Effects of Understory Liana Trachelospermum jasminoides on Distributions of Litterfall and Soil Organic Carbon in an Oak Forest in Central China


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Liana Survey
2.4. Litter Survey
2.5. Soil Microclimate and Respiration
2.6. Soil Organic Carbon and Total Nitrogen Content
2.7. Data Analysis
3. Results
3.1. T. jasminoides Characteristics and Biomass Distribution
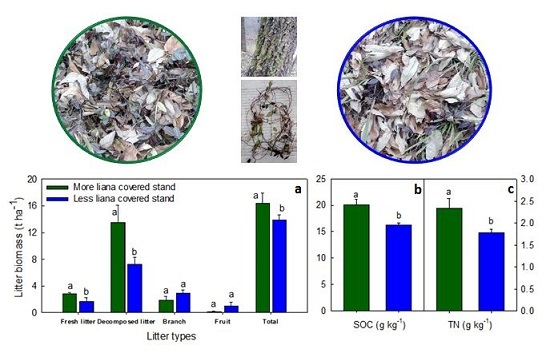
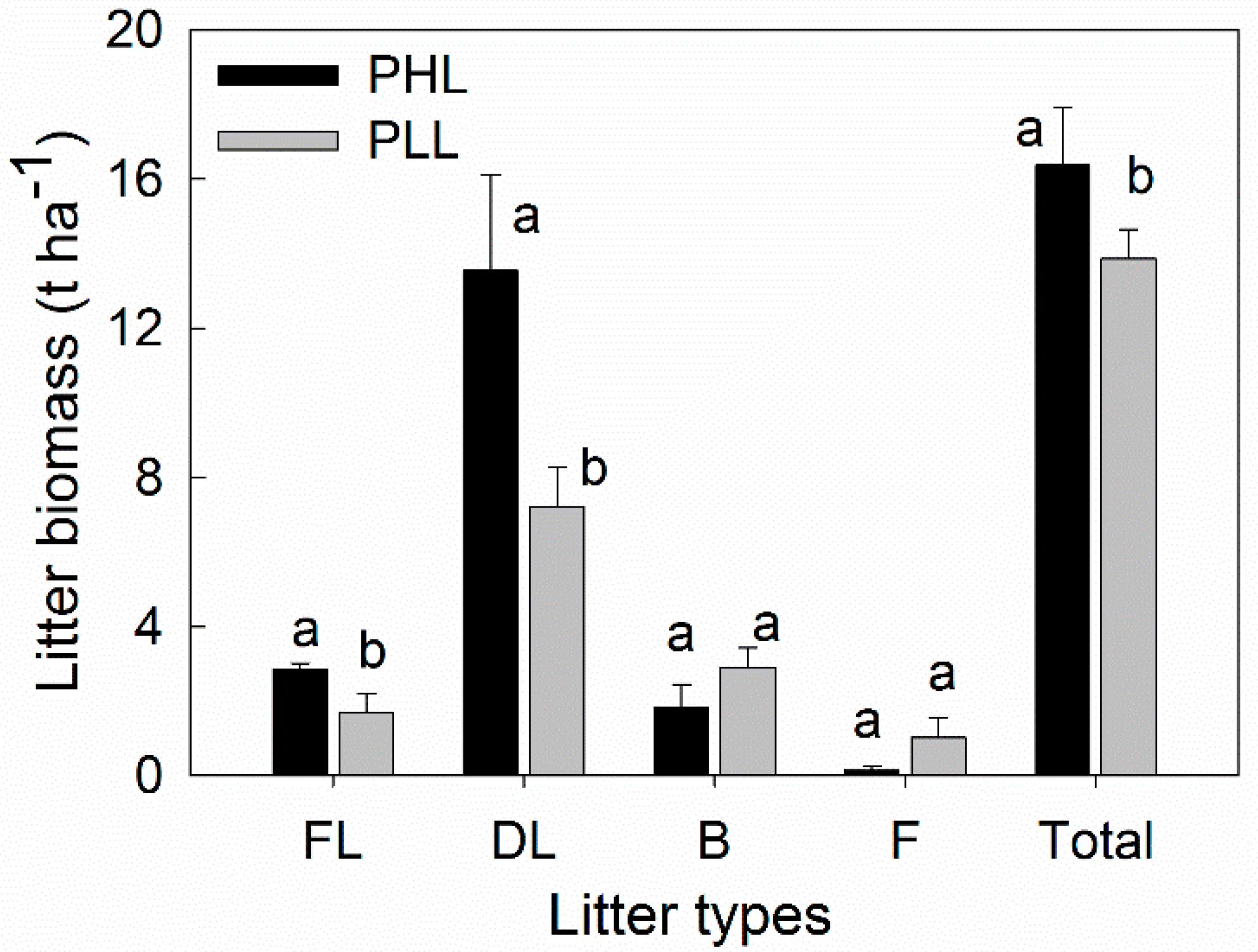
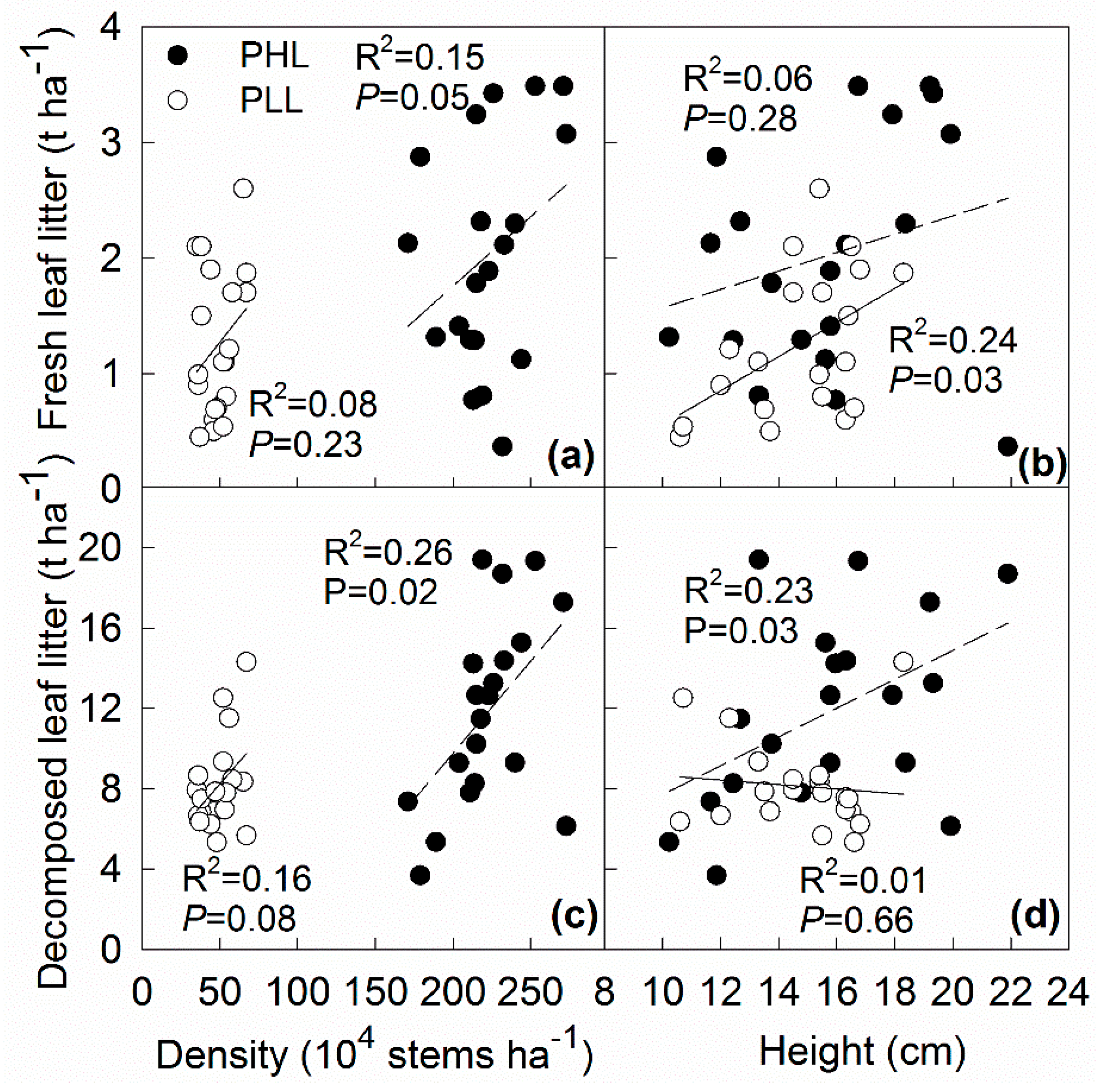
3.2. Effect of Liana on Litter Distribution
3.3. Effects of Liana on Soil Temperature and Moisture
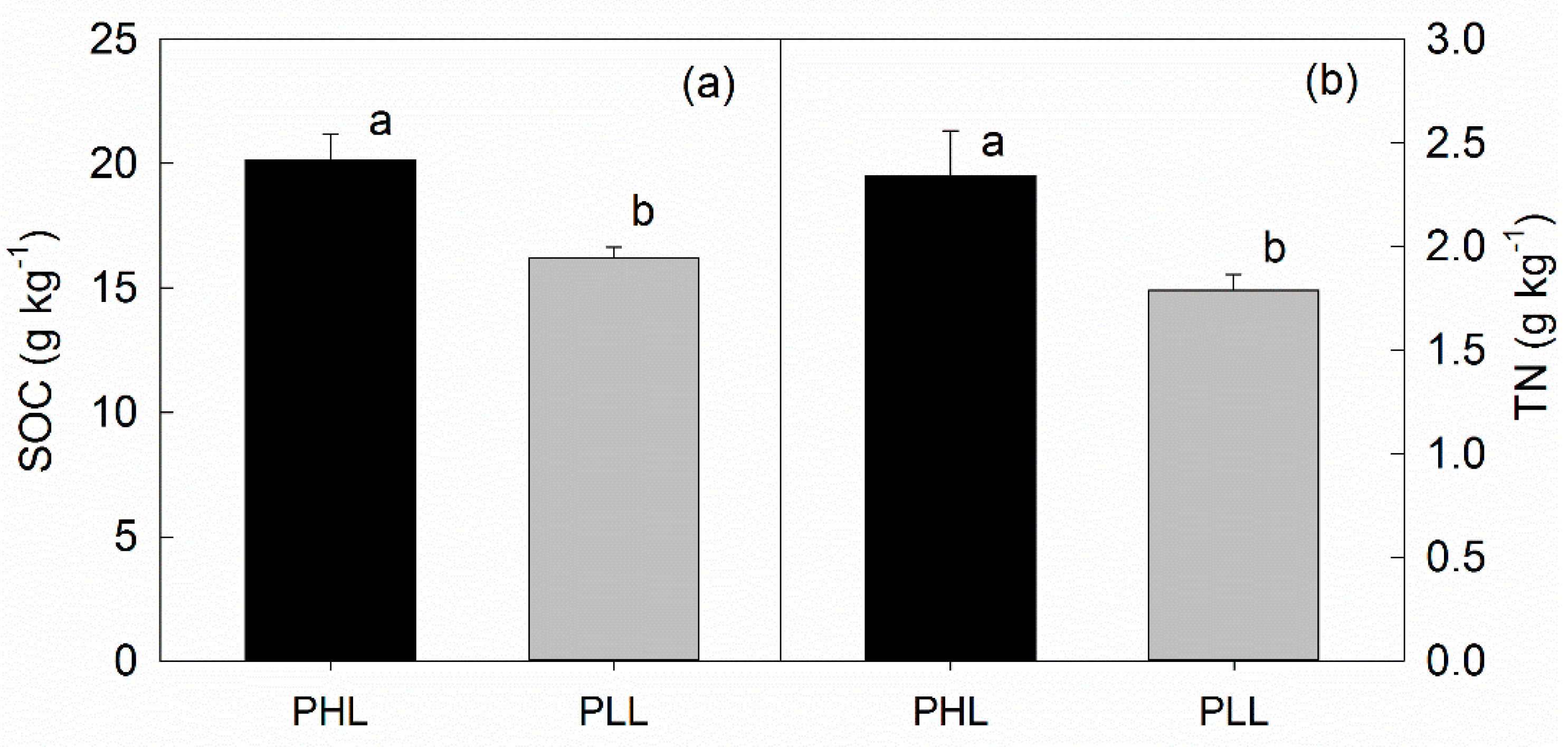
3.4. Effects of Liana on Soil Respiration, SOC, and TN
4. Discussion
4.1. Effect of Liana on Litter Distribution
4.2. Effects of Liana on SOC and TN
4.3. Implications for Forest C Storage and Ecological Restoration
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gerwing, J.J. Life history diversity among six species of canopy lianas in an old-growth forest of the eastern brazilian amazon. For. Ecol. Manag. 2004, 190, 57–72. [Google Scholar] [CrossRef]
- Addo-Fordjour, P.; El Duah, P.; Agbesi, D.K.K. Factors influencing liana species richness and structure following anthropogenic disturbance in a tropical forest, ghana. ISRN For. 2013, 2013, 11. [Google Scholar] [CrossRef]
- Vivek, P.; Parthasarathy, N. Liana community and functional trait analysis in tropical dry evergreen forest of india. J. Plant Ecol. 2015, 8, 501–512. [Google Scholar] [CrossRef]
- Campanello, P.I.; Gatti, M.G.; Ares, A.; Montti, L.; Goldstein, G. Tree regeneration and microclimate in a liana and bamboo-dominated semideciduous atlantic forest. For. Ecol. Manag. 2007, 252, 108–117. [Google Scholar] [CrossRef]
- Barry, K.E.; Schnitzer, S.A.; van Breugel, M.; Hall, J.S. Rapid liana colonization along a secondary forest chronosequence. Biotropica 2015, 47, 672–680. [Google Scholar] [CrossRef]
- Beaune, D. What would happen to the trees and lianas if apes disappeared? Oryx 2015, 49, 442–446. [Google Scholar] [CrossRef]
- Ibarra-Manríquez, G.; Martínez-Ramos, M. Landscape variation of liana communities in a neotropical rain forest. Plant Ecol. 2002, 160, 91–112. [Google Scholar] [CrossRef]
- Hättenschwiler, S. Liana seedling growth in response to fertilisation in a neotropical forest understorey. Basic Appl. Ecol. 2002, 3, 135–143. [Google Scholar] [CrossRef]
- Gehring, C.; Park, S.; Denich, M. Liana allometric biomass equations for amazonian primary and secondary forest. For. Ecol. Manag. 2004, 195, 69–83. [Google Scholar] [CrossRef]
- Pasquini, S.C.; Wright, S.J.; Santiago, L.S. Lianas always outperform tree seedlings regardless of soil nutrients: Results from a long-term fertilization experiment. Ecology 2015, 96, 1866–1876. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; van der Heijden, G.M.F.; Mascaro, J.; Carson, W.P. Lianas in gaps reduce carbon accumulation in a tropical forest. Ecology 2014, 95, 3008–3017. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Distribution and effects on tree growth of lianas and woody hemiepiphytes in a costa rican tropical wet forest. J. Trop. Ecol. 1990, 6, 321–331. [Google Scholar] [CrossRef]
- Wright, S.J.; Sun, I.F.; Pickering, M.; Fletcher, C.D.; Chen, Y.Y. Long-term changes in liana loads and tree dynamics in a malaysian forest. Ecology 2015, 96, 2748–2757. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, L.M.; Meiners, S.J. Impacts of temperate lianas on tree growth in young deciduous forests. For. Ecol. Manag. 2009, 259, 195–200. [Google Scholar] [CrossRef]
- Ichihashi, R.; Tateno, M. Biomass allocation and long-term growth patterns of temperate lianas in comparison with trees. New Phytol. 2015, 207, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; Bongers, F. The ecology of lianas and their role in forests. Trend Ecol. Evol. 2002, 17, 223–230. [Google Scholar] [CrossRef]
- Grauel, W.T.; Putz, F.E. Effects of lianas on growth and regeneration of Prioria copaifera in Darien, Panama. For. Ecol. Manag. 2004, 190, 99–108. [Google Scholar] [CrossRef]
- Laurance, W.F.; Andrade, A.S.; Magrach, A.; Camargo, J.L.C.; Valsko, J.J.; Campbell, M.; Fearnside, P.M.; Edwards, W.; Lovejoy, T.E.; Laurance, S.G. Long-term changes in liana abundance and forest dynamics in undisturbed amazonian forests. Ecology 2013, 95, 1604–1611. [Google Scholar] [CrossRef]
- Agnelli, A.; Bol, R.; Trumbore, S.; Dixon, L.; Cocco, S.; Corti, G. Carbon and nitrogen in soil and vine roots in harrowed and grass-covered vineyards. Agric. Ecosyst. Environ. 2014, 193, 70–82. [Google Scholar] [CrossRef]
- Londré, R.A.; Schnitzer, S.A. The distribution of lianas and their change in abundance in temperate forests over the past 45 years. Ecology 2006, 87, 2973–2978. [Google Scholar]
- Ladwig, L.M.; Meiners, S.J. Spatiotemporal dynamics of lianas during 50 years of succession to temperate forest. Ecology 2010, 91, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.H.; Li, H.M.; Yang, Y.P.; Chen, S.T.; Li, C.Z.; Shen, S.H. Effects of simulated nitrogen deposition on soil respiration in northern subtropical deciduous broad-leaved forest. Environ. Sci. 2010, 31, 1726–1732. [Google Scholar]
- Song, C.S. Scientific Survey of the Baotianman Mountial National Nature Reserve; China Forestry Publishing House: Beijing, China, 1994; pp. 32–35. [Google Scholar]
- Liu, Y.; Liu, S.; Wang, J.; Zhu, X.; Zhang, Y.; Liu, X. Variation in soil respiration under the tree canopy in a temperate mixed forest, central China, under different soil water conditions. Ecol. Res. 2014, 29, 133–142. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E.; Sparks, D.; Page, A.; Helmke, P.; Loeppert, R.; Soltanpour, P.; Tabatabai, M.; Johnston, C.; Sumner, M. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; Sparks, D., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bremner, J.; Sparks, D.; Page, A.; Helmke, P.; Loeppert, R.; Soltanpour, P.; Tabatabai, M.; Johnston, C.; Sumner, M. Nitrogen-Total. In Methods of Soil Analysis; Sparks, D., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Kreye, J.K.; Varner, J.M.; Dugaw, C.J. Spatial and temporal variability of forest floor duff characteristics in long-unburned pinus palustris forests. Can. J. For. Res. 2014, 44, 1477–1486. [Google Scholar] [CrossRef]
- Pérez-Salicrup, D.R.; Barker, M.G. Effect of liana cutting on water potential and growth of adult senna multijuga (caesalpinioideae) trees in a bolivian tropical forest. Oecologia 2000, 124, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Nepstad, D.C.; Tohver, I.M.; Ray, D.; Moutinho, P.; Cardinot, G. Mortality of large trees and lianas following experimental drought in an amazon forest. Ecology 2007, 88, 2259–2269. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.E.; Silver, W.L.; Fasth, B.; Chen, H.U.A.; Burke, I.C.; Parton, W.J.; Hart, S.C.; Currie, W.S. Long-term patterns of mass loss during the decomposition of leaf and fine root litter: An intersite comparison. Glob. Chang. Biol. 2009, 15, 1320–1338. [Google Scholar] [CrossRef]
- Nikolaidou, A.E.; Pavlatou-Ve, A.K.; Kostopoulou, S.K.; Mamolos, A.P.; Kalburtji, K.L. Litter quality and decomposition of vitis vinifera l. Residues under organic and conventional farming systems. Eur. J. Soil Biol. 2010, 46, 208–217. [Google Scholar] [CrossRef]
- Chen, M.M.; Zhu, Y.G.; Su, Y.H.; Chen, B.D.; Fu, B.J.; Marschner, P. Effects of soil moisture and plant interactions on the soil microbial community structure. Eur. J. Soil Biol. 2007, 43, 31–38. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Bastida, F.; Nicolás, C.; Moreno, J.L.; del Cerro, A.; Andrés, M. Influence of forest cover and herbaceous vegetation on the microbiological and biochemical properties of soil under mediterranean humid climate. Eur. J. Soil Biol. 2010, 46, 273–279. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Ickes, K.; Nilus, R.; Harms, K.E.; Burslem, D.F. Liana habitat associations and community structure in a bornean lowland tropical forest. Plant Ecol. 2006, 186, 203–216. [Google Scholar] [CrossRef]
- Pérez-Salicrup, D.R.; Sork, V.L.; Putz, F.E. Lianas and trees in a liana forest of amazonian bolivia1. Biotropica 2001, 33, 34–47. [Google Scholar] [CrossRef]
- Cox, P.M.; Pearson, D.; Booth, B.B.; Friedlingstein, P.; Huntingford, C.; Jones, C.D.; Luke, C.M. Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature 2013, 494, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Kuruppuarachchi, K.A.J.M.; Seneviratne, G.; Madurapperuma, B.D. Carbon sequestration in tropical forest stands: its control by plant, soil and climatic factors. Open J. For. 2016, 6, 59. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Height (cm) | Number of Leaf | Density (104 Individuals·ha−1) | Biomass (t·ha−1) | |||
|---|---|---|---|---|---|---|---|
| Leaf | Stem | Root | Total | ||||
| PHL | 21.89 ± 5.58 a | 4.3 ± 2.0 a | 232.4 ± 19.3 a | 0.19 ± 0.06 (8.8%) a | 0.33 ± 0.13 (15.3%) a | 1.64 ± 0.31 (75.9%) a | 2.16 ± 0.55 a |
| PLL | 26.37 ± 5.31 b | 5.9 ± 1.5 b | 40.2 ± 31.4 b | 0.05 ± 0.01 (10.5%) b | 0.07 ± 0.01 (14.6%) b | 0.36 ± 0.04 (75.0%) b | 0.48 ± 0.02 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Shang, Q.; Zhang, B.; Zhang, K.; Luan, J. Effects of Understory Liana Trachelospermum jasminoides on Distributions of Litterfall and Soil Organic Carbon in an Oak Forest in Central China. Sustainability 2017, 9, 1019. https://doi.org/10.3390/su9061019
Liu Y, Shang Q, Zhang B, Zhang K, Luan J. Effects of Understory Liana Trachelospermum jasminoides on Distributions of Litterfall and Soil Organic Carbon in an Oak Forest in Central China. Sustainability. 2017; 9(6):1019. https://doi.org/10.3390/su9061019
Chicago/Turabian StyleLiu, Yanchun, Qing Shang, Bo Zhang, Kesheng Zhang, and Junwei Luan. 2017. "Effects of Understory Liana Trachelospermum jasminoides on Distributions of Litterfall and Soil Organic Carbon in an Oak Forest in Central China" Sustainability 9, no. 6: 1019. https://doi.org/10.3390/su9061019
APA StyleLiu, Y., Shang, Q., Zhang, B., Zhang, K., & Luan, J. (2017). Effects of Understory Liana Trachelospermum jasminoides on Distributions of Litterfall and Soil Organic Carbon in an Oak Forest in Central China. Sustainability, 9(6), 1019. https://doi.org/10.3390/su9061019