
Perspectives and Progress in Bioethanol Processing and Social Economic Impacts

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
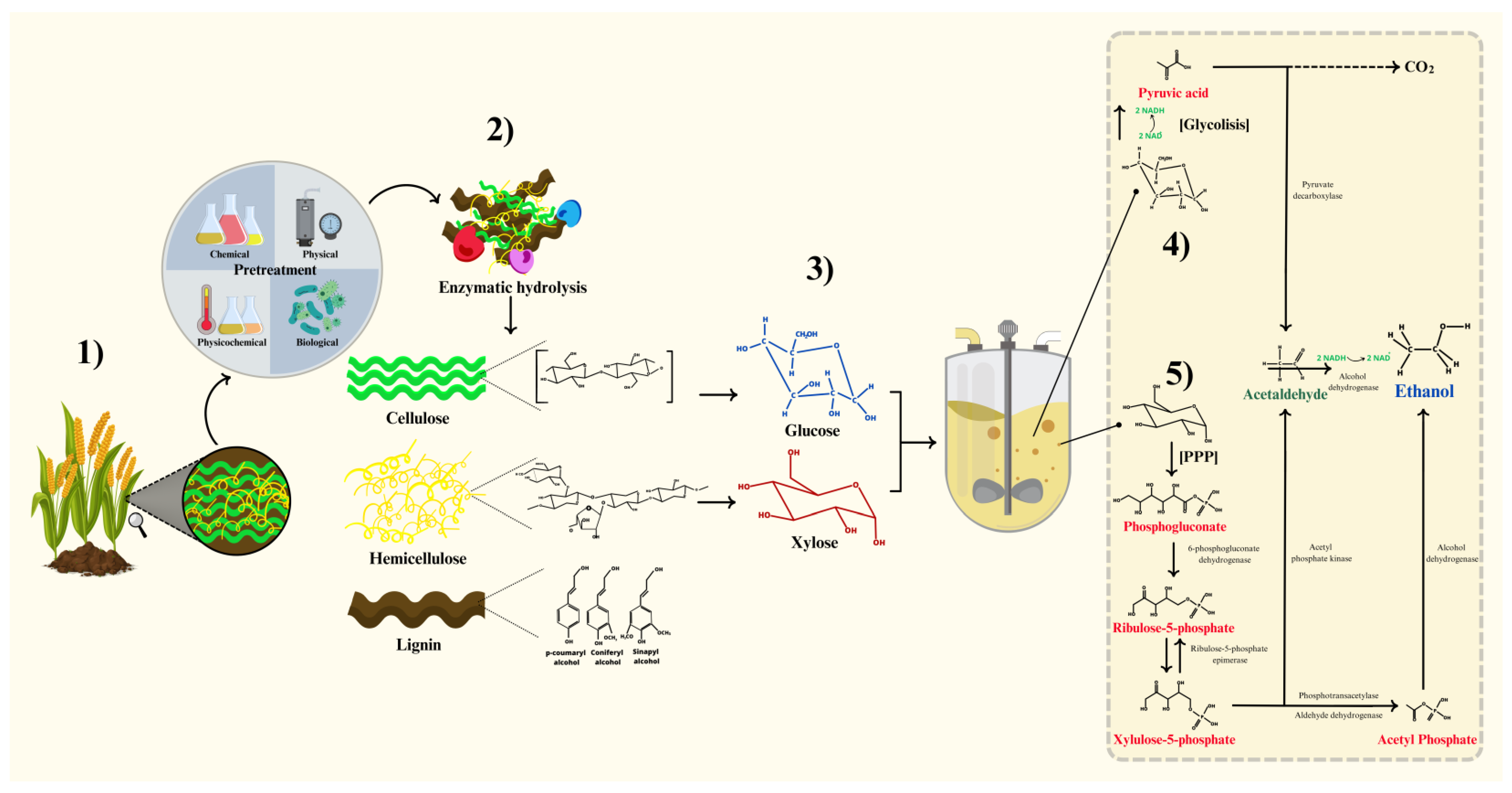
2. Breakdown of Lignocellulosic Substrates to Fermentable Sugars
2.1. Lignocellulosic Biomass Composition
2.2. Pretreatment Methods
3. Sugar Recovery from Pretreated Biomass through Hydrolysis
4. Metabolic Pathways, Settings, and Factors of Fermentation Process
4.1. Stress Factors and Inhibitors Affecting Ethanol Yield
4.2. Different Setups and Optimal Conditions of the Fermentation Process
4.3. Downstream Process: Ethanol Upgrading
5. Genetic Engineering Approaches to Improve Bioethanol Yield

{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Source | Modified Strain | Substrate | Bioethanol Concentration, g/L | Bioethanol Yield, g/g | Reference |
|---|---|---|---|---|---|---|
| XYL1, XYL2, XKS1, TAL1 | Scheffersomyce stipitis | S. cerevisiae MEC1122 | Glucose, xylose | 2.5 | 0.12 | [145] |
| PPP genes (TAL1, TKL1, RKI1, RPE1) | S. cerevisiae and Kluyveromyces marxianus | S. cerevisiae YK246 | Glucose, xylose | 48.6 | 0.45 | [146] |
| BvuXylA, XIqXylA, TAAXylA, araA, araB, araD | S. cerevisiae JUK36α | S. cerevisiae 36aS1.10.4 | Glucose, xylose | 54.11 | 0.44 | [141] |
| HXK2, and RSP5 | S. cerevisiae | S. cerevisiae IMS0629 | Glucose, xylose | - | 0.18 | [147] |
| PHO4 | S. cerevisiae MC15 | S. cerevisiae MF01-PHO4 | Glucose | 114.71 | - | [148] |
| PMA1, VMA1, VMA2, VMA4-8, VMA22 | S. cerevisiae | S. cerevisiae XUSAE57 | Glucose, xylose | - | 0.49 | [149] |
| GPD2, FPS1, ADH2, DLD3 | S. cerevisiae | S. cerevisiae SCGFAD | Glucose | 23,29 | - | [150] |
| SESTC | Ampullaria gigas Spix | S. cerevisiae wild type | Glucose | 7.53 | 0.377 | [151] |
| ADH2 and hyg | S. cerevisiae | S. cerevisiae ΔADH2 As2.4 | Glucose | 14.6 | - | [152] |
| noxE | Lactococcus lactiswas | S. cerevisiae JX123_noxE | Glucose, xylose | 55.5 | 0.433 | [153] |
| EG1, CBH1, Bglc | Trichoderma reesei | S. cerevisiae BY4743-4A | Glucose, xylose | 32.6 | 0.42 | [154] |
| EGII, CBHII, XynII, BGL, XylA | Trichoderma reesei, Aspergillus aculeatus and Aspergillus oryzae | S. cerevisiae Y5 | Xylose | 1.61 | 0.33 | [155] |
| BGL1, XYLA, XYN | Aspergillus aculeatus, Aspergillus oryzae and Trichoderma reesei | S. cerevisiae industrial strain | Glucose, xylose | 11.1 | 0.328 | [156] |
| XylA and Xyn | Aspergillus niger, Saccharophagus degradans | S. cerevisiae | Xylose | 6.0 | - | [157] |
| XYNII, XylA, BGL1 | Aspergillus aculeatus | S. cerevisiae MN8140/XBXX | Glucose, xylose | 8.2 | 0.32 | [158] |
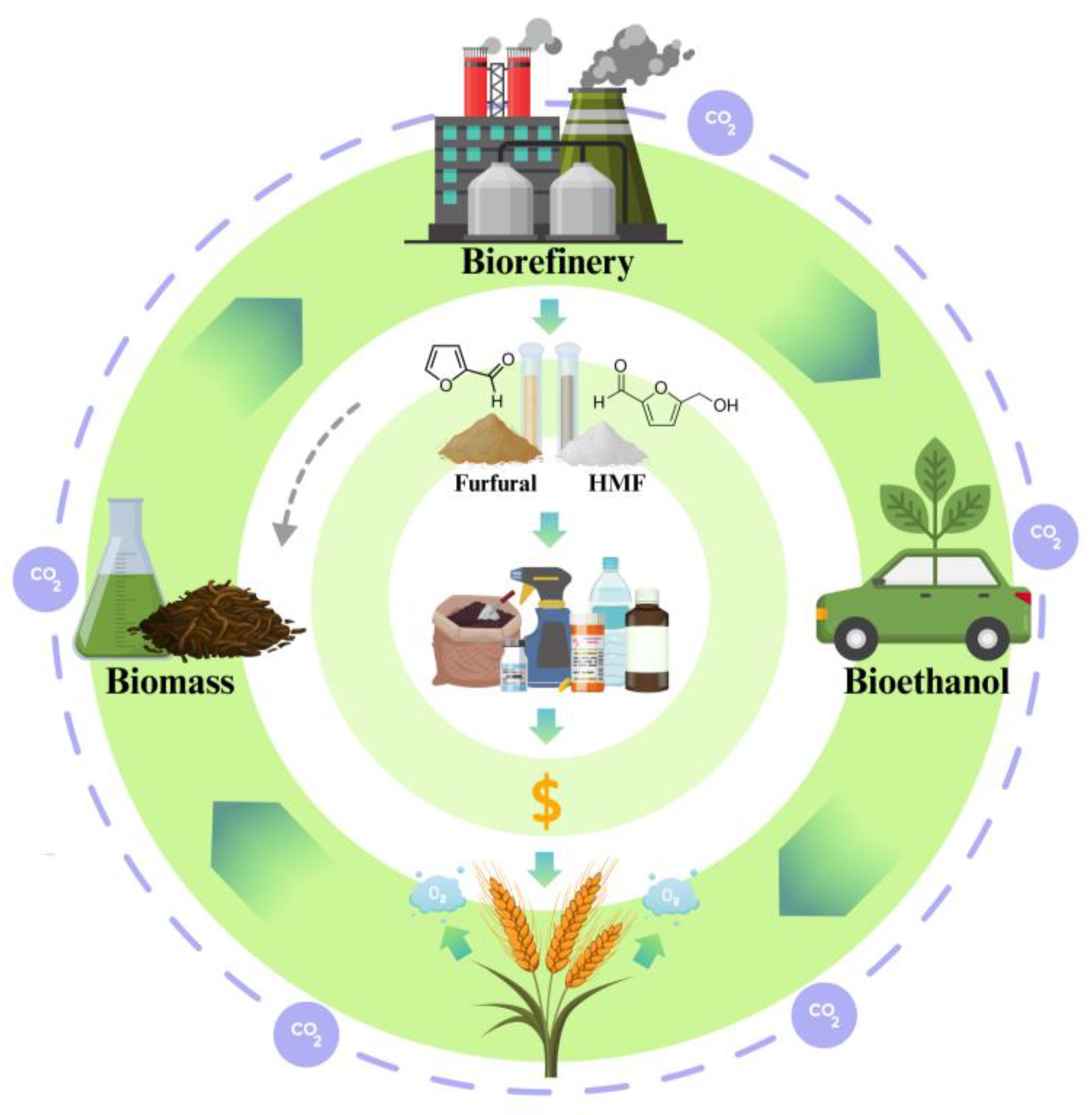
6. Challenges in Ethanol Conversion: Social, Economic, and Environmental Aspects
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sadigov, R. Rapid Growth of the World Population and Its Socioeconomic Results. Sci. World J. 2022, 2022, 8110229. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, Hannah, and Max Roser. “Cars, Planes, Trains: Where Do CO2 Emissions from Transport Come From?” Our World in Data. Available online: https://ourworldindata.org/co2-emissions-from-transport (accessed on 28 December 2023).
- Huang, Y.; Zhang, Y.; Deng, F.; Zhao, D.; Wu, R. Impacts of Built-Environment on Carbon Dioxide Emissions from Traffic: A Systematic Literature Review. Int. J. Environ. Res. Public Health 2022, 19, 16898. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.K.; Chatterjee, S.; Mazumder, P.B.; Gupta, S.K.; Sharma, S.; Vairale, M.G.; Datta, S.; Dwivedi, S.K.; Gupta, D.K. Bioethanol production from waste lignocelluloses: A review on microbial degradation potential. Chemosphere 2019, 231, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Kaur, M.; Punj, S.; Singh, K. Biomass as a sustainable resource for value-added modern materials: A review. Biofuels Bioprod. Biorefin. 2020, 14, 673–695. [Google Scholar] [CrossRef]
- Islam, K.; Wang, H.; Rehman, S.; Dong, C.; Hsu, H.-Y.; Lin, C.S.K.; Leu, S.-Y. Sustainability metrics of pretreatment processes in a waste derived lignocellulosic biomass biorefinery. Bioresour. Technol. 2020, 298, 122558. [Google Scholar] [CrossRef] [PubMed]
- Renewable Fuels Association. 2023 Ready. Set. Go! Renewable Fuels Association: Washington, DC, USA, 2023. [Google Scholar]
- Gonçalves, F.d.O.; Perna, R.F.; Lopes, E.S.; Tovar, L.P.; Filho, R.M.; Lopes, M.S. Strategies to Ensure Fuel Security in Brazil Considering a Forecast of Ethanol Production. Biomass 2023, 3, 1–17. [Google Scholar] [CrossRef]
- De Almeida, M.A.; Colombo, R. Production Chain of First-Generation Sugarcane Bioethanol: Characterization and Value-Added Application of Wastes. BioEnergy Res. 2023, 16, 924–939. [Google Scholar] [CrossRef]
- Carrillo-Nieves, D.; Alanís, M.J.R.; Quiroz, R.d.l.C.; Ruiz, H.A.; Iqbal, H.M.; Parra-Saldívar, R. Current status and future trends of bioethanol production from agro-industrial wastes in Mexico. Renew. Sustain. Energy Rev. 2019, 102, 63–74. [Google Scholar] [CrossRef]
- Santos, C.I.; Silva, C.C.; Mussatto, S.I.; Osseweijer, P.; van der Wielen, L.A.; Posada, J.A. Integrated 1st and 2nd generation sugarcane bio-refinery for jet fuel production in Brazil: Techno-economic and greenhouse gas emissions assessment. Renew. Energy 2018, 129, 733–747. [Google Scholar] [CrossRef]
- Aghaei, S.; Alavijeh, M.K.; Shafiei, M.; Karimi, K. A comprehensive review on bioethanol production from corn stover: Worldwide potential, environmental importance, and perspectives. Biomass Bioenergy 2022, 161, 106447. [Google Scholar] [CrossRef]
- Broda, M.; Yelle, D.J.; Serwańska, K. Bioethanol Production from Lignocellulosic Biomass—Challenges and Solutions. Molecules 2022, 27, 8717. [Google Scholar] [CrossRef]
- Maity, S.; Mallick, N. Trends and advances in sustainable bioethanol production by marine microalgae: A critical review. J. Clean. Prod. 2022, 345, 131153. [Google Scholar] [CrossRef]
- Ayodele, B.V.; Alsaffar, M.A.; Mustapa, S.I. An overview of integration opportunities for sustainable bioethanol production from first- and second-generation sugar-based feedstocks. J. Clean. Prod. 2020, 245, 118857. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.; Suely, A.; Boyce, A.; Faruq, G. Bioethanol production from renewable sources: Current perspectives and technological progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Duque, A.; Álvarez, C.; Doménech, P.; Manzanares, P.; Moreno, A.D. Advanced Bioethanol Production: From Novel Raw Materials to Integrated Biorefineries. Processes 2021, 9, 206. [Google Scholar] [CrossRef]
- Morales, M.; Arvesen, A.; Cherubini, F. Integrated process simulation for bioethanol production: Effects of varying lignocellulosic feedstocks on technical performance. Bioresour. Technol. 2021, 328, 124833. [Google Scholar] [CrossRef]
- El Akkari, M.; Ben Fradj, N.; Gabrielle, B.; Djomo, S.N. Spatially-explicit environmental assessment of bioethanol from miscanthus and switchgrass in France. Clean. Circ. Bioecon. 2023, 6, 100059. [Google Scholar] [CrossRef]
- Singh, A.; Chen, C.-W.; Patel, A.K.; Dong, C.-D.; Singhania, R.R. Subcritical Water Pretreatment for the Efficient Valorization of Sorghum Distillery Residue for the Biorefinery Platform. Bioengineering 2022, 10, 38. [Google Scholar] [CrossRef]
- Patel, A.; Shah, A.R. Integrated lignocellulosic biorefinery: Gateway for production of second generation ethanol and value added products. J. Bioresour. Bioprod. 2021, 6, 108–128. [Google Scholar] [CrossRef]
- Mujtaba, M.; Fraceto, L.F.; Fazeli, M.; Mukherjee, S.; Savassa, S.M.; de Medeiros, G.A.; Pereira, A.D.E.S.; Mancini, S.D.; Lipponen, J.; Vilaplana, F. Lignocellulosic biomass from agricultural waste to the circular economy: A review with focus on biofuels, biocomposites and bioplastics. J. Clean. Prod. 2023, 402, 136815. [Google Scholar] [CrossRef]
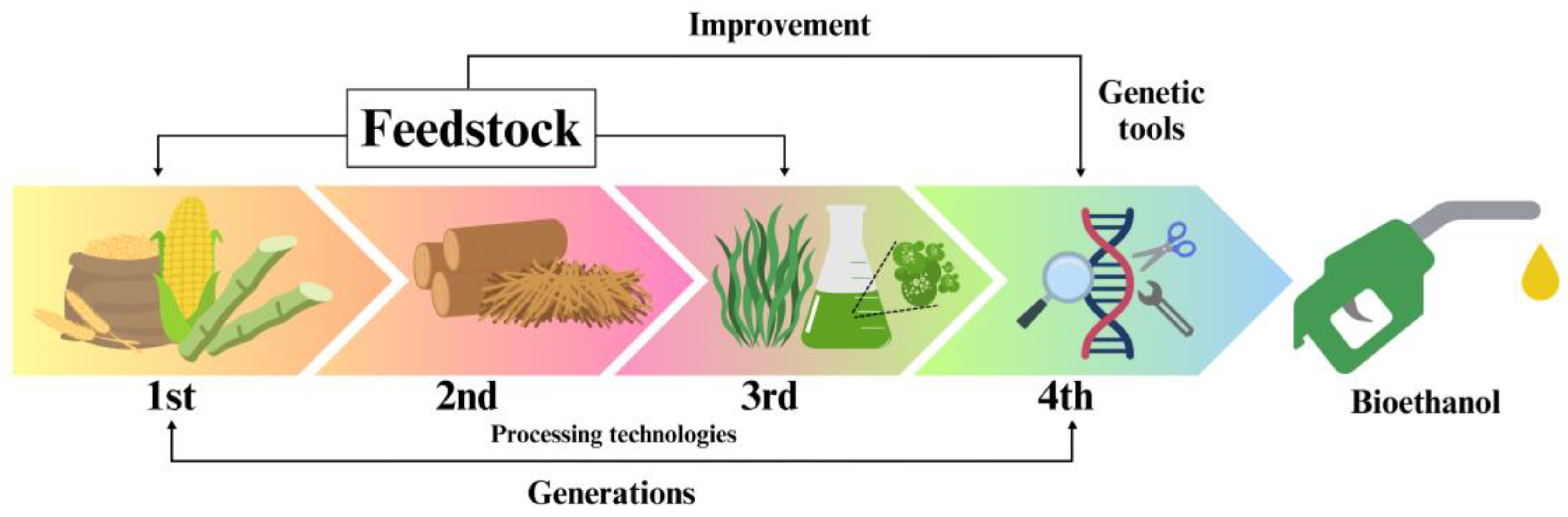
- Mat Aron, N.S.; Khoo, K.S.; Chew, K.W.; Show, P.L.; Chen, W.; Nguyen, T.H.P. Sustainability of the four generations of biofuels—A review. Int. J. Energy Res. 2020, 44, 9266–9282. [Google Scholar] [CrossRef]
- Müller, C.; Scapini, T.; Rempel, A.; Abaide, E.R.; Camargo, A.F.; Nazari, M.T.; Tadioto, V.; Bonatto, C.; Tres, M.V.; Zabot, G.L.; et al. Challenges and opportunities for third-generation ethanol production: A critical review. Eng. Microbiol. 2023, 3, 100056. [Google Scholar] [CrossRef]
- Anto, S.; Mukherjee, S.S.; Muthappa, R.; Mathimani, T.; Deviram, G.; Kumar, S.S.; Verma, T.N.; Pugazhendhi, A. Algae as green energy reserve: Technological outlook on biofuel production. Chemosphere 2020, 242, 125079. [Google Scholar] [CrossRef]
- Jambo, S.A.; Abdulla, R.; Azhar, S.H.M.; Marbawi, H.; Gansau, J.A.; Ravindra, P. A review on third generation bioethanol feedstock. Renew. Sustain. Energy Rev. 2016, 65, 756–769. [Google Scholar] [CrossRef]
- Ramachandra, T.; Hebbale, D. Bioethanol from macroalgae: Prospects and challenges. Renew. Sustain. Energy Rev. 2020, 117, 109479. [Google Scholar] [CrossRef]
- Sarwer, A.; Hamed, S.M.; Osman, A.I.; Jamil, F.; Al-Muhtaseb, A.H.; Alhajeri, N.S.; Rooney, D.W. Algal biomass valorization for biofuel production and carbon sequestration: A review. Environ. Chem. Lett. 2022, 20, 2797–2851. [Google Scholar] [CrossRef]
- Dave, N.; Selvaraj, R.; Varadavenkatesan, T.; Vinayagam, R. A critical review on production of bioethanol from macroalgal biomass. Algal Res. 2019, 42, 101606. [Google Scholar] [CrossRef]
- Chowdhury, H.; Loganathan, B. Third-generation biofuels from microalgae: A review. Curr. Opin. Green Sustain. Chem. 2019, 20, 39–44. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Ho, S.-H. Microalgae as a solution of third world energy crisis for biofuels production from wastewater toward carbon neutrality: An updated review. Chemosphere 2022, 291, 132863. [Google Scholar] [CrossRef]
- Bhatia, L.; Bachheti, R.K.; Garlapati, V.K.; Chandel, A.K. Third-generation biorefineries: A sustainable platform for food, clean energy, and nutraceuticals production. Biomass Convers. Biorefin. 2022, 12, 4215–4230. [Google Scholar] [CrossRef]
- Alalwan, H.A.; Alminshid, A.H.; Aljaafari, H.A. Promising evolution of biofuel generations. Subject review. Renew. Energy Focus 2019, 28, 127–139. [Google Scholar] [CrossRef]
- Sarwer, A.; Hussain, M.; Al-Muhtaseb, A.H.; Inayat, A.; Rafiq, S.; Khurram, M.S.; Ul-Haq, N.; Shah, N.S.; Din, A.A.; Ahmad, I.; et al. Suitability of Biofuels Production on Commercial Scale from Various Feedstocks: A Critical Review. ChemBioEng Rev. 2022, 9, 423–441. [Google Scholar] [CrossRef]
- Olguin-Maciel, E.; Singh, A.; Chable-Villacis, R.; Tapia-Tussell, R.; Ruiz, H.A. Consolidated Bioprocessing, an Innovative Strategy towards Sustainability for Biofuels Production from Crop Residues: An Overview. Agronomy 2020, 10, 1834. [Google Scholar] [CrossRef]
- Mehmood, M.A.; Shahid, A.; Malik, S.; Wang, N.; Javed, M.R.; Haider, M.N.; Verma, P.; Ashraf, M.U.F.; Habib, N.; Syafiuddin, A.; et al. Advances in developing metabolically engineered microbial platforms to produce fourth-generation biofuels and high-value biochemicals. Bioresour. Technol. 2021, 337, 125510. [Google Scholar] [CrossRef]
- Osman, A.I.; Qasim, U.; Jamil, F.; Al-Muhtaseb, A.H.; Abu Jrai, A.; Al-Riyami, M.; Al-Maawali, S.; Al-Haj, L.; Al-Hinai, A.; Al-Abri, M.; et al. Bioethanol and biodiesel: Bibliometric mapping, policies and future needs. Renew. Sustain. Energy Rev. 2021, 152, 111677. [Google Scholar] [CrossRef]
- Abdullah, B.; Muhammad, S.A.F.S.; Shokravi, Z.; Ismail, S.; Kassim, K.A.; Mahmood, A.N.; Aziz, M.A. Fourth generation biofuel: A review on risks and mitigation strategies. Renew. Sustain. Energy Rev. 2019, 107, 37–50. [Google Scholar] [CrossRef]
- Dietrich, K.; Dumont, M.-J.; Del Rio, L.F.; Orsat, V. Sustainable PHA production in integrated lignocellulose biorefineries. New Biotechnol. 2019, 49, 161–168. [Google Scholar] [CrossRef]
- Azeez, M.A.; Orege, J.I. Bamboo, Its Chemical Modification and Products. In Bamboo—Current and Future Prospects; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Gabhane, J.; William, S.P.; Gadhe, A.; Rath, R.; Vaidya, A.N.; Wate, S. Pretreatment of banana agricultural waste for bio-ethanol production: Individual and interactive effects of acid and alkali pretreatments with autoclaving, microwave heating and ultrasonication. Waste Manag. 2014, 34, 498–503. [Google Scholar] [CrossRef]
- Díaz, M.J.; Moya, M.; Castro, E. Bioethanol Production from Steam-Exploded Barley Straw by Co-Fermentation with Escherichia coli SL100. Agronomy 2022, 12, 874. [Google Scholar] [CrossRef]
- Nigam, P.S.N.; Gupta, N.; Anthwal, A. Pre-treatment of Agro-Industrial Residues. In Biotechnology for Agro-Industrial Residues Utilisation; Springer: Dordrecht, The Netherlands, 2009; pp. 13–33. [Google Scholar] [CrossRef]
- Saini, J.K.; Saini, R.; Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. 3 Biotech 2015, 5, 337–353. [Google Scholar] [CrossRef]
- Soni, S.K.; Sharma, A.; Soni, R. Microbial Enzyme Systems in the Production of Second Generation Bioethanol. Sustainability 2023, 15, 3590. [Google Scholar] [CrossRef]
- Chandra, R.; Takeuchi, H.; Hasegawa, T. Methane production from lignocellulosic agricultural crop wastes: A review in context to second generation of biofuel production. Renew. Sustain. Energy Rev. 2012, 16, 1462–1476. [Google Scholar] [CrossRef]
- Kim, M.; Day, D.F. Composition of sugar cane, energy cane, and sweet sorghum suitable for ethanol production at Louisiana sugar mills. J. Ind. Microbiol. Biotechnol. 2011, 38, 803–807. [Google Scholar] [CrossRef]
- Xu, P.; Cheng, S.; Han, Y.; Zhao, D.; Li, H.; Wang, Y.; Zhang, G.; Chen, C. Natural Variation of Lignocellulosic Components in Miscanthus Biomass in China. Front. Chem. 2020, 8, 595143. [Google Scholar] [CrossRef]
- Rajendran, K.; Drielak, E.; Varma, V.S.; Muthusamy, S.; Kumar, G. Updates on the pretreatment of lignocellulosic feedstocks for bioenergy production—A review. Biomass Convers. Biorefin. 2018, 8, 471–483. [Google Scholar] [CrossRef]
- Du, B.; Sharma, L.N.; Becker, C.; Chen, S.; Mowery, R.A.; van Walsum, G.P.; Chambliss, C.K. Effect of varying feedstock–pretreatment chemistry combinations on the formation and accumulation of potentially inhibitory degradation products in biomass hydrolysates. Biotechnol. Bioeng. 2010, 107, 430–440. [Google Scholar] [CrossRef]
- García-Cubero, M.T.; González-Benito, G.; Indacoechea, I.; Coca, M.; Bolado, S. Effect of ozonolysis pretreatment on enzymatic digestibility of wheat and rye straw. Bioresour. Technol. 2009, 100, 1608–1613. [Google Scholar] [CrossRef]
- Monlau, F.; Barakat, A.; Steyer, J.P.; Carrere, H. Comparison of seven types of thermo-chemical pretreatments on the structural features and anaerobic digestion of sunflower stalks. Bioresour. Technol. 2012, 120, 241–247. [Google Scholar] [CrossRef]
- Brigham, C. Biopolymers. In Green Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 753–770. [Google Scholar] [CrossRef]
- Damayanti, D.; Supriyadi, D.; Amelia, D.; Saputri, D.R.; Devi, Y.L.L.; Auriyani, W.A.; Wu, H.S. Conversion of Lignocellulose for Bioethanol Production, Applied in Bio-Polyethylene Terephthalate. Polymers 2021, 13, 2886. [Google Scholar] [CrossRef]
- Frassoldati, A.; Ranzi, E. Modeling of Thermochemical Conversion of Biomasses. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Hokkanen, S.; Bhatnagar, A.; Sillanpää, M. A review on modification methods to cellulose-based adsorbents to improve adsorption capacity. Water Res. 2016, 91, 156–173. [Google Scholar] [CrossRef]
- Banu, J.R.; Preethi; Kavitha, S.; Tyagi, V.K.; Gunasekaran, M.; Karthikeyan, O.P.; Kumar, G. Lignocellulosic biomass based biorefinery: A successful platform towards circular bioeconomy. Fuel 2021, 302, 121086. [Google Scholar] [CrossRef]
- Zhou, X.; Li, W.; Mabon, R.; Broadbelt, L.J. A Critical Review on Hemicellulose Pyrolysis. Energy Technol. 2017, 5, 52–79. [Google Scholar] [CrossRef]
- Bajpai, P. Wood and Fiber Fundamentals. In Biermann’s Handbook of Pulp and Paper; Elsevier: Amsterdam, The Netherlands, 2018; pp. 19–74. [Google Scholar] [CrossRef]
- Malik, K.; Sharma, P.; Yang, Y.; Zhang, P.; Zhang, L.; Xing, X.; Yue, J.; Song, Z.; Nan, L.; Yujun, S.; et al. Lignocellulosic biomass for bioethanol: Insight into the advanced pretreatment and fermentation approaches. Ind. Crops Prod. 2022, 188, 115569. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, A. Lignin; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Akpan, E.I. Chemistry and Structure of Lignin. In Sustainable Lignin for Carbon Fibers: Principles, Techniques, and Applications; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–50. [Google Scholar] [CrossRef]
- Lu, Y.; Lu, Y.-C.; Hu, H.-Q.; Xie, F.-J.; Wei, X.-Y.; Fan, X. Structural Characterization of Lignin and Its Degradation Products with Spectroscopic Methods. J. Spectrosc. 2017, 2017, 8951658. [Google Scholar] [CrossRef]
- Hou, J.; Tang, J.; Chen, J.; Zhang, Q. Quantitative Structure-Toxicity Relationship analysis of combined toxic effects of lignocellulose-derived inhibitors on bioethanol production. Bioresour. Technol. 2019, 289, 121724. [Google Scholar] [CrossRef]
- Kordala, N.; Lewandowska, M.; Bednarski, W. Effect of the method for the elimination of inhibitors present in Miscanthus giganteus hydrolysates on ethanol production effectiveness. Biomass Convers. Biorefin. 2023, 13, 2089–2097. [Google Scholar] [CrossRef]
- Kim, D. Physico-Chemical Conversion of Lignocellulose: Inhibitor Effects and Detoxification Strategies: A Mini Review. Molecules 2018, 23, 309. [Google Scholar] [CrossRef]
- Jeswani, H.K.; Chilvers, A.; Azapagic, A. Environmental sustainability of biofuels: A review. Proc. R Soc. A Math. Phys. Eng. Sci. 2020, 476, 20200351. [Google Scholar] [CrossRef]
- Rezania, S.; Oryani, B.; Cho, J.; Talaiekhozani, A.; Sabbagh, F.; Hashemi, B.; Rupani, P.F.; Mohammadi, A.A. Different pretreatment technologies of lignocellulosic biomass for bioethanol production: An overview. Energy 2020, 199, 117457. [Google Scholar] [CrossRef]
- Das, N.; Jena, P.K.; Padhi, D.; Mohanty, M.K.; Sahoo, G. A comprehensive review of characterization, pretreatment and its applications on different lignocellulosic biomass for bioethanol production. Biomass Convers. Biorefin. 2023, 13, 1503–1527. [Google Scholar] [CrossRef]
- Hideno, A.; Inoue, H.; Tsukahara, K.; Fujimoto, S.; Minowa, T.; Inoue, S.; Endo, T.; Sawayama, S. Wet disk milling pretreatment without sulfuric acid for enzymatic hydrolysis of rice straw. Bioresour. Technol. 2009, 100, 2706–2711. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.R.; Fujimoto, S.; Hirata, S.; Hassan, M.A. Ball Milling Pretreatment of Oil Palm Biomass for Enhancing Enzymatic Hydrolysis. Appl. Biochem. Biotechnol. 2014, 173, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, W.; Xia, Q.; Guo, B.; Wang, Q.; Liu, S.; Liu, Y.; Li, J.; Yu, H. Efficient Cleavage of Lignin–Carbohydrate Complexes and Ultrafast Extraction of Lignin Oligomers from Wood Biomass by Microwave-Assisted Treatment with Deep Eutectic Solvent. ChemSusChem 2017, 10, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- John, I.; Pola, J.; Appusamy, A. Optimization of Ultrasonic Assisted Saccharification of Sweet Lime Peel for Bioethanol Production Using Box–Behnken Method. Waste Biomass Valorization 2019, 10, 441–453. [Google Scholar] [CrossRef]
- Bonfiglio, F.; Cagno, M.; Rey, F.; Torres, M.; Böthig, S.; Menéndez, P.; Mussatto, S.I. Pretreatment of switchgrass by steam explosion in a semi-continuous pre-pilot reactor. Biomass Bioenergy 2019, 121, 41–47. [Google Scholar] [CrossRef]
- De Carvalho, D.M.; de Queiroz, J.H.; Colodette, J.L. Assessment of alkaline pretreatment for the production of bioethanol from eucalyptus, sugarcane bagasse and sugarcane straw. Ind. Crops Prod. 2016, 94, 932–941. [Google Scholar] [CrossRef]
- Hilares, R.T.; Swerts, M.P.; Ahmed, M.A.; Ramos, L.; da Silva, S.S.; Santos, J.C. Organosolv Pretreatment of Sugar Cane Bagasse for Bioethanol Production. Ind. Eng. Chem. Res. 2017, 56, 3833–3838. [Google Scholar] [CrossRef]
- Hossain, A.; Rahaman, M.S.; Yelle, D.; Shang, H.; Sun, Z.; Renneckar, S.; Dong, J.; Tulaphol, S.; Sathitsuksanoh, N. Effects of polyol-based deep eutectic solvents on the efficiency of rice straw enzymatic hydrolysis. Ind. Crops Prod. 2021, 167, 113480. [Google Scholar] [CrossRef]
- Singh, A.; Bishnoi, N.R. Enzymatic hydrolysis optimization of microwave alkali pretreated wheat straw and ethanol production by yeast. Bioresour. Technol. 2012, 108, 94–101. [Google Scholar] [CrossRef]
- Ziaei-Rad, Z.; Pazouki, M.; Fooladi, J.; Azin, M.; Gummadi, S.N.; Allahverdi, A. Investigation of a robust pretreatment technique based on ultrasound-assisted, cost-effective ionic liquid for enhancing saccharification and bioethanol production from wheat straw. Sci. Rep. 2023, 13, 446. [Google Scholar] [CrossRef]
- Yang, J.; Lu, X.; Zhang, Y.; Xu, J.; Yang, Y.; Zhou, Q. A facile ionic liquid approach to prepare cellulose fiber with good mechanical properties directly from corn stalks. Green Energy Environ. 2020, 5, 223–231. [Google Scholar] [CrossRef]
- Padilha, C.E.d.A.; Nogueira, C.d.C.; Souza, D.F.d.S.; de Oliveira, J.A.; dos Santos, E.S. Valorization of green coconut fibre: Use of the black liquor of organolsolv pretreatment for ethanol production and the washing water for production of rhamnolipids by Pseudomonas aeruginosa ATCC 27583. Ind. Crops Prod. 2019, 140, 111604. [Google Scholar] [CrossRef]
- Martínez-Patiño, J.C.; Ruiz, E.; Romero, I.; Cara, C.; López-Linares, J.C.; Castro, E. Combined acid/alkaline-peroxide pretreatment of olive tree biomass for bioethanol production. Bioresour. Technol. 2017, 239, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Lee, Y.Y.; Sunwoo, C.; Kim, J.S. Pretreatment of Corn Stover by Low-Liquid Ammonia Recycle Percolation Process. Appl. Biochem. Biotechnol. 2006, 133, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.; Greetham, D.; Mos, M.; Squance, M.; Kam, J.; Du, C. Exploring the Bioethanol Production Potential of Miscanthus Cultivars. Appl. Sci. 2021, 11, 9949. [Google Scholar] [CrossRef]
- Kumar, M.N.; Ravikumar, R.; Sankar, M.K.; Thenmozhi, S. New insight into the effect of fungal mycelia present in the bio-pretreated paddy straw on their enzymatic saccharification and optimization of process parameters. Bioresour. Technol. 2018, 267, 291–302. [Google Scholar] [CrossRef]
- Lee, O.K.; Oh, Y.-K.; Lee, E.Y. Bioethanol production from carbohydrate-enriched residual biomass obtained after lipid extraction of Chlorella sp. KR-1. Bioresour. Technol. 2015, 196, 22–27. [Google Scholar] [CrossRef]
- Guo, H.; Daroch, M.; Liu, L.; Qiu, G.; Geng, S.; Wang, G. Biochemical features and bioethanol production of microalgae from coastal waters of Pearl River Delta. Bioresour. Technol. 2013, 127, 422–428. [Google Scholar] [CrossRef]
- Kumar, V.; Yadav, S.K.; Kumar, J.; Ahluwalia, V. A critical review on current strategies and trends employed for removal of inhibitors and toxic materials generated during biomass pretreatment. Bioresour. Technol. 2020, 299, 122633. [Google Scholar] [CrossRef]
- Zhao, J.; Lee, J.; Wang, D. A Critical Review on Water Overconsumption in Lignocellulosic Biomass Pretreatment for Ethanol Production through Enzymic Hydrolysis and Fermentation. Energy Fuels 2023, 37, 2667–2680. [Google Scholar] [CrossRef]
- Ranjithkumar, M.; Uthandi, S.; Kumar, P.S.; Muniraj, I.; Thanabal, V.; Rajarathinam, R. Highly crystalline cotton spinning wastes utilization: Pretreatment, optimized hydrolysis and fermentation using Pleurotus florida for bioethanol production. Fuel 2022, 308, 122052. [Google Scholar] [CrossRef]
- Li, F.; Liu, Y.; Jia, J.; Yu, H. Combination of biological pretreatment with deep eutectic solvent pretreatment for enhanced enzymatic saccharification of Pinus massoniana. Bioresour. Technol. 2023, 380, 129110. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Zhang, Y.; Yong, C.; Qu, P.; Huang, H.; Xu, Y. Biological fermentation pretreatment accelerated the depolymerization of straw fiber and its mechanical properties as raw material for mulch film. J. Clean. Prod. 2021, 284, 124688. [Google Scholar] [CrossRef]
- Uzuner, S.; Cekmecelioglu, D. Enzymes in the Beverage Industry. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 29–43. [Google Scholar] [CrossRef]
- Suganuma, S.; Nakajima, K.; Kitano, M.; Yamaguchi, D.; Kato, H.; Hayashi, S.; Hara, M. Hydrolysis of Cellulose by Amorphous Carbon Bearing SO3H, COOH, and OH Groups. J. Am. Chem. Soc. 2008, 130, 12787–12793. [Google Scholar] [CrossRef] [PubMed]
- Chen, H. Lignocellulose biorefinery feedstock engineering. In Lignocellulose Biorefinery Engineering, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 37–86. [Google Scholar] [CrossRef]
- Wada, M.; Ike, M.; Tokuyasu, K. Enzymatic hydrolysis of cellulose I is greatly accelerated via its conversion to the cellulose II hydrate form. Polym. Degrad. Stab. 2010, 95, 543–548. [Google Scholar] [CrossRef]
- Vasić, K.; Knez, Ž.K.; Leitgeb, M. Bioethanol Production by Enzymatic Hydrolysis from Different Lignocellulosic Sources. Molecules 2021, 26, 753. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, M.; Pande, A.; Marathe, A. Different particle size study of castor deoiled cake for biofuel production with an environmental sustainability perspective. Heliyon 2022, 8, e09710. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Semwal, S.; Satlewal, A.; Christopher, J.; Gupta, R.P.; Kumar, R.; Puri, S.K.; Ramakumar, S. The impact of particle size of cellulosic residue and solid loadings on enzymatic hydrolysis with a mass balance. Fuel 2019, 245, 514–520. [Google Scholar] [CrossRef]
- Gao, W.; Li, Z.; Liu, T.; Wang, Y. Production of high-concentration fermentable sugars from lignocellulosic biomass by using high solids fed-batch enzymatic hydrolysis. Biochem. Eng. J. 2021, 176, 108186. [Google Scholar] [CrossRef]
- Modenbach, A.A.; Nokes, S.E. Enzymatic hydrolysis of biomass at high-solids loadings—A review. Biomass Bioenergy 2013, 56, 526–544. [Google Scholar] [CrossRef]
- Amândio, M.S.T.; Rocha, J.M.S.; Xavier, A.M.R.B. Enzymatic Hydrolysis Strategies for Cellulosic Sugars Production to Obtain Bioethanol from Eucalyptus globulus Bark. Fermentation 2023, 9, 241. [Google Scholar] [CrossRef]
- Constantino, A.; Rodrigues, B.; Leon, R.; Barros, R.; Raposo, S. Alternative chemo-enzymatic hydrolysis strategy applied to different microalgae species for bioethanol production. Algal Res. 2021, 56, 102329. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, C.; Tong, S.; Cui, Z.; Liu, P. Enhanced Enzymatic Hydrolysis and Structural Features of Corn Stover by NaOH and Ozone Combined Pretreatment. Molecules 2018, 23, 1300. [Google Scholar] [CrossRef] [PubMed]
- Ostadjoo, S.; Hammerer, F.; Dietrich, K.; Dumont, M.-J.; Friščić, T.; Auclair, K. Efficient Enzymatic Hydrolysis of Biomass Hemicellulose in the Absence of Bulk Water. Molecules 2019, 24, 4206. [Google Scholar] [CrossRef] [PubMed]
- Godbey, W.T. Fermentation, Beer, and Biofuels. In An Introduction to Biotechnology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 331–351. [Google Scholar] [CrossRef]
- Spector, M.P. Metabolism, Central (Intermediary). In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 242–264. [Google Scholar] [CrossRef]
- D’Amore, T.; Panchal, C.J.; Russell, I.; Stewart, G.G. A Study of Ethanol Tolerance in Yeast. Crit. Rev. Biotechnol. 1989, 9, 287–304. [Google Scholar] [CrossRef]
- Tse, T.J.; Wiens, D.J.; Reaney, M.J.T. Production of Bioethanol—A Review of Factors Affecting Ethanol Yield. Fermentation 2021, 7, 268. [Google Scholar] [CrossRef]
- Li, C.; Liu, Q.; Wang, Y.; Yang, X.; Chen, S.; Zhao, Y.; Wu, Y.; Li, L. Salt stress improves thermotolerance and high-temperature bioethanol production of multi-stress-tolerant Pichia kudriavzevii by stimulating intracellular metabolism and inhibiting oxidative damage. Biotechnol. Biofuels 2021, 14, 222. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Karimi, K. Fermentation Inhibitors in Ethanol Processes and Different Strategies to Reduce Their Effects. In Biofuels; Elsevier: Amsterdam, The Netherlands, 2011; pp. 287–311. [Google Scholar] [CrossRef]
- Varize, C.S.; Bücker, A.; Lopes, L.D.; Christofoleti-Furlan, R.M.; Raposo, M.S.; Basso, L.C.; Stambuk, B.U. Increasing Ethanol Tolerance and Ethanol Production in an Industrial Fuel Ethanol Saccharomyces cerevisiae Strain. Fermentation 2022, 8, 470. [Google Scholar] [CrossRef]
- Rastogi, M.; Shrivastava, S. Recent advances in second generation bioethanol production: An insight to pretreatment, saccharification and fermentation processes. Renew. Sustain. Energy Rev. 2017, 80, 330–340. [Google Scholar] [CrossRef]
- Sharma, S.; Nair, A.; Sarma, S.J. Biorefinery concept of simultaneous saccharification and co-fermentation: Challenges and improvements. Chem. Eng. Process. Process Intensif. 2021, 169, 108634. [Google Scholar] [CrossRef]
- Chu, Q.; Li, X.; Ma, B.; Xu, Y.; Ouyang, J.; Zhu, J.; Yu, S.; Yong, Q. Bioethanol production: An integrated process of low substrate loading hydrolysis-high sugars liquid fermentation and solid state fermentation of enzymatic hydrolysis residue. Bioresour. Technol. 2012, 123, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Gomes, D.; Cruz, M.; de Resende, M.; Ribeiro, E.; Teixeira, J.; Domingues, L. Very High Gravity Bioethanol Revisited: Main Challenges and Advances. Fermentation 2021, 7, 38. [Google Scholar] [CrossRef]
- Cruz, M.L.; de Resende, M.M.; Ribeiro, E.J. Improvement of ethanol production in fed-batch fermentation using a mixture of sugarcane juice and molasse under very high-gravity conditions. Bioprocess Biosyst. Eng. 2021, 44, 617–625. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chang, K.-S.; Chen, C.-Y.; Hsu, C.-L.; Chang, T.-C.; Jang, H.-D. Enhancement of the Efficiency of Bioethanol Production by Saccharomyces cerevisiae via Gradually Batch-Wise and Fed-Batch Increasing the Glucose Concentration. Fermentation 2018, 4, 45. [Google Scholar] [CrossRef]
- Kumari, M.; Gupta, S.K. Response surface methodological (RSM) approach for optimizing the removal of trihalomethanes (THMs) and its precursor’s by surfactant modified magnetic nanoadsorbents (sMNP)—An endeavor to diminish probable cancer risk. Sci. Rep. 2019, 9, 18339. [Google Scholar] [CrossRef]
- Assabjeu, A.C.; Noubissié, E.; Desobgo, S.C.Z.; Ali, A. Optimization of the enzymatic hydrolysis of cellulose of triplochiton scleroxylon sawdust in view of the production of bioethanol. Sci. Afr. 2020, 8, e00438. [Google Scholar] [CrossRef]
- El-Mekkawi, S.A.; Abdo, S.M.; Samhan, F.A.; Ali, G.H. Optimization of some fermentation conditions for bioethanol production from microalgae using response surface method. Bull. Natl. Res. Cent. 2019, 43, 164. [Google Scholar] [CrossRef]
- Žnidaršič-Plazl, P. The Promises and the Challenges of Biotransformations in Microflow. Biotechnol. J. 2019, 14, e1800580. [Google Scholar] [CrossRef]
- Hernández-Beltrán, J.U.; Fontalvo, J.; Hernández-Escoto, H. Fed-batch enzymatic hydrolysis of plantain pseudostem to fermentable sugars production and the impact of particle size at high solids loadings. Biomass Convers. Biorefin. 2021, 11, 2975–2982. [Google Scholar] [CrossRef]
- Van Zyl, W.H.; Lynd, L.R.; den Haan, R.; McBride, J.E. Consolidated Bioprocessing for Bioethanol Production Using Saccharomyces cerevisiae. In Biofuels; Springer: Berlin/Heidelberg, Germany, 2007; pp. 205–235. [Google Scholar] [CrossRef]
- Maleki, F.; Changizian, M.; Zolfaghari, N.; Rajaei, S.; Noghabi, K.A.; Zahiri, H.S. Consolidated bioprocessing for bioethanol production by metabolically engineered Bacillus subtilis strains. Sci. Rep. 2021, 11, 13731. [Google Scholar] [CrossRef]
- Pinaki, D.; Lhakpa, W.; Joginder, S. Simultaneous Saccharification and Fermentation (SSF), An Efficient Process for Bio-Ethanol Production: An Overview. Biosci. Biotechnol. Res. Asia 2015, 12, 87–100. [Google Scholar] [CrossRef]
- Derman, E.; Abdulla, R.; Marbawi, H.; Sabullah, M.K.; Gansau, J.A.; Ravindra, P. Simultaneous Saccharification and Fermentation of Empty Fruit Bunches of Palm for Bioethanol Production Using a Microbial Consortium of S. cerevisiae and T. harzianum. Fermentation 2022, 8, 295. [Google Scholar] [CrossRef]
- Adekunle, A.E.; Rabeya, T.; Jehadin, F.; Asad, M.A.; Ayodele, O.O.; Islam, S. Compressed hot water pretreatment enhanced bioethanol production from corn stalk. Bioresour. Technol. Rep. 2020, 12, 100595. [Google Scholar] [CrossRef]
- Fetyan, N.A.H.; El-Sayed, A.E.-K.B.; Ibrahim, F.M.; Attia, Y.A.; Sadik, M.W. Bioethanol production from defatted biomass of Nannochloropsis oculata microalgae grown under mixotrophic conditions. Environ. Sci. Pollut. Res. 2022, 29, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Jugwanth, Y.; Sewsynker-Sukai, Y.; Kana, E.G. Valorization of sugarcane bagasse for bioethanol production through simultaneous saccharification and fermentation: Optimization and kinetic studies. Fuel 2020, 262, 116552. [Google Scholar] [CrossRef]
- Chohan, N.A.; Aruwajoye, G.; Sewsynker-Sukai, Y.; Kana, E.G. Valorisation of potato peel wastes for bioethanol production using simultaneous saccharification and fermentation: Process optimization and kinetic assessment. Renew. Energy 2020, 146, 1031–1040. [Google Scholar] [CrossRef]
- Kumar, D.; Surya, K.; Verma, R. Bioethanol production from apple pomace using co-cultures with Saccharomyces cerevisiae in solid-state fermentation. J. Microbiol. Biotechnol. Food Sci. 2020, 9, 742–745. [Google Scholar] [CrossRef]
- Efunwoye, O.O.; Oluwole, O.R. Bioethanol Production from Pineapple Waste by Solid State Fermentation Method. 2019. Available online: www.nsmjournal.org (accessed on 19 December 2023).
- Prasoulas, G.; Gentikis, A.; Konti, A.; Kalantzi, S.; Kekos, D.; Mamma, D. Bioethanol Production from Food Waste Applying the Multienzyme System Produced On-Site by Fusarium oxysporum F3 and Mixed Microbial Cultures. Fermentation 2020, 6, 39. [Google Scholar] [CrossRef]
- Camargos, C.V.; Moraes, V.D.; de Oliveira, L.M.; Guidini, C.Z.; Ribeiro, E.J.; Santos, L.D. High Gravity and Very High Gravity Fermentation of Sugarcane Molasses by Flocculating Saccharomyces cerevisiae: Experimental Investigation and Kinetic Modeling. Appl. Biochem. Biotechnol. 2021, 193, 807–821. [Google Scholar] [CrossRef]
- Granjo, J.F.; Nunes, D.S.; Duarte, B.P.; Oliveira, N.M. A comparison of process alternatives for energy-efficient bioethanol downstream processing. Sep. Purif. Technol. 2020, 238, 116414. [Google Scholar] [CrossRef]
- Bisgaard, T.; Mauricio-Iglesias, M.; Huusom, J.K.; Gernaey, K.V.; Dohrup, J.; Petersen, M.A.; Abildskov, J. Adding Value to Bioethanol through a Purification Process Revamp. Ind. Eng. Chem. Res. 2017, 56, 5692–5704. [Google Scholar] [CrossRef]
- Jhariya, U.; Dafale, N.A.; Srivastava, S.; Bhende, R.S.; Kapley, A.; Purohit, H.J. Understanding Ethanol Tolerance Mechanism in Saccharomyces cerevisiae to Enhance the Bioethanol Production: Current and Future Prospects. BioEnergy Res. 2021, 14, 670–688. [Google Scholar] [CrossRef]
- Wu, M.; Li, H.; Wei, S.; Wu, H.; Wu, X.; Bao, X.; Hou, J.; Liu, W.; Shen, Y. Simulating Extracellular Glucose Signals Enhances Xylose Metabolism in Recombinant Saccharomyces cerevisiae. Microorganisms 2020, 8, 100. [Google Scholar] [CrossRef]
- Kwak, S.; Jin, Y.-S. Production of fuels and chemicals from xylose by engineered Saccharomyces cerevisiae: A review and perspective. Microb. Cell Factories 2017, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, T.; Peng, B.; Geng, A. Enhanced ethanol production from industrial lignocellulose hydrolysates by a hydrolysate-cofermenting Saccharomyces cerevisiae strain. Bioprocess Biosyst. Eng. 2019, 42, 883–896. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, S.; Gao, X.; Li, M.; Li, D.; Lu, W. Enhanced protopanaxadiol production from xylose by engineered Yarrowia lipolytica. Microb. Cell Factories 2019, 18, 83. [Google Scholar] [CrossRef]
- Cunha, J.T.; Soares, P.O.; Baptista, S.L.; Costa, C.E.; Domingues, L. Engineered Saccharomyces cerevisiae for lignocellulosic valorization: A review and perspectives on bioethanol production. Bioengineered 2020, 11, 883–903. [Google Scholar] [CrossRef]
- Li, H.; Shen, Y.; Wu, M.; Hou, J.; Jiao, C.; Li, Z.; Liu, X.; Bao, X. Engineering a wild-type diploid Saccharomyces cerevisiae strain for second-generation bioethanol production. Bioresour. Bioprocess. 2016, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Romaní, A.; Pereira, F.; Johansson, B.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae ethanol strains PE-2 and CAT-1 for efficient lignocellulosic fermentation. Bioresour. Technol. 2015, 179, 150–158. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Sahara, T.; Ohgiya, S.; Kamagata, Y.; Fujimori, K.E. Systematic optimization of gene expression of pentose phosphate pathway enhances ethanol production from a glucose/xylose mixed medium in a recombinant Saccharomyces cerevisiae. AMB Express 2018, 8, 139. [Google Scholar] [CrossRef]
- Papapetridis, I.; Verhoeven, M.D.; Wiersma, S.J.; Goudriaan, M.; van Maris, A.A.J.; Pronk, J.T. Laboratory evolution for forced glucose-xylose co-consumption enables identification of mutations that improve mixed-sugar fermentation by xylose-fermenting Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, foy056. [Google Scholar] [CrossRef]
- Wu, R.; Chen, D.; Cao, S.; Lu, Z.; Huang, J.; Lu, Q.; Chen, Y.; Chen, X.; Guan, N.; Wei, Y.; et al. Enhanced ethanol production from sugarcane molasses by industrially engineered Saccharomyces cerevisiae via replacement of the PHO4 gene. RSC Adv. 2020, 10, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.K.; Enkh-Amgalan, T.; Gong, G.; Um, Y.; Lee, S. Improved bioconversion of lignocellulosic biomass by Saccharomyces cerevisiae engineered for tolerance to acetic acid. GCB Bioenergy 2020, 12, 90–100. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, S.; Lu, S.; Jiang, S.; Jiang, S.; Deng, Y.; Lu, J.; Wang, H.; Zhou, Y. Ethanol yield improvement in Saccharomyces cerevisiae GPD2 Delta FPS1 Delta ADH2 Delta DLD3 Delta mutant and molecular mechanism exploration based on the metabolic flux and transcriptomics approaches. Microb. Cell Factories 2022, 21, 160. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wu, Y.; Zheng, Z.; Cao, L.; Zhu, X.; Mu, D.; Jiang, S. CRISPR-Cas9 Approach Constructing Cellulase sestc-Engineered Saccharomyces cerevisiae for the Production of Orange Peel Ethanol. Front. Microbiol. 2018, 9, 2436. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Zhang, W.; Liu, T.; Tan, G.; Li, H.; Huang, Z. Improvement of Ethanol Production in Saccharomyces cerevisiae by High-Efficient Disruption of the ADH2 Gene Using a Novel Recombinant TALEN Vector. Front. Microbiol. 2016, 7, 1067. [Google Scholar] [CrossRef]
- Lee, Y.-G.; Jin, Y.-S.; Cha, Y.-L.; Seo, J.-H. Bioethanol production from cellulosic hydrolysates by engineered industrial Saccharomyces cerevisiae. Bioresour. Technol. 2017, 228, 355–361. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Wang, L.; Yang, R.; Hou, P.; Liu, J. Direct bioethanol production from wheat straw using xylose/glucose co-fermentation by co-culture of two recombinant yeasts. J. Ind. Microbiol. Biotechnol. 2017, 44, 453–464. [Google Scholar] [CrossRef]
- Chen, L.; Du, J.-L.; Zhan, Y.-J.; Li, J.-A.; Zuo, R.-R.; Tian, S. Consolidated bioprocessing for cellulosic ethanol conversion by cellulase–xylanase cell-surfaced yeast consortium. Prep. Biochem. Biotechnol. 2018, 48, 653–661. [Google Scholar] [CrossRef]
- Cunha, J.T.; Romaní, A.; Inokuma, K.; Johansson, B.; Hasunuma, T.; Kondo, A.; Domingues, L. Consolidated bioprocessing of corn cob-derived hemicellulose: Engineered industrial Saccharomyces cerevisiae as efficient whole cell biocatalysts. Biotechnol. Biofuels 2020, 13, 138. [Google Scholar] [CrossRef]
- Sasaki, Y.; Takagi, T.; Motone, K.; Kuroda, K.; Ueda, M. Enhanced direct ethanol production by cofactor optimization of cell surface-displayed xylose isomerase in yeast. Biotechnol. Prog. 2017, 33, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Hasunuma, T.; Hori, Y.; Yamada, R.; Kondo, A. Direct ethanol production from hemicellulosic materials of rice straw by use of an engineered yeast strain codisplaying three types of hemicellulolytic enzymes on the surface of xylose-utilizing Saccharomyces cerevisiae cells. J. Biotechnol. 2012, 158, 203–210. [Google Scholar] [CrossRef]
- Chin, Y.-W.; Kang, W.-K.; Jang, H.W.; Turner, T.L.; Kim, H.J. CAR1 deletion by CRISPR/Cas9 reduces formation of ethyl carbamate from ethanol fermentation by Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2016, 43, 1517–1525. [Google Scholar] [CrossRef]
- Xue, T.; Liu, K.; Chen, D.; Yuan, X.; Fang, J.; Yan, H.; Huang, L.; Chen, Y.; He, W. Improved bioethanol production using CRISPR/Cas9 to disrupt the ADH2 gene in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2018, 34, 154. [Google Scholar] [CrossRef]
- Liu, K.; Yuan, X.; Liang, L.; Fang, J.; Chen, Y.; He, W.; Xue, T. Using CRISPR/Cas9 for multiplex genome engineering to optimize the ethanol metabolic pathway in Saccharomyces cerevisiae. Biochem. Eng. J. 2019, 145, 120–126. [Google Scholar] [CrossRef]
- Wang, L.; Li, B.; Su, R.-R.; Wang, S.-P.; Xia, Z.-Y.; Xie, C.-Y.; Tang, Y.-Q. Screening novel genes by a comprehensive strategy to construct multiple stress-tolerant industrial Saccharomyces cerevisiae with prominent bioethanol production. Biotechnol. Biofuels Bioprod. 2022, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Jiang, S.; Jiang, S.; Lu, S.; Zheng, Z.; Chen, J.; Wu, W.; Jiang, S. CRISPR-Cas9 Approach Constructed Engineered Saccharomyces cerevisiae with the Deletion of GPD2, FPS1, and ADH2 to Enhance the Production of Ethanol. J. Fungi 2022, 8, 703. [Google Scholar] [CrossRef] [PubMed]
- Gronchi, N.; De Bernardini, N.; Cripwell, R.A.; Treu, L.; Campanaro, S.; Basaglia, M.; Foulquié-Moreno, M.R.; Thevelein, J.M.; Van Zyl, W.H.; Favaro, L.; et al. Natural Saccharomyces cerevisiae Strain Reveals Peculiar Genomic Traits for Starch-to-Bioethanol Production: The Design of an Amylolytic Consolidated Bioprocessing Yeast. Front. Microbiol. 2022, 12, 768562. [Google Scholar] [CrossRef]
- Zhang, K.; Lu, X.; Li, Y.; Jiang, X.; Liu, L.; Wang, H. New technologies provide more metabolic engineering strategies for bioethanol production in Zymomonas mobilis. Appl. Microbiol. Biotechnol. 2019, 103, 2087–2099. [Google Scholar] [CrossRef]
- Wang, S.; Tian, R.; Liu, B.; Wang, H.; Liu, J.; Li, C.; Li, M.; Evivie, S.E.; Li, B. Effects of carbon concentration, oxygen, and controlled pH on the engineering strain Lactiplantibacillus casei E1 in the production of bioethanol from sugarcane molasses. AMB Express 2021, 11, 95. [Google Scholar] [CrossRef]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Huang, C.-C. Kluyveromyces marxianus: Current State of Omics Studies, Strain Improvement Strategy and Potential Industrial Implementation. Fermentation 2020, 6, 124. [Google Scholar] [CrossRef]
- Huang, J.; Xia, T.; Li, G.; Li, X.; Li, Y.; Wang, Y.; Wang, Y.; Chen, Y.; Xie, G.; Bai, F.-W.; et al. Overproduction of native endo-β-1,4-glucanases leads to largely enhanced biomass saccharification and bioethanol production by specific modification of cellulose features in transgenic rice. Biotechnol. Biofuels 2019, 12, 11. [Google Scholar] [CrossRef]
- Schmatz, A.A.; Tyhoda, L.; Brienzo, M. Sugarcane biomass conversion influenced by lignin. Biofuels Bioprod. Biorefin. 2020, 14, 469–480. [Google Scholar] [CrossRef]
- Bewg, W.P.; Poovaiah, C.; Lan, W.; Ralph, J.; Coleman, H.D. RNAi downregulation of three key lignin genes in sugarcane improves glucose release without reduction in sugar production. Biotechnol. Biofuels 2016, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Meneses, L.; Ferreira, J.A.; Mushtaq, M.; Karimi, S.; Orupõld, K.; Kikas, T. Genetic modification of cereal plants: A strategy to enhance bioethanol yields from agricultural waste. Ind. Crops Prod. 2020, 150, 112408. [Google Scholar] [CrossRef]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.-G.; Ventura, V.; et al. RNAi: What is its position in agriculture? J. Pest Sci. 2004, 93, 1125–1130. [Google Scholar] [CrossRef]
- De Souza, W.R.; Pacheco, T.F.; Duarte, K.E.; Sampaio, B.L.; Molinari, P.A.d.O.; Martins, P.K.; Santiago, T.R.; Formighieri, E.F.; Vinecky, F.; Ribeiro, A.P.; et al. Silencing of a BAHD acyltransferase in sugarcane increases biomass digestibility. Biotechnol. Biofuels 2019, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Poovaiah, C.R.; Bewg, W.P.; Lan, W.; Ralph, J.; Coleman, H.D. Sugarcane transgenics expressing MYB transcription factors show improved glucose release. Biotechnol. Biofuels 2016, 9, 143. [Google Scholar] [CrossRef]
- Huang, L.; Liu, Y.; Su, S.; Lai, C.; Wu, C.; Chao, T.; Yang, Y. Genetic engineering of transitory starch accumulation by knockdown of OsSEX4 in rice plants for enhanced bioethanol production. Biotechnol. Bioeng. 2020, 117, 933–944. [Google Scholar] [CrossRef]
- Ai, Y.; Feng, S.; Wang, Y.; Lu, J.; Sun, M.; Hu, H.; Hu, Z.; Zhang, R.; Liu, P.; Peng, H.; et al. Integrated genetic and chemical modification with rice straw for maximum bioethanol production. Ind. Crops Prod. 2021, 173, 114133. [Google Scholar] [CrossRef]
- Voorend, W.; Nelissen, H.; Vanholme, R.; De Vliegher, A.; Van Breusegem, F.; Boerjan, W.; Roldán-Ruiz, I.; Muylle, H.; Inzé, D. Overexpression of GA20-OXIDASE1 impacts plant height, biomass allocation and saccharification efficiency in maize. Plant Biotechnol. J. 2016, 14, 997–1007. [Google Scholar] [CrossRef]
- Morales-Vera, R.; Vásquez-Ibarra, L.; Scott, F.; Puettmann, M.; Gustafson, R. Life Cycle Assessment of Bioethanol Production: A Case Study from Poplar Biomass Growth in the U.S. Pacific Northwest. Fermentation 2022, 8, 734. [Google Scholar] [CrossRef]
- Ingrao, C.; Matarazzo, A.; Gorjian, S.; Adamczyk, J.; Failla, S.; Primerano, P.; Huisingh, D. Wheat-straw derived bioethanol production: A review of Life Cycle Assessments. Sci. Total. Environ. 2021, 781, 146751. [Google Scholar] [CrossRef]
- Angili, T.S.; Grzesik, K.; Rödl, A.; Kaltschmitt, M. Life Cycle Assessment of Bioethanol Production: A Review of Feedstock, Technology and Methodology. Energies 2021, 14, 2939. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations—FAO. Land Evaluation towards a Revised Framework; Food and Agriculture Organization of the United Nations—FAO: Rome, Italy, 2007. [Google Scholar]
- Hauschild, M.Z.; Rosenbaum, R.K.; Olsen, S.I. (Eds.) Life Cycle Assessment—Theory and Practice; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Li, W.-C.; Zhang, S.-J.; Xu, T.; Sun, M.-Q.; Zhu, J.-Q.; Zhong, C.; Li, B.-Z.; Yuan, Y.-J. Fractionation of corn stover by two-step pretreatment for production of ethanol, furfural, and lignin. Energy 2020, 195, 117076. [Google Scholar] [CrossRef]
- Hossain, N.; Zaini, J.; Mahlia, T.M.I. Life cycle assessment, energy balance and sensitivity analysis of bioethanol production from microalgae in a tropical country. Renew. Sustain. Energy Rev. 2019, 115, 109371. [Google Scholar] [CrossRef]
- Devi, A.; Bajar, S.; Kour, H.; Kothari, R.; Pant, D.; Singh, A. Lignocellulosic Biomass Valorization for Bioethanol Production: A Circular Bioeconomy Approach. BioEnergy Res. 2022, 15, 1820–1841. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.-S.; Li, X.-L.; Xu, H.-J.; Fu, Y. Conversion of 5-hydroxymethylfurfural to chemicals: A review of catalytic routes and product applications. Fuel Process. Technol. 2020, 209, 106528. [Google Scholar] [CrossRef]
- Yao, H.; Wang, Y.; Liu, J.; Xu, M.; Ma, P.; Ji, J.; You, Z. Review on Applications of Lignin in Pavement Engineering: A Recent Survey. Front. Mater. 2022, 8, 803524. [Google Scholar] [CrossRef]
- López-Ortega, M.G.; Guadalajara, Y.; Junqueira, T.L.; Sampaio, I.L.; Bonomi, A.; Sánchez, A. Sustainability analysis of bioethanol production in Mexico by a retrofitted sugarcane industry based on the Brazilian expertise. Energy 2021, 232, 121056. [Google Scholar] [CrossRef]
- Bonomi, A.; Cavalett, O. Virtual Biorefinery; da Cunha, P., Lima, M.A.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Souza, A.; Watanabe, M.D.B.; Cavalett, O.; Ugaya, C.M.L.; Bonomi, A. Social life cycle assessment of first and second-generation ethanol production technologies in Brazil. Int. J. Life Cycle Assess. 2018, 23, 617–628. [Google Scholar] [CrossRef]
- García, C.A.; Manzini, F. Environmental and economic feasibility of sugarcane ethanol for the Mexican transport sector. Sol. Energy 2012, 86, 1063–1069. [Google Scholar] [CrossRef]
- Wang, C.; Malik, A.; Wang, Y.; Chang, Y.; Pang, M.; Zhou, D. Understanding the resource-use and environmental impacts of bioethanol production in China based on a MRIO-based hybrid LCA model. Energy 2020, 203, 117877. [Google Scholar] [CrossRef]
- Niphadkar, S.; Bagade, P.; Ahmed, S. Bioethanol production: Insight into past, present and future perspectives. Biofuels 2018, 9, 229–238. [Google Scholar] [CrossRef]
- Gupta, S.; Chen, H.; Hazen, B.T.; Kaur, S.; Gonzalez, E.D.S. Circular economy and big data analytics: A stakeholder perspective. Technol. Forecast. Soc. Chang. 2019, 144, 466–474. [Google Scholar] [CrossRef]
- Tsegaye, B.; Balomajumder, C.; Roy, P. Microbial delignification and hydrolysis of lignocellulosic biomass to enhance biofuel production: An overview and future prospect. Bull. Natl. Res. Cent. 2019, 43, 51. [Google Scholar] [CrossRef]
- Castillo, R.M.; Feng, K.; Sun, L.; Guilhoto, J.; Pfister, S.; Miralles-Wilhelm, F.; Hubacek, K. The land-water nexus of biofuel production in Brazil: Analysis of synergies and trade-offs using a multiregional input-output model. J. Clean. Prod. 2019, 214, 52–61. [Google Scholar] [CrossRef]
| Lignocellulosic Biomass | Cellulose, % | Hemicellulose, % | Lignin, % | Reference |
|---|---|---|---|---|
| Bamboo | 47 | 23 | 28 | [40] |
| Banana waste | 28.92 | 25.23 | 10.56 | [41] |
| Barley straw | 38.43 | 28.55 | 16.26 | [42] |
| Corn cobs | 33.6 | 37.2 | 19.3 | [43] |
| Corn stover | 35–40 | 17–35 | 7–18 | [44] |
| Corn stalk | 34.5 | 27.6 | 8.7 | [45] |
| Grass | 25–40 | 35–50 | 10–30 | [46] |
| Hardwood stems | 40–55 | 24–40 | 18–25 | [47] |
| Miscanthus | 38.38 | 24.23 | 17.66 | [48] |
| Municipal solid waste | 33–49 | 9–16 | 10–14 | [21] |
| Oat straw | 31–37 | 27–38 | 14–19 | [49] |
| Pinewood | 40 | 28.5 | 27.7 | [50] |
| Rice straw | 28–36 | 23–28 | 12–15 | [44] |
| Rye straw | 30.9 | 21.5 | 22.1 | [51] |
| Softwood stems | 45–50 | 25–35 | 25–35 | [41] |
| Sugarcane bagasse | 42–48 | 19–25 | 20–42 | [41] |
| Sunflower stalk | 34.6 | 21 | 30 | [52] |
| Sweet sorghum bagasse | 34–45 | 18–27 | 14–21 | [44] |
| Wheat straw | 33–38 | 26–32 | 17–19 | [43] |
| Pretreatment | Feedstocks | Pretreatment Conditions | Pretreatment Results | Limitations | Bioethanol Yield | Reference |
|---|---|---|---|---|---|---|
| Mechanical | Rice straw | Dry ball milling Loading charge biomass 50 g 60 min 1700 rpm | Sugar yields: Glucose (331 mg/g rice straw) Xylose (74 mg/g rice straw) Arabinose (14 mg/g rice straw) | High consumption of energy. No solubilization of lignin. | * | [70] |
| Oil palm biomass | Ball milling Loading charge biomass 20 g 50 min 250 rpm | Size reduction (88.2%) Crystallinity reduction (46.6%) Sugar yield: Glucose 36.1% Xylose 56.4% | Inhibitors produced as acetic acid (1.3 mg/ g of biomass). High energy consumption. | * | [71] | |
| Physical | Wood biomass | Microwave treatment/DES Choline chloride/oxalic acid 80 °C 800 W 3 min | Lignin removed 80% Glucose yield 22.3%. | Inhibitors such as HMF are produced. | * | [72] |
| Sweet lime peel | 2% v/v H2SO4 Loading charge of 6% 60 min 750 W 20 kHz. | 181.5 mg sugars/g biomass | Relative low loading biomass charge High biomass content increases viscosity resulting in low chemical reaction. | 64% in acidic pH | [73] | |
| Switchgrass | Steam explosion 200 °C 10 min | Sugar yield: Glucose 50.9% Xylose 28% Lignin removed 50% | Inhibitors such as formic acid, acetic acid, HMF, and furfural. | * | [74] | |
| Chemical | Sugarcane bagasse | Alkaline 15% NaOH 140 °C 1 h | Reducing sugars 5.29 g/L Lignin removed 90% | Glucose released is negatively affected by the amount of hemicellulose and lignin present. | 0.1 g ethanol/g biomass and 0.88 g/Lh | |
| Sugarcane bagasse | Organosolv Glycerol–acid 15% 130 °C 60 min | Glucan 65.8% | High concentration of glycerol–acid is required to improve hydrolysis. No significant influence on the lignin content. | 0.38 g ethanol/g biomass and 0.57 g/Lh | [75] | |
| Rice straw | Acid 1%H2SO4 10%(w/v) rice straw 100 °C 2 h | Reducing sugars 14 g/L Lignin removed 11.7% Xylose 83% | More recycling times for acid treatment hydrolysate decrease xylose yield. | Concentration of 40.6 g/L yield of 86.4% | [76] | |
| Rice straw | DES: choline chloride-ethylene glycol 150 °C 24 h Biomass loading 5% w/w | Lignin removed 74% Glucan digestibility 87% | Residual DES decreases the enzymatic hydrolysis of cellulose. | * | [77] | |
| Wheat straw | Alkaline-Microwave 2.75% NaOH Solid loading 10% (w/v) 23 min 100 °C | Lignin removed 60% Total carbohydrates 82% | High energy consumption. Rapid oxidation of carbohydrates in alkaline conditions. | 48 g ethanol/g sugar consumed | [78] | |
| Wheat straw | Ultrasound-assisted ionic liquid Triethylamine hydrogen sulfate ([TBA][H2SO4]) Sonicated at 24 kHz 130 °C 30 min | Saccharification yield 76.1% Lignin removed 74.9% | Sodium azide is used and represents a metabolic inhibitor. Ultrasound is not effective on a bigger scale. | 42 g/L | [79] | |
| Corn stalks | Ionic liquid 150 °C 11.5 h 2.5% arginine 420 MPa | Lignin removed 92% Purity of cellulose reach 85% | Arginine can inhibit cellulose degradation. Good method for fabrication of cellulose fiber but not ethanol. | * | [80] | |
| Green coconut shells | Organosolv 80% (w/w) glycerol 1%(w/w) sulfuric acid 121 °C | Glucose 49 g/L Lignin removed 60% | Inhibitors such as furfural and HMF are formed. | 29.6 g/L | [81] | |
| Olive tree biomass | Combined acid–alkaline 2.4% H2SO4 130 °C 84 min Peroxide: 7% H2O2 80 °C 90 min NaOH until 11.5 pH | Solubilization of hemicellulose 71% Lignin removed 80% Cellulose highly accessible 72% | Overliming method is needed for removing degradation products from lignocellulosic hydrolysates. Presence of acetic acid and furfural as potential inhibitors. | 15 g ethanol/100 g biomass | [82] | |
| Oil palm trunk | Alkaline Peroxide 3% H2O2 70 °C 30 min | Lignin removed 58% Cellulose extraction 74%. | High phenolic compounds released. Black liquor released with high amounts of tannins and gallic acid. | * | [83] | |
| Corn stover | Ammonia Recycle Percolation Process 170 °C 10% ammonia (v/w) 1 h | Lignin removed 70% Xylan removed 47% | High temperature and energy required to improve pretreatment performance. High operation cost. | 19.4 g/L | [83] | |
| Miscanthus (Miscanthus giganteus) | Acid diluted; 1% H2SO4 (v/v), 1:10 solid ratio (w/v), 121 °C for 30 min. | Xylose 24 g/L | Presence of furfural and HMF. | 13.58 g/L; 0.148 g bioethanol/g dry biomass | [84] | |
| Biological | Paddy straw | White rot fungi Pleurotus florida 5% inoculum 25–29 °C Biomass loading 10% (w/v) 28 days | Saccharification efficiency of 75% | High time required. No convenience in an industrial stage. Fungal residues limit enzymatic hydrolysis. | * | [85] |
| Chlorella sp. KR-1 | Polygalacturonase, amyloglucosidase, cellulase, and β-glucanase (simultaneous) pH 5.5; and 45 °C; 0.3 N HCl. | 28.5 g of sugar released/L of hydrolysate | Inhibitors such as furfurals, HMF, and formic acid might be produced. | Ethanol yield of 0.4 g/g of fermentable sugar | [86] | |
| Scenedesmus abundans | H2SO4/amyloglucosidase, α-amylase (simultaneous) | 10.752 g of total sugars/L and 5.730 g of glucose/L of the hydrolysate | Sugar content released depends on cultivation and pretreatment performance. | Ethanol yield of 0.1 g/g of algal biomass. | [87] |
| Fermentation | Microorganism/Enzyme | Substrate | Substrate Concentration | Inoculum Volume | Parameters | Ethanol Yield | Total Ethanol | Reference |
|---|---|---|---|---|---|---|---|---|
| LSF | S.cerevisiae X19G2 | Corn Stalk | 5% (w/v) | 1% (w/v) | 30 °C, 0 rpm, 96 h | - | 16.48 g/L | [128] |
| LSF | S.cerevisiae NRRLY-2034 | Defatted Nannochloropsis oculate biomass | 100% (v/v) | 2% (v/v) | 30 °C, 0 rpm, 96 h | 64.5% | 5.70–6.64 g/L | [129] |
| SScF | S.cerevisiae BY4743; cellulase (Celluclast 1.5 L) | Sugarcane bagasse | 10% (w/v) | 100 (U/g) cellulases 10% (v/v) S.cerevisiae | 40 °C, 120 rpm, 4.8 pH | 0.49 g/g | 4.88 g/L | [130] |
| SSCF | S.cerevisiae X19G2; glucoamylase | Potato peel wastes | 12.25% (w/v) | 10% (v/v) | 41.5 °C, 120 rpm, 5.5 pH, 24 h | 0.26–0.33% | 23.09–29.86 g/L | [131] |
| SSCF | S.cerevisiae (YSCII), T.harzianum, cellulase, β-glucosidase | Empty fruit bunches from palm | 45 (w/v) | 6.79%(v/v) | 30 °C, pH 4.8, 150 rpm, 72 h | 0.46 (g/g) | 9.65 g/L | [125] |
| SSCF and SSF | S.cerevisiae MTCC173, soil inoculum | Apple waste | - | 1% (w/v) S.cerevisiae; 1% (w/v) inoculum | 30 °C, 6 pH, 72 h, 70.89% (w/w) moisture | 49.64 g/L | - | [132] |
| SSF and SSCF | S.cerevisiae; A.niger | Pineapple waste | 15 g/mL | 1.64 × 105 (spores/mL) A.niger; 1.9 × 108 (cells/mL) S.cerevisiae | 30 °C, 4.1 pH, 7 days | 11.20% | - | [133] |
| SSF and SSCF | F.oxyspurum; C.cerevisiae; glucoamylase | Food waste | 0.1% (w/w) F.oxyspurum; 15 mg/g S.cerevisiae | 30 °C, 80 rpm, 94 h | - | 30.8 g/L | [134] | |
| VHG | S.cerevisiae C2/00 | Sugarcane molasses | 250.34 g/L | 30% (v/v) | 32 °C, 4.5 pH | 63.91% | 87.37 g/L | [135] |
| VHG; SSCF | L. sacchari LP175; Kluyveromyces marxianus DMKU-KS07; glucoamylases | Sugarcane bagasse | 250 g/L | 15% (v/v) L.sacchari and K.marxianus | 50 °C, 200 rpm, 6 h | 98.6 g/L | 118 g/L | [130] |
| Parameter | Collected Data | Reference |
|---|---|---|
| Plan lifetime | 25 years | [190] |
| Income tax rate | 30% | [190] |
| Sugarcane price | 34.5USD/TC (metric ton) | [190] |
| Ethanol price | 0.52 USD/L | [191] |
| Electricity price | 67.8 USD/MWh | [191] |
| Investment | 139 USD × 106 | [192] |
| Return of investment | 16% per year | [189] |
| Net present value | 66 USD × 106 | [190] |
| Co-products sold (sugar in a concept of biorefinery) | 0.49 USD/ kg | [190] |
| Greenhouse gases emitted | 1.67 × 105 gCO2 eq/ TC (metric ton of sugarcane) | [190] |
| Parameter | 1G Mexico | 2G China | 3G Brunei | Reference |
|---|---|---|---|---|
| Feedstock type | Sugarcane | Wheat straw | Algal biomass (Chlorella vulgaris) | [183,184,189] |
| Bioethanol produced | 77 L/metric ton of sugarcane | - | 57,087.5805 gallons per year/210 tons biomass per year | [184,189] |
| CO2 emitted | 1.67 × 102 kgCO2 eq/metric ton of sugarcane (GHG not only CO2) | 1.21 tons/USD 1420 of ethanol produced | 2.59 kg/gallon of bioethanol | [183,184,189] |
| Energy input | 16 kWh/metric ton of sugarcane | - | 3822 GJ per year | [183,184] |
| Water usage | 55.6 L/metric ton of sugarcane | 1281 m3/USD 1420 of ethanol produced | 2 m3/GJ−1 | [183,184,189,190] |
| Land usage | - | 0.13 ha/USD 1420 of ethanol produced | 2 m2/GJ−1 | [183,184] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaverino-Gutiérrez, M.A.; Wong, A.Y.C.-H.; Ibarra-Muñoz, L.A.; Chávez, A.C.F.; Sosa-Martínez, J.D.; Tagle-Pedroza, A.S.; Hernández-Beltran, J.U.; Sánchez-Muñoz, S.; Santos, J.C.d.; da Silva, S.S.; et al. Perspectives and Progress in Bioethanol Processing and Social Economic Impacts. Sustainability 2024, 16, 608. https://doi.org/10.3390/su16020608
Yaverino-Gutiérrez MA, Wong AYC-H, Ibarra-Muñoz LA, Chávez ACF, Sosa-Martínez JD, Tagle-Pedroza AS, Hernández-Beltran JU, Sánchez-Muñoz S, Santos JCd, da Silva SS, et al. Perspectives and Progress in Bioethanol Processing and Social Economic Impacts. Sustainability. 2024; 16(2):608. https://doi.org/10.3390/su16020608
Chicago/Turabian StyleYaverino-Gutiérrez, Mario Alberto, Alán Yazid Chávez-Hita Wong, Lizbeth Alejandra Ibarra-Muñoz, Ana Cristina Figueroa Chávez, Jazel Doménica Sosa-Martínez, Ana Sofia Tagle-Pedroza, Javier Ulises Hernández-Beltran, Salvador Sánchez-Muñoz, Julio César dos Santos, Silvio Silvério da Silva, and et al. 2024. "Perspectives and Progress in Bioethanol Processing and Social Economic Impacts" Sustainability 16, no. 2: 608. https://doi.org/10.3390/su16020608
APA StyleYaverino-Gutiérrez, M. A., Wong, A. Y. C.-H., Ibarra-Muñoz, L. A., Chávez, A. C. F., Sosa-Martínez, J. D., Tagle-Pedroza, A. S., Hernández-Beltran, J. U., Sánchez-Muñoz, S., Santos, J. C. d., da Silva, S. S., & Balagurusamy, N. (2024). Perspectives and Progress in Bioethanol Processing and Social Economic Impacts. Sustainability, 16(2), 608. https://doi.org/10.3390/su16020608