
Bird Species Use of Bioenergy Croplands in Illinois, USA—Can Advanced Switchgrass Cultivars Provide Suitable Habitats for Breeding Grassland Birds?

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
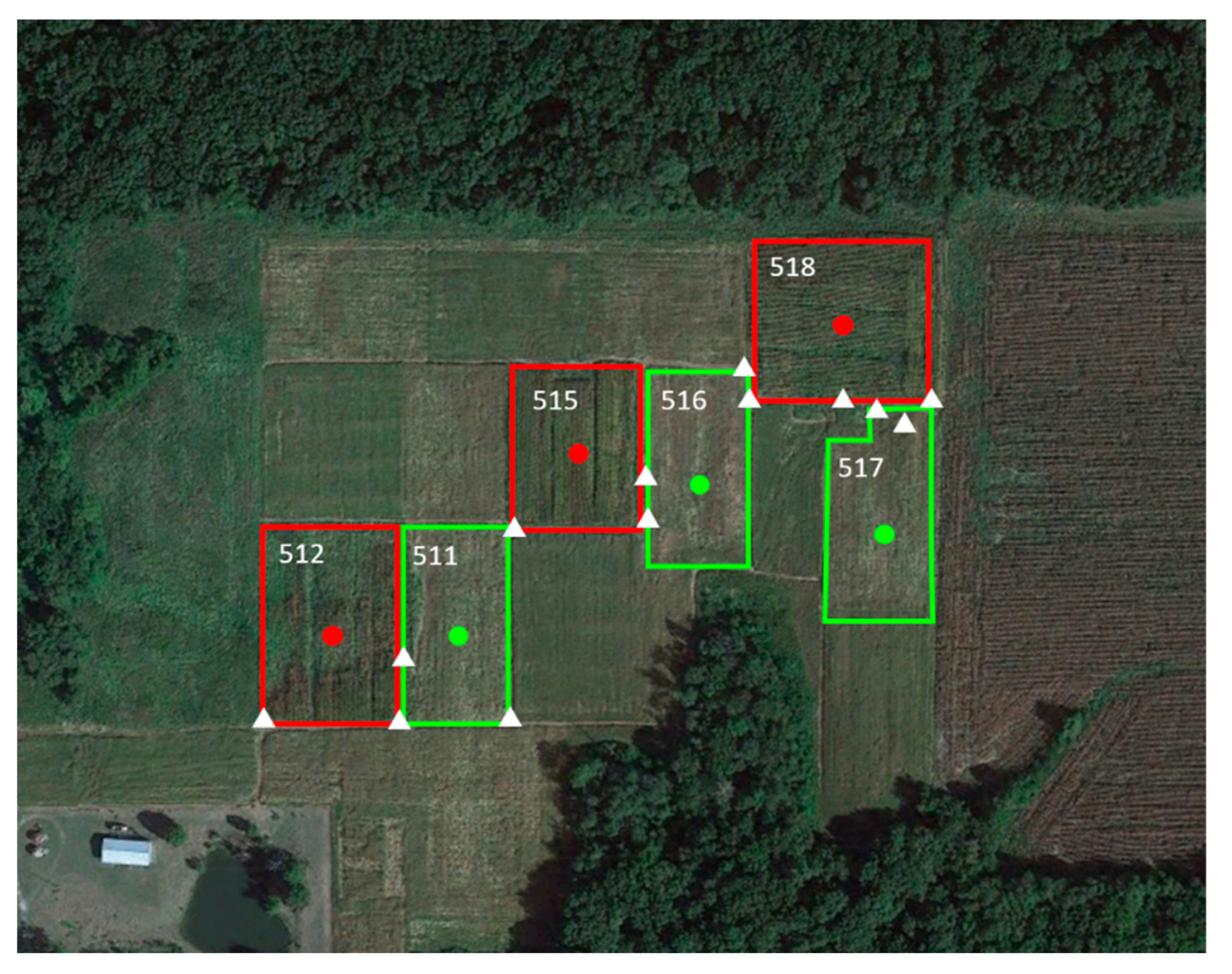
2.1. Study Sites
2.2. Point Counts
2.3. Acoustic Monitoring
2.4. Data Processing and Statistical Analyses
3. Results
3.1. Brighton Site
3.1.1. Point Counts
3.1.2. Acoustic Monitoring
3.2. Urbana Site
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langholtz, M.H.; Stokes, B.J.; Eaton, L.M. Billion-Ton Report: Advancing Domestic Resources for a Thriving Bioeconomy; U.S. DOE Energy Efficiency and Renewable Energy Bioenergies Technologies Office: Washington, DC, USA; Oak Ridge National Laboratory: Oak Ridge, TN, USA, 2016; pp. 1–411.
- U.S. Department of Energy. 2023 Billion-Ton Report: An Assessment of U.S. Renewable Carbon Resources; Oak Ridge National Laboratory: Oak Ridge, TN, USA, 2024; ORNL/SPR-2024/3103. [CrossRef]
- Fargione, J.E.; Cooper, T.R.; Flaspohler, D.J.; Hill, J.; Lehman, C.; Tilman, D.; McCoy, T.; McLeod, S.; Nelson, E.J.; Oberhauser, K.S. Bioenergy and wildlife: Threats and opportunities for grassland conservation. BioScience 2009, 59, 767–777. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Liu, X.V. Environmental implications of higher ethanol production and use in the US: A literature review. Part I–Impacts on water, soil, and air quality. Renew. Sustain. Energy Rev. 2018, 81, 3140–3158. [Google Scholar] [CrossRef]
- Mumm, R.H.; Goldsmith, P.D.; Rausch, K.D.; Stein, H.H. Land usage attributed to corn ethanol production in the United States: Sensitivity to technological advances in corn grain yield, ethanol conversion, and co-product utilization. Biotechnol. Biofuels 2014, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture Economic Research Service. Feed Grains: Yearbook Tables. Available online: https://www.ers.usda.gov/data-products/feed-grains-database/feed-grains-yearbook-tables (accessed on 15 February 2024).
- Beckmann, M.; Gerstner, K.; Akin-Fajiye, M.; Ceaușu, S.; Kambach, S.; Kinlock, N.L.; Phillips, H.R.; Verhagen, W.; Gurevitch, J.; Klotz, S. Conventional land-use intensification reduces species richness and increases production: A global meta-analysis. Glob. Chang. Biol. 2019, 25, 1941–1956. [Google Scholar] [CrossRef]
- Robertson, B.A.; Doran, P.J.; Loomis, E.R.; Robertson, J.R.; Schemske, D.W. Avian use of perennial biomass feedstocks as post-breeding and migratory stopover habitat. PLoS ONE 2011, 6, e16941. [Google Scholar] [CrossRef] [PubMed]
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef]
- Lark, T.J.; Spawn, S.A.; Bougie, M.; Gibbs, H.K. Cropland expansion in the United States produces marginal yields at high costs to wildlife. Nat. Commun. 2020, 11, 4295. [Google Scholar] [CrossRef]
- Ssegane, H.; Negri, M.C.; Quinn, J.; Urgun-Demirtas, M. Multifunctional landscapes: Site characterization and field-scale design to incorporate biomass production into an agricultural system. Biomass Bioenergy 2015, 80, 179–190. [Google Scholar] [CrossRef]
- Ferrarini, A.; Serra, P.; Almagro, M.; Trevisan, M.; Amaducci, S. Multiple ecosystem services provision and biomass logistics management in bioenergy buffers: A state-of-the-art review. Renew. Sustain. Energy Rev. 2017, 73, 277–290. [Google Scholar] [CrossRef]
- Zumpf, C.; Quinn, J.; Cacho, J.; Grasse, N.; Negri, M.C.; Lee, D. Invertebrate and plant community diversity of an Illinois corn–soybean field with integrated shrub willow bioenergy buffers. Sustainability 2021, 13, 12280. [Google Scholar] [CrossRef]
- Mosier, S.; Córdova, S.; Robertson, G.P. Restoring soil fertility on degraded lands to meet food, fuel, and climate security needs via perennialization. Front. Sustain. Food Syst. 2021, 5, 706142. [Google Scholar] [CrossRef]
- Staie, B.; Kinzer, A.; Macknick, J.; Wang, Y.; Cortright, R.; Foust, T.; Ghantous, S.; Lamers, P.; Steward, D. Pathways for Agricultural Decarbonization in the United States; National Renewable Energy Laboratory (NREL): Golden, CO, USA, 2024.
- Nassauer, J.I.; Opdam, P. Design in science: Extending the landscape ecology paradigm. Landsc. Ecol. 2008, 23, 633–644. [Google Scholar] [CrossRef]
- Brandes, E.; McNunn, G.S.; Schulte, L.A.; Bonner, I.J.; Muth, D.J.; Babcock, B.A.; Sharma, B.; Heaton, E.A. Subfield profitability analysis reveals an economic case for cropland diversification. Environ. Res. Lett. 2016, 11, 014009. [Google Scholar] [CrossRef]
- Ssegane, H.; Negri, M.C. An integrated landscape designed for commodity and bioenergy crops for a tile-drained agricultural watershed. J. Environ. Qual. 2016, 45, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Bonner, I.J.; Cafferty, K.G.; Muth, D.J., Jr.; Tomer, M.D.; James, D.E.; Porter, S.A.; Karlen, D.L. Opportunities for energy crop production based on subfield scale distribution of profitability. Energies 2014, 7, 6509–6526. [Google Scholar] [CrossRef]
- Cacho, J.; Negri, M.; Zumpf, C.; Campbell, P. Introducing perennial biomass crops into agricultural landscapes to address water quality challenges and provide other environmental services. Wiley Interdiscip. Rev. Energy Environ. 2018, 7, e275. [Google Scholar] [CrossRef]
- Wang, M.; Han, J.; Dunn, J.B.; Cai, H.; Elgowainy, A. Well-to-wheels energy use and greenhouse gas emissions of ethanol from corn, sugarcane and cellulosic biomass for U.S. use. Environ. Res. Lett. 2012, 7, 045905. [Google Scholar] [CrossRef]
- Brambilla, M.; Gustin, M.; Cento, M.; Ilahiane, L.; Celada, C. Habitat, climate, topography and management differently affect occurrence in declining avian species: Implications for conservation in changing environments. Sci. Total Environ. 2020, 742, 140663. [Google Scholar] [CrossRef]
- Fisher, R.J.; Davis, S.K. From Wiens to Robel: A review of grassland-bird habitat selection. J. Wildl. Manag. 2010, 74, 265–273. [Google Scholar] [CrossRef]
- Pelosi, C.; Bonthoux, S.; Castellarini, F.; Goulard, M.; Ladet, S.; Balent, G. Is there an optimum scale for predicting bird species’ distribution in agricultural landscapes? J. Environ. Manag. 2014, 136, 54–61. [Google Scholar] [CrossRef]
- Kreig, J.A.; Parish, E.; Jager, H.I. Growing grasses in unprofitable areas of U.S. Midwest croplands could increase species richness. Biol. Conserv. 2021, 261, 109289. [Google Scholar] [CrossRef]
- Rosenburg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Douglass, D.J.T.; Waldinger, J.; Buckmire, Z.; Gibb, K.; Medina, J.P.; Sutcliffe, L.; Beckmann, C.; Collar, N.J.; Jansen, R.; Kamp, J.; et al. A global review identifies agriculture as the main threat to declining grassland birds. Ibis 2023, 165, 1107–1128. [Google Scholar] [CrossRef]
- With, K.A.; King, A.W.; Jensen, W.E. Remaining large grasslands may not be sufficient to prevent grassland bird declines. Biol. Conserv. 2008, 141, 3152–3167. [Google Scholar] [CrossRef]
- Lee, M.S.; Casler, M.; Lee, D.K. Registration of ‘Independence’ switchgrass. J. Plant Regist. 2024; in press. [Google Scholar] [CrossRef]
- Vogel, K.P.; Mitchell, R.; Casler, M.; Sarath, G. Registration of ‘Liberty’ switchgrass. J. Plant Regist. 2014, 8, 242–247. [Google Scholar] [CrossRef]
- Vogel, K.P.; Hopkins, A.; Moore, K.; Johnson, K.; Carlson, I. Registration of ‘Shawnee’ switchgrass. Crop Sci. 1996, 36, 1713. [Google Scholar] [CrossRef]
- Darras, K.; Batáry, P.; Furnas, B.; Celis-Murillo, A.; Van Wilgenburg, S.L.; Mulyani, Y.A.; Tscharntke, T. Comparing the sampling performance of sound recorders versus point counts in bird surveys: A meta-analysis. J. Appl. Ecol. 2018, 55, 2575–2586. [Google Scholar] [CrossRef]
- Ralph, C.J.; Geupel, G.R.; Pyle, P.; Martin, T.E.; DeSante, D.F. Handbook of Field Methods for Monitoring Landbirds; General Technical Report; PSW-GTR-144; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 1993.
- Kahl, S.; Wood, C.M.; Eibl, M.; Klinck, H. BirdNET: A deep learning solution for avian diversity monitoring. Ecol. Inform. 2021, 61, 101236. [Google Scholar] [CrossRef]
- Cornell Laboratory of Ornithology. eBird. Available online: https://ebird.org/home (accessed on 15 March 2024).
- Kahl, S. Species Range Model Details. Available online: https://github.com/kahst/BirdNET-Analyzer/discussions/234 (accessed on 15 March 2024).
- Pérez-Granados, C. BirdNET: Applications, performance, pitfalls and future opportunities. Ibis 2023, 165, 1068–1075. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Posit Team. RStudio: Integrated Development Environment for R; Posit Software, PBC: Boston, MA, USA, 2023. [Google Scholar]
- Kaschek, D. dMod: Dynamic Modeling and Parameter Estimation in ODE Models. Available online: https://cran.r-project.org/web/packages/dMod/index.html (accessed on 15 March 2024).
- Wickham, H. Stringr: Simple, Consistent Wrappers for Common String Operations. Available online: https://CRAN.R-project.org/package=stringr (accessed on 15 March 2024).
- Wickham, H.; Francois, R.; Henry, L.; Müller, K.; Vaughan, D. Dplyr: A Grammar of Data Manipulation. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 15 March 2024).
- SAS Institute. SAS for Windows, 9.4; SAS Institute, Inc.: Cary, NA, USA, 2016. [Google Scholar]
- Guzy, M.J.; Ritchison, G. Common Yellowthroat (Geothlypis trichas), Version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Herkert, J.R.; Vickery, P.D.; Kroodsma, D.E. Henslow’s Sparrow (Centronyx henslowii), Version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Namoi, N.; Archer, D.; Rosenstock, T.S.; Jang, C.; Lin, C.H.; Boe, A.; Lee, D. How profitable is switchgrass in Illinois, USA? An Economic Definition of Marginal Land. Grassl. Res. 2022, 1, 111–122. [Google Scholar] [CrossRef]
- Herkert, J.R.; Kroodsma, D.E.; Gibbs, J.P. Sedge Wren (Cistothorus stellaris), Version 1.0. In Birds of the World; Poole, A.F., Gill, F.B., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2021. [Google Scholar]
- Vickery, P.D. Grasshopper Sparrow (Ammodramus savannarum), Version 1.0. In Birds of the World; Poole, A.F., Gill, F.B., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Sousa, B.F.; Temple, S.A.; Basili, G.D. Dickcissel (Spiza americana), Version 2.0. In Birds of the World; Schulenberg, T.S., Keeney, B.K., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Robertson, B.A.; Doran, P.J.; Loomis, L.R.; Robertson, J.R.; Schemske, D.W. Perennial biomass feedstocks enhance avian diversity. GCB Bioenergy 2011, 3, 235–246. [Google Scholar] [CrossRef]
- Payne, R.B. Indigo Bunting (Passerina cyanea), Version 1.0. In Birds of the World; Poole, A.F., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Post van der Burg, M.; Otto, C.; MacDonald, G. Trending against the grain: Bird population responses to expanding energy portfolios in the U.S. Northern Great Plains. Ecol. Appl. 2023, 33, e2904. [Google Scholar] [CrossRef] [PubMed]
- Heard, L.P.; Allen, A.W.; Best, L.B.; Brady, S.J.; Burger, W.; Esser, A.J.; Hackett, E.; Johnson, D.H.; Pederson, R.L.; Reynolds, R.E.; et al. Technical Report: A Comprehensive Review of Farm Bill Contributions to Wildlife Conservation, 1985–2000; USDA/NRCS/WHMI-2000; U.S. Department of Agriculture, Natural Resource Conservation Service, Wildlife Habitat Management Institute: Washington, DC, USA, 2000.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of On-Plot Observations (Off-Plot Numbers in Parentheses) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2022 | All Years | |||||
| Species | CO | SW | CO | SW | CO | SW | CO | SW |
| Barn Swallow (Hirundo rustica) | 6 (1) | 6 (5) | 9 (2) | 15 (2) | 4 (3) | 6 (1) | 19 (6) | 27 (8) |
| Brown-headed Cowbird (Molothrus ater) | 42 (30) | 0 (42) | 16 (20) | 4 (32) | 12 (16) | 0 (8) | 70 (66) | 4 (82) |
| Common Yellowthroat (Geothlypis trichas) | 1 (21) | 0 (20) | 0 (26) | 0 (29) | 0 (27) | 8 (28) | 1 (74) | 8 (77) |
| Field Sparrow (Spizella pusilla) | 0 (32) | 0 (35) | 1 (18) | 3 (19) | 5 (32) | 6 (27) | 6 (82) | 9 (81) |
| Indigo Bunting (Passerina cyanea) | 13 (33) | 14 (32) | 38 (47) | 17 (50) | 13 (63) | 24 (59) | 64 (143) | 55 (141) |
| Purple Martin (Progne subis) | 1 (12) | 0 (13) | 9 (4) | 17 (1) | 5 (48) | 6 (30) | 15 (64) | 23 (44) |
| Blue Grosbeak (Passerina caerulea) | 1 (4) | 0 (8) | 1 (4) | 0 (3) | 3 (8) | 0 (3) | 5 (16) | 0 (14) |
| Cliff Swallow (Petrochelidon pyrrhonota) | 1 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (0) | 4 (0) | 3 (0) | 4 (0) |
| Dickcissel (Spiza americana) * | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (5) | 1 (3) | 0 (5) | 1 (3) |
| Eastern Phoebe (Sayornis phoebe) | 0 (4) | 0 (3) | 0 (2) | 0 (2) | 1 (1) | 0 (1) | 1 (7) | 0 (6) |
| Eastern Kingbird (Tyrannus tyrannus) | 0 (2) | 0 (0) | 1 (6) | 0 (3) | 2 (3) | 0 (8) | 3 (11) | 0 (8) |
| European Starling (Sturnus vulgaris) | 0 (0) | 0 (8) | 0 (6) | 0 (7) | 2 (1) | 0 (16) | 2 (7) | 0 (31) |
| Eurasian Tree Sparrow (Passer montanus) | 1 (0) | 0 (8) | 2 (0) | 0 (0) | 0 (0) | 0 (0) | 3 (0) | 0 (8) |
| House Sparrow (Passer domesticus) | 0 (2) | 0 (0) | 0 (1) | 0 (0) | 0 (4) | 1 (0) | 0 (7) | 1 (0) |
| Northern Rough-winged Swallow (Stelgidopteryx serripennis) | 0 (0) | 0 (0) | 0 (1) | 2 (0) | 0 (0) | 0 (0) | 0 (1) | 2 (0) |
| Northern Cardinal (Cardinalis cardinalis) | 0 (15) | 0 (20) | 0 (21) | 0 (22) | 1 (15) | 0 (13) | 1 (51) | 0 (55) |
| Red-winged Blackbird (Agelaius phoeniceus) | 0 (17) | 0 (10) | 1 (10) | 0 (15) | 1 (13) | 1 (12) | 2 (40) | 1 (37) |
| Ruby-throated Hummingbird (Archilochus colubris) | 0 (0) | 0 (1) | 1 (3) | 2 (2) | 0 (0) | 0 (0) | 1 (3) | 2 (3) |
| Song Sparrow (Melospiza melodia) | 0 (11) | 0 (11) | 4 (3) | 0 (5) | 0 (2) | 0 (3) | 4 (16) | 0 (19) |
| Tree Swallow (Tachycineta bicolor) | 2 (5) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (0) | 2 (5) | 2 (0) |
| Willow Flycatcher (Empidonax trailii) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0) | 0 (0) | 1 (0) | 0 (0) |
| Unidentified | 0 (5) | 0 (9) | 4 (3) | 3 (7) | 0 (0) | 0 (3) | 4 (8) | 3 (16) |
| Number of species b | 9 (14) | 2 (14) | 11 (16) | 7 (14) | 13 (15) | 10 (14) | 18 (18) | 13 (16) |
| Number of birds | 68 (194) | 20 (225) | 87 (177) | 63 (199) | 52 (241) | 59 (212) | 207 (612) | 142 (636) |
| Number of surveys c | 27 | 27 | 27 | 27 | 27 | 27 | 54 | 54 |
| Number of birds per survey | 2.5 (7.2) | 0.7 (8.3) | 3.2 (6.6) | 2.3 (7.4) | 1.9 (8.9) | 2.2 (7.9) | 3.8 (11.3) | 2.6 (11.8) |
| Mean Number of Detected Vocalizations/Plot/Day | ||||||||
|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2022 | All Years | |||||
| Species | CO | SW | CO | SW | CO | SW | CO | SW |
| Brown-headed Cowbird (Molothrus ater) | 8.93 | 1.02 | 8.61 | 1.26 | 1.69 | 0.32 | 6.81 | 0.91 |
| Common Yellowthroat (Geothlypis trichas) | 0.92 | 0.66 | 1.53 | 5.52 | 1.42 | 34.03 | 1.27 | 11.62 |
| Dickcissel (Spiza americana) * | 0.01 | 0.04 | 0.07 | 0.04 | 4.84 | 14.71 | 1.37 | 4.12 |
| Field Sparrow (Spizella pusilla) | 13.82 | 9.98 | 12.25 | 11.26 | 17.27 | 17.69 | 14.24 | 12.57 |
| Henslow’s Sparrow (Centronyx henslowii) * | 8.23 | 1.48 | 0.28 | 0.02 | 0.01 | 0.01 | 3.19 | 0.57 |
| Indigo Bunting (Passerina cyanea) | 215.17 | 91.58 | 166.04 | 136.18 | 273.64 | 118.36 | 214.44 | 114.46 |
| Purple Martin (Progne subis) | 1.41 | 1.90 | 4.79 | 6.55 | 0.54 | 1.35 | 2.34 | 3.35 |
| Sedge Wren (Cistothorus stellaris) * | 1.24 | 21.99 | 0.12 | 13.86 | 0.00 | 0.06 | 0.51 | 13.07 |
| Vesper Sparrow (Pooecetes gramineus) | 3.03 | 2.73 | 3.11 | 2.62 | 1.45 | 1.03 | 2.62 | 2.22 |
| Number of On-Plot Observations (Off-Plot Numbers in Parentheses) | |||
|---|---|---|---|
| Species a | Miscanthus | Prairie | Switchgrass |
| Barn Swallow (Hirundo rustica) | 3 (0) | 3 (0) | 5 (0) |
| Common Yellowthroat (Geothlypis trichas) | 0 (0) | 1 (0) | 0 (0) |
| Dickcissel (Spiza americana) * | 0 (0) | 2 (0) | 1 (1) |
| Ring-necked Pheasant (Phasianus colchicus) | 0 (0) | 1 (0) | 0 (0) |
| Red-winged Blackbird (Agelaius phoeniceus) | 2 (5) | 21 (0) | 0 (8) |
| Song Sparrow (Melospiza melodia) | 1 (0) | 1 (0) | 0 (0) |
| Tree Swallow (Tachycineta bicolor) | 1 (0) | 0 (0) | 1 (0) |
| Number of birds | 7 (5) | 29 (0) | 7 (1) |
| Mean Number of Detections/Day | |||
|---|---|---|---|
| Species | Miscanthus | Prairie | Switchgrass |
| Barn Swallow (Hirundo rustica) | 26.31 | 0.31 | 1.56 |
| Common Yellowthroat (Geothlypis trichas) | 22.10 | 15.93 | 23.10 |
| Dickcissel (Spiza americana) * | 1.40 | 92.42 | 55.81 |
| Red-winged Blackbird (Agelaius phoeniceus) | 9.15 | 9.76 | 12.32 |
| Song Sparrow (Melospiza melodia) | 17.94 | 0.93 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaGory, K.E.; Cacho, J.F.; Zumpf, C.R.; Lee, D.; Feinstein, J.; Dematties, D.; Walston, L.J.; Namoi, N.; Negri, M.C. Bird Species Use of Bioenergy Croplands in Illinois, USA—Can Advanced Switchgrass Cultivars Provide Suitable Habitats for Breeding Grassland Birds? Sustainability 2024, 16, 4807. https://doi.org/10.3390/su16114807
LaGory KE, Cacho JF, Zumpf CR, Lee D, Feinstein J, Dematties D, Walston LJ, Namoi N, Negri MC. Bird Species Use of Bioenergy Croplands in Illinois, USA—Can Advanced Switchgrass Cultivars Provide Suitable Habitats for Breeding Grassland Birds? Sustainability. 2024; 16(11):4807. https://doi.org/10.3390/su16114807
Chicago/Turabian StyleLaGory, Kirk E., Jules F. Cacho, Colleen R. Zumpf, DoKyoung Lee, Jeremy Feinstein, Dario Dematties, Leroy J. Walston, Nictor Namoi, and M. Cristina Negri. 2024. "Bird Species Use of Bioenergy Croplands in Illinois, USA—Can Advanced Switchgrass Cultivars Provide Suitable Habitats for Breeding Grassland Birds?" Sustainability 16, no. 11: 4807. https://doi.org/10.3390/su16114807