
Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling

 , , and
, , and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Feedstock | Cultivation | Yield [g/g] | Ref. |
|---|---|---|---|---|
| L. rhamnosus LA-04-01 a | Defatted rice brain hydrolysate | Batch | 0.95 | [14] |
| Continuous | 0.98 | |||
| L. paracasei 7BL a | Wood chips | Fed-Batch | 0.96 | [15] |
| Rice straw | 0.97 | |||
| L. casei G-02 a | Jerusalem artichoke | Fed-Batch (SSF) | 0.96 | [16] |
| L. agilis LPB 56 b | Soybean vinasse | Batch | 0.85 | [17] |
| L. plantarum NCIMB 8826 a | Delignified hardwood pulp | Batch (SSF) | 0.88 | [18] |
| L. paracasei LA104 a | Curcuma longa waste | Batch (SSCF) | 0.69 | [19] |
| L. coryniformis ATCC 25600 b | 0.65 | |||
| L. pentosus FL0421 a | Corn stover | Fed-Batch (SSF) | 0.66 | [20] |
| L. pentosus DSM20314 a | Wheat bran | Batch | 0.73 | [21] |
| L. delbrueckii NBRC 3202 b | Cassava fibrous waste | Batch | 0.50 | [22] |
| L. rhamosus ATCC 7469 a | Lignocellulosic mixture | Batch (SSF) | 0.97 | [23] |
| L. delbrueckii subsp. bulgarius ATCC 11842 b | Beechwood hydrolysate | Batch (SSF) | 0.69 | [24] |
| Pine hydrolysate | 0.40 | |||
| L. delbrueckii DSM 20074 b | Household bio-waste | Batch | 0.65 | [25] |
| L. casei DSM 20011 a | Agro-industrial waste | Batch | 0.78 | [26] |
| L. delbrueckii subsp. lactis DSMZ 20072 b | Brewers’ spent grain | Batch | 0.89 | [27] |
| L. brevis MTCC 4460 c | Cottonseed cake | Batch (SSCF) | 0.22 | [28] |
| Wheat straw | 0.49 | |||
| Sugarcane bagasse | 0.52 | |||
| L. buchneri NRRL B-30929 c | Elephant grass liquor | Batch | 0.50 | [29] |
| B. coagulans A107 b | Tapioca starch hydrolysate | Continuous | 0.80 | [7] |
| B. coagulans AD b | Corn stover hydrolysate | Continuous | 0.95 | [30] |
| B. coagulans LA204 b | Corncob | Fed-Batch (SSF) | 0.77 | [31] |
| B. coagulans LA1507 b | Sweet sorghum bagasse | Open-Fed-Batch (SSF) | 0.44 | [30] |
| E. faecalis RKY1 b | Molasses | Batch | 0.95 | [32] |
| E. faecium WH51-1 b | Corn steep water effluent | Batch | 0.89 | [33] |
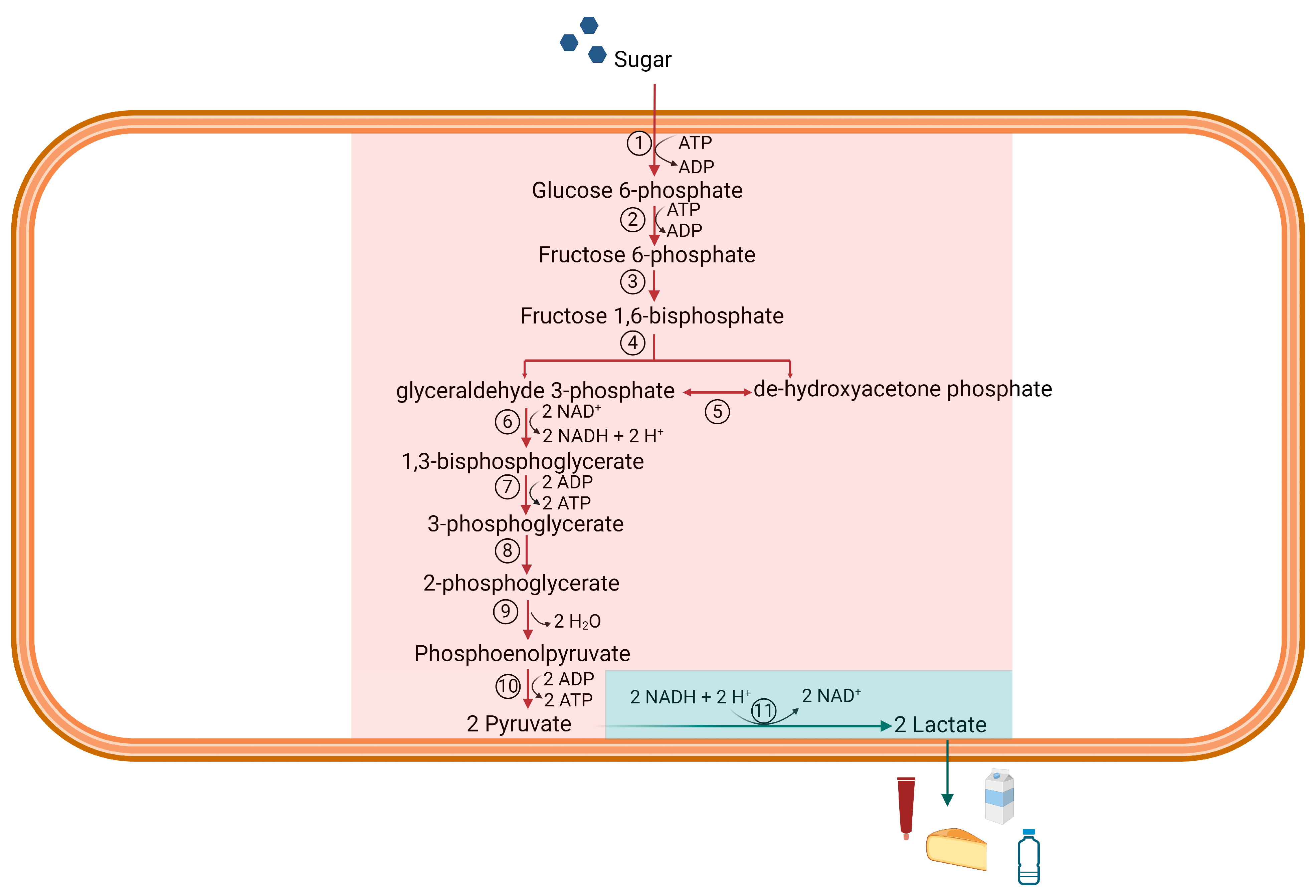
2. Materials and Methods
2.1. Raw Material
2.2. Pretreatment and Preparation of Sterile Press-Juice
2.3. Microorganism, Preculture, and Complex Medium Preparation
2.4. Cultivation in Different Percentages of Press-Juice
2.5. Bench-Scale Fermentation in Schott Bottles
2.6. Scaled-Up Fermentation in the Bioreactor
2.7. Ensiling of Different Varieties of Lolium perenne
2.8. Analytical Methods
2.9. Data Processing and Evaluation
3. Results and Discussion
3.1. Influence of Raw Material
3.2. Press-Juice Analysis
3.3. Bench-Scale Fermentation in Complex Medium
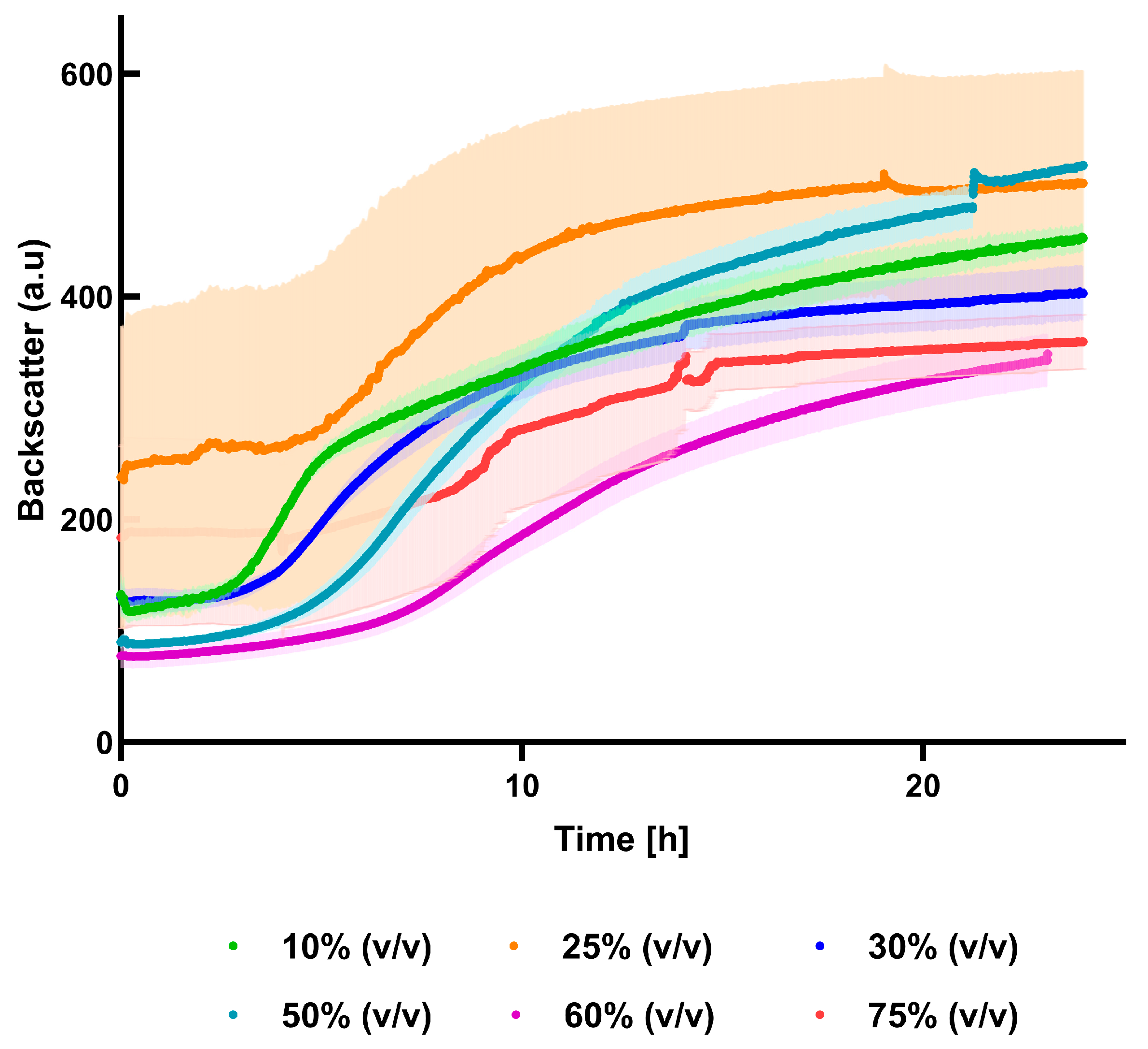
3.4. Cultivation in Different Percentages of Press-Juice
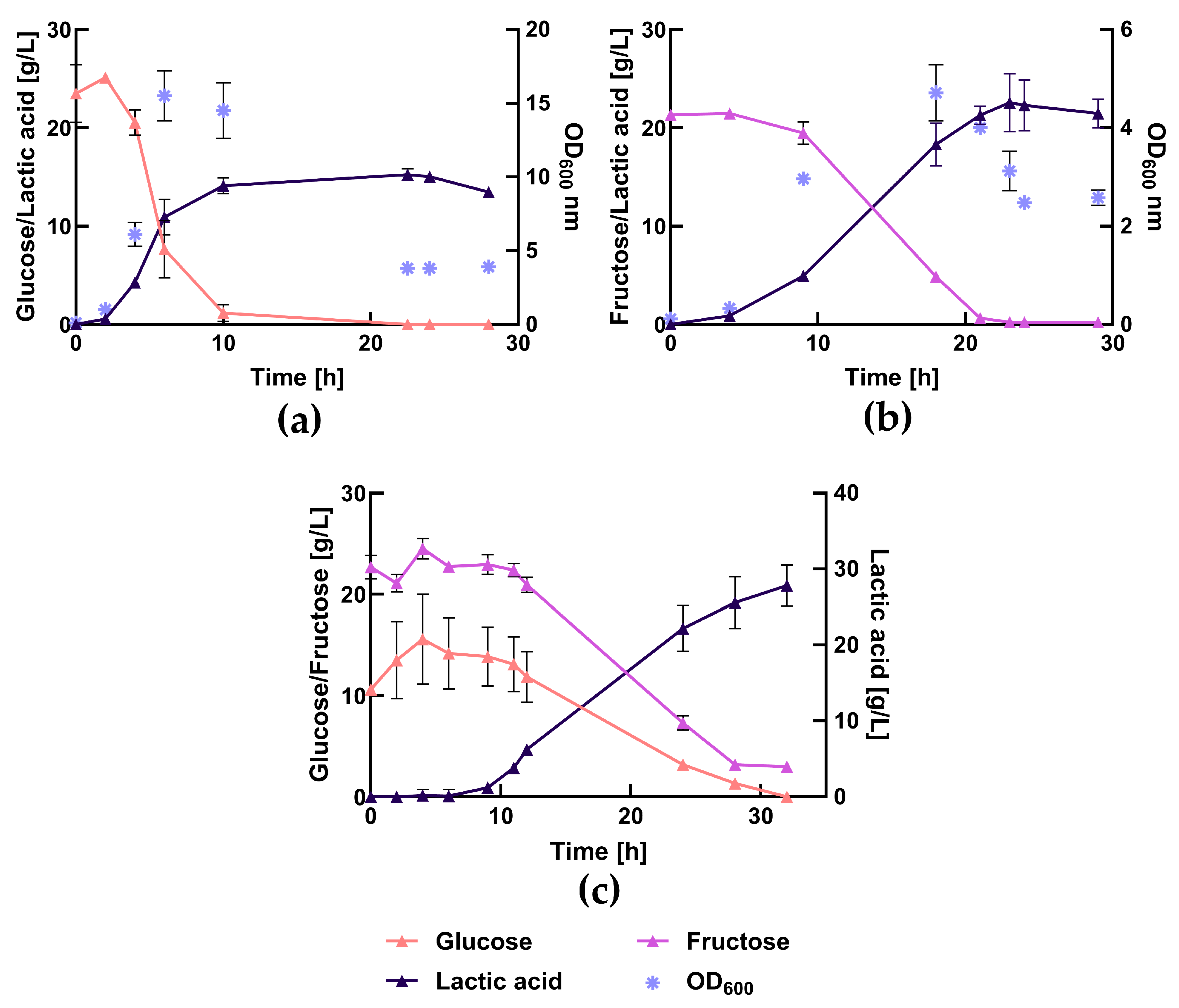
3.5. Bench-Scale Fermentation Using the Press-Juice
3.6. Comparison between Complex Medium and Press-Juice in the Bioreactor
3.7. Lactic Acid Yields from Ensiling Whole Grass Fractions of Different Varieties of Lolium perenne
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antar, M.; Lyu, D.; Nazari, M.; Shah, A.; Zhou, X.; Smith, D.L. Biomass for a Sustainable Bioeconomy: An Overview of World Biomass Production and Utilization. Renew. Sustain. Energy Rev. 2021, 139, 110691. [Google Scholar] [CrossRef]
- Yadav, V.G.; Yadav, G.D.; Patankar, S.C. The Production of Fuels and Chemicals in the New World: Critical Analysis of the Choice between Crude Oil and Biomass Vis-à-Vis Sustainability and the Environment. Clean Technol. Environ. Policy 2020, 22, 1757–1774. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Feng, Y.; Fu, J.; Guo, H.; Guo, Y.; Han, B.; Jiang, Z.; Kong, L.; Li, C.; Liu, H.; et al. Catalytic Conversion of Lignocellulosic Biomass into Chemicals and Fuels. Green Energy Environ. 2023, 8, 10–114. [Google Scholar] [CrossRef]
- McEniry, J.; O’Kiely, P. Developments in Grass-/Forage-Based Biorefineries. In Advances in Biorefineries: Biomass and Waste Supply Chain Exploitation; Waldron, K., Ed.; Woodhead Publishing: Sawston, UK, 2014; pp. 335–363. ISBN 9780857095213. [Google Scholar]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent Advances in Lactic Acid Production by Microbial Fermentation Processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Balla, E.; Daniilidis, V.; Karlioti, G.; Kalamas, T.; Stefanidou, M.; Bikiaris, N.D.; Vlachopoulos, A.; Koumentakou, I.; Bikiaris, D.N. Poly(Lactic Acid): A Versatile Biobased Polymer for the Future with Multifunctional Properties—From Monomer Synthesis, Polymerization Techniques and Molecular Weight Increase to PLA Applications. Polymers 2021, 13, 1822. [Google Scholar] [CrossRef]
- López-Gómez, J.P.; Alexandri, M.; Schneider, R.; Venus, J. A Review on the Current Developments in Continuous Lactic Acid Fermentations and Case Studies Utilising Inexpensive Raw Materials. Process. Biochem. 2019, 79, 1–10. [Google Scholar] [CrossRef]
- Alves de Oliveira, R.; Komesu, A.; Vaz Rossell, C.E.; Maciel Filho, R. Challenges and Opportunities in Lactic Acid Bioprocess Design—From Economic to Production Aspects. Biochem. Eng. J. 2018, 133, 219–239. [Google Scholar] [CrossRef]
- Varriale, L.; Ulber, R. Fungal-Based Biorefinery: From Renewable Resources to Organic Acids. ChemBioEng Rev. 2023, 10, 272–292. [Google Scholar] [CrossRef]
- Ajala, E.O.; Olonade, Y.O.; Ajala, M.A.; Akinpelu, G.S. Lactic Acid Production from Lignocellulose—A Review of Major Challenges and Selected Solutions. ChemBioEng Rev. 2020, 7, 38–49. [Google Scholar] [CrossRef]
- Di Lorenzo, R.D.; Serra, I.; Porro, D.; Branduardi, P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts 2022, 12, 234. [Google Scholar] [CrossRef]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production—Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Gobbetti, M. Lactobacillus spp.: General Characteristics. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Li, L.; Cai, D.; Wang, C.; Han, J.; Ren, W.; Zheng, J.; Wang, Z.; Tan, T. Continuous l-Lactic Acid Production from Defatted Rice Bran Hydrolysate Using Corn Stover Bagasse Immobilized Carrier. RSC Adv. 2015, 5, 18511–18517. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Yuan, S.F.; Wang, C.A.; Huang, Y.J.; Guo, G.L.; Hwang, W.S. Production of Optically Pure l-Lactic Acid from Lignocellulosic Hydrolysate by Using a Newly Isolated and d-Lactate Dehydrogenase Gene-Deficient Lactobacillus paracasei Strain. Bioresour. Technol. 2015, 198, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.Y.; Qian, H.; Zhang, W.G. Enhancement of l-Lactic Acid Production in Lactobacillus casei from Jerusalem Artichoke Tubers by Kinetic Optimization and Citrate Metabolism. J. Microbiol. Biotechnol. 2010, 20, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Karp, S.G.; Igashiyama, A.H.; Siqueira, P.F.; Carvalho, J.C.; Vandenberghe, L.P.S.; Thomaz-Soccol, V.; Coral, J.; Tholozan, J.L.; Pandey, A.; Soccol, C.R. Application of the Biorefinery Concept to Produce l-Lactic Acid from the Soybean Vinasse at Laboratory and Pilot Scale. Bioresour. Technol. 2011, 102, 1765–1772. [Google Scholar] [CrossRef]
- Hama, S.; Mizuno, S.; Kihara, M.; Tanaka, T.; Ogino, C.; Noda, H.; Kondo, A. Production of d-Lactic Acid from Hardwood Pulp by Mechanical Milling Followed by Simultaneous Saccharification and Fermentation Using Metabolically Engineered Lactobacillus Plantarum. Bioresour. Technol. 2015, 187, 167–172. [Google Scholar] [CrossRef]
- Nguyen, C.M.; Kim, J.S.; Nguyen, T.N.; Kim, S.K.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Kim, J.C. Production of l- and d-Lactic Acid from Waste Curcuma longa Biomass through Simultaneous Saccharification and Cofermentation. Bioresour. Technol. 2013, 146, 35–43. [Google Scholar] [CrossRef]
- Hu, J.; Lin, Y.; Zhang, Z.; Xiang, T.; Mei, Y.; Zhao, S.; Liang, Y.; Peng, N. High-Titer Lactic Acid Production by Lactobacillus pentosus FL0421 from Corn Stover Using Fed-Batch Simultaneous Saccharification and Fermentation. Bioresour. Technol. 2016, 214, 74–80. [Google Scholar] [CrossRef]
- Tirpanalan, Ö.; Reisinger, M.; Smerilli, M.; Huber, F.; Neureiter, M.; Kneifel, W.; Novalin, S. Wheat Bran Biorefinery—An Insight into the Process Chain for the Production of Lactic Acid. Bioresour. Technol. 2015, 180, 242–249. [Google Scholar] [CrossRef]
- Cingadi, S.; Srikanth, K.; Arun, E.V.R.; Sivaprakasam, S. Statistical Optimization of Cassava Fibrous Waste Hydrolysis by Response Surface Methodology and Use of Hydrolysate Based Media for the Production of Optically Pure d-Lactic Acid. Biochem. Eng. J. 2015, 102, 82–90. [Google Scholar] [CrossRef]
- Pontes, R.; Romaní, A.; Michelin, M.; Domingues, L.; Teixeira, J.; Nunes, J. l-Lactic Acid Production from Multi-Supply Autohydrolyzed Economically Unexploited Lignocellulosic Biomass. Ind. Crop. Prod. 2021, 170, 113775. [Google Scholar] [CrossRef]
- Karnaouri, A.; Asimakopoulou, G.; Kalogiannis, K.G.; Lappas, A.; Topakas, E. Efficient d-Lactic Acid Production by Lactobacillus delbrueckii subsp. bulgaricus through Conversion of Organosolv Pretreated Lignocellulosic Biomass. Biomass Bioenergy 2020, 140, 105672. [Google Scholar] [CrossRef]
- Tsapekos, P.; Alvarado-Morales, M.; Baladi, S.; Bosma, E.F.; Angelidaki, I. Fermentative Production of Lactic Acid as a Sustainable Approach to Valorize Household Bio-Waste. Front. Sustain. 2020, 1, 4. [Google Scholar] [CrossRef]
- Costa, S.; Summa, D.; Semeraro, B.; Zappaterra, F.; Rugiero, I.; Tamburini, E. Fermentation as a Strategy for Bio-Transforming Waste into Resources: Lactic Acid Production from Agri-Food Residues. Fermentation 2020, 7, 3. [Google Scholar] [CrossRef]
- Akermann, A.; Weiermüller, J.; Christmann, J.; Guirande, L.; Glaser, G.; Knaus, A.; Ulber, R. Brewers’ Spent Grain Liquor as a Feedstock for Lactate Production with Lactobacillus delbrueckii subsp. lactis. Eng. Life Sci. 2020, 20, 168–180. [Google Scholar] [CrossRef]
- Grewal, J.; Khare, S.K. One-Pot Bioprocess for Lactic Acid Production from Lignocellulosic Agro-Wastes by Using Ionic Liquid Stable Lactobacillus brevis. Bioresour. Technol. 2018, 251, 268–273. [Google Scholar] [CrossRef]
- Montipó, S.; Ballesteros, I.; Fontana, R.C.; Liu, S.; Martins, A.F.; Ballesteros, M.; Camassola, M. Integrated Production of Second Generation Ethanol and Lactic Acid from Steam-Exploded Elephant Grass. Bioresour. Technol. 2018, 249, 1017–1024. [Google Scholar] [CrossRef]
- Ahring, B.K.; Traverso, J.J.; Murali, N.; Srinivas, K. Continuous Fermentation of Clarified Corn Stover Hydrolysate for the Production of Lactic Acid at High Yield and Productivity. Biochem. Eng. J. 2016, 109, 162–169. [Google Scholar] [CrossRef]
- Zhang, Z.; Xie, Y.; He, X.; Li, X.; Hu, J.; Ruan, Z.; Zhao, S.; Peng, N.; Liang, Y. Comparison of High-Titer Lactic Acid Fermentation from NaOH-and NH3-H2O2-Pretreated Corncob by Bacillus coagulans Using Simultaneous Saccharification and Fermentation. Sci. Rep. 2016, 6, 37245. [Google Scholar] [CrossRef]
- Wee, Y.J.; Kim, J.N.; Yun, J.S.; Ryu, H.W. Utilization of Sugar Molasses for Economical L(+)-Lactic Acid Production by Batch Fermentation of Enterococcus faecalis. Enzyme Microb. Technol. 2004, 35, 568–573. [Google Scholar] [CrossRef]
- Selim, M.T.; Salem, S.S.; Fouda, A.; El-Gamal, M.S.; Abdel-Rahman, M.A. Use of Corn-Steep Water Effluent as a Promising Substrate for Lactic Acid Production by Enterococcus faecium Strain Wh51-1. Fermentation 2021, 7, 111. [Google Scholar] [CrossRef]
- Talukder, M.M.R.; Das, P.; Wu, J.C. Microalgae (Nannochloropsis salina) Biomass to Lactic Acid and Lipid. Biochem. Eng. J. 2012, 68, 109–113. [Google Scholar] [CrossRef]
- Kim, J.; Kim, Y.-M.; Lebaka, V.R.; Wee, Y.-J. Lactic Acid for Green Chemical Industry: Recent Advances in and Future Prospects for Production Technology, Recovery, and Applications. Fermentation 2022, 8, 609. [Google Scholar] [CrossRef]
- Büyükkileci, A.O.; Harsa, S. Batch Production of L(+) Lactic Acid from Whey by Lactobacillus casei (NRRL B-441). J. Chem. Technol. Biotechnol. 2004, 79, 1036–1040. [Google Scholar] [CrossRef]
- Hayek, S.A.; Ibrahim, S.A. Current Limitations and Challenges with Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2013, 2013, 73–87. [Google Scholar] [CrossRef]
- Pessione, A.; Lamberti, C.; Pessione, E. Proteomics as a Tool for Studying Energy Metabolism in Lactic Acid Bacteria. Mol. Biosyst. 2010, 6, 1419–1430. [Google Scholar] [CrossRef]
- Vasco-Correa, J.; Zuleta-Correa, A.; Gómez-León, J.; Pérez-Taborda, J.A. Advances in Microbial Pretreatment for Biorefining of Perennial Grasses. Appl. Microbiol. Biotechnol. 2023, 107, 5281–5300. [Google Scholar] [CrossRef]
- Baruah, J.; Nath, B.K.; Sharma, R.; Kumar, S.; Deka, R.C.; Baruah, D.C.; Kalita, E. Recent Trends in the Pretreatment of Lignocellulosic Biomass for Value-Added Products. Front. Energy Res. 2018, 6, 141. [Google Scholar] [CrossRef]
- Varriale, L.; Volkmar, M.; Weiermüller, J.; Ulber, R. Effects of Pretreatment on the Biocatalysis of Renewable Resources. Chem. Ing. Tech. 2022, 94, 1818–1826. [Google Scholar] [CrossRef]
- Gonçalves, M.C.P.; Romanelli, J.P.; Cansian, A.B.M.; Pucci, E.F.Q.; Guimarães, J.R.; Tardioli, P.W.; Saville, B.A. A Review on the Production and Recovery of Sugars from Lignocellulosics for Use in the Synthesis of Bioproducts. Ind. Crops Prod. 2022, 186, 115213. [Google Scholar] [CrossRef]
- Kongkeitkajorn, M.B.; Sae-Kuay, C.; Reungsang, A. Evaluation of Napier Grass for Bioethanol Production through a Fermentation Process. Processes 2020, 8, 567. [Google Scholar] [CrossRef]
- Taufikurahman; Sherly; Jessica; Delimanto, W.O. Production of Bioethanol from Napier Grass: Comparison in Pre-Treatment and Fermentation Methods. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 520. [Google Scholar] [CrossRef]
- Damborg, V.K.; Jensen, S.K.; Weisbjerg, M.R.; Adamsen, A.P.; Stødkilde, L. Screw-Pressed Fractions from Green Forages as Animal Feed: Chemical Composition and Mass Balances. Anim. Feed Sci. Technol. 2020, 261, 114401. [Google Scholar] [CrossRef]
- Varriale, L.; Geib, D.; Ulber, R. Short-Term Adaptation as a Tool to Improve Bioethanol Production Using Grass Press-Juice as Fermentation Medium. Appl. Microbiol. Biotechnol. 2024, 108, 393. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free Lactic Acid Production under Acidic Conditions by Lactic Acid Bacteria Strains: Challenges and Future Prospects. Appl. Microbiol. Biotechnol. 2018, 102, 5911–5924. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Fernandes, M.; Mancilha, I.M.; Roberto, I.C. Effects of Medium Supplementation and PH Control on Lactic Acid Production from Brewer’s Spent Grain. Biochem. Eng. J. 2008, 40, 437–444. [Google Scholar] [CrossRef]
- Si, H.; Liang, X.; Liu, S.; Xu, M.; Wang, J.; Hua, D. Efficient Production of l-Lactic Acid from Corn Straw Hydrolysate. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 514. [Google Scholar] [CrossRef]
- Hébert, E.M.; Raya, R.R.; De Giori, G.S. Nutritional Requirements of Lactobacillus delbrueckii subsp. lactis in a Chemically Defined Medium. Curr. Microbiol. 2004, 49, 341–345. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic Systems of Lactic Acid Bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Richter, K.; Träger, A. L(+)-Lactic Acid from Sweet Sorghum by Submerged and Solid-state Fermentations. Acta Biotechnol. 1994, 14, 367–378. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Cai, D.; Wang, B.; Wang, Z.; Qin, P.; Tan, T. Efficient l-Lactic Acid Production from Sweet Sorghum Bagasse by Open Simultaneous Saccharification and Fermentation. RSC Adv. 2016, 6, 35771–35777. [Google Scholar] [CrossRef]
- Iino, T.; Uchimura, T.; Komagata, K. The Effect of Sodium Acetate on the Growth Yield, the Production of l- and d-Lactic Acid, and the Activity of Some Enzymes of the Glycolytic Pathway of Lactobacillus Sakei NRIC 1071 and Lactobacillus plantarum NRIC 1067. J. Gen. Appl. Microbiol. 2002, 48, 91–102. [Google Scholar] [CrossRef]
- Coelho, L.F.; De Lima, C.J.B.; Rodovalho, C.M.; Bernardo, M.P.; Contiero, J. Lactic Acid Production by New Lactobacillus Plantarum LMISM6 Grown in Molasses: Optimization of Medium Composition. Braz. J. Chem. Eng. 2011, 28, 27–36. [Google Scholar] [CrossRef]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Emam-Djomeh, Z.; Kiani, H. Fermentation of Pomegranate Juice by Probiotic Lactic Acid Bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- Volkmar, M.; Maus, A.L.; Weisbrodt, M.; Bohlender, J.; Langsdorf, A.; Holtmann, D.; Ulber, R. Municipal Green Waste as Substrate for the Microbial Production of Platform Chemicals. Bioresour. Bioprocess 2023, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Boakye-Boaten, N.A.; Xiu, S.; Shahbazi, A.; Wang, L.; Li, R.; Schimmel, K. Uses of Miscanthus Press Juice within a Green Biorefinery Platform. Bioresour. Technol. 2016, 207, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Santamaria-Fernandez, M.; Ambye-Jensen, M.; Damborg, V.K.; Lübeck, M. Demonstration-Scale Protein Recovery by Lactic Acid Fermentation from Grass Clover—A Single Case of the Production of Protein Concentrate and Press Cake Silage for Animal Feeding Trials. Biofuels Bioprod. Biorefining 2019, 13, 502–513. [Google Scholar] [CrossRef]
- Rolfe, M.D.; Rice, C.J.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.D.S.; Alston, M.; Stringer, M.F.; Betts, R.P.; Baranyi, J.; et al. Lag Phase Is a Distinct Growth Phase That Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. J. Bacteriol. 2012, 194, 686–701. [Google Scholar] [CrossRef]
- Hamill, P.G.; Stevenson, A.; McMullan, P.E.; Williams, J.P.; Lewis, A.D.R.; Sudharsan, S.; Stevenson, K.E.; Farnsworth, K.D.; Khroustalyova, G.; Takemoto, J.Y.; et al. Microbial Lag Phase Can Be Indicative of, or Independent from, Cellular Stress. Sci. Rep. 2020, 10, 5948. [Google Scholar] [CrossRef]
- Petrut, S.; Rusu, E.; Tudorache, I.S.; Pelinescu, D.; Sarbu, I.; Stoica, I.; Vassu, T. Influence of Various Carbon Sources on Growth and Biomass Accumulation of Some Lactic Acid Bacteria Strains. Rev. Chim. 2019, 70, 2434–2438. [Google Scholar] [CrossRef]
- Chen, H.; Niu, J.; Qin, T.; Ma, Q.; Wang, L.; Shu, G. Optimization of the Medium for Lactobacillus Acidophilus by Plackett-Burman and Steepest Ascent Experiment. Acta Sci. Pol. Technol. Aliment. 2015, 14, 227–232. [Google Scholar] [CrossRef]
- Chen, P.T.; Hong, Z.S.; Cheng, C.L.; Ng, I.S.; Lo, Y.C.; Nagarajan, D.; Chang, J.S. Exploring Fermentation Strategies for Enhanced Lactic Acid Production with Polyvinyl Alcohol-Immobilized Lactobacillus Plantarum 23 Using Microalgae as Feedstock. Bioresour. Technol. 2020, 308, 123266. [Google Scholar] [CrossRef]
- De la Torre, I.; Acedos, M.G.; Ladero, M.; Santos, V.E. On the Use of Resting L. delbrueckii spp. Delbrueckii Cells for d-Lactic Acid Production from Orange Peel Wastes Hydrolysates. Biochem. Eng. J. 2019, 145, 162–169. [Google Scholar] [CrossRef]
- Erliana, W.H.; Widjaja, T.; Altway, A.; Pudjiastuti, L. Synthesis of Lactic Acid from Sugar Palm Trunk Waste (Arenga pinnata): Preliminary Hydrolysis and Fermentation Studies. Biodiversitas 2020, 21, 2281–2288. [Google Scholar] [CrossRef]
- Kung, L.; Shaver, R. Interpretation and Use of Silage Fermentation Analysis Reports. Focus Forage 2001, 3, 1–5. [Google Scholar]
- Johnson, H.E.; Merry, R.J.; Davies, D.R.; Kell, D.B.; Theodorou, M.K.; Griffith, G.W. Vacuum Packing: A Model System for Laboratory-Scale Silage Fermentations. J. Appl. Microbiol. 2005, 98, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Haag, N.L.; Nägele, H.J.; Fritz, T.; Oechsner, H. Effects of Ensiling Treatments on Lactic Acid Production and Supplementary Methane Formation of Maize and Amaranth—An Advanced Green Biorefining Approach. Bioresour. Technol. 2015, 178, 217–225. [Google Scholar] [CrossRef]
- Fazzino, F.; Luque, R.; Paone, E.; Padullà, A.; Sidari, R.; Calabrò, P.S. Long-Term Preservation of Orange Peel Waste for the Production of Acids and Biogas. ACS Sustain. Chem. Eng. 2022, 10, 13733–13741. [Google Scholar] [CrossRef]



| Variety | Ploidy | Breeder |
|---|---|---|
| Agaska | 2× | DLF (Roskilde, Denmark) |
| Honroso | 2× | DSV (Asendorf, Germany) |
| Arvicola | 4× | Feldsaaten Freudenberger (Krefeld, Germany) |
| Barmigo | 4× | Barenbrug (Nijmegen, The Netherlands) |
| Explosion | 4× | DSV (Asendorf, Germany) |
| Variety | Cut of the Year | Harvest Date | BBCH Stage | DM (t/ha) | CF (%) | CP (%) | WSCs (%) |
|---|---|---|---|---|---|---|---|
| Agaska a | 2 | 7 July 2021 | 49 | 2.5 ± 0.4 | 25.2 ± 1.4 | 11.6 ± 1.1 | 16.2 ± 0.6 |
| Arvicola a | 2 | 7 July 2021 | 31 | 2.5 ± 0.5 | 21.8 ± 1.4 | 10.9 ± 0.6 | 22.2 ± 0.9 |
| Barmigo a | 2 | 7 July 2021 | 45 | 2.5 ± 0.4 | 24.8 ± 0.2 | 10.5 ± 0.6 | 18.0 ± 0.3 |
| Explosion b | 1 | 3 June 2022 | 61 | 5.2 ± 0.0 | 25.9 ± 0.3 | 5.8 ± 0.7 | 22.6 ± 3.0 |
| Honroso b | 1 | 3 June 2022 | 59 | 4.3 ± 0.0 | 25.3 ± 1.6 | 7.9 ± 1.3 | 20.4 ± 0.3 |
| Arvicola | Agaska | Barmigo | |
|---|---|---|---|
| Total sugar [g/L] | 23.8 | 39.5 | 28.3 |
| Protein [mg/L] | 559.4 | 682.9 | 641.7 |
| Amino acids [mg/L] | n.a. | 3130.5 | 2560.9 |
| Cations [g/L] | 9.3 | 8.1 | 8.2 |
| Anions [g/L] | 6.0 | 5.9 | 4.8 |
| Sample | Glucose Content [% (m/m)] | Deviation | Fructose Content [% (m/m)] | Deviation |
|---|---|---|---|---|
| Honroso | 2.93% | 0.41% | 5.32% | 1.20% |
| Explosion | 5.09% | 0.79% | 7.56% | 3.00% |
| MRS Medium with Glucose 1 | MRS Medium with Fructose 1 | 75% (v/v) Press-Juice 2 | |
|---|---|---|---|
| Si [g/L] | 23.5 ± 2.9 | 21.3 ± 0.3 | 33.3 ± 0.84 |
| Sugar(s) remaining [g/L] | 0 ± 0.00 | 0.21 ± 00 | 2.9 ± 0.00 |
| Su [%] | 94.6 ± 4.5 | 77.2 ± 1.43 | 102.7 ± 3.5 |
| rS [g/Lh] | −2.23 ± 0.36 | −0.91 ± 0.03 | −1.03 ± 0.04 |
| CLA [g/L] | 13.47 ± 0.57 | 21.45 ± 1.45 | 27.81 ± 2.69 |
| S [g/g] | 0.59 ± 0.11 | 1.02 ± 0.08 | 0.84 ± 0.01 |
| YP/S [g/g] | 0.63 ± 0.02 | 1.02 ± 0.08 | 0.91 ± 0.07 |
| QLA [g/L h] | 0.48 ± 0.02 | 0.69 ± 0.00 | 0.87 ± 0.10 |
| Fermentation efficiency [%] | 51.3 ± 1.9 | 101.7 ± 8.4 | 91.4 ± 6.7 |
| YX/S [g/g] | 0.17 ± 0.02 | 0.12 ± 0.01 | n.a |
| QCELLS [g/L h] | 0.16 ± 0.00 | 0.09 ± 0.01 | n.a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varriale, L.; Hengsbach, J.-N.; Guo, T.; Kuka, K.; Tippkötter, N.; Ulber, R. Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling. Sustainability 2024, 16, 8054. https://doi.org/10.3390/su16188054
Varriale L, Hengsbach J-N, Guo T, Kuka K, Tippkötter N, Ulber R. Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling. Sustainability. 2024; 16(18):8054. https://doi.org/10.3390/su16188054
Chicago/Turabian StyleVarriale, Ludovica, Jan-Niklas Hengsbach, Tianyi Guo, Katrin Kuka, Nils Tippkötter, and Roland Ulber. 2024. "Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling" Sustainability 16, no. 18: 8054. https://doi.org/10.3390/su16188054
APA StyleVarriale, L., Hengsbach, J. -N., Guo, T., Kuka, K., Tippkötter, N., & Ulber, R. (2024). Sustainable Production of Lactic Acid Using a Perennial Ryegrass as Feedstock—A Comparative Study of Fermentation at the Bench- and Reactor-Scale, and Ensiling. Sustainability, 16(18), 8054. https://doi.org/10.3390/su16188054