

Are European Trees Prepared for Being Limited by Other Nutrients than Nitrogen?


{kind=link}
{kind=link}
Abstract
1. Background
2. The Spatial Distribution of Available Nutrients
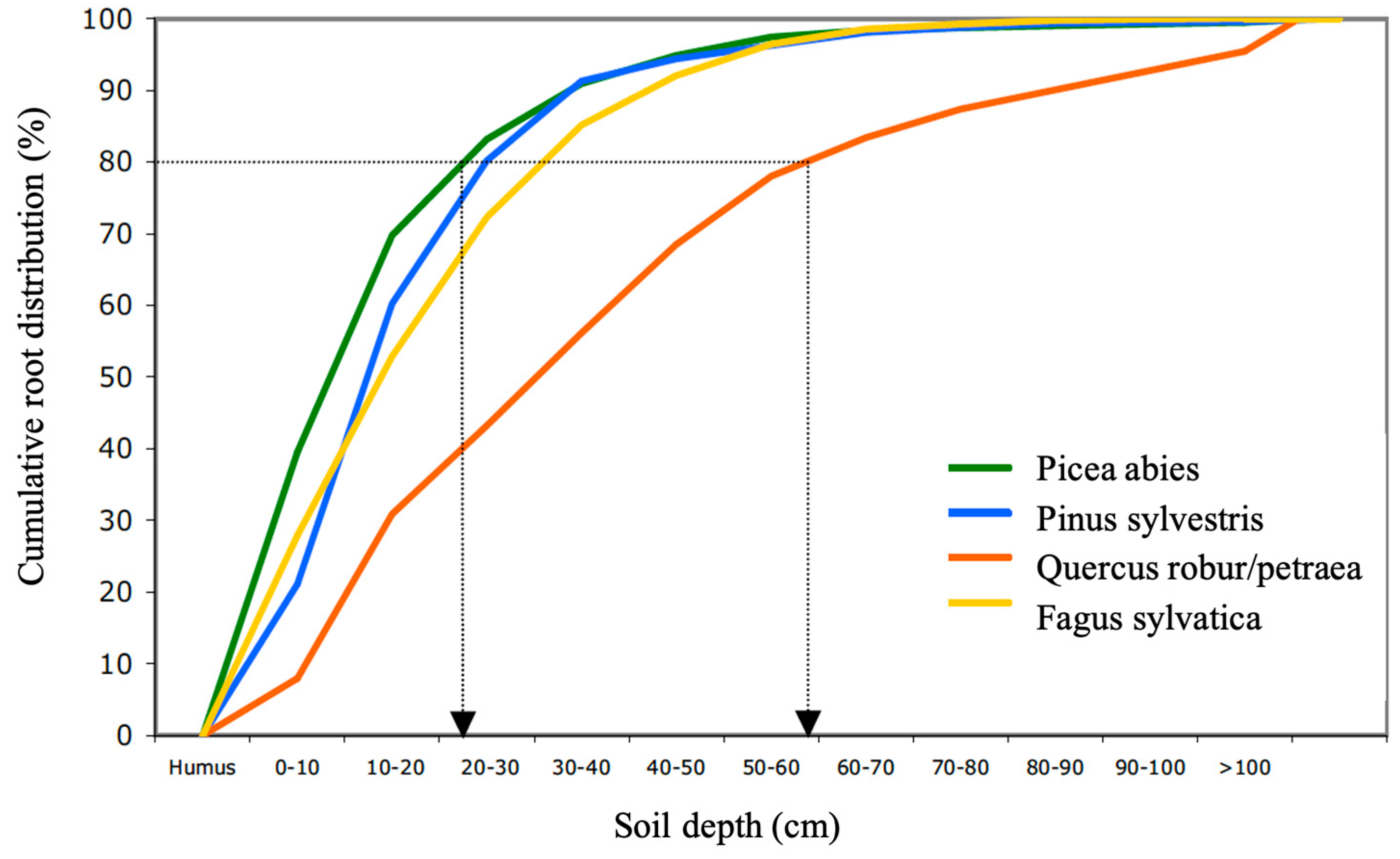
3. Root and Mycorrhiza’s Distribution and Growth
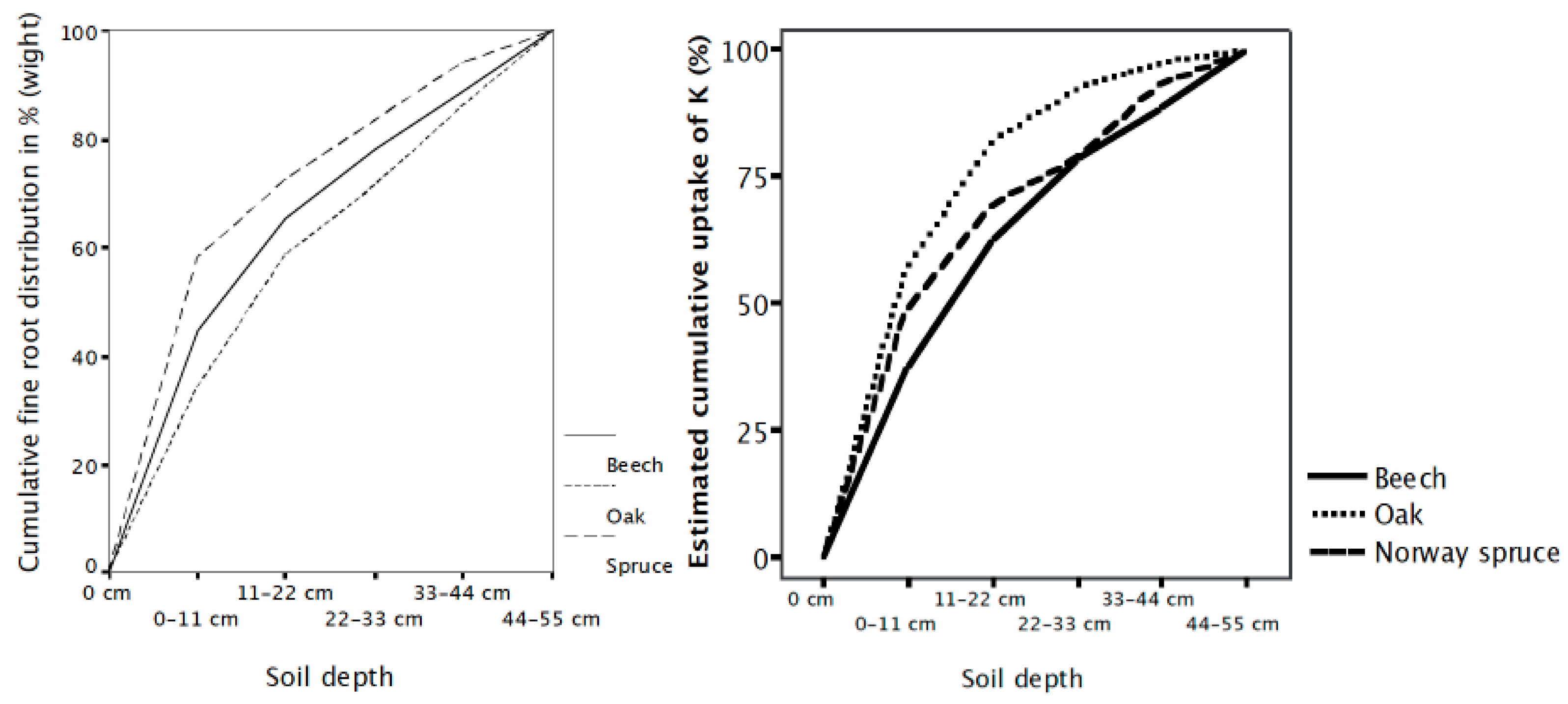
4. Root and Mycorrhiza Uptake Efficiency
5. Implications for Modeling Forest Nutrition
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sverdrup, H. Nutrient sustainability for Swedish forests. In Developing Principles and Models for Sustainable Forestry in Sweden; Sverdrup, H., Stjernquist, I., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; Volume 5. [Google Scholar]
- Krüger, I.S.T.; Potočić, N.; Ukonmaanaho, L.P.R. Increased Evidence of Nutrient Imbalances in Forest Trees across Europe, ICP Forests Brief 4; Thünen Institute of Forest Ecosystems: Braunschweig, Germany, 2020. [Google Scholar]
- Svenning, J.C.; Skov, F. Ice age legacies in the geographical distribution of tree species richness in Europe. Global Ecol. Biogeogr. 2007, 16, 234–245. [Google Scholar] [CrossRef]
- Bernasconi, S.M.; Bauder, A.; Bourdon, B.; Brunner, I.; Bünemann, E.; Chirs, I.; Derungs, N.; Edwards, P.; Farinotti, D.; Frey, B.; et al. Chemical and biological gradients along the Damma Glacier soil chronosequence (Switzerland). Vadose Zone J. 2011, 10, 867–883. [Google Scholar] [CrossRef]
- Binkley, D.; Högberg, P. Tamm Review: Revisiting the influence of nitrogen deposition on Swedish forests. For. Ecol. Manage. 2016, 368, 222–239. [Google Scholar] [CrossRef]
- De Vries, W.; Posch, M. Modelling the impact of nitrogen deposition, climate change and nutrient limitations on tree carbon sequestration in Europe for the period 1900–2050. Environ. Pollut. 2011, 159, 2289–2299. [Google Scholar] [CrossRef] [PubMed]
- Augustin, S.; Bolte, A.; Holzhausen, M.; Wolff, B. Exceedance of critical loads of nitrogen and sulphur and its relation to forest conditions. Eur. J. For. Res. 2005, 124, 289–300. [Google Scholar] [CrossRef]
- Waldner, P.; Thimonier, A.; Pannatier, E.G.; Etzold, S.; Schmitt, M.; Marchetto, A.; Rautio, P.; Derome, K.; Nieminen, T.M.; Nevalainen, S. Exceedance of critical loads and of critical limits impacts tree nutrition across Europe. Ann. For. Sci. 2015, 72, 929–939. [Google Scholar] [CrossRef]
- De Vries, W.; Vel, E.; Reinds, G.; Deelstra, H.; Klap, J.; Leeters, E.; Hendriks, C.; Kerkvoorden, M.; Landmann, G.; Herkendell, J. Intensive monitoring of forest ecosystems in Europe: 1. Objectives, set-up and evaluation strategy. For. Ecol. Manag. 2003, 174, 77–95. [Google Scholar] [CrossRef]
- Sardans, J.; Alonso, R.; Janssens, I.A.; Carnicer, J.; Vereseglou, S.; Rillig, M.C.; Fernández-Martínez, M.; Sanders, T.G.; Peñuelas, J. Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: Relationships with climate, N deposition and tree growth. Funct. Ecol. 2015, 30, 676–689. [Google Scholar] [CrossRef]
- Du, E.; van Doorn, M.; de Vries, W. Spatially divergent trends of nitrogen versus phosphorus limitation across European forests. Sci. Total Environ. 2021, 771, 145391. [Google Scholar] [CrossRef]
- Jonard, M.; Fürst, A.; Verstraeten, A.; Thimonier, A.; Timmermann, V.; Potočić, N.; Waldner, P.; Benham, S.; Hansen, K.; Merilä, P. Tree mineral nutrition is deteriorating in Europe. Glob. Chang. Biol. 2015, 21, 418–430. [Google Scholar] [CrossRef]
- Thelin, G. Ash Recycling to Spruce and Beech Stands Effects on Nutrients, Growth, Nitrogen Dynamics and Carbon Balance; Report 965; Värmeforsk Service AB: Stockholm, Sweden, 2006. (In Swedish) [Google Scholar]
- Akselsson, C.; Westling, O.; Alveteg, M.; Thelin, G.; Fransson, A.-M.; Hellsten, S. The influence of N load and harvest intensity on the risk of P limitation in Swedish forest soils. Sci. Total Environ. 2008, 404, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Terrer, C.; Pellegrini, A.F.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Ostertag, R.; DiManno, N.M. Detecting terrestrial nutrient limitation: A global meta-analysis of foliar nutrient concentrations after fertilization. Front. Earth Sci. 2016, 4, 23. [Google Scholar] [CrossRef]
- Talkner, U.; Meiwes, K.J.; Potočić, N.; Seletković, I.; Cools, N.; De Vos, B.; Rautio, P. Phosphorus nutrition of beech (Fagus sylvatica L.) is decreasing in Europe. Ann. For. Sci. 2015, 72, 919–928. [Google Scholar] [CrossRef]
- Braun, S.; Schindler, C.; Rihm, B. Foliar nutrient concentrations of European beech in Switzerland: Relations with nitrogen deposition, ozone, climate and soil chemistry. Front. For. Glob. Chang. 2020, 3, 33. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Wang, L.; Katzensteiner, K.; Schume, H.; Van Loo, M.; Godbold, D.L. Potassium fertilization affects the distribution of fine roots but does not change ectomycorrhizal community structure. Ann. For. Sci. 2016, 73, 691–702. [Google Scholar] [CrossRef]
- Sarkkola, S.; Ukonmaanaho, L.; Nieminen, T.M.; Laiho, R.; Laurén, A.; Finér, L.; Nieminen, M. Should harvest residues be left on site in peatland forests to decrease the risk of potassium depletion? For. Ecol. Manag. 2016, 374, 136–145. [Google Scholar] [CrossRef]
- Penuelas, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sardans, J. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Chang. Biol. 2020, 26, 1962–1985. [Google Scholar] [CrossRef]
- Schmitz, A.; Sanders, T.G.; Bolte, A.; Bussotti, F.; Dirnböck, T.; Johnson, J.; Peñuelas, J.; Pollastrini, M.; Prescher, A.-K.; Sardans, J. Responses of forest ecosystems in Europe to decreasing nitrogen deposition. Environ. Pollut. 2019, 244, 980–994. [Google Scholar] [CrossRef]
- Göttlein, A. Grenzwertbereiche für die ernährungsdiagnostische Einwertung der Hauptbaumarten Fichte, Kiefer, Eiche, Buche. Allg. Forst. Jagdztg. 2015, 186, 110–116. [Google Scholar]
- Braun, S.; Tresch, S.; Augustin, S. Soil solution in Swiss forest stands: A 20 year’s time series. PLoS ONE 2020, 15, e0227530. [Google Scholar] [CrossRef] [PubMed]
- Falkengren-Grerup, U.; Linnermark, N.; Tyler, G. Changes in acidity and cation pools of southern Sweden soils between 1949 and 1985. Chemosphere 1987, 16, 2239–2248. [Google Scholar] [CrossRef]
- Falkengren-Grerup, U.; Brunet, J.; Diekmann, M. Nitrogen mineralisation in deciduous forest soils in south Sweden in gradients of soil acidity and deposition. Environ. Pollut. 1998, 102, 415–420. [Google Scholar] [CrossRef]
- Jönsson, U.; Rosengren, U.; Thelin, G.; Nihlgård, B. Acidification-induced chemical changes in coniferous forest soils in southern Sweden. Environ. Pollut. 2003, 123, 75–83. [Google Scholar] [CrossRef]
- Göransson, H.; Rosengren, U.; Thelin, G.; Nihlgård, B. Effects on soil chemistry and nutrient uptake by admixture of birch in Norway spruce stands. In Sustainable Forestry in Temperate Regions; Lund University: Lund, Sweden, 2002. [Google Scholar]
- Johnson, J.; Graf Pannatier, E.; Carnicelli, S.; Cecchini, G.; Clarke, N.; Cools, N.; Hansen, K.; Meesenburg, H.; Nieminen, T.M.; Pihl-Karlsson, G. The response of soil solution chemistry in European forests to decreasing acid deposition. Glob. Chang. Biol. 2018, 24, 3603–3619. [Google Scholar] [CrossRef]
- Naturvårdsverket. Bara Naturlig Föryngring; Naturvårdsverket: Stockholm, Sweden, 2022; ISBN 978-91-620-7069-4.
- Mylona, S. Sulphur dioxide emissions in Europe 1880–1991 and their effect on sulphur concentrations and depositions. Tellus B 1996, 48, 662–689. [Google Scholar] [CrossRef]
- Fowler, D.; Smith, R.; Muller, J.; Cape, J.N.; Sutton, M.; Erisman, J.W.; Fagerli, H. Long term trends in sulphur and nitrogen deposition in Europe and the cause of non-linearities. Water Air Soil Pollut. Focus 2007, 7, 41–47. [Google Scholar] [CrossRef]
- Cools, N.; De Vos, B. Availability and evaluation of European forest soil monitoring data in the study on the effects of air pollution on forests. Iforest-Biogeosci. For. 2011, 4, 205. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef]
- Solberg, S.; Dobbertin, M.; Reinds, G.J.; Lange, H.; Andreassen, K.; Fernandez, P.G.; Hildingsson, A.; de Vries, W. Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: A stand growth approach. For. Ecol. Manag. 2009, 258, 1735–1750. [Google Scholar] [CrossRef]
- Sverdrup, H.; Stjernquist, I. Developing Principles and Models for Sustainable Forestry in Sweden; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Kint, V.; Aertsen, W.; Campioli, M.; Vansteenkiste, D.; Delcloo, A.; Muys, B. Radial growth change of temperate tree species in response to altered regional climate and air quality in the period 1901–2008. Clim. Chang. 2012, 115, 343–363. [Google Scholar] [CrossRef]
- Wall, A. Risk analysis of effects of whole-tree harvesting on site productivity. For. Ecol. Manag. 2012, 282, 175–184. [Google Scholar] [CrossRef]
- Clarke, N.; Kiær, L.P.; Kjønaas, O.J.; Bárcena, T.G.; Vesterdal, L.; Stupak, I.; Finér, L.; Jacobson, S.; Armolaitis, K.; Lazdina, D. Effects of intensive biomass harvesting on forest soils in the Nordic countries and the UK: A meta-analysis. For. Ecol. Manag. 2021, 482, 118877. [Google Scholar] [CrossRef]
- Roy, S.; Leban, J.-M.; Zeller, B.; van der Heijden, G.; Reichard, A.; Gehin, M.-C.; Santenoise, P.; Saint-Andre, L. Removing harvest residues from hardwood stands affects tree growth, wood density and stem wood nutrient concentration in European beech (Fagus sylvatica) and oak (Quercus spp.). For. Ecosyst. 2022, 9, 100014. [Google Scholar] [CrossRef]
- Rosengren, U.; Göransson, H.; Jönsson, U.; Stjernquist, I.; Thelin, G.; Wallander, H. Functional Biodiversity Aspects on the Nutrient Sustainability in Forests—Importance of Root Distribution. J. Sustain. For. 2005, 21, 75–98. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.-D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Germon, A.; Laclau, J.-P.; Robin, A.; Jourdan, C. Tamm Review: Deep fine roots in forest ecosystems: Why dig deeper? For. Ecol. Manag. 2020, 466, 118135. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Uhlig, D.; Amelung, W.; Von Blanckenburg, F. Mineral nutrients sourced in deep regolith sustain long-term nutrition of mountainous temperate forest ecosystems. Glob. Biogeochem. Cycles 2020, 34, e2019GB006513. [Google Scholar] [CrossRef]
- Holloway, J.M.; Dahlgren, R.A. Nitrogen in rock: Occurrences and biogeochemical implications. Glob. Biogeochem. Cycles 2002, 16, 65-1–65-17. [Google Scholar] [CrossRef]
- Augusto, L.; Turpault, M.-P.; Ranger, J. Impact of forest tree species on feldspar weathering rates. Geoderma 2000, 96, 215–237. [Google Scholar] [CrossRef]
- Stendahl, J.; Akselsson, C.; Melkerud, P.-A.; Belyazid, S. Pedon-scale silicate weathering: Comparison of the PROFILE model and the depletion method at 16 forest sites in Sweden. Geoderma 2013, 211, 65–74. [Google Scholar] [CrossRef]
- Sverdrup, H.; Warfvinge, P. Calculating field weathering rates using a mechanistic geochemical model PROFILE. Appl. Geochem. 1993, 8, 273–283. [Google Scholar] [CrossRef]
- Rosengren, U.; Stjernquist, I. Gå på Djupet: Om Rotdjup och Rotproduktion i Olika Skogstyper; SUFOR Report; Plant Ecology and Systematics: Lund, Sweden, 2004. [Google Scholar]
- Wallander, H.; Göransson, H.; Rosengren, U. Production, standing biomass and ∂15N/∂13C abundance of ectomycorrhizal mycelia at different soil depths in spruce forests and mixed (spruce-oak) forests in southern Sweden. Oecologia 2004, 139, 89–97. [Google Scholar] [CrossRef]
- Leake, J.R. Is diversity of ectomycorrhizal fungi important for ecosystem function? New Phytol. 2001, 152, 1–3. [Google Scholar] [CrossRef]
- Göransson, H. The Vertical Distribution of Roots, Mycorrhizal Mycelia and Nutrient Acquisition in Mature Forest Trees; Lund University: Lund, Sweden, 2006. [Google Scholar]
- Göransson, H.; Wallander, H.; Ingerslev, M.; Rosengren, U. Estimating the relative nutrient uptake from different soil depth of Quercus robur, Fagus sylvatica and Picea abies (L.) Karst. Plant Soil 2006, 286, 87–97. [Google Scholar] [CrossRef]
- Becerra, A.; Bartoloni, N.; Cofré, N.; Soteras, F.; Cabello, M. Arbuscular mycorrhizal fungi in saline soils: Vertical distribution at different soil depth. Braz. J. Microbiol. 2014, 45, 585–594. [Google Scholar] [CrossRef]
- de Araujo Pereira, A.P.; Santana, M.C.; Bonfim, J.A.; de Lourdes Mescolotti, D.; Cardoso, E.J.B.N. Digging deeper to study the distribution of mycorrhizal arbuscular fungi along the soil profile in pure and mixed Eucalyptus grandis and Acacia mangium plantations. Appl. Soil Ecol. 2018, 128, 1–11. [Google Scholar] [CrossRef]
- Köble, R.; Seufert, G. Novel maps for forest tree species in Europe. In Proceedings of the 8th European Symposium on the Physico-Chemical Behaviour of Air Pollutants: “A Changing Atmosphere”, Torino, Italy, 17–20 September 2001. [Google Scholar]
- von Zoth, R.; Block, J. Untersuchungen anWurzelballen sturmgeworfener Bäume in Rheinland-Pfalz. Forst Holz 1992, 47, 566–571. [Google Scholar]
- Ericsson, T. Growth and shoot: Root ratio of seedlings in relation to nutrient availability. Plant Soil 1995, 168–169, 205–214. [Google Scholar] [CrossRef]
- Ericsson, T.; Rytter, L.; Vapaavuori, E. Physiology of carbon allocation in trees. Biomass Bioenergy 1996, 11, 115–127. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, A.; Wallander, H.; Carlsson, M.L.; Huss-Danell, K. Fungal Biomass in Roots and Extramatrical Mycelium in Relation to Macronutrients and Plant Biomass of Ectomycorrhizal Pinus sylvestris and Alnus incana. New Phytol. 1995, 131, 443–451. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants--an economic analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Li, W.; Jin, C.; Guan, D.; Wang, Q.; Wang, A.; Yuan, F.; Wu, J. The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Biol. Biochem. 2015, 82, 112–118. [Google Scholar] [CrossRef]
- Zhao, X.; Tian, Q.; Huang, L.; Lin, Q.; Wu, J.; Liu, F. Fine-root functional trait response to nitrogen deposition across forest ecosystems: A meta-analysis. Sci. Total Environ. 2022, 844, 157111. [Google Scholar] [CrossRef]
- Ma, X.; Zhu, B.; Nie, Y.; Liu, Y.; Kuzyakov, Y. Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen deposition: A meta-analysis. Soil Biol. Biochem. 2021, 163, 108418. [Google Scholar] [CrossRef]
- Luo, M.; Moorhead, D.L.; Ochoa-Hueso, R.; Mueller, C.W.; Ying, S.C.; Chen, J. Nitrogen loading enhances phosphorus limitation in terrestrial ecosystems with implications for soil carbon cycling. Funct. Ecol. 2022, 36, 2845–2858. [Google Scholar] [CrossRef]
- Steingrobe, B. A sensitivity analysis for assessing the relevance of fine-root turnover for P and K uptake. J. Plant Nutr. Soil Sci. 2005, 168, 496–502. [Google Scholar] [CrossRef]
- Bahr, A.; Ellström, M.; Akselsson, C.; Ekblad, A.; Mikusinska, A.; Wallander, H. Growth of ectomycorrhizal fungal mycelium along a Norway spruce forest nitrogen deposition gradient and its effect on nitrogen leakage. Soil Biol. Biochem. 2013, 59, 38–48. [Google Scholar] [CrossRef]
- Nilsson, L.O.; Wallander, H. Production of external mycelium by ectomycorrhizal fungi in Norway spruce forest was reduced in response to nitrogen fertilization. New Phytol. 2003, 158, 409–416. [Google Scholar] [CrossRef]
- Nilsson, L.O.; Bååth, E.; Falkengren-Grerup, U.; Wallander, H. Growth of ectomycorrhizal mycelia and composition of soil microbial communities in oak forest soils along a nitrogen deposition gradient. Oecologia 2007, 153, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Lilleskov, E.A.; Kuyper, T.W.; Bidartondo, M.I.; Hobbie, E.A. Atmospheric nitrogen deposition impacts on the structure and function of forest mycorrhizal communities: A review. Environ. Pollut. 2019, 246, 148–162. [Google Scholar] [CrossRef]
- Averill, C.; Dietze, M.C.; Bhatnagar, J.M. Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks. Glob. Chang. Biol. 2018, 24, 4544–4553. [Google Scholar] [CrossRef]
- Lehmann, J. Subsoil root activity in tree-based cropping systems. Plant Soil 2003, 255, 319–331. [Google Scholar] [CrossRef]
- Soethe, N.; Lehmann, J.; Engels, C. The vertical pattern of rooting and nutrient uptake at different altitudes of a south Ecuadorian montane forest. Plant Soil 2006, 286, 287–299. [Google Scholar] [CrossRef]
- Brandtberg, P.-O.; Bengtsson, J.; Lundkvist, H. Distribution of the capacity to take up nutrients by Betula spp. and Picea abies in mixed stands. For. Ecol. Manag. 2004, 198, 193–208. [Google Scholar] [CrossRef]
- Göransson, H.; Ingerslev, M.; Wallander, H. The vertical distribution of N and K uptake in relation to root distribution and root uptake capacity in mature Quercus robur, Fagus sylvatica and Picea abies stands. Plant Soil 2008, 306, 129–137. [Google Scholar] [CrossRef]
- Newman, E.I.; Andrews, R.E. Uptake of phosphorus and potassium in relation to root growth and root density. Plant Soil 1973, 38, 49–69. [Google Scholar] [CrossRef]
- Göransson, H.; Rosengren, U.; Wallander, H.; Fransson, A.-M.; Thelin, G. Nutrient acquisition from different soil depths by Pedunculate oak. Trees 2005, 20, 292–298. [Google Scholar] [CrossRef]
- Thelin, G.; Sverdrup, H.; Holmqvist, J.; Rosengren, U.; Linden, M. Sustainability in spruce and mixed-species stands. In Developing Principles and Models for Sustainable Forestry in Sweden; Sverdrup, H., Stjernquist, I., Eds.; Managing forest ecosystems; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; Volume 5, pp. 337–354. [Google Scholar]
- da Silva, E.V.; Bouillet, J.P.; de Moraes Gonçalves, J.L.; Junior, C.H.A.; Trivelin, P.C.O.; Hinsinger, P.; Jourdan, C.; Nouvellon, Y.; Stape, J.L.; Laclau, J.P. Functional specialization of Eucalyptus fine roots: Contrasting potential uptake rates for nitrogen, potassium and calcium tracers at varying soil depths. Funct. Ecol. 2011, 25, 996–1006. [Google Scholar] [CrossRef]
- Göransson, H.; Fransson, A.-M.; Jönsson-Belyazid, U. Do oaks have different strategies for uptake of N, K and P depending on soil depth? Plant Soil 2007, 297, 119–125. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in Sugar maple root respiration with root diameter and soil depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Bergkvist, B.; Folkeson, L. The influence of tree species on acid deposition, proton budgets and element fluxes in south Swedish forest ecosystems. Ecol. Bull. 1995, 44, 90–99. [Google Scholar]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Rosling, A.; Landeweert, R.; Lindahl, B.D.; Larsson, K.-H.; Kuyper, T.W.; Taylor, A.F.S.; Finlay, R.D. Vertical distribution of ectomycorrhizal fungal taxa in a podzol soil profile. New Phytol. 2003, 159, 775–783. [Google Scholar] [CrossRef]
- Dickie, I.A.; Xu, B.; Koide, R.T. Vertical niche differentiation of ectomycorrhizal hyphae in soil as shown by T-RFLP analysis. New Phytol. 2002, 156, 527–535. [Google Scholar] [CrossRef]
- Dighton, J.; Mason, P.P.; Poskitt, J.M. Field use of 32P to measure phosphate uptake by birch mycorrhizas. New Phytol. 1990, 116, 655–661. [Google Scholar] [CrossRef]
- Dighton, J.; Poskitt, J.M.; Brown, T.K. Phosphate influx into ectomycorrhizal and saprophytic fungal hyphae in relations to phosphate supply: A potential method for selection of efficient mycorrhizal species. Mycol. Res. 1993, 97, 355–358. [Google Scholar] [CrossRef]
- Wallander, H.; Wickman, T.; Jacks, G. Apatite as a P source in mycorrhizal and non-mycorrhizal Pinus sylvestris seedlings. Plant Soil 1997, 196, 123–131. [Google Scholar] [CrossRef]
- Mensah, J.A.; Koch, A.M.; Antunes, P.M.; Kiers, E.T.; Hart, M.; Bücking, H. High functional diversity within species of arbuscular mycorrhizal fungi is associated with differences in phosphate and nitrogen uptake and fungal phosphate metabolism. Mycorrhiza 2015, 25, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.M.; Hanson, P.J.; Iversen, C.M.; Kumar, J.; Walker, A.P.; Wullschleger, S.D. Root structural and functional dynamics in terrestrial biosphere models–evaluation and recommendations. New Phytol. 2015, 205, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Sverdrup, H.; Thelin, G.; Robles, M.; Stjernquist, I.; Sörensen, J. Assesing nutrient sustainability of forest production for different tree species considering Ca, Mg, K, N and P at Björnstorp Estate, Sweden. Biogeochemistry 2006, 81, 219–238. [Google Scholar] [CrossRef]
- Wang, B.; McCormack, M.L.; Ricciuto, D.M.; Yang, X.; Iversen, C.M. Embracing fine-root system complexity in terrestrial ecosystem modeling. Glob. Chang. Biol. 2023, 29, 2871–2885. [Google Scholar] [CrossRef]
- Wallman, P.; Svensson, M.G.E.; Sverdrup, H. ForSAFE-an integrated process-oriented forest model for long-term sustainability assessments. For. Ecol. Manag. 2005, 207, 19–36. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandén, H. Are European Trees Prepared for Being Limited by Other Nutrients than Nitrogen? Sustainability 2024, 16, 7980. https://doi.org/10.3390/su16187980
Sandén H. Are European Trees Prepared for Being Limited by Other Nutrients than Nitrogen? Sustainability. 2024; 16(18):7980. https://doi.org/10.3390/su16187980
Chicago/Turabian StyleSandén, Hans. 2024. "Are European Trees Prepared for Being Limited by Other Nutrients than Nitrogen?" Sustainability 16, no. 18: 7980. https://doi.org/10.3390/su16187980
APA StyleSandén, H. (2024). Are European Trees Prepared for Being Limited by Other Nutrients than Nitrogen? Sustainability, 16(18), 7980. https://doi.org/10.3390/su16187980