
Tracing Water Recharge and Transport in the Root-Zone Soil of Different Vegetation Types in the Poyang Lake Floodplain Wetland (China) Using Stable Isotopes
Abstract
:1. Introduction
2. Materials and Methods
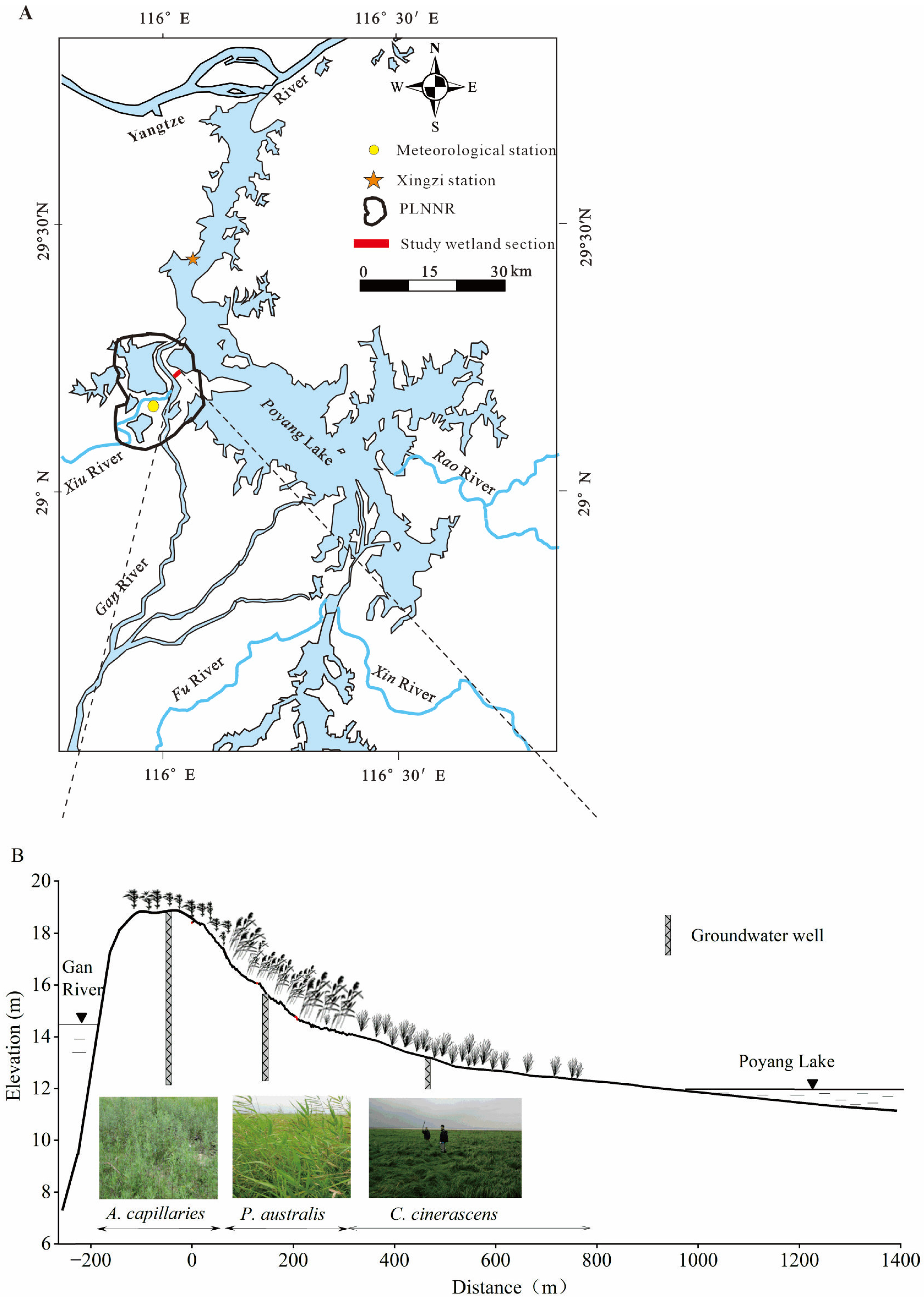
2.1. Study Area
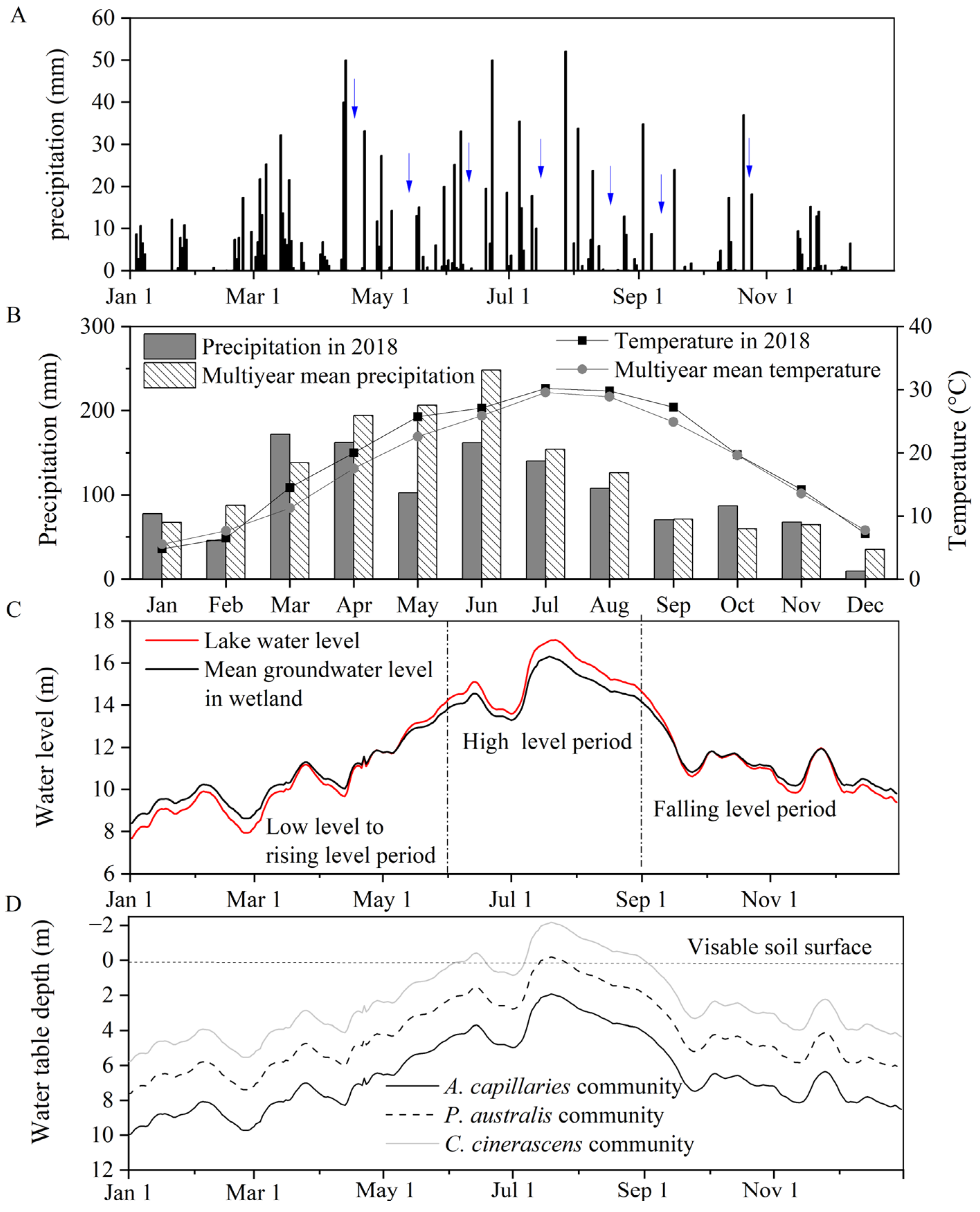
2.2. Source of Hydrological Data
2.3. Field Sampling and Laboratory Testing
2.4. Line-Conditioned Excess Calculation
2.5. Calculation of Water Supply Ratio
2.6. Statistical Analysis
3. Results
3.1. Characteristics of Hydrometeorological Parameters
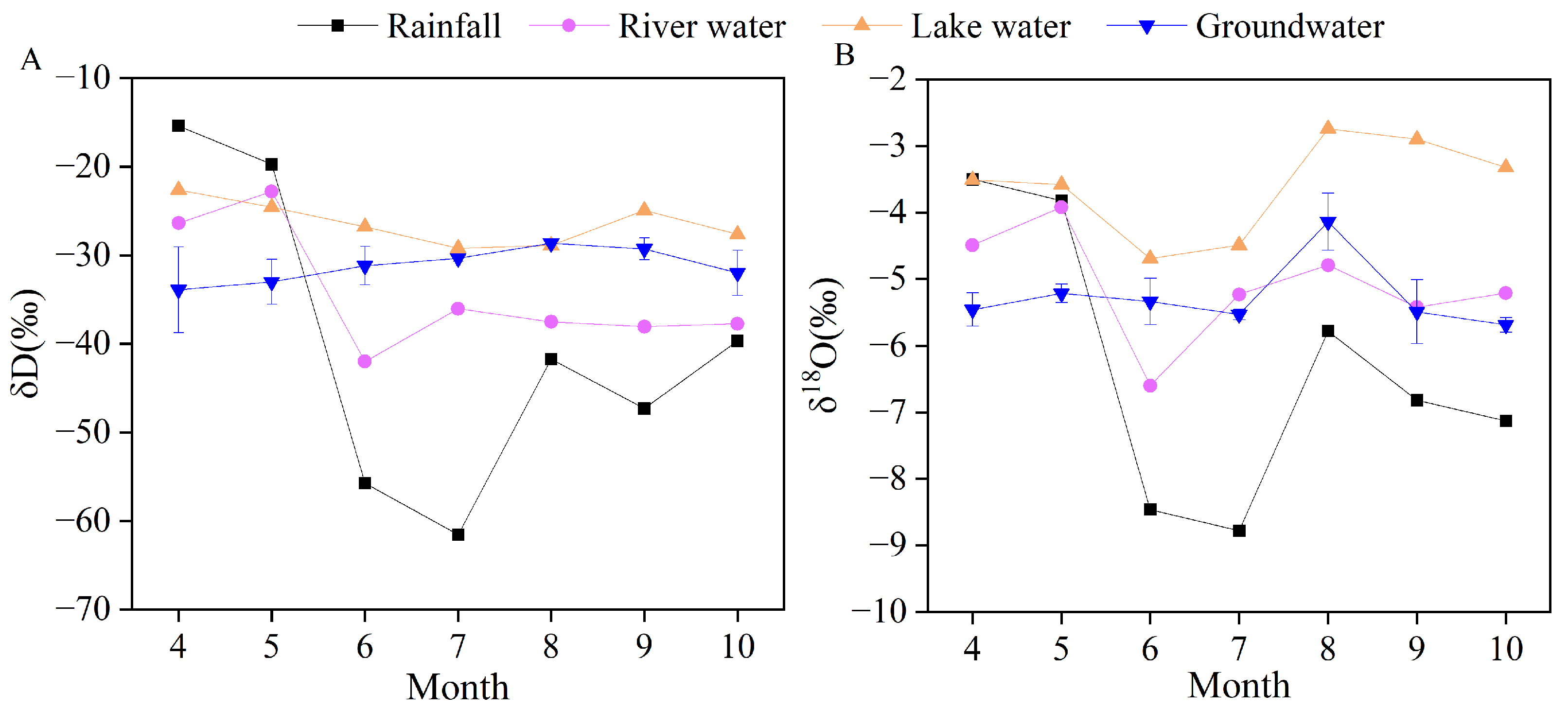
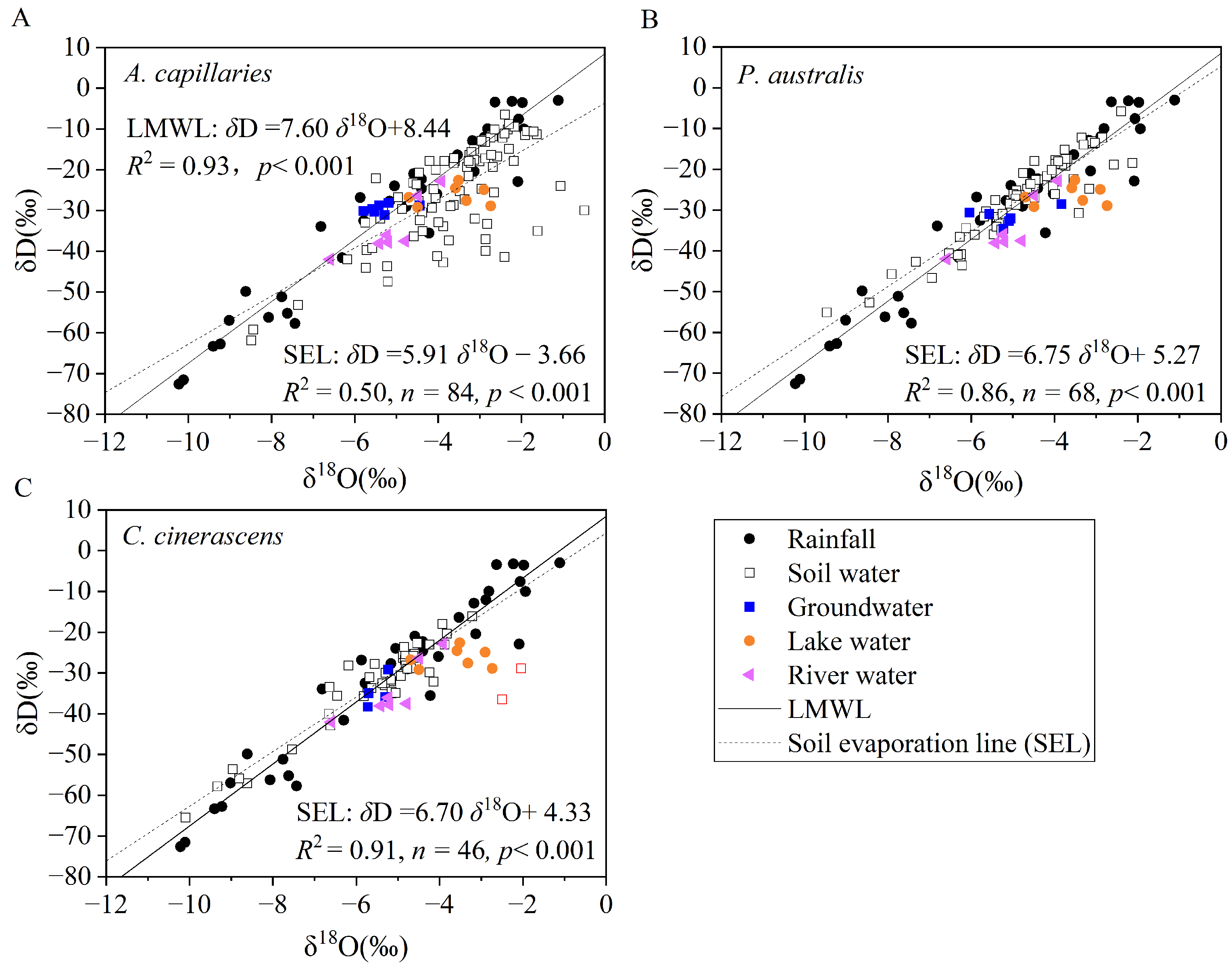
3.2. Isotopic Compositions of Different Water Sources
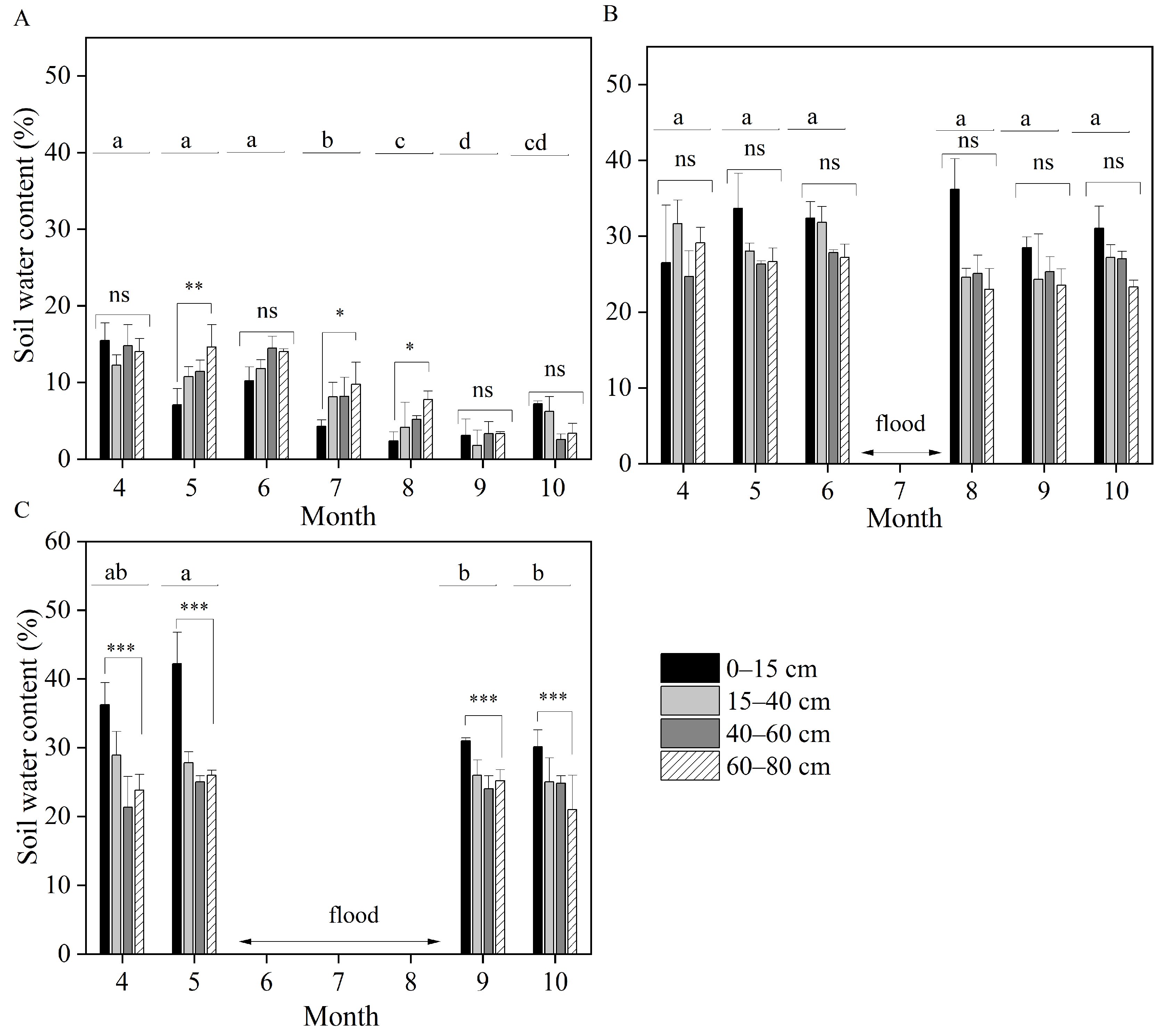
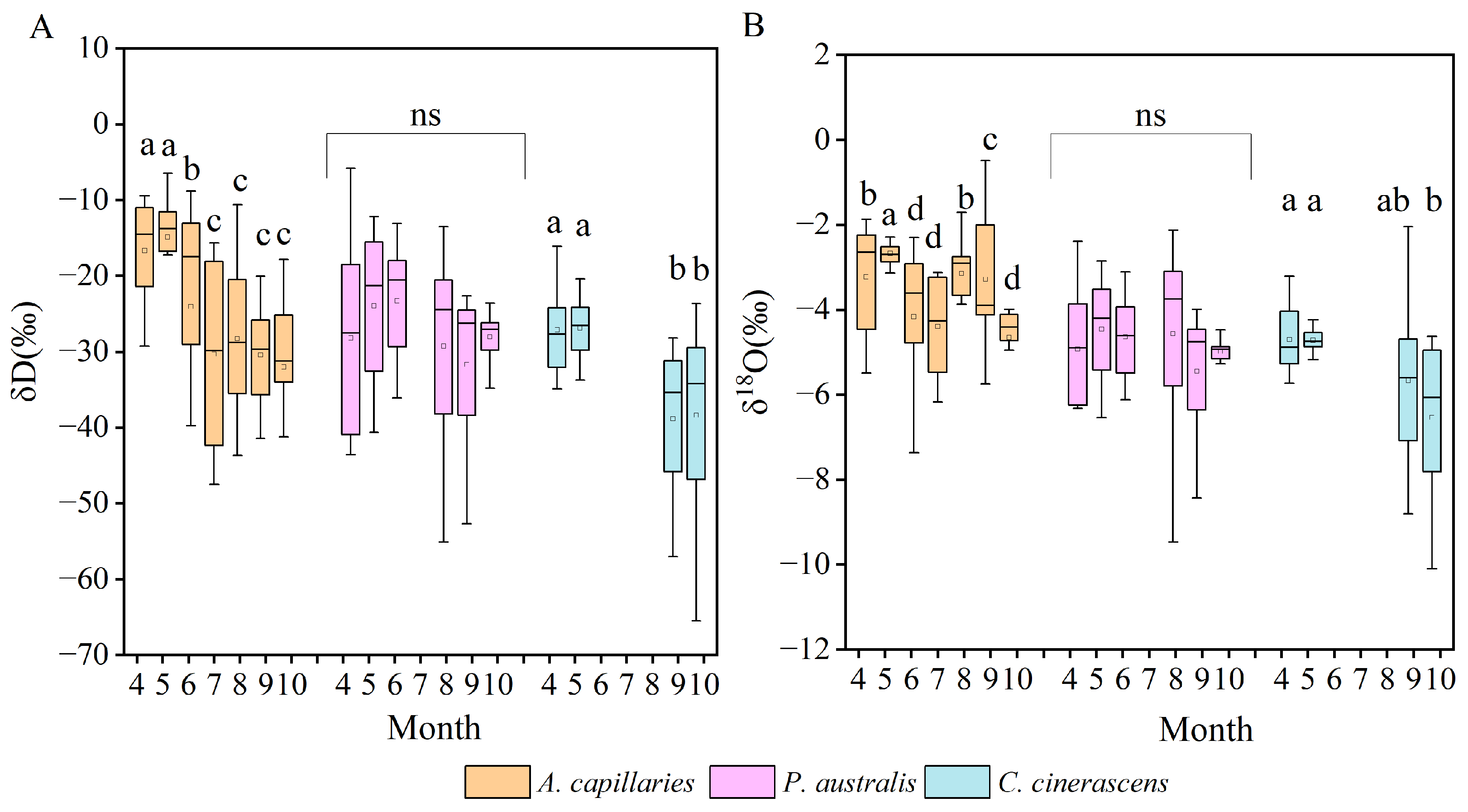
3.3. Isotopic Compositions of Root-Zone Soil Water
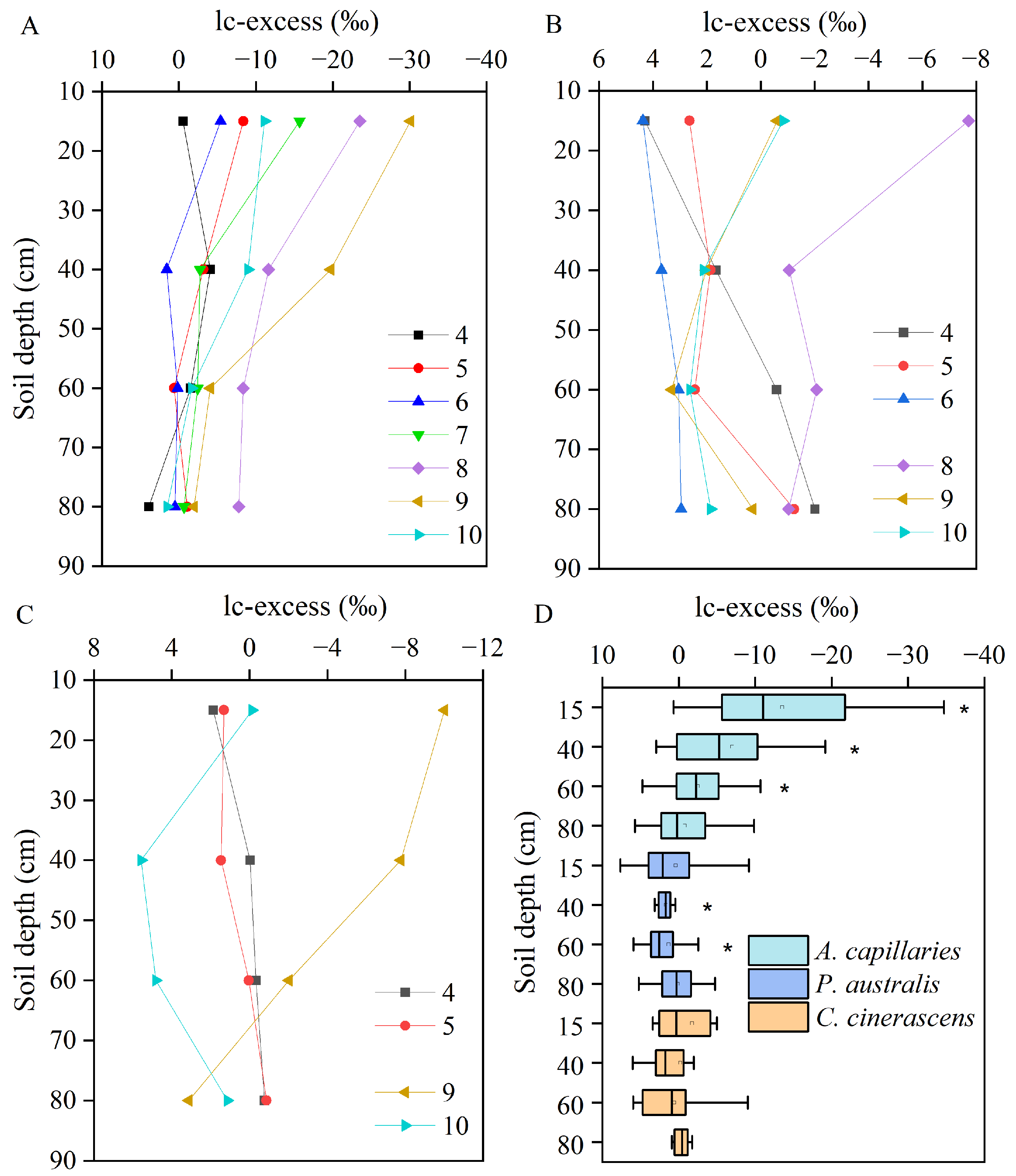
3.4. Line-Conditioned Excess of Soil Water
3.5. Contribution of Rainfall and Groundwater to Soil Water
4. Discussion
4.1. Mechanisms of Rainfall Infiltration, Mixing, and Piston Flow
4.2. Evidence for Preferential Flow and Its Formation Mechanisms
4.3. Evaporation Fractionation Depth of Soil Water
4.4. Groundwater Recharge and Capillary Flow
4.5. Hydrological Connection at the Profile Scale
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Henszey, R.J.; Pfeiffer, K.; Keough, J.R. Linking surface- and ground-water levels to riparian grassland species along the Platte River in Central Nebraska, USA. Wetlands 2004, 24, 665–687. [Google Scholar] [CrossRef]
- Hammersmark, C.T.; Rains, M.C.; Wickland, A.C.; Mount, J.F. Vegetation and water-table relationships in a hydrologically restored riparian meadow. Wetlands 2009, 29, 785–797. [Google Scholar] [CrossRef]
- Helliwell, J.R.; Sturrock, C.J.; Mairhofer, S.; Craigon, J.; Ashton, R.W.; Miller, A.J.; Whalley, W.R.; Mooney, S.J. The emergent rhizosphere: Imaging the development of the porous architecture at the root-soil interface. Sci. Rep. 2017, 7, 14875. [Google Scholar] [CrossRef]
- Jiang, B.F.; Cui, B.L.; Wang, Y.; Wang, Y.X.; Li, D.S.; Wang, L.S.; Li, X.Y. Stable-isotope tracing of vadose-zone water transport in Achnatherum splendens grassland of the Qinghai Lake Basin, NE Qinghai–Tibet Plateau, China. Catena 2021, 200, 105088. [Google Scholar] [CrossRef]
- Sprenger, M.; Leistert, H.; Gimbel, K.; Weiler, M. Illuminating hydrological processes at the soil-vegetation-atmosphere interface with water stable isotopes. Rev. Geophys. 2016, 54, 674–704. [Google Scholar] [CrossRef]
- Sprenger, M.; Tetzlaff, D.; Soulsby, C. Soil water stable isotopes reveal evaporation dynamics at the soil–plant–atmosphere interface of the critical zone. Hydrol. Earth Syst. Sci. 2017, 21, 3839–3858. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, Y.H.; Zhou, S.J.; Pan, Y.Y.; Wang, R.Q.; Li, Z.; Zhang, Z.M.; Zhang, M.X. Cracks and root channels promote both static and dynamic vertical hydrological connectivity in the Yellow River Delta. J. Clean. Prod. 2022, 367, 132972. [Google Scholar] [CrossRef]
- Zhang, M.D.; Wang, H.R.; Wang, C.Y.; Wang, K.Y.; Zhou, Y.K.; Bai, X.L. Water use strategy of Carex cinerascens and its response to water condition changes in Poyang Lake Wetland, China. Ecol. Indic. 2023, 146, 109863. [Google Scholar] [CrossRef]
- Gazis, C.; Feng, X.H. A stable isotope study of soil water: Evidence for mixing and preferential flow paths. Geoderma 2004, 119, 97–111. [Google Scholar] [CrossRef]
- Demand, D.; Blume, T.; Weiler, M. Spatio-temporal relevance and controls of preferential flow at the landscape scale. Hydrol. Earth Syst. Sci. 2019, 23, 4869–4889. [Google Scholar] [CrossRef]
- Song, X.F.; Wang, P.; Yu, J.J.; Liu, X.; Liu, J.R.; Yuan, R.Q. Relationships between precipitation, soil water and groundwater at Chongling catchment with the typical vegetation cover in the Taihang mountainous region, China. Environ. Earth. Sci. 2011, 62, 787–796. [Google Scholar] [CrossRef]
- Wan, H.; Liu, W.G. An isotope study (18O and D) of water movements on the Loess Plateau of China in arid and semiarid climates. Ecol. Eng. 2016, 93, 226–233. [Google Scholar] [CrossRef]
- Luo, Z.; Niu, J.; Zhang, L.; Chen, X.; Zhang, W.; Xie, B.Y.; Du, J.; Zhu, Z.J.; Wu, S.S.; Li, X. Roots-enhanced preferential flows in deciduous and coniferous forest soils revealed by dual-tracer experiments. J. Environ. Qual. 2018, 48, 136–146. [Google Scholar] [CrossRef]
- Bogner, C.; Gaul, D.; Kolb, A.; Schmiedinger, I.; Huwe, B. Investigating flow mechanisms in a forest soil by mixed-effects modelling. Eur. J. Soil. Sci. 2020, 61, 1079–1090. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.H.; Xie, L.M.; Zhao, S.Q.; Dai, L.Y.; Zhang, Z.M. Effect of soil characteristics on preferential flow of Phragmites australis community in Yellow River delta. Ecol. Indic. 2021, 125, 107486. [Google Scholar] [CrossRef]
- Dai, L.Y.; Zhang, Y.H.; Liu, Y.; Xie, L.M. Assessing hydrological connectivity of wetlands by dye-tracing experiment. Ecol. Indic. 2020, 119, 106840. [Google Scholar] [CrossRef]
- Boswell, J.S.; Olyphant, G.A. Modeling the hydrologic response of groundwater dominated wetlands to transient boundary conditions: Implications for wetland restoration. J. Hydrol. 2007, 332, 467–476. [Google Scholar] [CrossRef]
- Elci, A.; Molz, F.J. Identification of Lateral Macropore Flow in a Forested Riparian Wetland through Numerical Simulation of a Subsurface Tracer Experiment. Water Air Soil. Pollut. 2009, 197, 149–164. [Google Scholar] [CrossRef]
- Chen, X.; Chen, L.; Stone, M.C.; Acharya, K. Assessing connectivity between the river channel and floodplains during high flows using hydrodynamic modeling and particle tracking analysis. J. Hydrol. 2020, 583, 124609. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, Y.; Wan, H.; Deng, Z.M.; Pan, G.Y.; Xia, J. Using stable hydrogen and oxygen isotopes to study water movement in soil-plant-atmosphere continuum at Poyang Lake wetland, China. Wetl. Ecol. Manag. 2017, 25, 221–234. [Google Scholar] [CrossRef]
- Sun, H.M.; Hu, Z.; Zhang, J.; Wu, W.Z.; Liang, S.; Lu, S.Y.; Liu, H.Q. Determination of hydraulic flow patterns in constructed wetlands using hydrogen and oxygen isotopes. J. Mol. Liq. 2016, 223, 775–780. [Google Scholar] [CrossRef]
- Li, Y.L.; Yao, J.; Zhao, G.Z.; Zhang, Q. Evidences of hydraulic relationships between groundwater and lake water across the large floodplain wetland of Poyang Lake, China. Water Sci. Technol. Water Supply. 2018, 18, 698–712. [Google Scholar] [CrossRef]
- Tan, Z.Q.; Li, Y.L.; Xu, X.L.; Yao, J.; Zhang, Q. Mapping inundation dynamics in a heterogeneous floodplain: Insights from integrating observations and modeling approach. J. Hydrol. 2019, 572, 148–159. [Google Scholar] [CrossRef]
- Xu, X.L.; Zhang, Q.; Li, Y.L.; Li, X.H. Evaluating the influence of water table depth on transpiration of two vegetation communities in a lake floodplain wetland. Hydrol. Res. 2016, 47, 293–312. [Google Scholar] [CrossRef]
- Feng, W.J.; Mariotte, P.; Xu, L.G.; Buttler, A.; Bragazza, L.; Jiang, J.H.; Santonja, M. Seasonal variability of groundwater level effects on the growth of Carex cinerascens in lake wetlands. Ecol. Evol. 2020, 10, 517–526. [Google Scholar] [CrossRef]
- Feng, L.; Hu, C.; Chen, X.; Zhao, X. Dramatic inundation changes of China’s two largest freshwater lakes linked to the Three Gorges Dam. Environ. Sci. Technol. 2013, 47, 9628–9634. [Google Scholar] [CrossRef]
- Zhang, Q.; Ye, X.C.; Werner, A.D.; Li, Y.L.; Yao, J.; Li, X.H. An investigation of enhanced recessions in Poyang Lake: Comparison of Yangtze River and local catchment impacts. J. Hydrol. 2014, 517, 425–434. [Google Scholar] [CrossRef]
- Mei, X.F.; Dai, Z.J.; Fagherazzi, S.; Chen, J.Y. Dramatic variations in emergent wetland area in China’s largest freshwater lake, Poyang Lake. Adv. Water Resour. 2016, 96, 1–10. [Google Scholar] [CrossRef]
- Zhan, L.C.; Chen, J.S.; Zhang, S.Y.; Li, L.; Huang, D.W.; Wang, T. Isotopic signatures of precipitation, surface water, and groundwater interactions, Poyang Lake Basin, China. Environ. Earth. Sci. 2016, 75, 1307. [Google Scholar] [CrossRef]
- Wu, H.W.; Huang, Q.; Fu, C.S.; Song, F.; Liu, J.Z.; Li, J. Stable isotope signatures of river and lake water from Poyang Lake, China: Implications for river–lake interactions. J. Hydrol. 2021, 592, 125619. [Google Scholar] [CrossRef]
- Landwehr, J.M.; Coplen, T.B.; Stewart, D.W. Spatial, seasonal, and source variability in the stable oxygen and hydrogen isotopic composition of tap waters throughout the USA. Hydrol. Process. 2014, 28, 5382–5422. [Google Scholar] [CrossRef]
- Yeh, H.F.; Lin, H.I.; Lee, C.H.; Hsu, K.C.; Wu, C.S. Identifying seasonal groundwater recharge using environmental stable isotopes. Water 2014, 6, 2849–2861. [Google Scholar] [CrossRef]
- Du, K.; Zhang, B.Y.; Li, L.J. Soil Water Dynamics Under Different Land Uses in Loess Hilly Region in China by Stable Isotopic Tracing. Water 2021, 13, 242. [Google Scholar] [CrossRef]
- Zheng, W.B.; Wang, S.Q.; Sprenger, M.; Liu, B.X.; Cao, J.S. Response of soil water movement and groundwater recharge to extreme precipitation in a headwater catchment in the North China Plain. J. Hydrol. 2019, 576, 466–477. [Google Scholar] [CrossRef]
- Gehrels, J.C.; Peeters, J.E.M.; De Vries, J.J.; Dekkers, M. The mechanism of soil water movement as inferred from 18O stable isotope studies. Hydrolog Sci. J. 1998, 43, 579–594. [Google Scholar] [CrossRef]
- Orlowski, N.; Kraft, P.; Pferdmenges, J.; Breuer, L. Exploring water cycle dynamics through sampling multitude stable water isotope pools in a small developed landscape of Germany. Hydrol. Earth Syst. Sci. 2015, 12, 1809–1853. [Google Scholar]
- Zhang, Y.H.; Chen, J.H.; Zhang, J.C.; Zhang, Z.M.; Zhang, M.X. Novel indicator for assessing wetland degradation based on the index of hydrological connectivity and its correlation with the root-soil interface. Ecol. Indic. 2021, 133, 108392. [Google Scholar] [CrossRef]
- Thomas, E.M.; Lin, H.; Duffy, C.J.; Sullivan, P.L.; Holmes, G.H.; Brantley, S.L.; Jin, L.X. Spatiotemporal patterns of water stable isotope compositions at the shale hills critical zone observatory: Linkages to subsurface hydrologic processes. Vadose Zone J. 2013, 4, 1–16. [Google Scholar] [CrossRef]
- Wang, J.X.; Zhang, M.J.; Argiriou, A.A.; Wang, S.J.; Qu, D.Y.; Zhang, Y.; Su, P.Y. Recharge and Infiltration Mechanisms of Soil Water in the Floodplain Revealed by Water-Stable Isotopes in the Upper Yellow River. Sustainability 2021, 13, 9369. [Google Scholar] [CrossRef]
- Wiekenkamp, I.; Huisman, J.A.; Bogena, H.R.; Lin, H.S.; Vereecken, H. Spatial and temporal occurrence of preferential flow in a forested headwater catchment. J. Hydrol. 2016, 534, 139–149. [Google Scholar] [CrossRef]
- Kang, W.M.; Tian, J.; Bogena, H.R.; Lai, Y.; Xue, D.X.; He, C.S. Soil moisture observations and machine learning reveal preferential flow mechanisms in the Qilian Mountains. Geoderma 2023, 438, 116626. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, W.J.; Chen, C.F.; Liu, J.Q.; Yuan, Z.Q.; Jin, B.C.; Yu, X.Y. Effects of three morphometric features of roots on soil water flow behavior in three sites in China. Geoderma 2018, 320, 161–171. [Google Scholar] [CrossRef]
- Porporato, A.; D’odorico, P.; Laio, F.; Ridolfi, L.; Rodriguez-Iturbe, I. Ecohydrology of water-controlled ecosystems. Adv. Water Resour. 2002, 25, 1335–1348. [Google Scholar] [CrossRef]
- Hardie, M.A.; Cotching, W.E.; Doyle, R.B.; Holz, G.; Lisson, S.; Mattern, K. Effect of antecedent soil moisture on preferential flow in a texture-contrast soil. J. Hydrol. 2011, 398, 191–201. [Google Scholar] [CrossRef]
- Merdun, H.; Meral, R.; Riza Demirkiran, A. Effect of the initial soil moisture content on the spatial distribution of the water retention. Eurasian Soil. Sci. 2008, 41, 1098–1106. [Google Scholar] [CrossRef]
- Edwards, W.M.; Shipitalo, M.J.; Owens, L.B.; Dick, W.A. Factors affecting preferential flow of water and atrazine through earthworm burrows under continuous no-till corn. J. Environ. Qual. 1993, 22, 453–457. [Google Scholar] [CrossRef]
- Liu, H.; Lin, H. Frequency and Control of Subsurface Preferential Flow: From Pedon to Catchment Scales. Soil. Sci. Soc. Am. J. 2015, 79, 362–377. [Google Scholar] [CrossRef]
- Ghestem, M.; Sidle, R.C.; Stokes, A. The influence of plant root system on subsurface flow: Implications for slope stability. BioScience 2011, 61, 869–879. [Google Scholar] [CrossRef]
- Zimmermann, U.; Ehhalt, D.; Munnich, K.O. Soil water movement and evaporation: Changes in the isotopic composition of the water. In Proceedings of the IAEA Symposium on Isotope Hydrology, Vienna, Austria, 14–18 November 1966; IAEA: Vienna, Austria, 1967; pp. 567–585. [Google Scholar]
- Rothfuss, Y.; Merz, S.; Vanderborght, J.; Hermes, N.; Weuthen, A.; Pohlmeier, A.; Vereecken, H.; Brueggemann, N. Long-term and high-frequency non-destructive monitoring of water stable isotope profiles in an evaporating soil column. Hydrol. Earth Syst. Sci. 2015, 19, 4067–4080. [Google Scholar] [CrossRef]
- McCutcheon, R.J.; McNamara, J.P.; Kohn, M.J.; Evans, S.L. An Evaluation of the Ecohydrological Separation Hypothesis in a Semiarid Catchment. Hydrol. Process. 2016, 31, 783–799. [Google Scholar] [CrossRef]
- Simonin, K.A.; Link, P.; Rempe, D.; Miller, S.; Oshun, J.; Bode, C.; Dietrich, W.E.; Fung, I.; Dawson, T.E. Vegetation induced changes in the stable isotope composition of near surface humidity. Ecohydrology 2014, 7, 936–949. [Google Scholar] [CrossRef]
- Midwood, A.J.; Boutton, T.W.; Archer, S.R.; Watts, S.E. Water use by woody plants on contrasting soils in a savanna parkland: Assessment with δ2H and δ18O. Plant Soil. 1998, 205, 13–24. [Google Scholar] [CrossRef]
- Woessner, W.W. Stream and fluvial plain ground water interactions: Rescaling hydrogeologic thought. Groundwater 2000, 38, 423–429. [Google Scholar] [CrossRef]
- Yeh, H.F.; Lee, C.H.; Hsu, K.C. Oxygen and hydrogen isotopes for the characteristics of groundwater recharge: A case study from the Chih-Pen Creek basin, Taiwan. Environ. Earth Sci. 2011, 62, 393–402. [Google Scholar] [CrossRef]
- Pringle, C. What is hydrologic connectivity and why is it ecologically important? Hydrol. Process. 2003, 17, 2685–2689. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the Design of Ecological Field Experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Community | Ecotype | Wusong Base Level (m) | Companion Species |
|---|---|---|---|
| A. capillaries | Mesophyte | 17–18.5 | Cynodon dactylon an Eriophorum angustifolium |
| P. australis | Emergent plant | 15–16 | Triarrhena lutarioriparia, C. cinerascens, and Polygonum hydropiper |
| C. cinerascens | Hygrophyte | 12–14 | Artemisia selengensis and Phalaris arundinacea |
| Vegetation Community | Plant Coverage (%) | Belowground Biomass (g/0.25 m2) | Height (cm) | Depth (cm) | Sand (%) | Silt (%) | Clay (%) | Bulk Density (g/cm3) |
|---|---|---|---|---|---|---|---|---|
| A. capillaries | 50–60 | 83 ± 12 | 55 ± 8 | 0–15 | 92.8 | 6.2 | 1.0 | 1.33 |
| 15–40 | 91.2 | 7.8 | 1.0 | 1.26 | ||||
| 40–60 | 87.1 | 8.6 | 4.3 | 1.32 | ||||
| 60–80 | 82.3 | 10.2 | 7.5 | 1.37 | ||||
| P. australis | 100 | 586 ± 121 | 150 ± 10 | 0–15 | 13.6 | 75.4 | 11.0 | 1.30 |
| 15–40 | 12.4 | 65.3 | 22.3 | 1.24 | ||||
| 40–60 | 11.2 | 68.3 | 20.5 | 1.26 | ||||
| 60–80 | 10.2 | 62.1 | 27.7 | 1.27 | ||||
| C. cinerascens | 95–100 | 429 ± 104 | 63 ± 7 | 0–15 | 29.2 | 55.4 | 15.4 | 1.19 |
| 15–40 | 33.7 | 50.3 | 16.0 | 1.23 | ||||
| 40–60 | 40.2 | 44.1 | 15.7 | 1.25 | ||||
| 60–80 | 33.2 | 47.6 | 19.2 | 1.30 |
| Hydrological Period | Month | δ18O (‰) | Contribution Ratio (%) | |||
|---|---|---|---|---|---|---|
| Rainfall | Groundwater | Soil Water | Rainfall | Groundwater | ||
| Water level rising period | April | −3.49 | −5.23 | −4.93 | 17.3 | 82.7 |
| May | −3.82 | −5.05 | −4.45 | 48.6 | 51.4 | |
| High water level period | June | −8.46 | −5.09 | −4.64 | \ | \ |
| August | −5.78 | −3.83 | −4.57 | 37.7 | 62.3 | |
| Water level falling period | September | −6.82 | −6.04 | −5.45 | \ | \ |
| October | −7.13 | −5.57 | −4.99 | \ | \ | |
| Hydrological Period | Month | δ18O (‰) | Contribution Ratio (%) | |||
|---|---|---|---|---|---|---|
| Rainfall | Groundwater | Soil Water | Rainfall | Groundwater | ||
| Water level rising period | April | −3.49 | −5.72 | −4.71 | 45.6 | 54.4 |
| May | −3.82 | −5.30 | −4.72 | 39.3 | 60.7 | |
| Water level falling period | September | −6.82 | −5.24 | −5.67 | 27.2 | 72.8 |
| October | −7.13 | −5.70 | −6.54 | 58.4 | 41.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Zhao, J.; Wu, G.; Li, Y.; Hou, L. Tracing Water Recharge and Transport in the Root-Zone Soil of Different Vegetation Types in the Poyang Lake Floodplain Wetland (China) Using Stable Isotopes. Sustainability 2024, 16, 1755. https://doi.org/10.3390/su16051755
Xu X, Zhao J, Wu G, Li Y, Hou L. Tracing Water Recharge and Transport in the Root-Zone Soil of Different Vegetation Types in the Poyang Lake Floodplain Wetland (China) Using Stable Isotopes. Sustainability. 2024; 16(5):1755. https://doi.org/10.3390/su16051755
Chicago/Turabian StyleXu, Xiuli, Jun Zhao, Guangdong Wu, Yunliang Li, and Lili Hou. 2024. "Tracing Water Recharge and Transport in the Root-Zone Soil of Different Vegetation Types in the Poyang Lake Floodplain Wetland (China) Using Stable Isotopes" Sustainability 16, no. 5: 1755. https://doi.org/10.3390/su16051755