Predicting Tropical Tree Species Richness from Normalized Difference Vegetation Index Time Series: The Devil Is Perhaps Not in the Detail


Abstract
:1. Introduction
2. Materials and Methods
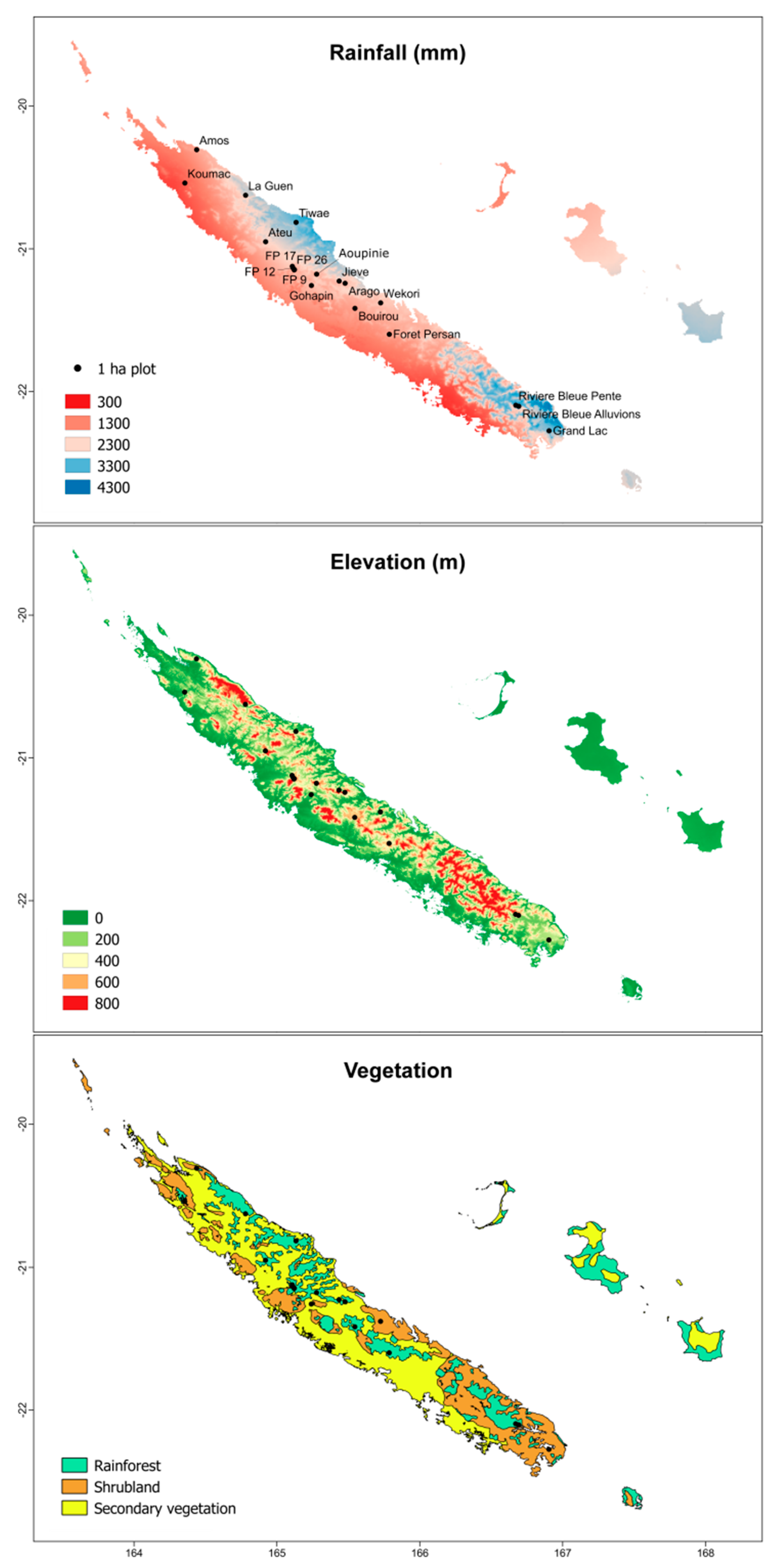
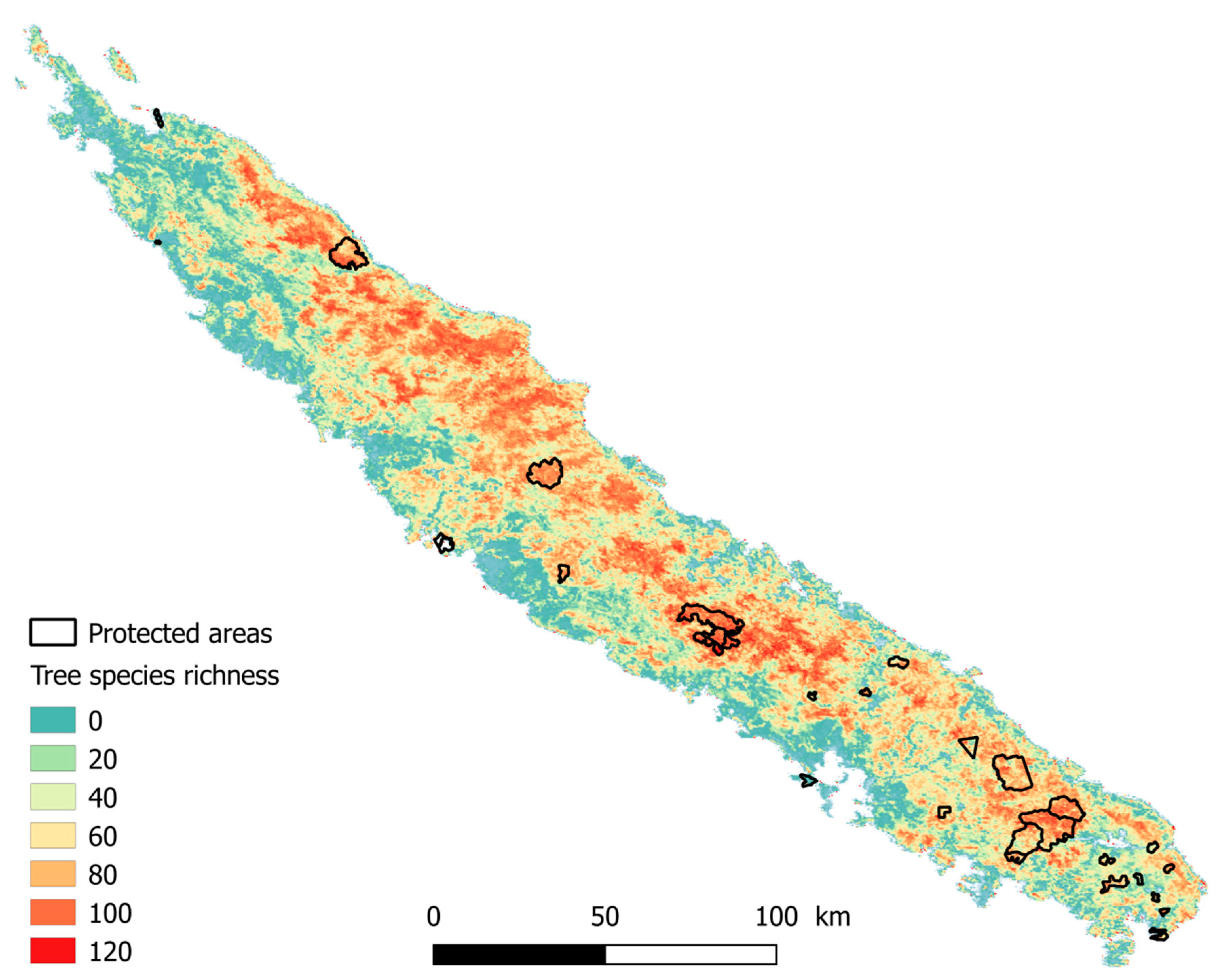
2.1. Study Area
2.2. Field Data
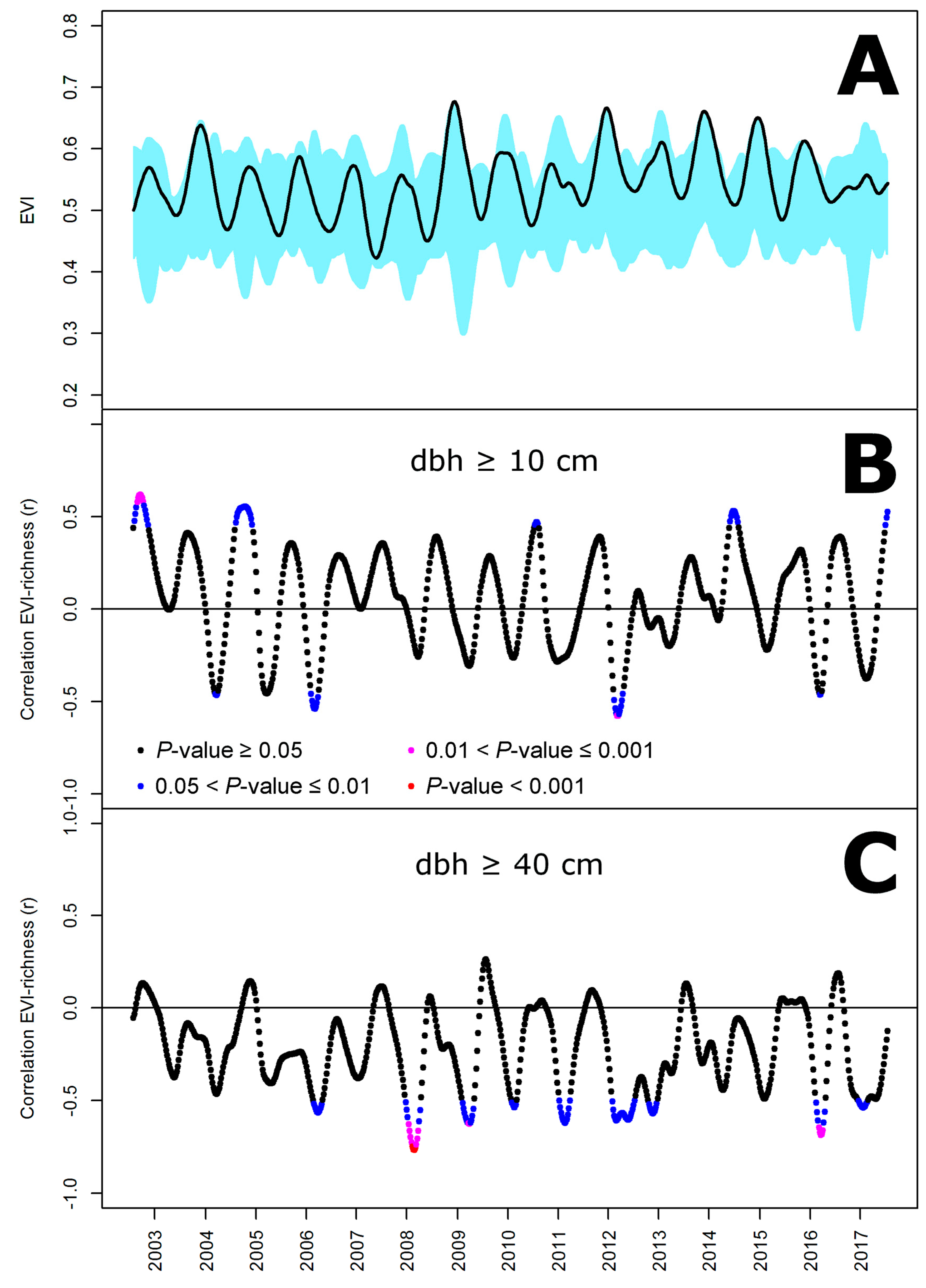
2.3. Remote Sensing and Statistical Analysis
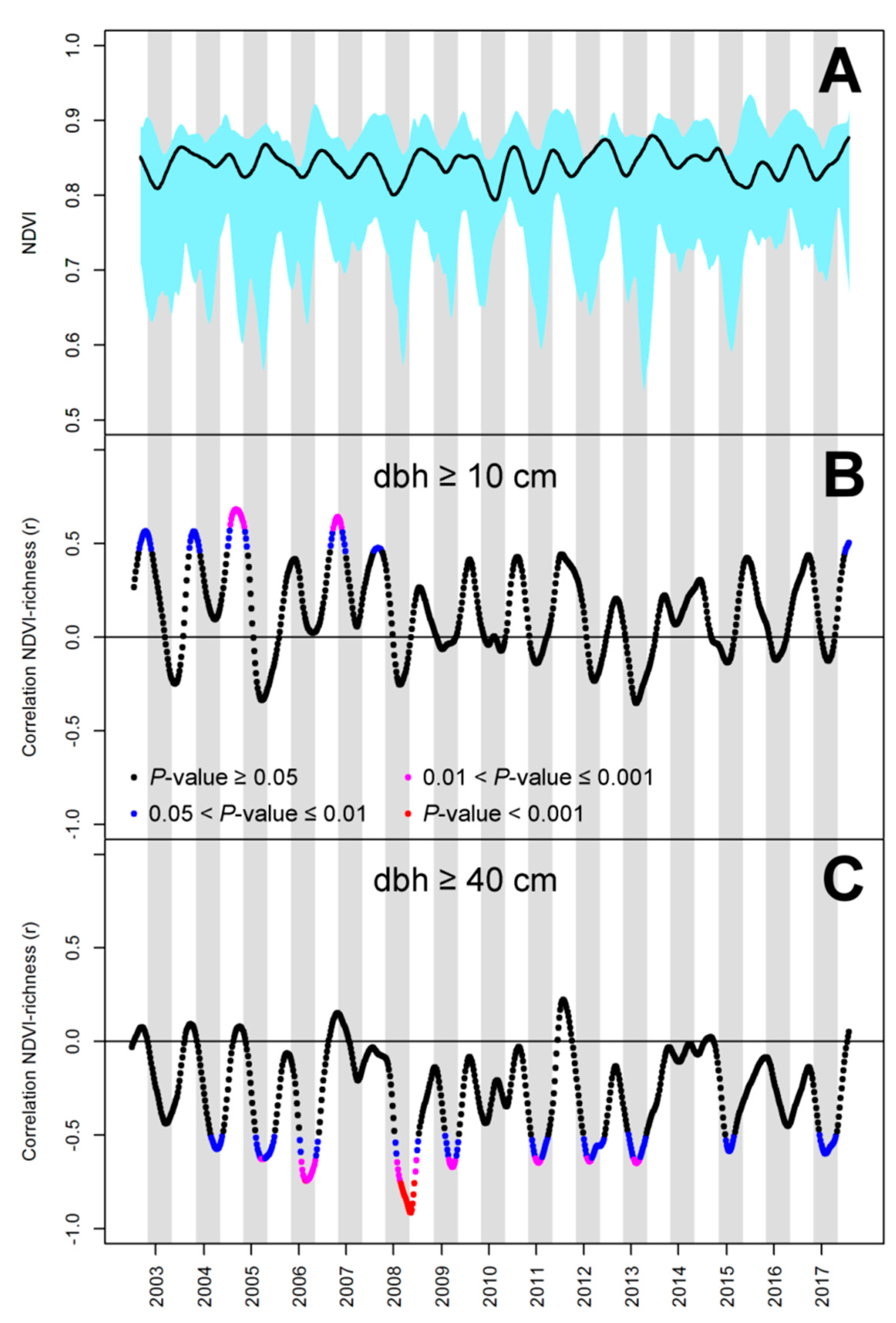
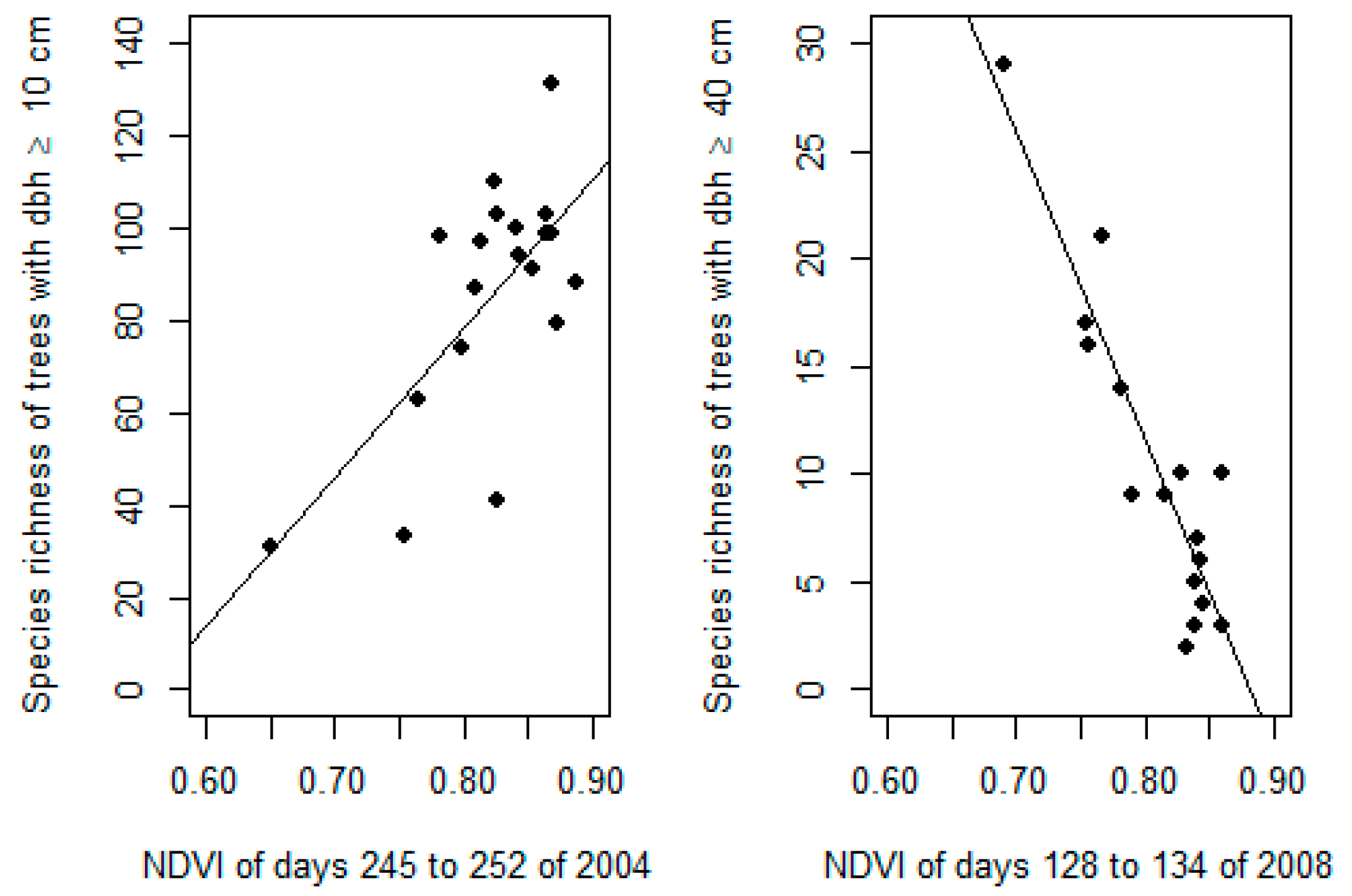
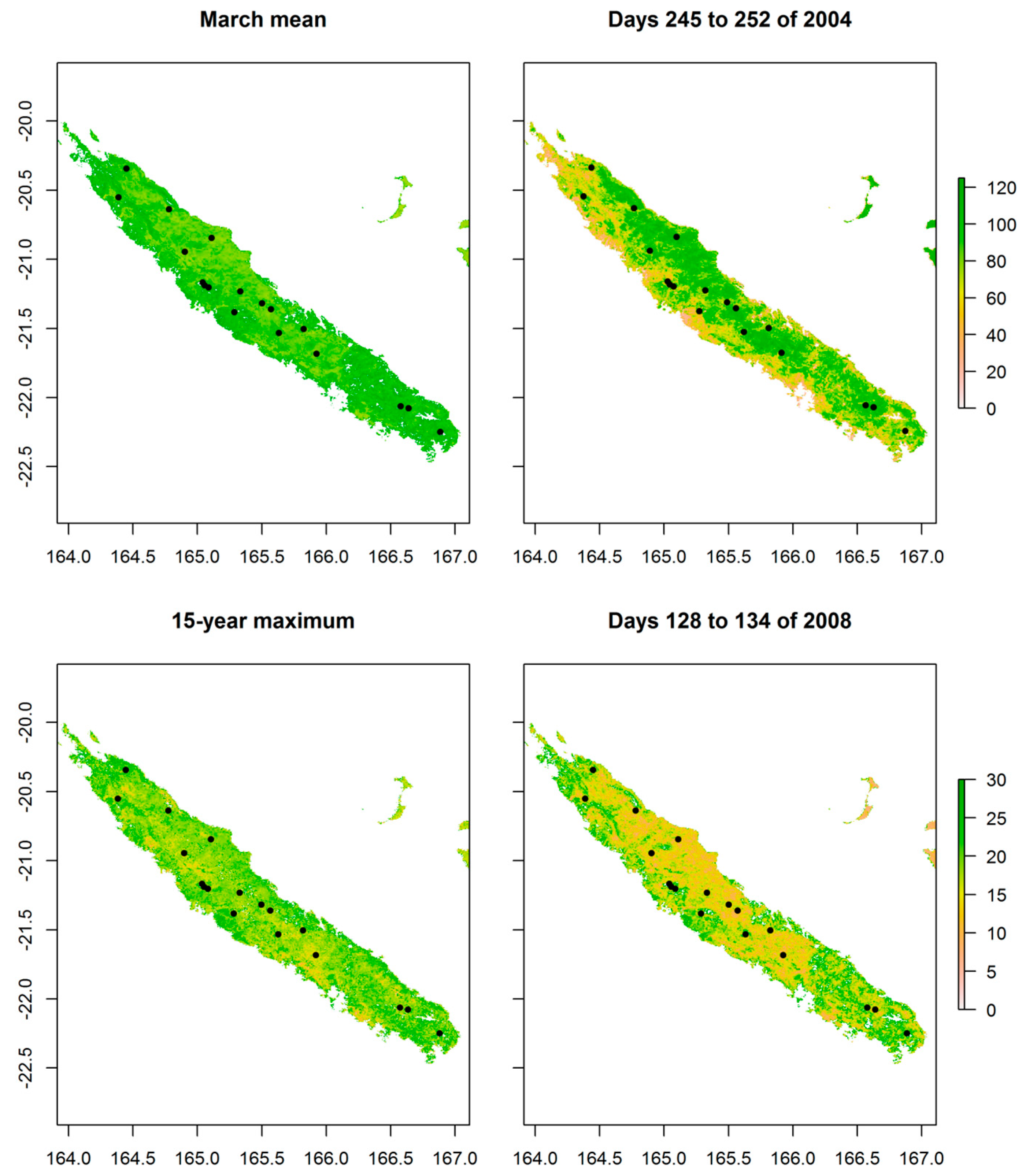
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistics | SR (dbh ≥ 10 cm) | SR (dbh ≥ 40 cm) |
|---|---|---|
| Full time-series statistics | ||
| 15-year mean | 0.07 | −0.37 |
| 15-year median | 0.10 | −0.35 |
| 15-year variance | −0.51 * | 0.11 |
| 15-year 1st quartile | 0.16 | −0.36 |
| 15-year 3rd quartile | −0.01 | −0.29 |
| 15-year minimum | 0.19 | −0.51 * |
| 15-year maximum | −0.21 | −0.26 |
| Seasonal statistics | ||
| Wet season mean | −0.15 | −0.50 * |
| Wet season median | −0.13 | −0.48 |
| Wet season variance | −0.25 | 0.37 |
| 15-year 1st quartile | −0.17 | −0.58 * |
| 15-year 3rd quartile | −0.17 | −0.40 |
| Wet season minimum | 0.17 | −0.53 * |
| Wet season maximum | −0.23 | −0.28 |
| Dry season mean | 0.29 | −0.15 |
| Dry season median | 0.29 | −0.15 |
| Dry season variance | −0.46 * | −0.00 |
| 15-year 1st quartile | 0.34 | −0.16 |
| 15-year 3rd quartile | 0.24 | −0.10 |
| Dry season minimum | 0.38 | −0.31 |
| Dry season maximum | 0.06 | −0.17 |
| Annual statistics | ||
| 2002 mean | 0.52 * | 0.09 |
| 2002 variance | 0.04 | 0.25 |
| 2003 mean | 0.25 | −0.21 |
| 2003 variance | −0.13 | 0.09 |
| 2004 mean | 0.18 | −0.25 |
| 2004 variance | −0.59 * | 0.02 |
| 2005 mean | −0.04 | −0.36 |
| 2005 variance | −0.19 | 0.05 |
| 2006 mean | −0.04 | −0.41 |
| 2006 variance | −0.35 | 0.07 |
| 2007 mean | 0.16 | −0.24 |
| 2007 variance | −0.01 | −0.19 |
| 2008 mean | 0.08 | −0.46 |
| 2008 variance | 0.08 | 0.19 |
| 2009 mean | −0.07 | −0.37 |
| 2009 variance | −0.04 | 0.55 * |
| 2010 mean | −0.05 | −0.31 |
| 2010 variance | −0.56 * | −0.23 |
| 2011 mean | 0.07 | −0.27 |
| 2011 variance | −0.56 * | −0.10 |
| 2012 mean | −0.24 | −0.66 ** |
| 2012 variance | 0.16 | 0.19 |
| 2013 mean | 0.03 | −0.23 |
| 2013 variance | −0.39 | −0.10 |
| 2014 mean | 0.24 | −0.27 |
| 2014 variance | −0.36 | −0.11 |
| 2015 mean | 0.09 | −0.17 |
| 2015 variance | −0.00 | 0.14 |
| 2016 mean | 0.01 | −0.41 |
| 2016 variance | −0.31 | 0.34 |
| 2017 mean | −0.13 | −0.55 * |
| 2017 variance | −0.47 * | 0.11 |
| Monthly statistics | ||
| January mean | −0.16 | −0.49 |
| January variance | −0.12 | 0.47 |
| February mean | −0.32 | −0.59 * |
| February variance | 0.02 | 0.40 |
| March mean | −0.37 | −0.61 * |
| March variance | 0.16 | 0.39 |
| April mean | −0.20 | −0.52 * |
| April variance | 0.10 | 0.20 |
| May mean | 0.03 | −0.33 |
| May variance | −0.00 | 0.03 |
| June mean | 0.23 | −0.13 |
| June variance | −0.23 | 0.14 |
| July mean | 0.35 | −0.05 |
| July variance | −0.44 | 0.44 |
| August mean | 0.37 | −0.06 |
| August variance | −0.39 | 0.19 |
| September mean | 0.35 | −0.09 |
| September variance | −0.51 * | −0.09 |
| October mean | 0.27 | −0.17 |
| October variance | −0.54 * | −0.06 |
| November mean | 0.16 | −0.24 |
| November variance | −0.35 | 0.28 |
| December mean | −0.00 | −0.36 |
| December variance | −0.22 | 0.48 |
References
- Bradley, B.A.; Fleishman, E. Can remote sensing of land cover improve species distribution modelling? J. Biogeogr. 2008, 35, 1158–1159. [Google Scholar] [CrossRef]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.-N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Waring, R.H.; Coops, N.C.; Ohmann, J.L.; Sarr, D.A. Interpreting woody plant richness from seasonal ratios of photosynthesis. Ecology 2002, 83, 2964–2970. [Google Scholar] [CrossRef]
- Fairbanks, D.H.K.; McGwire, K.C. Patterns of floristic richness in vegetation communities of California: Regional scale analysis with multi-temporal NDVI. Glob. Ecol. Biogeogr. 2004, 13, 221–235. [Google Scholar] [CrossRef]
- Gillespie, T.W. Predicting woody-plant species richness in tropical dry forests: A case study from South Florida. Ecol. Appl. 2005, 15, 27–37. [Google Scholar] [CrossRef]
- Feeley, K.J.; Gillespie, T.W.; Terborgh, J.W. The utility of spectral indices from Landsat ETM+ for measuring the structure and composition of tropical dry forests. Biotropica 2005, 37, 508–519. [Google Scholar] [CrossRef]
- Cayuela, L.; Benayas, J.M.; Justel, A.; Salas-Rey, J. Modelling tree diversity in a highly fragmented tropical montane landscape. Glob. Ecol. Biogeogr. 2006, 15, 602–613. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Saatchi, S.; Pau, S.; Bohlman, S.; Giorgi, A.P.; Lewis, S. Towards quantifying tree species richness in tropical forests. Int. J. Remote Sens. 2008, 30, 1629–1634. [Google Scholar] [CrossRef]
- Wolf, J.A.; Fricker, G.A.; Meyer, V.; Hubbell, S.P.; Gillespie, T.W.; Saatchi, S.S. Plant species richness is associated with canopy height and topography in a Neotropical forest. Remote Sens. 2012, 4, 4010–4021. [Google Scholar] [CrossRef]
- Fricker, G.A.; Wolfe, G.; Saatchi, S.S.; Gillespie, T.W. Predicting variation of tree species richness in tropical forest landscapes using high spatial resolution remote sensing. Ecol. Appl. 2015, 25, 1776–1789. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Gamon, J.A.; Montgomery, R.A.; Townsend, P.A.; Zygielbaum, A.I.; Bitan, K.; Tilman, D.; Cavender-Bares, J. Seasonal variation in the NDVI-species richness relationship in a prairie grassland experiment (Cedar Creek). Remote Sens. 2016, 8, 128. [Google Scholar] [CrossRef]
- Cumming, S.G.; Stralberg, D.; Lefevre, K.L.; Sólymos, P.; Bayne, E.M.; Fang, S.; Fontaine, T.; Mazerolle, D.; Schmiegelow, F.K.A.; Song, S.J. Climate and vegetation hierarchically structure patterns of songbird distribution in the Canadian boreal region. Ecography 2014, 37, 137–151. [Google Scholar] [CrossRef]
- Youngentob, K.N.; Yoon, H.J.; Stein, J.; Lindenmayer, B.D.; Held, A.A. Where the wild things are: Using remotely sensed forest productivity to assess arboreal marsupial species richness and abundance. Divers. Distrib. 2015, 21, 977–990. [Google Scholar] [CrossRef]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jedrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen successes in animal ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- Pau, S.; Gillespie, T.W.; Wolkovich, E.M. Dissecting NDVI-species richness relationships in Hawaiian dry forests. J. Biogeogr. 2012, 39, 1678–1686. [Google Scholar] [CrossRef]
- Šímová, I.; Storch, D. The enigma of terrestrial primary productivity: Measurements, models, scales and the diversity-productivity relationship. Ecography 2017, 40, 239–252. [Google Scholar] [CrossRef]
- Pouteau, R.; Bayle, E.; Blanchard, E.; Birnbaum, P.; Cassan, J.-J.; Hequet, V.; Ibanez, T.; Vandrot, H. Accounting for the indirect area effect in stacked species distribution models to map species richness in a montane biodiversity hotspot. Divers. Distrib. 2015, 21, 1929–1938. [Google Scholar] [CrossRef]
- Météo-France. Atlas Climatique de la Nouvelle-Calédonie; Météo-France: Noumea, New Caledonia, 2007. [Google Scholar]
- Jaffré, T. The relationship between ecological diversity and floristic diversity in New Caledonia. Biodivers. Lett. 1993, 1, 82–87. [Google Scholar] [CrossRef]
- Jaffré, T.; Rigault, F.; Munzinger, J. La végétation. In Atlas de la Nouvelle-Calédonie; Bonvallot, J., Gay, J.-C., Habert, E., Eds.; IRD-Congrès de la Nouvelle-Calédonie: Marseille, France, 2012; pp. 73–76. [Google Scholar]
- Morat, P.; Jaffré, T.; Tronchet, F.; Munzinger, J.; Pillon, Y.; Veillon, J.-M.; Chapolin, M. The taxonomic database ‘FLORICAL’ and characteristics of the indigenous flora of New Caledonia. Adansonia 2012, 3, 177–219. [Google Scholar]
- Jaffré, T.; Veillon, J.-M. Étude floristique et structurale de deux forêts denses humides sur roches ultrabasiques en Nouvelle-Calédonie. Adansonia 1991, 4, 243–273. [Google Scholar]
- Jaffré, T.; Veillon, J.-M. Structural and floristic characteristics of a rain forest on schist in New Caledonia: A comparison with an ultramafic rain forest. Adansonia 1995, 17, 201–226. [Google Scholar]
- Ibanez, T.; Blanchard, E.; Hequet, V.; Keppel, G.; Laidlaw, M.; Pouteau, R.; Vandrot, H.; Birnbaum, P. High endemism and stem density distinguish New Caledonian from other high-diversity rainforests in the Southwest Pacific. Ann. Bot. 2017, 121, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [PubMed]
- University of Natural Resources and Life Sciences, Vienna. Available online: http://ivfl-info.boku.ac.at/ (accessed on 4 January 2018).
- Vuolo, F.; Mattiuzzi, M.; Klisch, A.; Atzberger, C. Data service platform for MODIS Vegetation Indices time series processing at BOKU Vienna: Current status and future perspectives. Proc. SPIE 2012, 8538. [Google Scholar] [CrossRef]
- Wiens, J.J. The causes of species richness patterns across space, time, and clades and the role of “ecological limits”. Q. Rev. Biol. 2011, 86, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Marks, C.O.; Muller-Landau, H.C.; Tilman, D. Tree diversity, tree height and environmental harshness in eastern and western North America. Ecol. Lett. 2016, 19, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Marks, C.O.; Muller-Landau, H.C.; Tilman, D. Tree diversity in relation to maximum tree height: Evidence for the harshness hypothesis of species diversity gradients. Ecol. Lett. 2017, 20, 398–399. [Google Scholar] [CrossRef] [PubMed]
- Slik, J.W.; Paoli, G.; McGuire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large trees drive forest aboveground biomass variation in moist lowland forests across the tropics. Glob. Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Bastin, J.F.; Barbier, N.; Réjou-Méchain, M.; Fayolle, A.; Gourlet-Fleury, S.; Maniatis, D.; de Haulleville, T.; Baya, F.; Beeckman, H.; Beina, D.; et al. Seeing Central African forests through their largest trees. Sci. Rep. 2015, 5, 13156. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.; Birnbaum, P.; Ibanez, T.; Boutreux, T.; Antin, C.; Ploton, P.; Vincent, G.; Pouteau, R.; Vandrot, H.; Hequet, V.; et al. Contrasted allometries between stem diameter, crown area and tree height in five tropical biogeographic areas. Trees Struct. Fonct. 2016, 6, 1953–1968. [Google Scholar] [CrossRef]





| Plot | Elevation (m) | SR (dbh ≥ 10 cm) | SR (dbh ≥ 40 cm) | Data Source |
|---|---|---|---|---|
| Amos | 486 | 94 | 7 | [26] |
| Aoupinie | 884 | 87 | 29 | [26] |
| Arago | 488 | 110 | 16 | [26] |
| Ateou | 780 | 88 | 17 | [26] |
| Bouirou | 533 | 103 | 10 | [26] |
| Jieve | 370 | 98 | 14 | [26] |
| Foret Persan | 435 | 99 | NA | [24] |
| Foret Plate 9 | 508 | 91 | 9 | IAC/UMR AMAP |
| Foret Plate 12 | 513 | 100 | 10 | [26] |
| Foret Plate 17 | 454 | 63 | 3 | IAC/UMR AMAP |
| Foret Plate 26 | 485 | 65 | 5 | IAC/UMR AMAP |
| Gohapin | 272 | 41 | 4 | IAC/UMR AMAP |
| Grand Lac | 273 | 97 | 21 | IAC/UMR AMAP |
| Koumac | 45 | 31 | 9 | IAC/UMR AMAP |
| La Guen | 573 | 79 | 6 | [26] |
| Riviere Bleue Alluvions | 159 | 103 | NA | [25] |
| Riviere Bleue Pente | 176 | 131 | NA | [25] |
| Tiwae | 244 | 99 | 2 | [26] |
| Wekori | 62 | 74 | 3 | IAC/UMR AMAP |
| Statistics | SR (dbh ≥ 10 cm) | SR (dbh ≥ 40 cm) |
|---|---|---|
| Full time-series statistics | ||
| 15-year mean | 0.13 | −0.47 |
| 15-year median | 0.21 | 0.16 |
| 15-year variance | −0.20 | −0.42 |
| 15-year 1st quartile | 0.16 | 0.15 |
| 15-year 3rd quartile | 0.23 | 0.10 |
| 15-year minimum | −0.01 | −0.45 |
| 15-year maximum | −0.52 * | 0.00 |
| Seasonal statistics | ||
| Wet season mean | −0.05 | −0.66 ** |
| Wet season median | 0.01 | 0.15 |
| Wet season variance | −0.01 | 0.61 * |
| 15-year 1st quartile | −0.06 | 0.07 |
| 15-year 3rd quartile | 0.07 | 0.06 |
| Wet season minimum | −0.02 | −0.61 * |
| Wet season maximum | −0.45 | 0.01 |
| Dry season mean | 0.29 | −0.21 |
| Dry season median | 0.34 | 0.17 |
| Dry season variance | −0.28 | 0.24 |
| 15-year 1st quartile | 0.31 | 0.21 |
| 15-year 3rd quartile | 0.36 | 0.16 |
| Dry season minimum | 0.01 | −0.33 |
| Dry season maximum | −0.51 * | 0.01 |
| Annual statistics | ||
| 2002 mean | 0.51 * | 0.19 |
| 2002 variance | −0.10 | 0.16 |
| 2003 mean | 0.15 | 0.22 |
| 2003 variance | 0.31 | 0.16 |
| 2004 mean | 0.47 * | 0.28 |
| 2004 variance | −0.34 | −0.28 |
| 2005 mean | −0.06 | 0.02 |
| 2005 variance | −0.05 | −0.25 |
| 2006 mean | 0.37 | 0.13 |
| 2006 variance | −0.44 | −0.14 |
| 2007 mean | 0.34 | 0.10 |
| 2007 variance | 0.13 | 0.37 |
| 2008 mean | −0.02 | −0.00 |
| 2008 variance | 0.21 | 0.45 |
| 2009 mean | 0.13 | 0.09 |
| 2009 variance | 0.09 | −0.14 |
| 2010 mean | 0.12 | 0.15 |
| 2010 variance | 0.47 * | 0.07 |
| 2011 mean | 0.23 | 0.24 |
| 2011 variance | −0.04 | 0.04 |
| 2012 mean | −0.06 | 0.11 |
| 2012 variance | 0.24 | 0.20 |
| 2013 mean | −0.11 | −0.11 |
| 2013 variance | 0.28 | 0.27 |
| 2014 mean | 0.12 | 0.23 |
| 2014 variance | 0.36 | −0.14 |
| 2015 mean | 0.18 | 0.02 |
| 2015 variance | −0.05 | 0.07 |
| 2016 mean | 0.20 | −0.04 |
| 2016 variance | −0.03 | −0.11 |
| 2017 mean | 0.14 | −0.13 |
| 2017 variance | −0.08 | 0.07 |
| Monthly statistics | ||
| January mean | −0.06 | −0.62 * |
| January variance | −0.19 | 0.48 |
| February mean | −0.13 | −0.72 ** |
| February variance | 0.02 | 0.65 ** |
| March mean | −0.21 | −0.78 *** |
| March variance | 0.23 | 0.71 ** |
| April mean | −0.06 | −0.68 ** |
| April variance | 0.26 | 0.65 ** |
| May mean | 0.06 | −0.58 * |
| May variance | 0.32 | 0.60 * |
| June mean | 0.24 | −0.36 |
| June variance | 0.36 | 0.59 * |
| July mean | 0.35 | −0.17 |
| July variance | 0.39 | 0.53 * |
| August mean | 0.21 | −0.17 |
| August variance | −0.27 | 0.21 |
| September mean | 0.16 | −0.11 |
| September variance | −0.31 | 0.20 |
| October mean | 0.30 | −0.15 |
| October variance | −0.34 | 0.21 |
| November mean | 0.24 | −0.28 |
| November variance | −0.37 | 0.27 |
| December mean | 0.08 | −0.47 |
| December variance | −0.32 | 0.37 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouteau, R.; Gillespie, T.W.; Birnbaum, P. Predicting Tropical Tree Species Richness from Normalized Difference Vegetation Index Time Series: The Devil Is Perhaps Not in the Detail. Remote Sens. 2018, 10, 698. https://doi.org/10.3390/rs10050698
Pouteau R, Gillespie TW, Birnbaum P. Predicting Tropical Tree Species Richness from Normalized Difference Vegetation Index Time Series: The Devil Is Perhaps Not in the Detail. Remote Sensing. 2018; 10(5):698. https://doi.org/10.3390/rs10050698
Chicago/Turabian StylePouteau, Robin, Thomas W. Gillespie, and Philippe Birnbaum. 2018. "Predicting Tropical Tree Species Richness from Normalized Difference Vegetation Index Time Series: The Devil Is Perhaps Not in the Detail" Remote Sensing 10, no. 5: 698. https://doi.org/10.3390/rs10050698
APA StylePouteau, R., Gillespie, T. W., & Birnbaum, P. (2018). Predicting Tropical Tree Species Richness from Normalized Difference Vegetation Index Time Series: The Devil Is Perhaps Not in the Detail. Remote Sensing, 10(5), 698. https://doi.org/10.3390/rs10050698