
Prediction of Adaptability of Typical Vegetation Species in Flood Storage Areas under Future Climate Change: A Case in Hongze Lake FDZ, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Investigation and Collection of Occurrence Data of Species
2.3. Environmental Variables
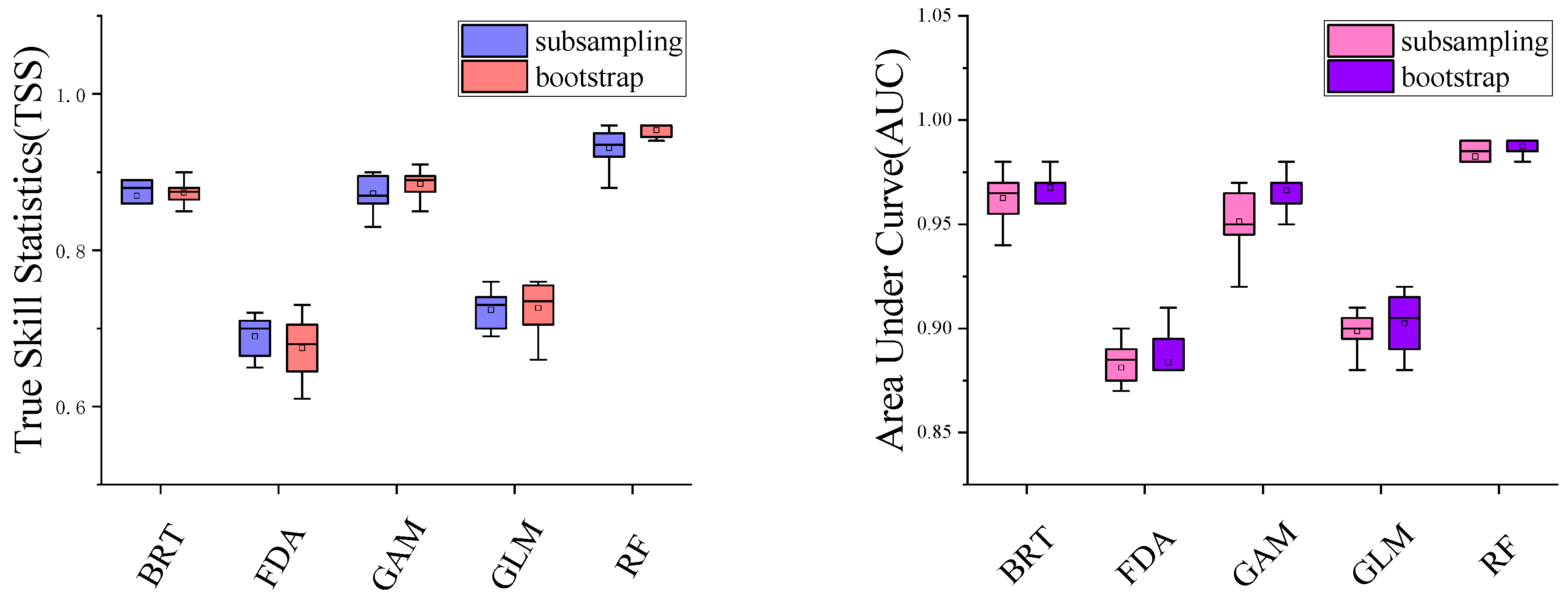
2.4. Model Simulation and Model Evaluation
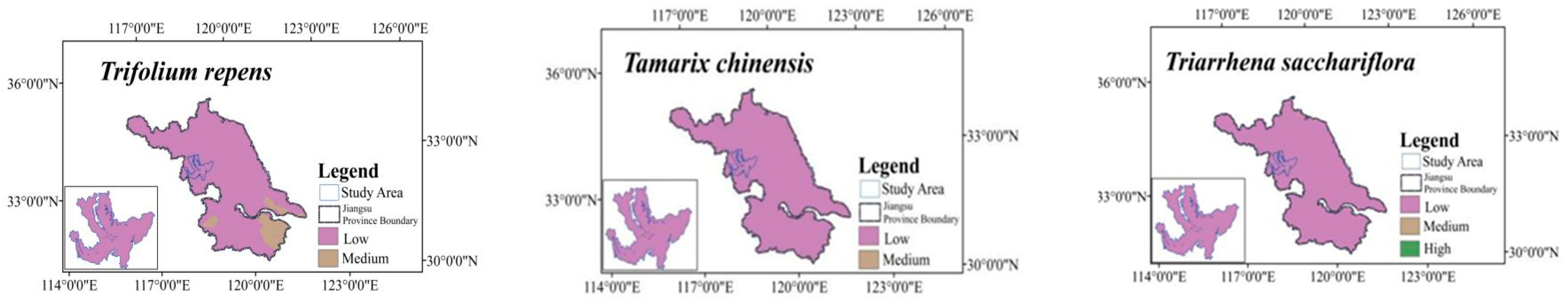
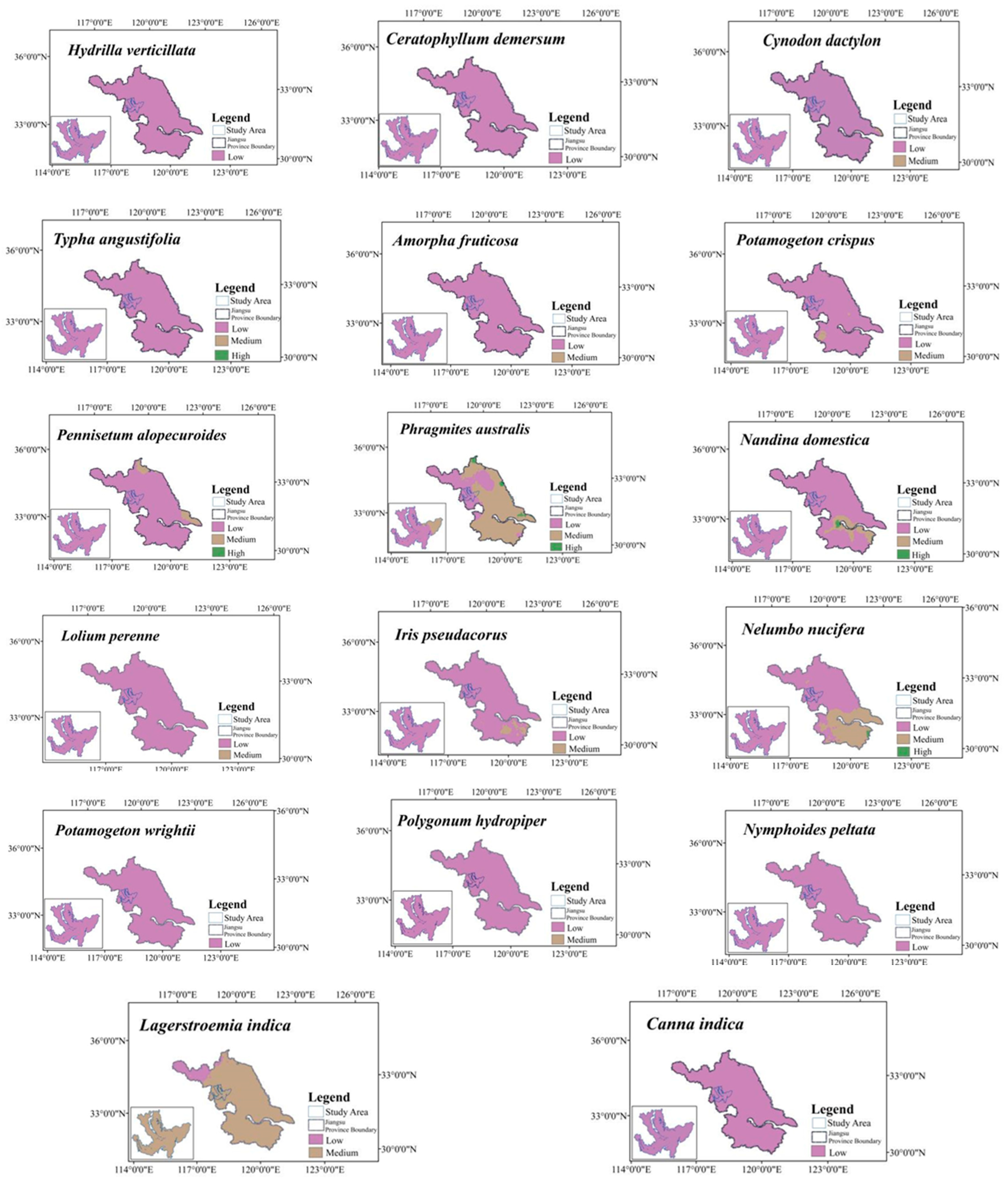
2.5. Potential Distribution under Current Climate Conditions
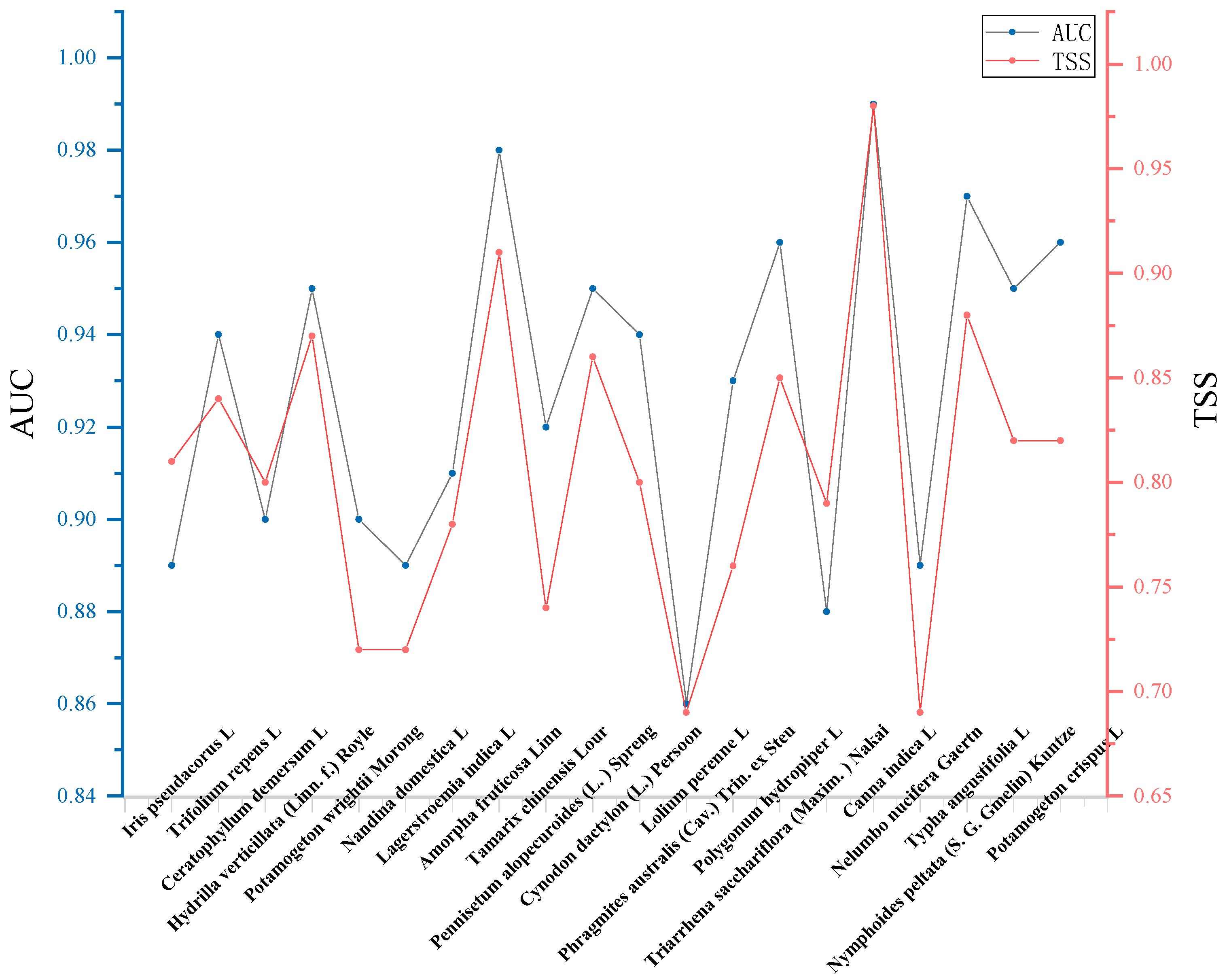
Model Accuracy
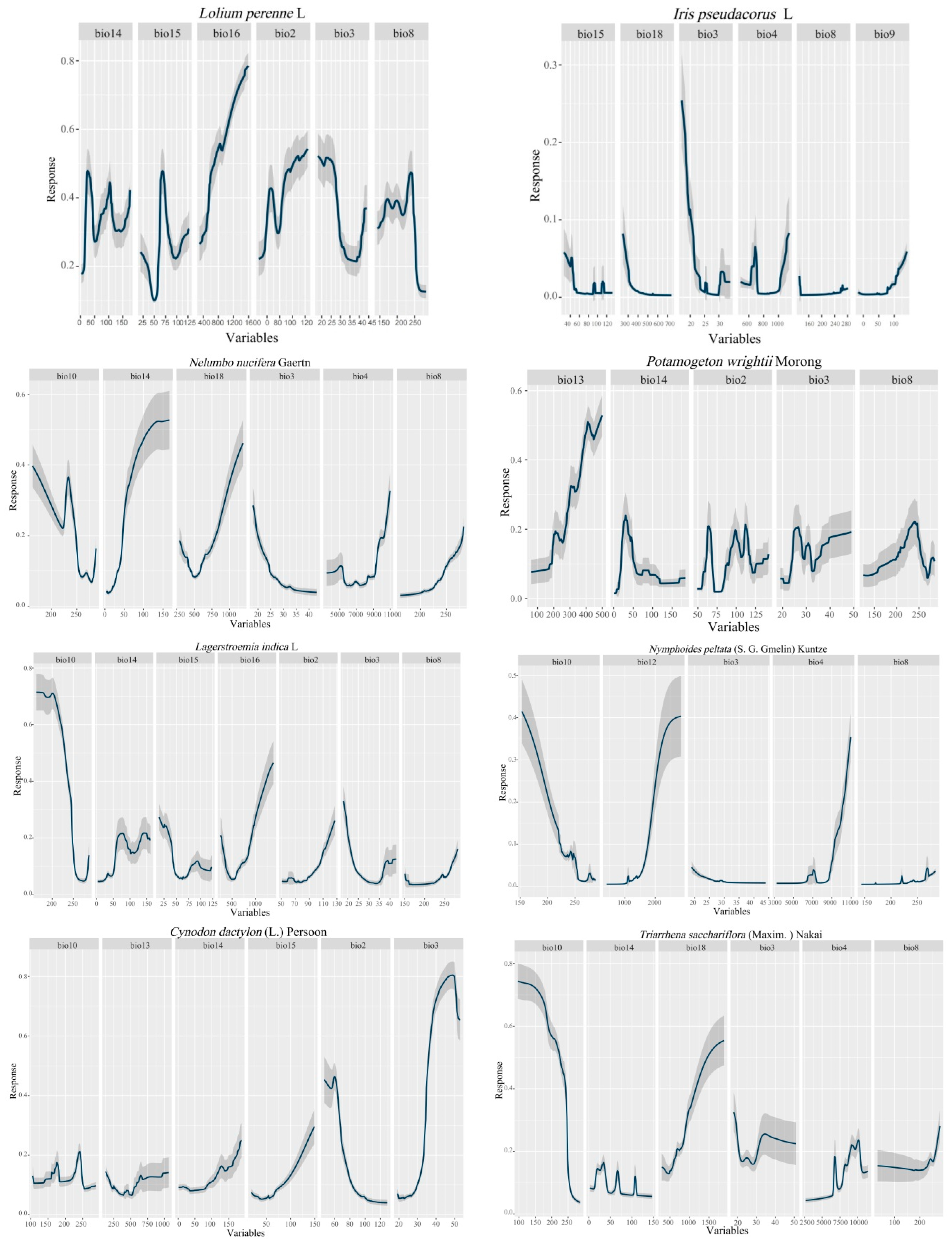
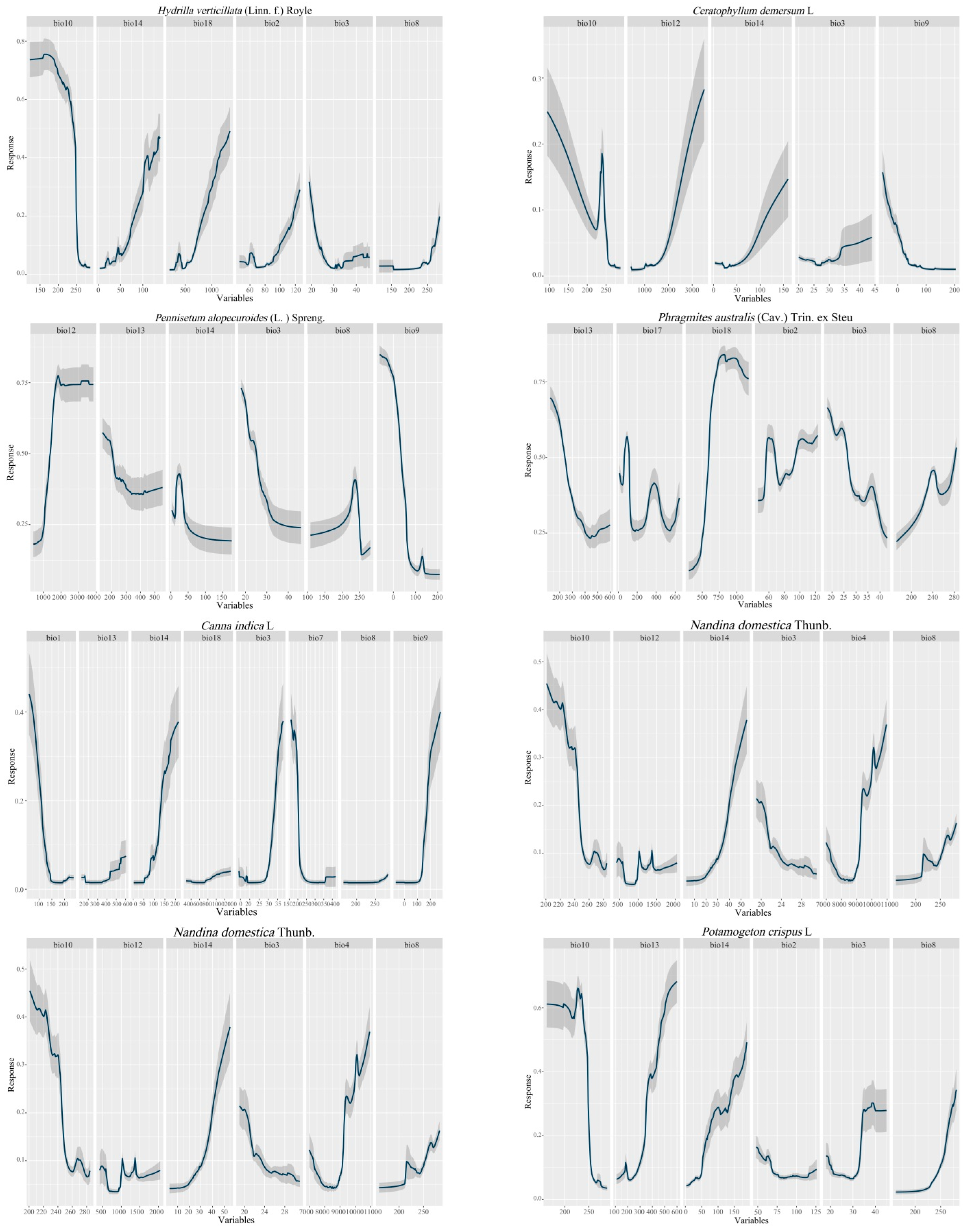
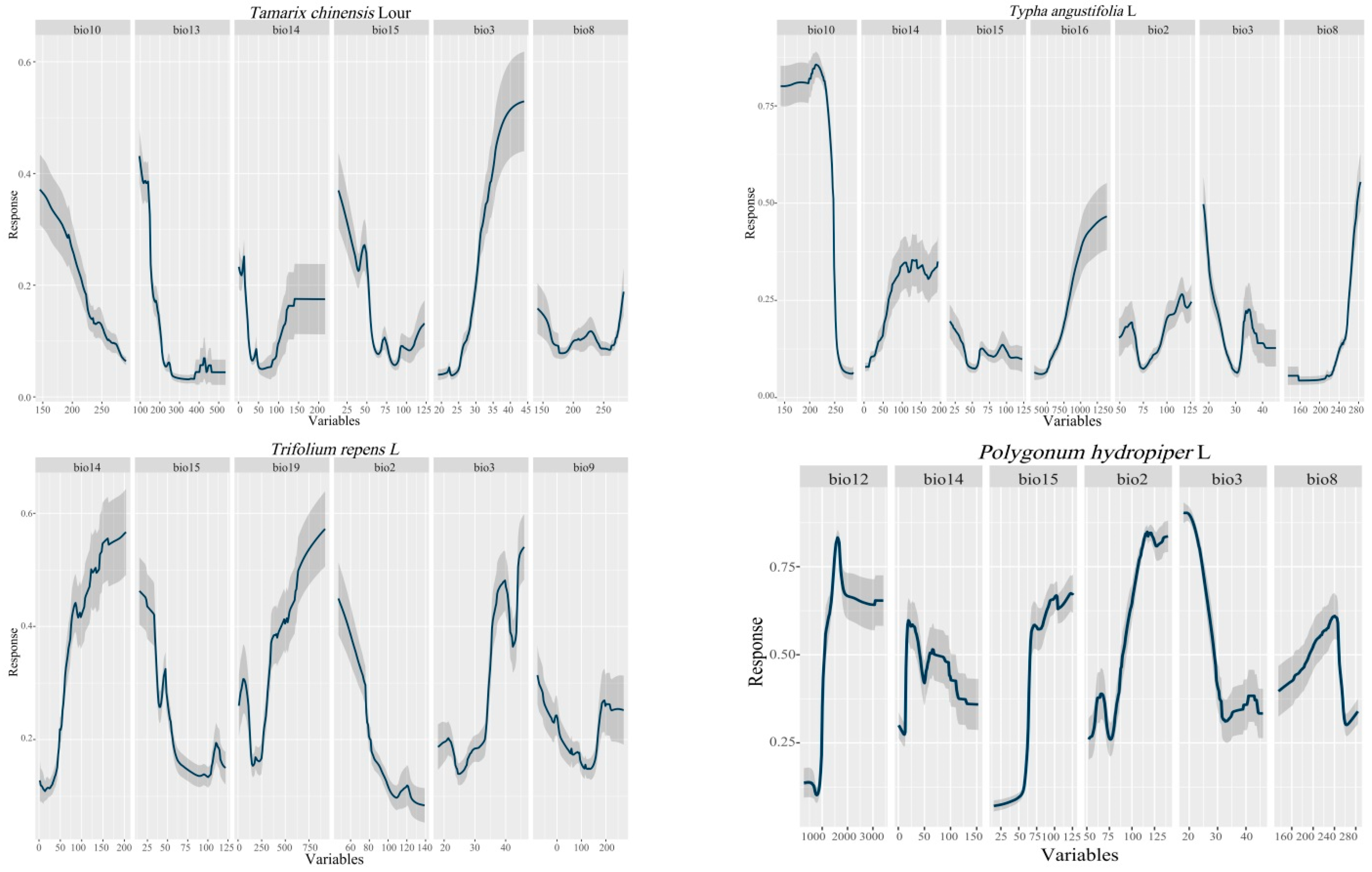
2.6. Environmental Variable Weight and Response Curve
2.7. Typical Vegetation Adaptability Patterns under Current Climatic Conditions
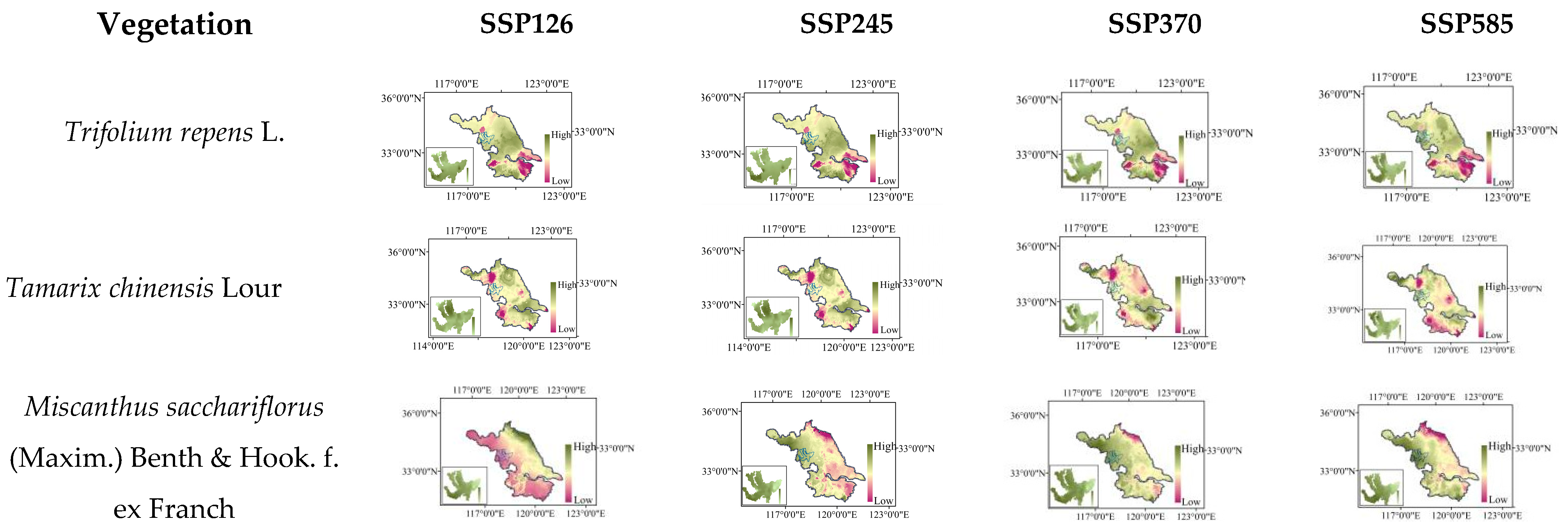
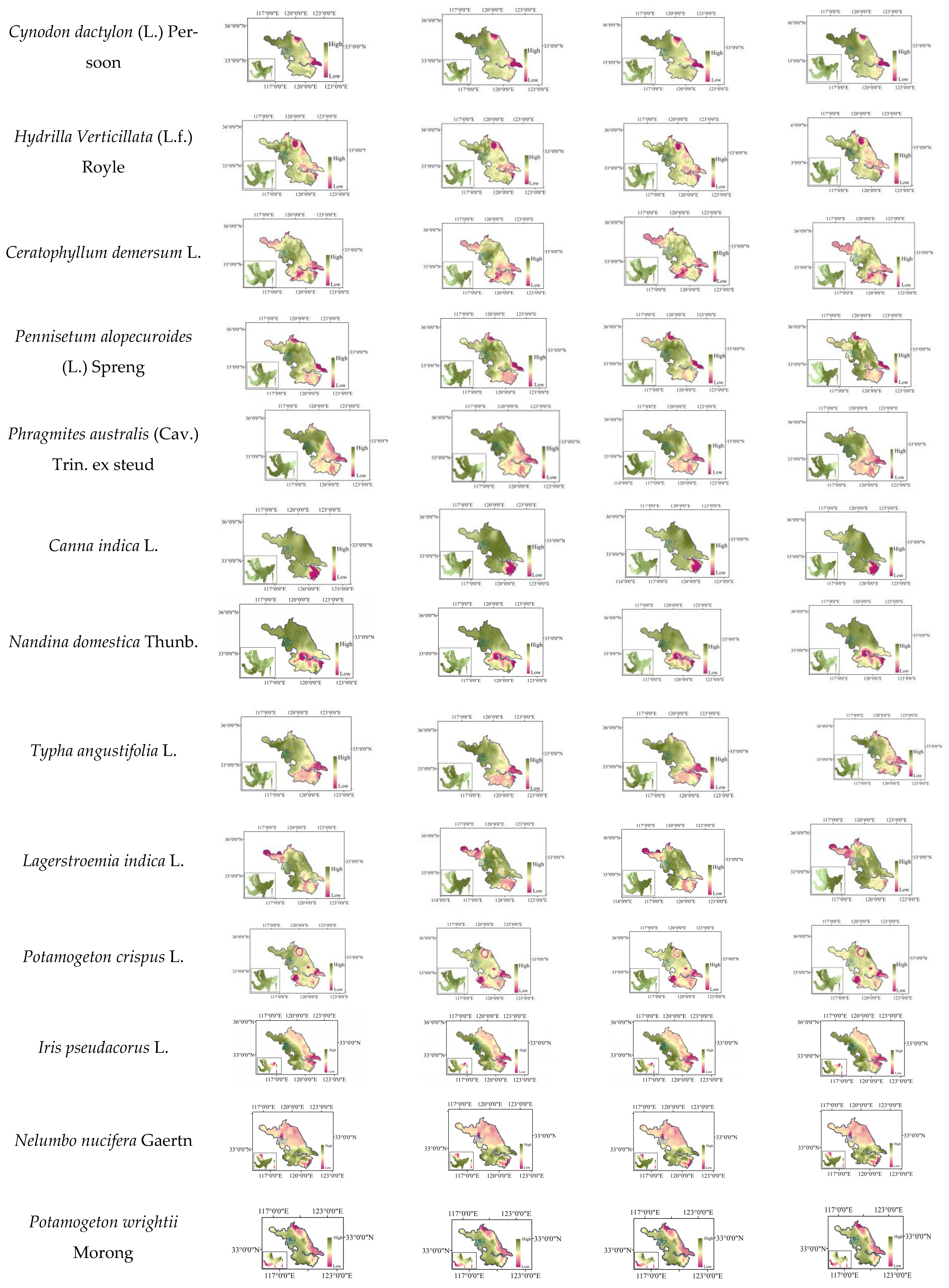
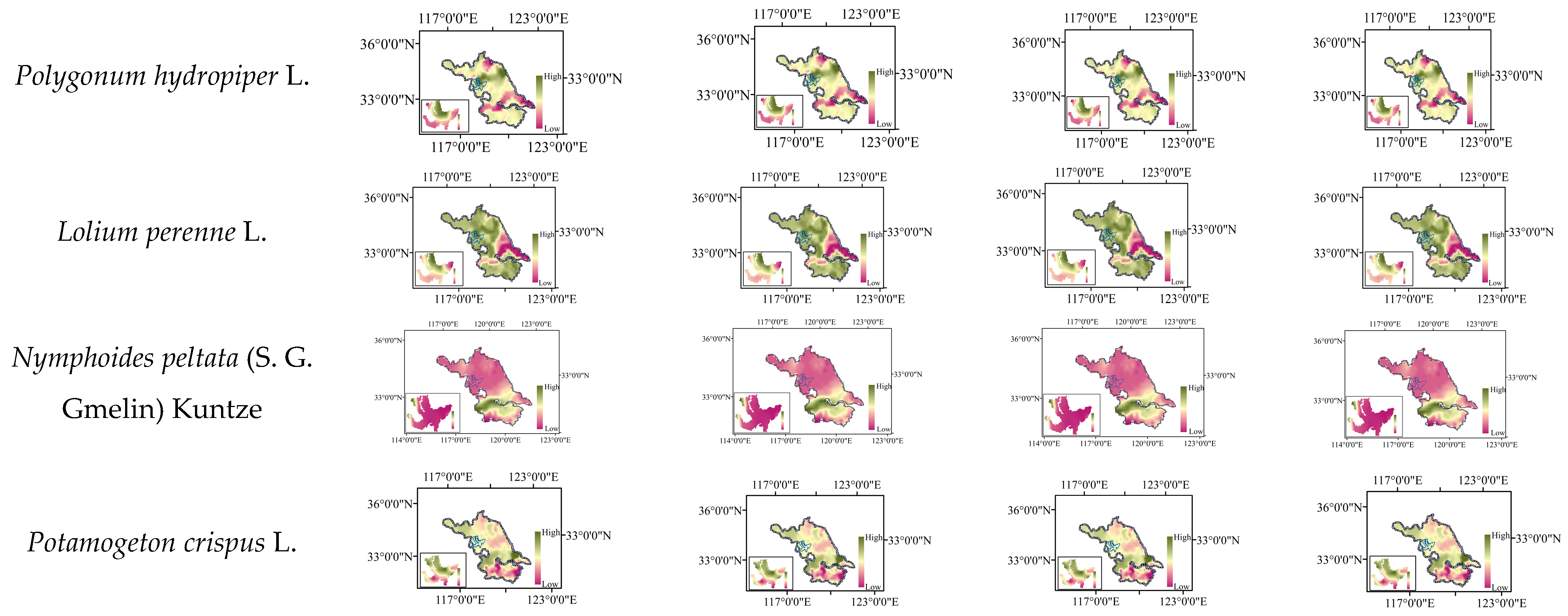
2.8. Distribution Changes under Future Climate Scenarios
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, W.; Li, C.; Xiang, L. Research on Ecological Restoration of Flood Storage and Detention Areas. Water Res. Hydropower Eng. 2003, 7, 61–63. [Google Scholar] [CrossRef]
- Wang, K. Plant Ecological Restoration Technology for Flood Storage and Detention Areas: A Case Study of the Waste Grass Levee Flood Storage and Detention Area in Quanjiao County. Anhui For. Sci. Technol. 2016, 42, 27–29. [Google Scholar]
- Li, C.; Dong, Q.; Guo, Y.; Zhuang, L. Evaluation of vegetation construction techniques in degraded areas of Hongze Lake wetland. J. Anhui Agric. Sci. 2022, 50, 75–81. [Google Scholar] [CrossRef]
- Gong, L.J.; Tian, B.X.; Li, Y.G.; Wu, S. Phenological Changes of Soybean in Response to Climate Conditions in Frigid Region in China over the Past Decades. Int. J. Plant Prod. 2021, 15, 363–375. [Google Scholar] [CrossRef]
- Isbell, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Diaz, S.; Hector, A.; Mace, G.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.E.; et al. Linking the influence and dependence of people on biodiversity across scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Lei, G.C.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–274. [Google Scholar] [CrossRef]
- Meng, H.H.; Zhou, S.S.; Li, L.; Tan, Y.H.; Li, J.W.; Li, J. Conflict between biodiversity conservation and economic growth: Insight into rare plants in tropical China. Biodivers. Conserv. 2019, 28, 523–537. [Google Scholar] [CrossRef]
- Moraitis, M.L.; Valavanis, V.D.; Karakassis, I. Modelling the effects of climate change on the distribution of benthic indicator species in the Eastern Mediterranean Sea. Sci. Total Environ. 2019, 667, 16–24. [Google Scholar] [CrossRef]
- Pacifici, M.; Visconti, P.; Butchart, S.H.M.; Watson, J.E.M.; Cassola, F.M.; Rondinini, C. Species’ traits influenced their response to recent climate change. Nat. Clim. Chang. 2018, 8, 750. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- McCullagh, P. Sampling bias and logistic models. J. R. Stat. Soc. Ser. B Methodol. 2008, 70, 643–664. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araujo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Coetzee, B.W.T.; Robertson, M.P.; Erasmus, B.F.N.; van Rensburg, B.J.; Thuiller, W. Ensemble models predict Important Bird Areas in southern Africa will become less effective for conserving endemic birds under climate change. Glob. Ecol. Biogeogr. 2009, 18, 701–710. [Google Scholar] [CrossRef]
- Naimi, B.; Araujo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, R. Analysis of the purification effect and influencing factors of aquatic vegetation on the water environment of Hongze Lake lakeside wetland. Agric. Sci. Technol. 2012, 13, 211–216. [Google Scholar] [CrossRef]
- Shi, Q.; Hua, J.; Chen, C. The Erosion Prevention Effect of Different Plant Communities on the Embankment of Hongze Lake. Bull. Soil Water Conserv. 2022, 42, 42–48. [Google Scholar] [CrossRef]
- Yuan, X.; Yuan, Y. Research on the Planning and Design of Returning the Hongze Lake to the Lake. Water Conserv. Technical Superv. 2022, 133–143. [Google Scholar]
- Chen, C.; Zhang, M.; Xu, M.; Wang, L. Ecological Risk Assessment of Hongze Lake Basin. People’s Yangtze River 2021, 52, 36–40. [Google Scholar] [CrossRef]
- Tian, Z.; Jiang, D. Analysis of Climate Simulation Capability of CCSM4 with Different Resolutions for East Asia and China. Atmos. Sci. 2013, 37, 171–186. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Riahi, K.; Ebi, K.L.; Hallegatte, S.; Carter, T.R.; Mathur, R.; van Vuuren, D.P. A new scenario framework for climate change research: The concept of shared socioeconomic pathways. Clim. Chang. 2014, 122, 387–400. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carre, G.; Marquez, J.R.G.; Gruber, B.; Lafourcade, B.; Leitao, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Jiang, Y.B.; Wang, T.J.; Wu, Y.P.; Hu, R.G.; Huang, K.; Shao, X.M. Past distribution of epiphyllous liverworts in China: The usability of historical data. Ecol. Evol. 2018, 8, 7436–7450. [Google Scholar] [CrossRef] [PubMed]
- Ao, Y.L.; Li, H.Q.; Zhu, L.P.; Ali, S.; Yang, Z.G. The linear random forest algorithm and its advantages in machine learning assisted logging regression modeling. J. Pet. Sci. Eng. 2019, 174, 776–789. [Google Scholar] [CrossRef]
- Payne, R.W. Developments from analysis of variance through to generalized linear models and beyond. Ann. Appl. Biol. 2014, 164, 11–17. [Google Scholar] [CrossRef]
- Alahuhta, J.; Antikainen, H.; Hjor, J.; Helm, A.; Heino, J. Current climate overrides historical effects on species richness and range size of freshwater plants in Europe and North America. J. Ecol. 2020, 108, 1262–1275. [Google Scholar] [CrossRef]
- Alavipanah, S.; Schreyer, J.; Haase, D.; Lakes, T.; Qureshi, S. The effect of multi-dimensional indicators on urban thermal conditions. J. Clean. Prod. 2018, 177, 115–123. [Google Scholar] [CrossRef]
- Fraker, M.E.; Keitzer, S.C.; Sinclair, J.S.; Aloysius, N.R.; Dippold, D.A.; Yen, H.; Arnold, J.G.; Daggupati, P.; Johnson, M.V.V.; Martin, J.F.; et al. Projecting the effects of agricultural conservation practices on stream fish communities in a changing climate. Sci. Total Environ. 2020, 747, 141112. [Google Scholar] [CrossRef]
- Guo, G.H.; Wu, Z.F.; Xiao, R.B.; Chen, Y.B.; Liu, X.N.; Zhang, X.S. Impacts of urban biophysical composition on land surface temperature in urban heat island clusters. Landsc. Urban Plan. 2015, 135, 1–10. [Google Scholar] [CrossRef]
- Carslaw, D.C.; Taylor, P.J. Analysis of air pollution data at a mixed source location using boosted regression trees. Atmos. Environ. 2009, 43, 3563–3570. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Hu, Y.F.; Dai, Z.X.; Guldmann, J.M. Modeling the impact of 2D/3D urban indicators on the urban heat island over different seasons: A boosted regression tree approach. J. Environ. Manag. 2020, 266, 110424. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Botstein, D.; Brown, P. Supervised harvesting of expression trees. Genome Biol. 2001, 2, RESEARCH0003. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.; Tibshirani, R. Generalized Additive Models: Some Applications. J. Am. Stat. Assoc. 1987, 82, 371–386. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Edwards, T.C.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R. Generalized additive models for medical research. Stat. Methods Med. Res. 1995, 4, 187–196. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Wang, Z.; Fan, B. Prediction of potential suitable growth areas for Chinese jujube under the background of climate change. Sci. Silvae Sin. 2021, 57, 158–168. [Google Scholar] [CrossRef]
- Luo, M.; Wang, H.; Lv, Z. Evaluating the Performance of Biomod2 and MaxEnt Distribution Prediction Models Using Giant Panda Data. Chin. J. Appl. Ecol. 2017, 28, 4001–4006. [Google Scholar] [CrossRef]
- Guo, L.F.; Gao, Y.; He, P.; He, Y.; Meng, F.Y. Modeling for Predicting the Potential Geographical Distribution of Three Ephedra Herbs in China. Plants-Basel 2023, 12, 787. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.Q.; Zhang, M.X.; Chen, Y.; Wang, A.X.; Li, M.H. Applying Biomod2 for modeling of species suitable habitats:a case study of Paeonia lactiflora in China. Zhongguo Zhong Yao Za Zhi. 2022, 47, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Qian, Y.; Wang, G. Characteristics and Existing Problems of Hongze Lake Wetland Ecosystem. Environ. Sci. Technol. 2010, 33, 1–5. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, K.; Sun, L.; Zhang, J.; Li, S.; Hu, Z.; Wang, R.; Li, X.; Xiong, Q.; Jie, W. Effects of Trifolium repens invasion on soil animals in urban lawn ecosystems. J. Ecol. 2018, 38, 8489–8499. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, C.; Fu, J.; Liu, L.; Li, X. Effects of Aspergillus spinosus on the quality and yield of perennial ryegrass forage. J. Ecol. 2019, 38, 3667–3672. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Liu, Y.; Yangfu, L.; Zhou, J. Effect of Propionic Acid on the Silage Quality and Microbial Flora of Hybrid Pennisetum. J. Anim. Nutr. 2023, 35, 5404–5417. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, P.; Fu, Y.; Wei, S. Effect of Tamarix shrub grass community on surface soil improvement in mudflat of Binzhou Port. Wetl. Sci. 2023, 21, 541–548. [Google Scholar] [CrossRef]
- Shao, T.; Yan, X.; Jia, B.; Xu, L.; Ma, Z.; Ren, Y.; Long, X. Study on the Improvement of Coastal Viscous Saline-alkali Land by Planting Herbal Plant Dogtooth Root. Soils 2023, 25, 1–7. Available online: https://kns.cnki.net/kcms2/detail/32.1118.P.20230629.0918.002.html (accessed on 9 January 2023).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation | Iris pseudacorus L. | Ceratophyllum demeser L. | Hydrilla verticillata (Linn. f.) Royle | Potamogeton wrightii Morong |
|---|---|---|---|---|
| Number of occurrences | 246 | 234 | 222 | 186 |
| Vegetation | Cynodon typist (L.) Persoon | Lolium perenne L. | Phragmites australis (Cav.) Trin. ex steu | Polygonum hydropiper L. |
| Number of occurrences | 352 | 164 | 1084 | 454 |
| Vegetation | Lagerstroemia indica L. | Amorpha fruticosa Linn | Tamarix chinensis Lour | Trifolium repens L. |
| Number of occurrences | 163 | 550 | 257 | 602 |
| Vegetation | Canna indica L. | Nelumbo nucifera Gaertn | Typha angustifolia L. | Nymphoides peltata (S. G. Gmelin) Kuntze |
| Number of occurrences | 250 | 191 | 379 | 444 |
| Vegetation | Nandina domestica Thunb | Triarrhena sacchariflora (Maxim.) Nakai | Pennisetum alopecuroides (L.) Spreng | Potamogeton crispus L. |
| Number of occurrences | 126 | 249 | 509 | 335 |
| Data sources | on-the-spot investigation + (GBIF, http://www.gbif.org, accessed on 13 February 2023), (CVH, http://www.cvh.org/, accessed on 13 February 2023), (http://www.nsii.org/, accessed on 15 February 2023) | |||
| Environment Variables | Variable Description |
|---|---|
| Bio1 | Annual Mean Temperature (°C) |
| Bio2 | Mean diurnal range (mean of monthly (max temp–min temp)) daily average temperature range (°C). |
| Bio3 | Isothermicity (Bio 2/Bio 7) (×100) Isothermicity |
| Bio4 | Temperature seasonality (standard deviation × 100) coefficient of seasonal variation of temperature. |
| Bio5 | Max Temperature of Warmest Month Maximum temperature in hottest month (°C) |
| Bio6 | Min Temperature of Coldest Month Minimum temperature in coldest month (°C) |
| Bio7 | Temperature annual range (Bio 5–Bio 6) Temperature annual range (°C) |
| Bio8 | Mean temperature of wet quarter (°C) |
| Bio9 | Mean Temperature of Driest Quarter Average temperature in the driest quarter (°C) |
| Bio10 | Mean temperature of warm quarter average temperature in the warmest quarter (°C) |
| Bio11 | Mean temperature of coldest quarter average temperature (°C) |
| Bio12 | Annual Precipitation (mm) |
| Bio13 | Precipitation of Wettest Month in the wettest month (mm) |
| Bio14 | Precipitation of Driest Month (mm) |
| Bio15 | Seasonal variation of precipitation (coefficient of variation) (mm) |
| Bio16 | Precipitation in the driest quarter of precision of wet quarter (mm) |
| Bio17 | Precipitation in the wettest quarter of precision of drill quarter (mm) |
| Bio18 | The warmest quarterly precipitation of Precipitation of Warmest Quarter (mm) |
| Bio19 | The coldest quarterly precipitation of Precipitation of Coldest Quarter (mm) |
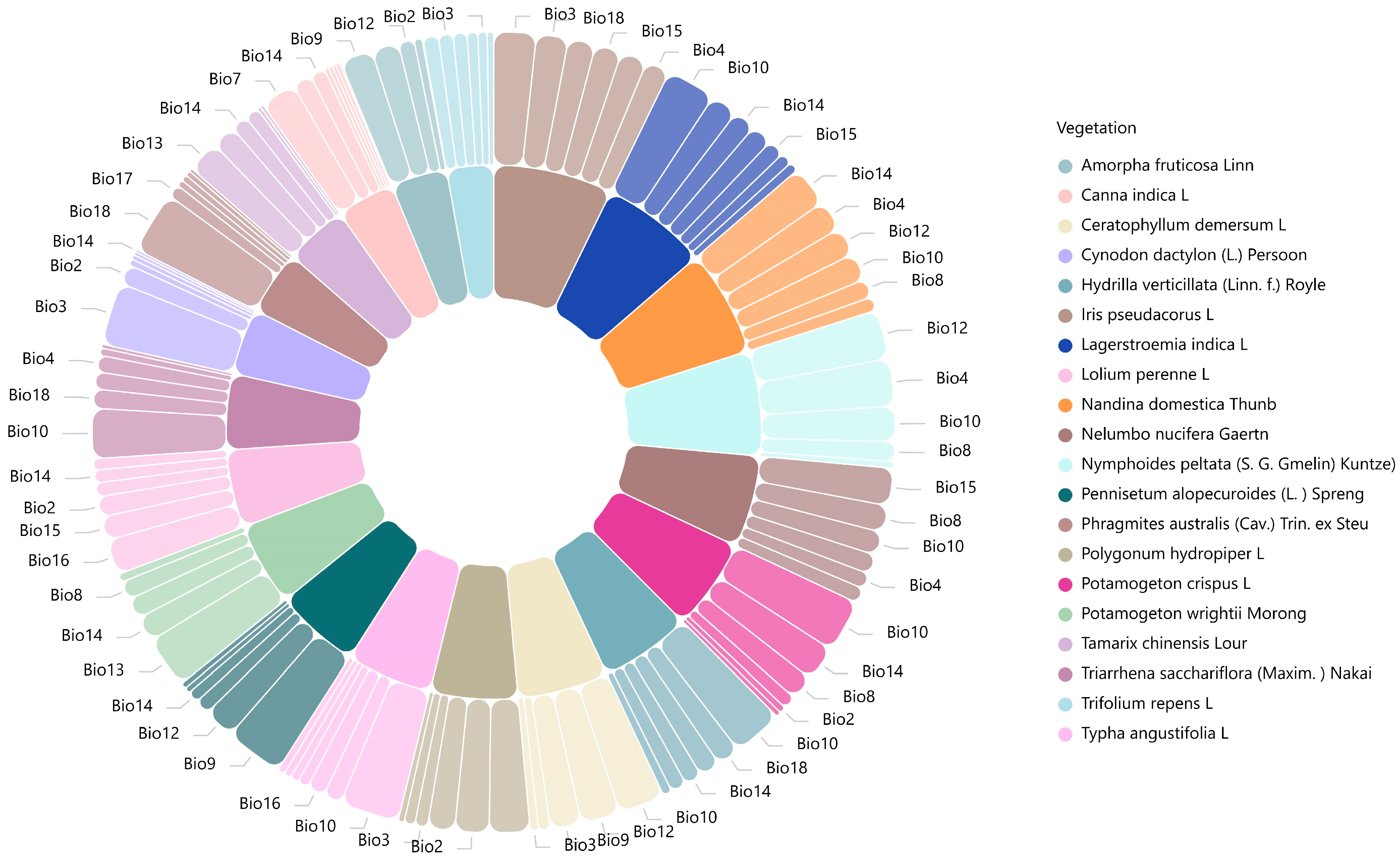
| Vegetation Name | Bio1 | Bio2 | Bio3 | Bio4 | Bio5 | Bio6 | Bio7 | Bio8 | Bio9 | Bio10 | Bio11 | Bio12 | Bio13 | Bio14 | Bio15 | Bio16 | Bio17 | Bio18 | Bio19 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bio | ||||||||||||||||||||
| Iris pseudacorus L. | 47.7 | 27.5 | 29.9 | 30.8 | 30.1 | 36 | ||||||||||||||
| Ceratophyllum demersum L. | 9.3 | 35.1 | 52.3 | 44.1 | 13.3 | |||||||||||||||
| Hydrilla verticillata (Linn. f.) Royle | 23.7 | 10.2 | 17.8 | 57.1 | 19.5 | 26.3 | ||||||||||||||
| Potamogeton wrightii Morong | 23.5 | 9.4 | 20.9 | 58.7 | 28.6 | |||||||||||||||
| Nandina domestica Thunb | 15.6 | 34.7 | 21.1 | 31.2 | 32.3 | 45.6 | ||||||||||||||
| Lagerstroemia indica L. | 13.6 | 10.7 | 24.6 | 56.2 | 25.4 | 19.1 | 31 | |||||||||||||
| Amorpha fruticosa Linn | 0.9 | 36.8 | 30.6 | 17.8 | 9 | |||||||||||||||
| Tamarix chinensis Lour | 27.5 | 3.7 | 4.1 | 36.6 | 18.9 | 16 | ||||||||||||||
| Trifolium repens L. | 16.4 | 10.5 | 7.4 | 17.6 | 12.1 | 15.6 | ||||||||||||||
| Pennisetum alopecuroides (L.) Spreng | 6.5 | 16.9 | 66.1 | 35.4 | 6.5 | 13 | ||||||||||||||
| Cynodon dactylon (L.) Persoon | 22.6 | 71 | 7 | 3.8 | 3 | 5.3 | ||||||||||||||
| Lolium perenne L. | 22.8 | 12.8 | 15.7 | 14.3 | 26.8 | 39.9 | ||||||||||||||
| Phragmitesaustralis (Cav.) Trin. ex steu | 7.2 | 3.8 | 7.4 | 6 | 14.7 | 70.2 | ||||||||||||||
| Polygonum hydropiper L. | 38.6 | 14.4 | 6.9 | 45.8 | 12.1 | 30.6 | ||||||||||||||
| Nelumbo nucifera Gaertn | 4.3 | 18.7 | 8.3 | 56.6 | 19.2 | 21.7 | ||||||||||||||
| Triarrhena sacchariflora (Maxim) Nakai | 4.2 | 4.2 | 40 | 4.6 | 21.8 | 4.9 | 17.2 | 2 | ||||||||||||
| Canna indica L. | 17.3 | 17.7 | 30.5 | 26.4 | 42.2 | 22.1 | ||||||||||||||
| Typha angustifolia L. | 8.4 | 9.4 | 20.6 | 66.9 | 12.3 | 8.1 | 20.2 | |||||||||||||
| Nymphoides peltata (S. G. Gmelin) Kuntze | 7.3 | 53.5 | 22.6 | 39.7 | 55.7 | |||||||||||||||
| Potamogeton crispus L. | 7.5 | 5.7 | 27.9 | 59.4 | 39.7 | 14.9 | ||||||||||||||
| Vegetation Name | Change Range of Suitable Area | |||
|---|---|---|---|---|
| SSP126 | SSP245 | SSP370 | SSP585 | |
| Iris pseudacorus L. | −2~2.9 | −2~3.0 | −2~2.9 | −2~3.1 |
| Ceratophyllum demersum L. | 8.7~10.6 | 8.9~11.9 | 8.3~10.4 | 7.9~11.0 |
| Hydrilla verticillate (Linn. f.) Royle | 4.7~7.4 | 4.0~7.3 | 5.0~7.6 | 4.7~7.8 |
| Potamogeton wrightii Morong | 10.2~18.2 | 12.9~19.3 | 9.2~16.4 | 8.8~18.5 |
| Nandina domestica Thunb | 5.9~10.8 | 5.7~8.9 | 5.5~10.7 | 4.5~9.8 |
| Lagerstroemia indica L. | 17.6~24.4 | 16.2~24.2 | 18.1~24.8 | 16.7~24.0 |
| Amorpha fruticose Linn | 53.7~65.9 | 55.6~65.9 | 53.5~65.7 | 51.7~64.5 |
| Tamarix chinensis Lour | 42.2~46.8 | 41.5~46.4 | 44.8~48.8 | 44.3~49.2 |
| Trifolium repens L. | 35.5~44.4 | 35.3~48.2 | 35.4~44.4 | 37.7~45.8 |
| Pennisetum alopecuroides (L.) Spreng | 11.0~18.0 | 4.7~18.2 | 10.4~17.8 | 3.7~16.9 |
| Cynodon dactylon (L.) Pers | 46.3~48.9 | 46.2~49.0 | 46.6~49.4 | 47.2~49.8 |
| Lolium perenne L. | 34.3~37.9 | 35.0~38.0 | 33.3~38.3 | 30.9~37.4 |
| Phragmites australis (Cav.) Trin. ex steu | 0.2~12.6 | 0.3~13.8 | 0.0~12.6 | 0.0~12.6 |
| Polygonum hydropiper L. | 22.3~28.1 | 22.3~28.1 | 13.3~27.4 | 12.9~28.0 |
| Nelumbo nucifera Gaertn | −10.0~−2.5 | −9.8~−2.5 | −10.3~−2.5 | −10.2~−2.8 |
| Triarrhena sacchariflora (Maxim.) Nakai | 4.2~7.4 | 4.2~6.5 | 4.2~6.5 | 4.0~7.4 |
| Canna indica L. | 72.8~73.3 | 73.3~74.6 | 72.6~73.4 | 72.7~73.5 |
| Typha angustifolia L. | 5.8~9.8 | 5.9~10.3 | 6.2~10.0 | 6.5~10.0 |
| Nymphoides peltate (S. G. Gmelin) Kuntze | 0.4~1.0 | 0.5~1.3 | 0.3~1.0 | 0.3~1.0 |
| Potamogeton crispus L. | 22.7~26.8 | 25.2~28.1 | 22.8~26.1 | 22.9~25.8 |
| Vegetation SSPs | Future (~2040s) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | SSP126 | SSP 245 | SSP 370 | SSP 585 | |||||||||||||
| Classification of Suitable Habitat | Low | Medium | High | Most | Low | Medium | High | Most | Low | Medium | High | Most | Low | Medium | High | Most | |
| Trifolium repens L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Tamarix chinensis Lour | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Triarrhena sacchariflora (Maxim.) Nakai | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Cynodon dactylon (L.) Persoon | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Hydrilla verticillata (Linn. f.) Royle | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Ceratophyllum demersum L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Pennisetum alopecuroides (L.) Spreng | 12.42 | 23.42 | 14.12 | 21.72 | 13.97 | 21.87 | 20.77 | 15.07 | |||||||||
| Phragmites australis (Cav.) Trin. ex steu | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Canna indica L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Nandina domestica Thunb | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Typha angustifolia L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Amorpha fruticosa Linn | 1.04 | 34.78 | 0.66 | 35.18 | 0.91 | 34.93 | 18.33 | 17.51 | |||||||||
| Potamogeton crispus L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Lolium perenne L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Iris pseudacorus L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Nelumbo nucifera Gaertn | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Potamogeton wrightii Morong | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Polygonum hydropiper L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Nymphoides peltata (S. G. Gmelin) Kuntze | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
| Lagerstroemia indica L. | 35.84 | 35.84 | 35.84 | 35.84 | |||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Cheng, J.; Jiang, Y.; Liu, N.; Wang, K. Prediction of Adaptability of Typical Vegetation Species in Flood Storage Areas under Future Climate Change: A Case in Hongze Lake FDZ, China. Sustainability 2024, 16, 6331. https://doi.org/10.3390/su16156331
Wang L, Cheng J, Jiang Y, Liu N, Wang K. Prediction of Adaptability of Typical Vegetation Species in Flood Storage Areas under Future Climate Change: A Case in Hongze Lake FDZ, China. Sustainability. 2024; 16(15):6331. https://doi.org/10.3390/su16156331
Chicago/Turabian StyleWang, Liang, Jilin Cheng, Yushan Jiang, Nian Liu, and Kai Wang. 2024. "Prediction of Adaptability of Typical Vegetation Species in Flood Storage Areas under Future Climate Change: A Case in Hongze Lake FDZ, China" Sustainability 16, no. 15: 6331. https://doi.org/10.3390/su16156331