– | ||
| FL | 0.000847 | 1.481578 | –– | –– | –– | ||
| AB | 0.000146 | 2.603533 | –– | –– | –– | ||
| Populus hybrid Androscoggin [46] | ST | −2.01 | 2.14 | –– | –– | –– | |
| B | −5.22 | 2.76 | –– | –– | –– | ||
| FL | –– | –– | –– | –– | –– | –– | |
| AB | −2.14 | 2.26 | –– | –– | –– | ||
| Padus serotina [47] | ST | −95.36 | 18.78 | −0.84 | 0.02 | –– | |
| B | 0.04 | 0.01 | –– | –– | –– | ||
| FL | 0.049727 | 1.868954 | –– | –– | –– | ||
| AB | −2.56 | 2.13 | 0.5 | 1.008 * | –– | ||
| Variant 1 | Forest Floor Plant Biomass [Mg ha−1] | Shrub Biomass [Mg ha−1] | Tree Density [pcs ha−1] | Dbh [cm] | Height [m] | Stem Biomass [Mg ha−1] | Branch Biomass [Mg ha−1] | Foliage Biomass [Mg ha−1] | Aboveground Biomass [Mg ha−1] | Belowground Biomass [Mg ha−1] | Tree Biomass [Mg ha−1] | Total Biomass [Mg ha−1] 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S-BR | 0.38 ± 0.14 a 2 | 0.75 ± 0.50 ab | 800 ± 122 a | 10.74 ± 0.76 a | 9.49 ± 0.58 a | 18.91 ± 5.45 a | 3.95 ± 1.26 a | 1.43 ± 0.63 a | 31.14 ± 7.37 a | 7.79 ± 1.84 a | 38.93 ± 9.21 a | 40.06 ± 9.29 a |
| S-TS | 0.33 ± 0.04 a | 0.68 ± 0.26 ab | 1050 ± 65 a | 10.21 ± 0.51 a | 10.63 ± 0.31 a | 13.58 ± 1.96 a | 3.38 ± 0.39 a | 0.91 ± 0.10 a | 30.18 ± 4.66 a | 7.55 ± 1.17 a | 37.73 ± 5.83 a | 38.74 ± 5.92 a |
| Rb-TS | 2.34 ± 0.43 b | 0.15 ± 0.15 a | 1250 ± 253 a | 10.10 ± 0.78 a | 9.71 ± 0.42 a | 13.99 ± 3.76 a | 5.72 ± 1.45 a | 1.33 ± 0.29 a | 20.03 ± 5.28 a | 5.01 ± 1.32 a | 25.03 ± 6.60 a | 27.52 ± 6.33 a |
| Re-TS | 1.63 ± 0.68 ab | 3.90 ± 1.63 b | 1450 ± 312 a | 8.78 ± 0.53 a | 9.15 ± 0.56 a | 25.10 ± 4.06 a | 3.71 ± 0.85 a | 1.66 ± 0.39 a | 30.19 ± 5.24 a | 7.55 ± 1.31 a | 37.74 ± 6.54 a | 43.27 ± 8.29 a |
| Variant 1 | C Stock in Soil [Mg ha−1] | C Stock in Biomass [Mg ha−1] | Ecosystem C Stock [Mg ha−1] | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Litter | 0–10 cm | 10–30 cm | Total Soil | Forest Floor Plants | Shrubs | Tree Aboveground | Tree Roots | Trees | Total Biomass 2 | ||
| S-BR | 5.91 ± 0.76 b 2 | 149.97 ± 17.29 b | 351.77 ± 50.74 b | 507.66 ± 60.32 b | 0.16 ± 0.06 a | 0.35 ± 0.24 ab | 11.31 ± 3.31 a | 3.65 ± 0.88 a | 14.97 ± 4.14 a | 15.48 ± 4.24 a | 523.14 ± 62.75 c |
| S-TS | 3.07 ± 0.38 a | 26.10 ± 1.64 a | 26.88 ± 7.02 a | 56.05 ± 8.40 a | 0.13 ± 0.02 a | 0.33 ± 0.13 ab | 8.21 ± 1.13 a | 3.42 ± 0.54 a | 11.63 ± 1.67 a | 12.09 ± 1.72 a | 68.13 ± 9.07 a |
| Rb-TS | 5.31 ± 0.55 ab | 58.76 ± 17.79 a | 44.13 ± 12.54 a | 108.19 ± 17.29 a | 0.97 ± 0.18 b | 0.07 ± 0.07 a | 9.66 ± 2.56 a | 2.20 ± 0.60 a | 11.86 ± 3.15 a | 12.89 ± 3.05 a | 121.08 ± 16.15 b |
| Re-TS | 4.59 ± 0.52 ab | 35.39 ± 4.10 a | 34.78 ± 2.25 a | 74.76 ± 3.75 a | 0.68 ± 0.28 ab | 1.80 ± 0.76 b | 14.17 ± 2.39 a | 3.50 ± 0.60 a | 17.67 ± 2.99 a | 20.15 ± 3.80 a | 94.90 ± 2.58 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woś, B.; Misebo, A.M.; Ochał, W.; Klamerus-Iwan, A.; Pająk, M.; Sierka, E.; Kompała-Bąba, A.; Bujok, M.; Bierza, W.; Józefowska, A.; et al. Biodiversity Characteristics and Carbon Sequestration Potential of Successional Woody Plants versus Tree Plantation under Different Reclamation Treatments on Hard-Coal Mine Heaps––A Case Study from Upper Silesia. Sustainability 2024, 16, 4793. https://doi.org/10.3390/su16114793
Woś B, Misebo AM, Ochał W, Klamerus-Iwan A, Pająk M, Sierka E, Kompała-Bąba A, Bujok M, Bierza W, Józefowska A, et al. Biodiversity Characteristics and Carbon Sequestration Potential of Successional Woody Plants versus Tree Plantation under Different Reclamation Treatments on Hard-Coal Mine Heaps––A Case Study from Upper Silesia. Sustainability. 2024; 16(11):4793. https://doi.org/10.3390/su16114793
Chicago/Turabian StyleWoś, Bartłomiej, Amisalu Milkias Misebo, Wojciech Ochał, Anna Klamerus-Iwan, Marek Pająk, Edyta Sierka, Agnieszka Kompała-Bąba, Michał Bujok, Wojciech Bierza, Agnieszka Józefowska, and et al. 2024. "Biodiversity Characteristics and Carbon Sequestration Potential of Successional Woody Plants versus Tree Plantation under Different Reclamation Treatments on Hard-Coal Mine Heaps––A Case Study from Upper Silesia" Sustainability 16, no. 11: 4793. https://doi.org/10.3390/su16114793
APA StyleWoś, B., Misebo, A. M., Ochał, W., Klamerus-Iwan, A., Pająk, M., Sierka, E., Kompała-Bąba, A., Bujok, M., Bierza, W., Józefowska, A., Bujak, J., & Pietrzykowski, M. (2024). Biodiversity Characteristics and Carbon Sequestration Potential of Successional Woody Plants versus Tree Plantation under Different Reclamation Treatments on Hard-Coal Mine Heaps––A Case Study from Upper Silesia. Sustainability, 16(11), 4793. https://doi.org/10.3390/su16114793