
A Study of the Effect of Biochar Additive on the Manure–Compost–Soil Process and Its Bacterial Succession

Abstract
1. Introduction
2. Materials and Methods
2.1. Aerobic Composting and Cucumber Potting Experiment
2.2. Basic Physical and Chemical Indicators
2.3. Microbial Diversity Determination and Analysis
2.3.1. DNA Extraction and Purification
2.3.2. High-Throughput Sequencing
2.4. Statistics and Data Analysis
3. Results and Discussion
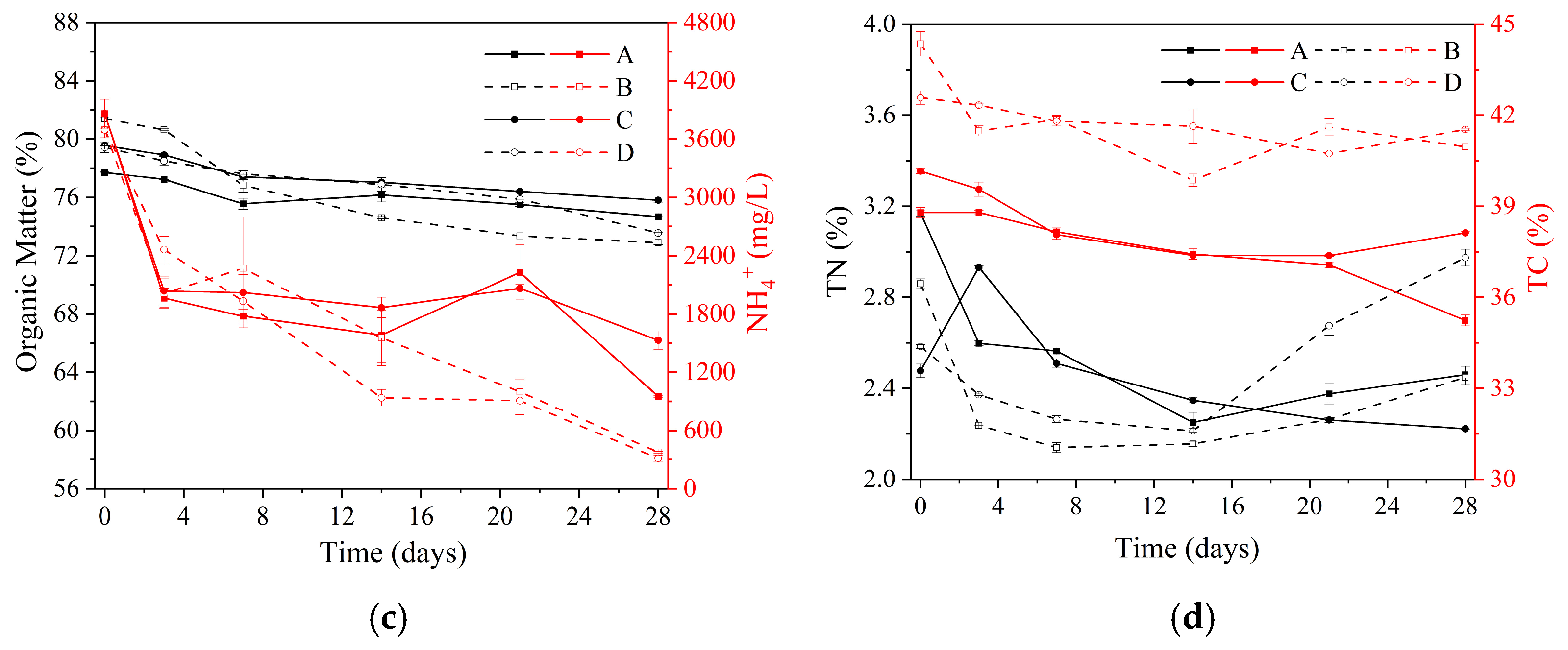
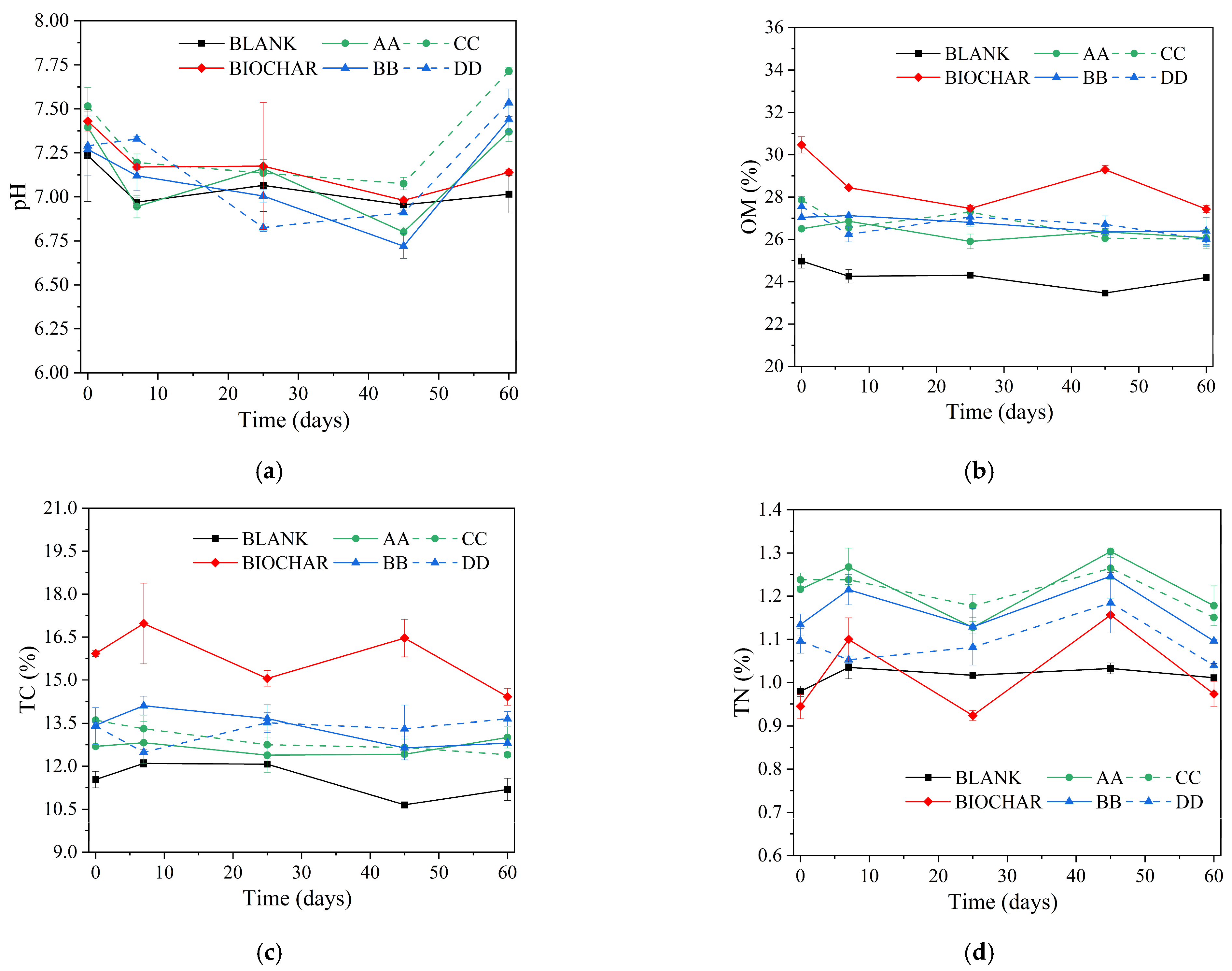
3.1. Physicochemical Changes in the Manure–Compost–Soil Process
3.1.1. Manure–Compost during Aerobic Composting
3.1.2. Compost–Soil in the Potting Experiment
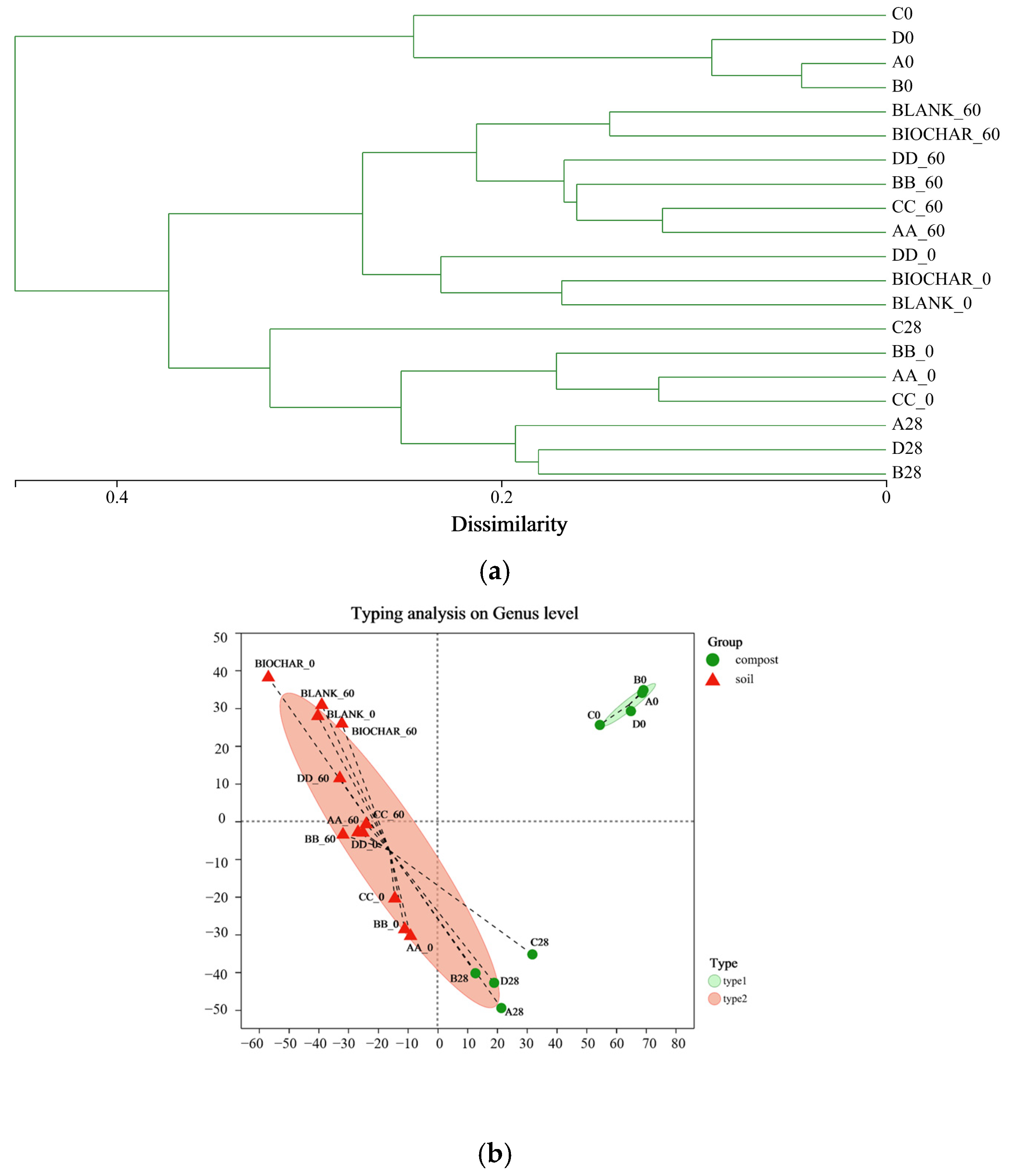
3.2. Analysis of the Bacterial Diversity in the Manure–Compost–Soil Process
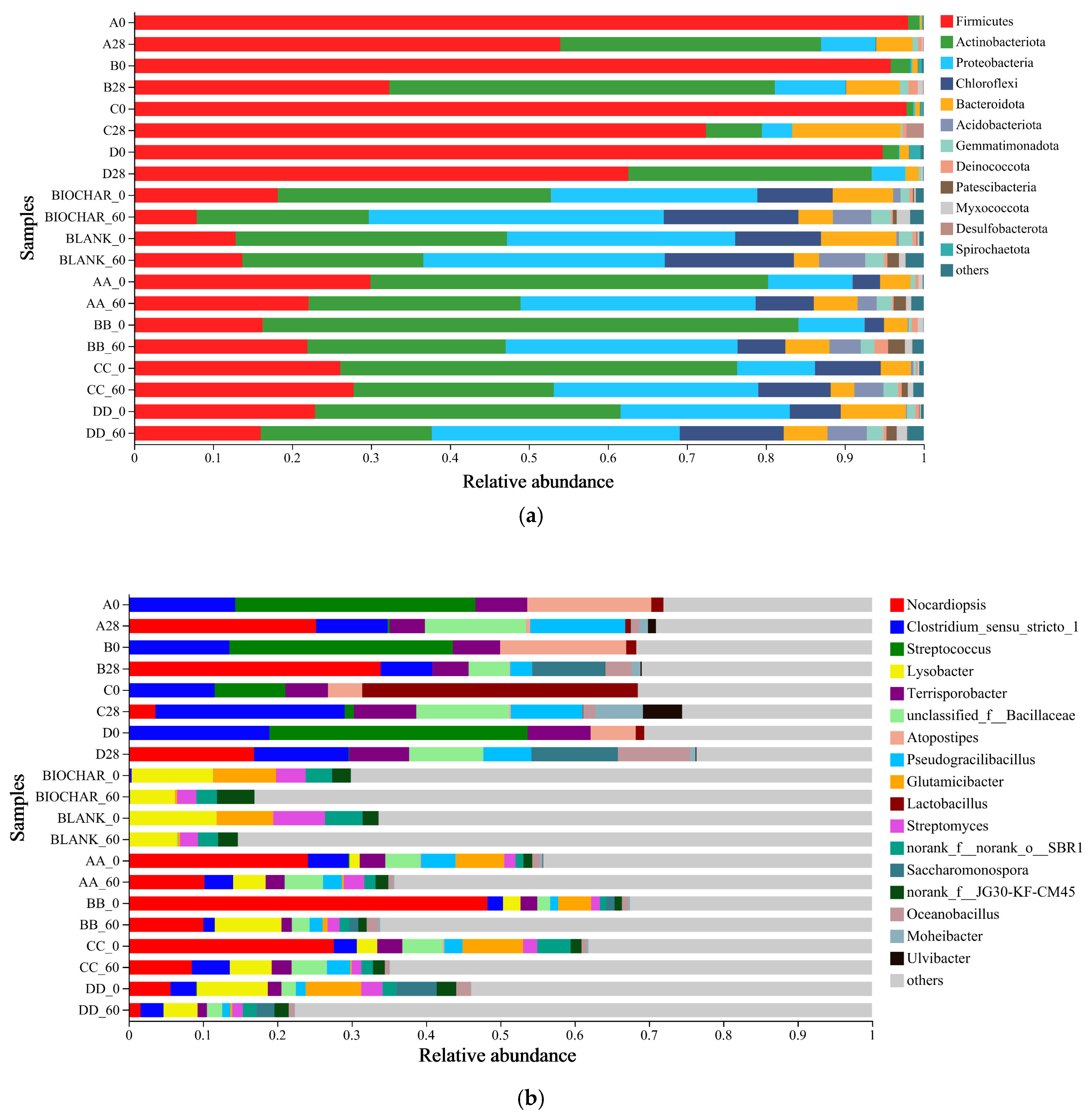
3.3. Bacterial Community Composition in the Manure–Compost–Soil Process
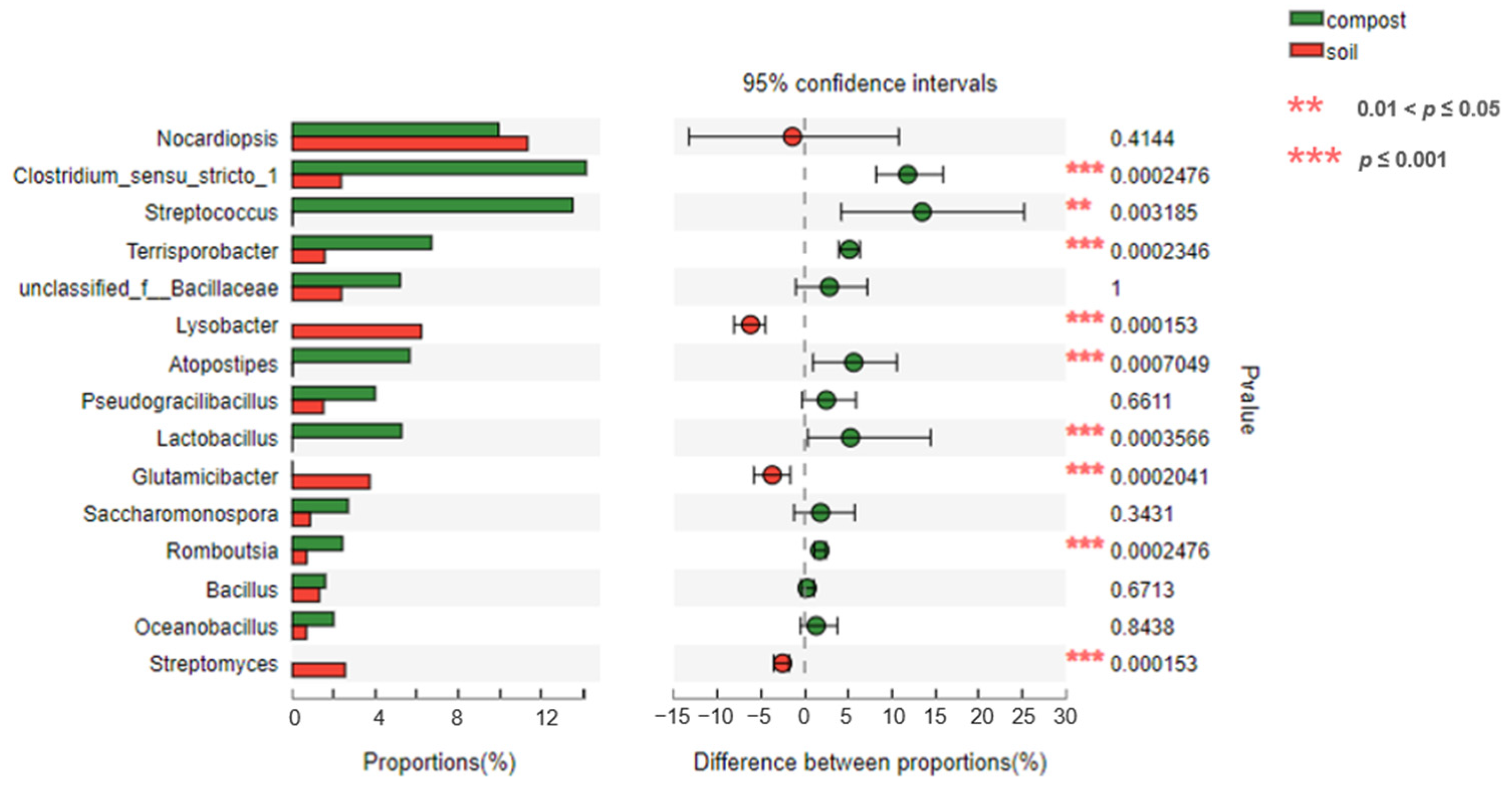
3.4. Difference Analysis of Bacteria from Pig Manure–Compost to Compost–Soil
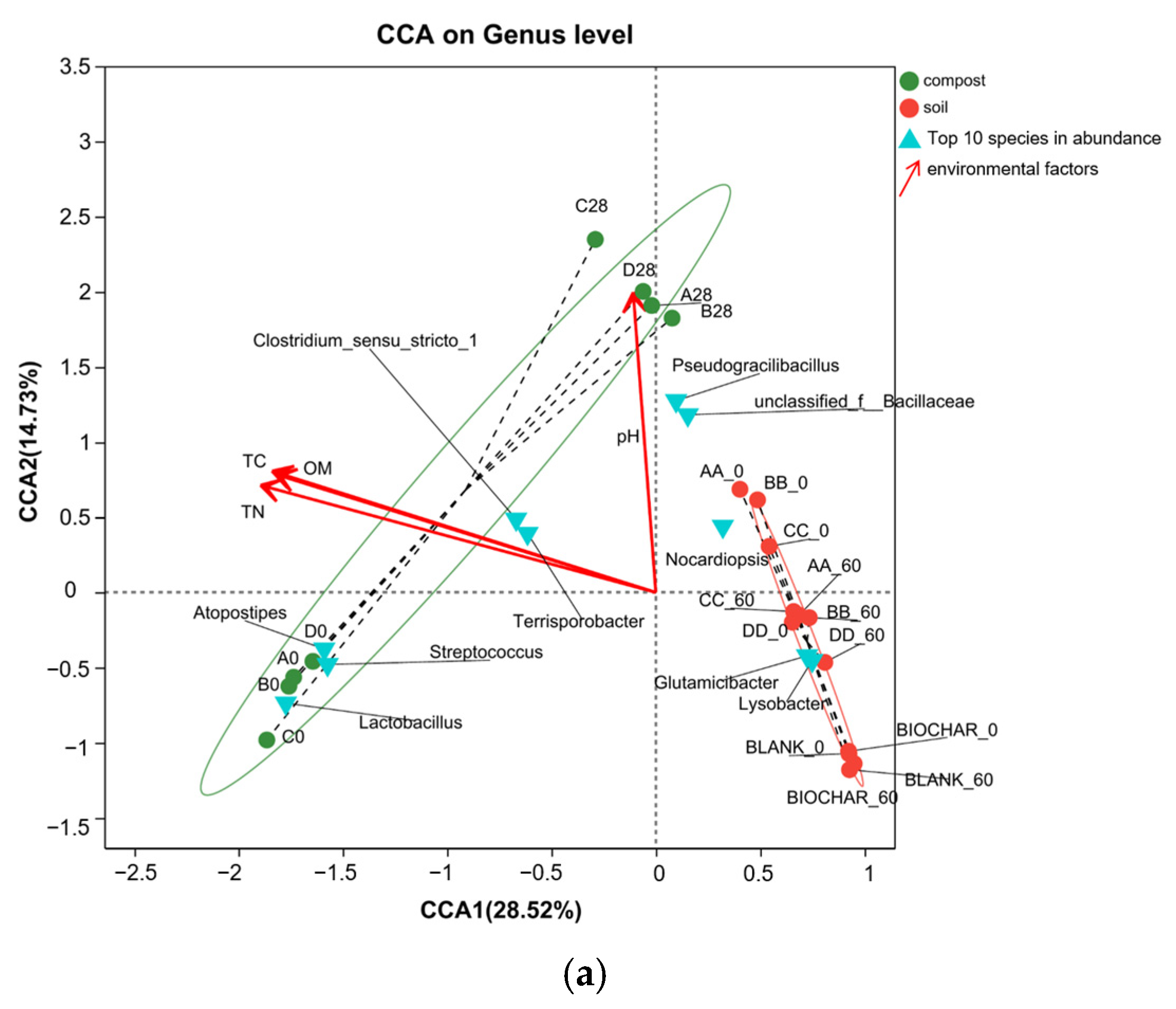
3.5. Correlation Analysis of Bacteria and Physicochemical Properties in the Manure–Compost–Soil Process
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| Properties | abbreviations |
| moisture content, % | MC |
| organic matter, % | OM |
| total carbon, % | TC |
| total nitrogen, % | TN |
| Initial aerobic composting samples on Day 0 of A | A0 |
| Initial aerobic composting samples on Day 0 of B | B0 |
| Initial aerobic composting samples on Day 0 of C | C0 |
| Initial aerobic composting samples on Day 0 of D | D0 |
| Final aerobic composting samples on Day 28 of A | A28 |
| Final aerobic composting samples on Day 28 of B | B28 |
| Final aerobic composting samples on Day 28 of C | C28 |
| Final aerobic composting samples on Day 28 of D | D28 |
| Initial soil samples of potting experiment on Day 0 of BLANK | BLANK_0 |
| Initial soil samples of potting experiment on Day 0 of BIOCHAR | BIOCHAR_0 |
| Initial soil samples of potting experiment on Day 0 of AA | AA_0 |
| Initial soil samples of potting experiment on Day 0 of BB | BB_0 |
| Initial soil samples of potting experiment on Day 0 of CC | CC_0 |
| Initial soil samples of potting experiment on Day 0 of DD | DD_0 |
| Final soil samples of potting experiment on Day 60 of BLANK | BLANK_60 |
| Final soil samples of potting experiment on Day 60 of BIOCHAR | BIOCHAR_60 |
| Final soil samples of potting experiment on Day 60 of AA | AA_60 |
| Final soil samples of potting experiment on Day 60 of BB | BB_60 |
| Final soil samples of potting experiment on Day 60 of CC | CC_60 |
| Final soil samples of potting experiment on Day 60 of DD | DD_60 |
References
- Ma, S.; Sun, X.; Fang, C.; He, X.; Han, L.; Huang, G. Exploring the mechanisms of decreased methane during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane. Waste Manag. 2018, 78, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, B.; Nguyen, D.D.; Chaudhary, D.K.; Chang, S.W.; Kim, J.; Lee, S.R.; Shin, J.; Jeon, B.; Chung, S.; Lee, J. Influence of biochar on physico-chemical and microbial community during swine manure composting process. J. Environ. Manag. 2019, 232, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Haug, R. The Practical Handbook of Compost Engineering; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Zhou, S.; Chang, T.; Zhang, Y.; Shaghaleh, H.; Zhang, J.; Yang, X.; Qin, H.; Talpur, M.M.A.; Alhaj Hamoud, Y. Organic fertilizer compost alters the microbial composition and network structure in strongly acidic soil. Appl. Soil. Ecol. 2024, 195, 105263. [Google Scholar] [CrossRef]
- Morra, L.; Bilotto, M.; Baldantoni, D.; Alfani, A.; Baiano, S. A seven-year experiment in a vegetable crops sequence: Effects of replacing mineral fertilizers with Biowaste compost on crop productivity, soil organic carbon and nitrates concentrations. Sci. Hortic. 2021, 290, 110534. [Google Scholar] [CrossRef]
- Scotti, R.; Pane, C.; Spaccini, R.; Palese, A.M.; Piccolo, A.; Celano, G.; Zaccardelli, M. On-farm compost: A useful tool to improve soil quality under intensive farming systems. Appl. Soil. Ecol. 2016, 107, 13–23. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Zhang, Z.; Wang, Q.; Shen, F.; Li, R.; Li, D.; Ren, X.; Wang, M.; Chen, H.; Zhao, J. New insight with the effects of biochar amendment on bacterial diversity as indicators of biomarkers support the thermophilic phase during sewage sludge composting. Bioresour. Technol. 2017, 238, 589–601. [Google Scholar] [CrossRef]
- He, X.; Yin, H.; Han, L.; Cui, R.; Fang, C.; Huang, G. Effects of biochar size and type on gaseous emissions during pig manure/wheat straw aerobic composting: Insights into multivariate-microscale characterization and microbial mechanism. Bioresour. Technol. 2019, 271, 375–382. [Google Scholar] [CrossRef]
- Ma, S.; Fang, C.; Sun, X.; Han, L.; He, X.; Huang, G. Bacterial community succession during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane under slight positive pressure. Bioresour. Technol. 2018, 259, 221–227. [Google Scholar] [CrossRef]
- Babin, D.; Leoni, C.; Neal, A.L.; Sessitsch, A.; Smalla, K. Editorial to the Thematic Topic “Towards a more sustainable agriculture through managing soil microbiomes”. FEMS Microbiol. Ecol. 2021, 97, fiab094. [Google Scholar] [CrossRef]
- Fierer, N.; Wood, S.A.; Bueno De Mesquita, C.P. How microbes can, and cannot, be used to assess soil health. Soil. Biol. Biochem. 2021, 153, 108111. [Google Scholar] [CrossRef]
- Maron, P.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Karimi, B.; Bernard, L.; Dequiedt, S.; Terrat, S.; et al. High Microbial Diversity Promotes Soil Ecosystem Functioning. Appl. Environ. Microbiol. 2018, 84, e02738-17. [Google Scholar] [CrossRef] [PubMed]
- Bol, R.; Amelung, W.; Friedrich, C.; Ostle, N. Tracing dung-derived carbon in temperate grassland using 13C natural abundance measurements. Soil. Biol. Biochem. 2000, 32, 1337–1343. [Google Scholar] [CrossRef]
- Duan, M.; Li, H.; Gu, J.; Tuo, X.; Sun, W.; Qian, X.; Wang, X. Effects of biochar on reducing the abundance of oxytetracycline, antibiotic resistance genes, and human pathogenic bacteria in soil and lettuce. Environ. Pollut. 2017, 224, 787–795. [Google Scholar] [CrossRef]
- Mawof, A.; Prasher, S.; Bayen, S.; Nzediegwu, C. Effects of Biochar and Biochar-Compost Mix as Soil Amendments on Soil Quality and Yield of Potatoes Irrigated with Wastewater. J. Soil. Sci. Plant Nutr. 2021, 21, 2600–2612. [Google Scholar] [CrossRef]
- Purakayastha, T.J.; Bera, T.; Bhaduri, D.; Sarkar, B.; Mandal, S.; Wade, P.; Kumari, S.; Biswas, S.; Menon, M.; Pathak, H.; et al. A review on biochar modulated soil condition improvements and nutrient dynamics concerning crop yields: Pathways to climate change mitigation and global food security. Chemosphere 2019, 227, 345–365. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.K.; Lin, C.; Hoang, H.G.; Sanderson, P.; Dang, B.T.; Bui, X.T.; Nguyen, N.S.H.; Vo, D.N.; Tran, H.T. Evaluate the role of biochar during the organic waste composting process: A critical review. Chemosphere 2022, 299, 134488. [Google Scholar] [CrossRef]
- Sanchez-Monedero, M.A.; Cayuela, M.L.; Roig, A.; Jindo, K.; Mondini, C.; Bolan, N. Role of biochar as an additive in organic waste composting. Bioresour. Technol. 2018, 247, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl. Soil. Ecol. 2017, 119, 156–170. [Google Scholar] [CrossRef]
- Fu, T.; Shangguan, H.; Wu, J.; Tang, J.; Yuan, H.; Zhou, S. Insight into the synergistic effects of conductive biochar for accelerating maturation during electric field-assisted aerobic composting. Bioresour. Technol. 2021, 337, 125359. [Google Scholar] [CrossRef]
- Guo, X.; Liu, H.; Zhang, J. The role of biochar in organic waste composting and soil improvement: A review. Waste Manag. 2020, 102, 884–899. [Google Scholar] [CrossRef]
- Kung, C.; Mu, J.E. Prospect of China’s renewable energy development from pyrolysis and biochar applications under climate change. Renew. Sustain. Energy Rev. 2019, 114, 109343. [Google Scholar] [CrossRef]
- Sun, X.; Ma, S.; Han, L.; Li, R.; Schlick, U.; Chen, P.; Huang, G. The effect of a semi-permeable membrane-covered composting system on greenhouse gas and ammonia emissions in the Tibetan Plateau. J. Clean. Prod. 2018, 204, 778–787. [Google Scholar] [CrossRef]
- Xiao, L.; Feng, L.R.; Yuan, G.D.; Wei, J. Low-cost field production of biochars and their properties. Environ. Geochem. Health 2020, 42, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Tao, X.; Du, Y.; Li, M.; Yang, S.; Zhang, W.; Yang, C.; Li, H.; Wang, X.; Chen, R. Biochar improves the humification process during pig manure composting: Insights into roles of the bacterial community and metabolic functions. J. Environ. Manag. 2024, 355, 120463. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.C.; Wang, H.Y.; Gai, X.P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil. Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, J.; Han, L.; Ma, S.; Sun, X.; Huang, G. Role and multi-scale characterization of bamboo biochar during poultry manure aerobic composting. Bioresour. Technol. 2017, 241, 190–199. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yin, H.; Fang, C.; Xiong, J.; Han, L.; Yang, Z.; Huang, G. Metagenomic and q-PCR analysis reveals the effect of powder bamboo biochar on nitrous oxide and ammonia emissions during aerobic composting. Bioresour. Technol. 2021, 323, 124567. [Google Scholar] [CrossRef]
- He, X.; Chen, L.; Han, L.; Liu, N.; Cui, R.; Yin, H.; Huang, G. Evaluation of biochar powder on oxygen supply efficiency and global warming potential during mainstream large-scale aerobic composting. Bioresour. Technol. 2017, 245, 309–317. [Google Scholar] [CrossRef]
- Xiong, J.; Ma, S.; He, X.; Han, L.; Huang, G. Nitrogen transformation and dynamic changes in related functional genes during functional-membrane covered aerobic composting. Bioresour. Technol. 2021, 332, 125087. [Google Scholar] [CrossRef]
- Fang, C.; Yuan, X.; Liao, K.; Qu, H.; Han, L.; He, X.; Huang, G. Micro-aerobic conditions based on membrane-covered improves the quality of compost products: Insights into fungal community evolution and dissolved organic matter characteristics. Bioresour. Technol. 2022, 362, 127849. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and Experimental Application of a Novel Non-Degenerate Universal Primer Set that Amplifies Prokaryotic 16S rRNA Genes with a Low Possibility to Amplify Eukaryotic rRNA Genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- López-Cano, I.; Roig, A.; Cayuela, M.L.; Alburquerque, J.A.; Sánchez-Monedero, M.A. Biochar improves N cycling during composting of olive mill wastes and sheep manure. Waste Manag. 2016, 49, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, M.; Alburquerque, J.A.; Sánchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. Biochar accelerates organic matter degradation and enhances N mineralisation during composting of poultry manure without a relevant impact on gas emissions. Bioresour. Technol. 2015, 192, 272–279. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresour. Technol. 2018, 247, 190–199. [Google Scholar] [CrossRef]
- Xie, T.; Zhang, Z.; Zhang, D.; Wei, C.; Lin, Y.; Feng, R.; Nan, J.; Feng, Y. Effect of hydrothermal pretreatment and compound microbial agents on compost maturity and gaseous emissions during aerobic composting of kitchen waste. Sci. Total Environ. 2023, 854, 158712. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y. Elimination of phytotoxicity during co-composting of spent pig-manure sawdust litter and pig sludge. Bioresour. Technol. 1998, 65, 43–49. [Google Scholar] [CrossRef]
- Kong, Y.; Zhang, J.; Yang, Y.; Liu, Y.; Zhang, L.; Wang, G.; Liu, G.; Dang, R.; Li, G.; Yuan, J. Determining the extraction conditions and phytotoxicity threshold for compost maturity evaluation using the seed germination index method. Waste Manag. 2023, 171, 502–511. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresour. Technol. 2014, 171, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Ottani, F.; Parenti, M.; Santunione, G.; Moscatelli, G.; Kahn, R.; Pedrazzi, S.; Allesina, G. Effects of different gasification biochar grain size on greenhouse gases and ammonia emissions in municipal aerated composting processes. J. Environ. Manag. 2023, 331, 117257. [Google Scholar] [CrossRef]
- Yu, H.; Xie, B.; Khan, R.; Shen, G. The changes in carbon, nitrogen components and humic substances during organic-inorganic aerobic co-composting. Bioresour. Technol. 2019, 271, 228–235. [Google Scholar] [CrossRef]
- Naeem, M.A.; Khalid, M.; Aon, M.; Abbas, G.; Amjad, M.; Murtaza, B.; Khan, W.; Ahmad, N. Combined application of biochar with compost and fertilizer improves soil properties and grain yield of maize. J. Plant Nutr. 2018, 41, 112–122. [Google Scholar] [CrossRef]
- Liu, H.; Awasthi, M.K.; Zhang, Z.; Syed, A.; Bahkali, A.H. Evaluation of gases emission and enzyme dynamics in sheep manure compost occupying with peach shell biochar. Environ. Pollut. 2024, 351, 124065. [Google Scholar] [CrossRef] [PubMed]
- Aina Najwa Mohd Nor Azman, N.; Inderakusumowati Md Khalid, P.; Aishah Saidina Amin, N.; Yamani Zakaria, Z.; Mohammad Zainol, M.; Ilham, Z.; Phaiboonsilpa, N.; Asmadi, M. Effects of biochar, compost, and composted biochar soil amendments on okra plant growth. Mater. Today Proc. 2023; in press. [Google Scholar] [CrossRef]
- Neville, F.; Seyfaee, A. Real-Time Monitoring of in Situ Polyethyleneimine-Silica Particle Formation. Langmuir 2013, 29, 14681–14690. [Google Scholar] [CrossRef]
- He, X.; Han, L.; Fu, B.; Du, S.; Liu, Y.; Huang, G. Effect and microbial reaction mechanism of rice straw biochar on pore methane production during mainstream large-scale aerobic composting in China. J. Clean. Prod. 2019, 215, 1223–1232. [Google Scholar] [CrossRef]
- Mao, H.; Lv, Z.; Sun, H.; Li, R.; Zhai, B.; Wang, Z.; Awasthi, M.K.; Wang, Q.; Zhou, L. Improvement of biochar and bacterial powder addition on gaseous emission and bacterial community in pig manure compost. Bioresour. Technol. 2018, 258, 195–202. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.; Chen, F.; Zhang, T.; Kong, W. Preparation of two types plant biochars and application in soil quality improvement. Sci. Total Environ. 2024, 906, 167334. [Google Scholar] [CrossRef]
- Bennur, T.; Kumar, A.R.; Zinjarde, S.; Javdekar, V. Nocardiopsis species: Incidence, ecological roles and adaptations. Microbiol. Res. 2015, 174, 33–47. [Google Scholar] [CrossRef]
- Ibrahim, A.H.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S.; Gulder, T.A.M.; Abdelmohsen, U.R. Natural Product Potential of the Genus Nocardiopsis. Mar. Drugs 2018, 16, 147. [Google Scholar] [CrossRef]
- Jepsen, S.E.; Krause, M.; Gruttner, H. Reduction of Fecal Streptococcus and Salmonella by selected treatment methods for sludge and organic waste. Water Sci. Technol. 1997, 36, 203–210. [Google Scholar] [CrossRef]
- Li, X.; Shi, X.; Lu, M.; Zhao, Y.; Li, X.; Peng, H.; Guo, R. Succession of the bacterial community and functional characteristics during continuous thermophilic composting of dairy manure amended with recycled ceramsite. Bioresour. Technol. 2019, 294, 122044. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gu, J.; Wang, X.; Song, Z.; Dai, X.; Guo, H.; Yu, J.; Zhao, W.; Lei, L. Enhanced removal of antibiotic resistance genes and mobile genetic elements during swine manure composting inoculated with mature compost. J. Hazard Mater. 2021, 411, 125135. [Google Scholar] [CrossRef] [PubMed]
- Exposito, R.G.; Postma, J.; Raaijmakers, J.M.; De Bruijn, I. Diversity and Activity of Lysobacter Species from Disease Suppressive Soils. Front. Microbiol. 2015, 6, 1243. [Google Scholar]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic micro-organisms. J. Appl. Microbiol. 2018, 124, 15–27. [Google Scholar] [CrossRef]
- Qin, S.; Feng, W.; Zhang, Y.; Wang, T.; Xiong, Y.; Xing, K. Diversity of Bacterial Microbiota of Coastal Halophyte Limonium sinense and Amelioration of Salinity Stress Damage by Symbiotic Plant Growth-Promoting Actinobacterium Glutamicibacter halophytocola KLBMP 5180. Appl. Environ. Microb. 2018, 84, e01533-18. [Google Scholar] [CrossRef]
- Cordovez, V.; Carrion, V.J.; Etalo, D.W.; Mumm, R.; Zhu, H.; van Wezel, G.P.; Raaijmakers, J.M. Diversity and functions of volatile organic compounds produced by Streptomyces from a disease-suppressive soil. Front. Microbiol. 2015, 6, 1081. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Group Name | Ingredients a |
|---|---|---|
| Aerobic Composting | Control groups (A and C) | Swine manure and wheat straw (Ratio of 5:1) |
| Experimental groups (B and D) | Swine manure, wheat straw, and wheat straw biochar (Ratio of 10:2:1) | |
| Potting b | BLANK | 0.25 kg nutrient soil |
| BIOCHAR | 0.225 kg nutrient soil + 0.025 kg biochar | |
| AA | 0.225 kg nutrient soil + 0.025 kg compost from the Group A composting | |
| BB | 0.225 kg nutrient soil + 0.025 kg biochar-based compost from Group B | |
| CC | 0.225 kg nutrient soil + 0.025 kg compost from Group C | |
| DD | 0.225 kg nutrient soil + 0.025 kg biochar-based compost from Group D |
| Name | MC a | pH a | OM b | TC b | TN b |
|---|---|---|---|---|---|
| Pig manure | 66.00 ± 0.72 | / | 74.70 ± 0.01 | 37.41 ± 0.14 | 3.19 ± 0.01 |
| Wheat straw | 6.07 ± 0.14 | / | 89.81 ± 0.03 | 43.43 ± 0.09 | 0.99 ± 0.06 |
| Biochar | 3.21 ± 0.15 | 10.21 ± 0.07 | 80.99 ± 0.28 | 70.60 ± 0.16 | 1.08 ± 0.01 |
| Black soil | 2.71 ± 0.15 | 6.87 ± 0.01 | 4.51 ± 0.06 | 12.60 ± 0.24 | 1.12 ± 0.02 |
| Vermiculite | 5.62 ± 0.03 | 7.25 ± 0.01 | 25.05 ±0.68 | 0.16 ± 0.01 | 0.20 ± 0.01 |
| Compost A | 13.32 ± 0.06 | 7.96 ± 0.01 | 65.57 ± 0.07 | 33.36 ± 0.59 | 3.97 ± 0.06 |
| Compost C | 13.38 ±0.15 | 8.07 ± 0.03 | 65.30 ± 0.22 | 32.51 ± 0.02 | 3.86 ± 0.01 |
| Compost B | 10.80 ± 0.36 | 8.54 ± 0.04 | 67.98 ± 0.09 | 41.41 ± 0.15 | 3.34 ± 0.01 |
| Compost D | 10.31 ± 0.10 | 8.15 ± 0.03 | 71.47 ± 0.28 | 43.20 ± 0.06 | 2.67 ± 0.02 |
| Sample Name | Chao | Shannon | Simpson | Shannoneven | Simpsoneven |
|---|---|---|---|---|---|
| A0 | 252 | 3.314 | 0.111 | 0.599 | 0.036 |
| C0 | 243 | 3.131 | 0.155 | 0.570 | 0.026 |
| B0 | 294 | 3.542 | 0.098 | 0.623 | 0.035 |
| D0 | 260 | 3.273 | 0.128 | 0.589 | 0.030 |
| A28 | 199 | 3.840 | 0.060 | 0.725 | 0.084 |
| B28 | 136 | 3.231 | 0.123 | 0.658 | 0.060 |
| C28 | 268 | 4.092 | 0.058 | 0.732 | 0.064 |
| D28 | 145 | 3.430 | 0.068 | 0.689 | 0.101 |
| BLANK_0 | 635 | 5.449 | 0.014 | 0.844 | 0.114 |
| AA_0 | 351 | 4.466 | 0.045 | 0.762 | 0.063 |
| BB_0 | 278 | 3.372 | 0.199 | 0.599 | 0.018 |
| CC_0 | 330 | 4.515 | 0.031 | 0.778 | 0.096 |
| DD_0 | 436 | 4.967 | 0.019 | 0.817 | 0.121 |
| BIOCHAR_0 | 1035 | 5.739 | 0.014 | 0.827 | 0.070 |
| BLANK_60 | 896 | 6.259 | 0.004 | 0.921 | 0.298 |
| AA_60 | 761 | 5.830 | 0.008 | 0.879 | 0.171 |
| BB_60 | 856 | 5.771 | 0.014 | 0.855 | 0.083 |
| CC_60 | 661 | 5.739 | 0.008 | 0.884 | 0.179 |
| DD_60 | 987 | 6.288 | 0.004 | 0.912 | 0.243 |
| BIOCHAR_60 | 758 | 6.145 | 0.004 | 0.927 | 0.332 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; He, L.; Jiao, F.; Zhang, K.; He, X. A Study of the Effect of Biochar Additive on the Manure–Compost–Soil Process and Its Bacterial Succession. Sustainability 2024, 16, 7910. https://doi.org/10.3390/su16187910
He X, He L, Jiao F, Zhang K, He X. A Study of the Effect of Biochar Additive on the Manure–Compost–Soil Process and Its Bacterial Succession. Sustainability. 2024; 16(18):7910. https://doi.org/10.3390/su16187910
Chicago/Turabian StyleHe, Xiang, Li He, Feiyang Jiao, Kan Zhang, and Xueqin He. 2024. "A Study of the Effect of Biochar Additive on the Manure–Compost–Soil Process and Its Bacterial Succession" Sustainability 16, no. 18: 7910. https://doi.org/10.3390/su16187910
APA StyleHe, X., He, L., Jiao, F., Zhang, K., & He, X. (2024). A Study of the Effect of Biochar Additive on the Manure–Compost–Soil Process and Its Bacterial Succession. Sustainability, 16(18), 7910. https://doi.org/10.3390/su16187910