Microphytobenthos Biomass and Diversity Mapping at Different Spatial Scales with a Hyperspectral Optical Model

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Hyperspectral Measurements and Image Processing
2.2. Laboratory Experimentations
2.3. Ground and Airborne Field Studies
2.3.1. Muddy Site
2.3.2. Sandy Site
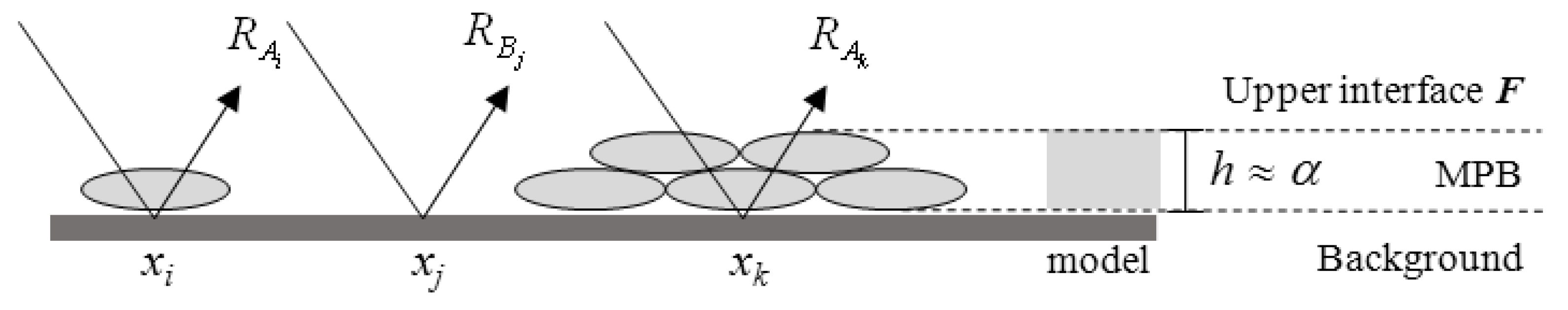
2.4. The MicroPhytoBenthos Optical Model
2.4.1. Optical Properties
2.4.2. Background Estimation
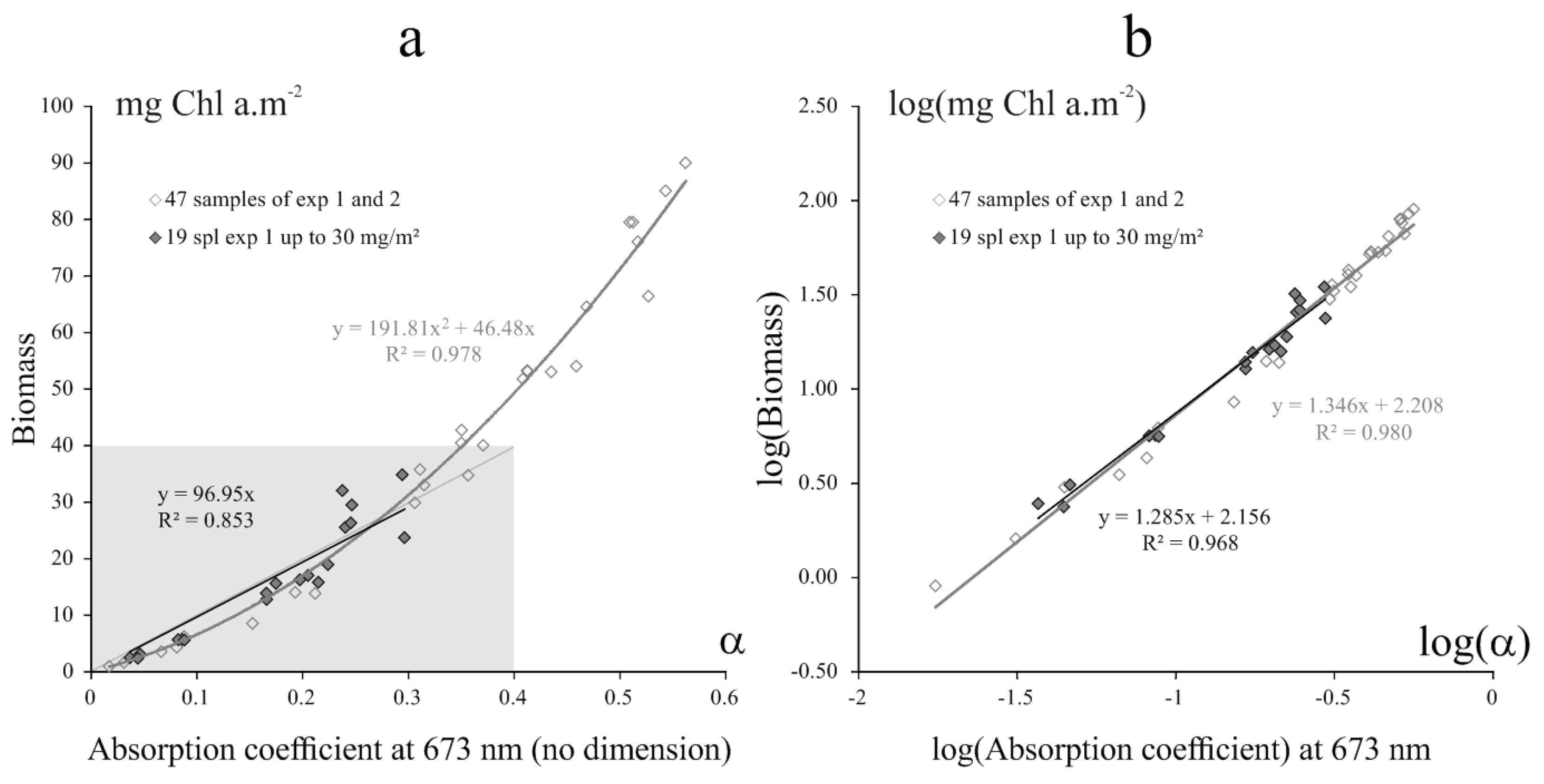
2.4.3. Biomass Estimation
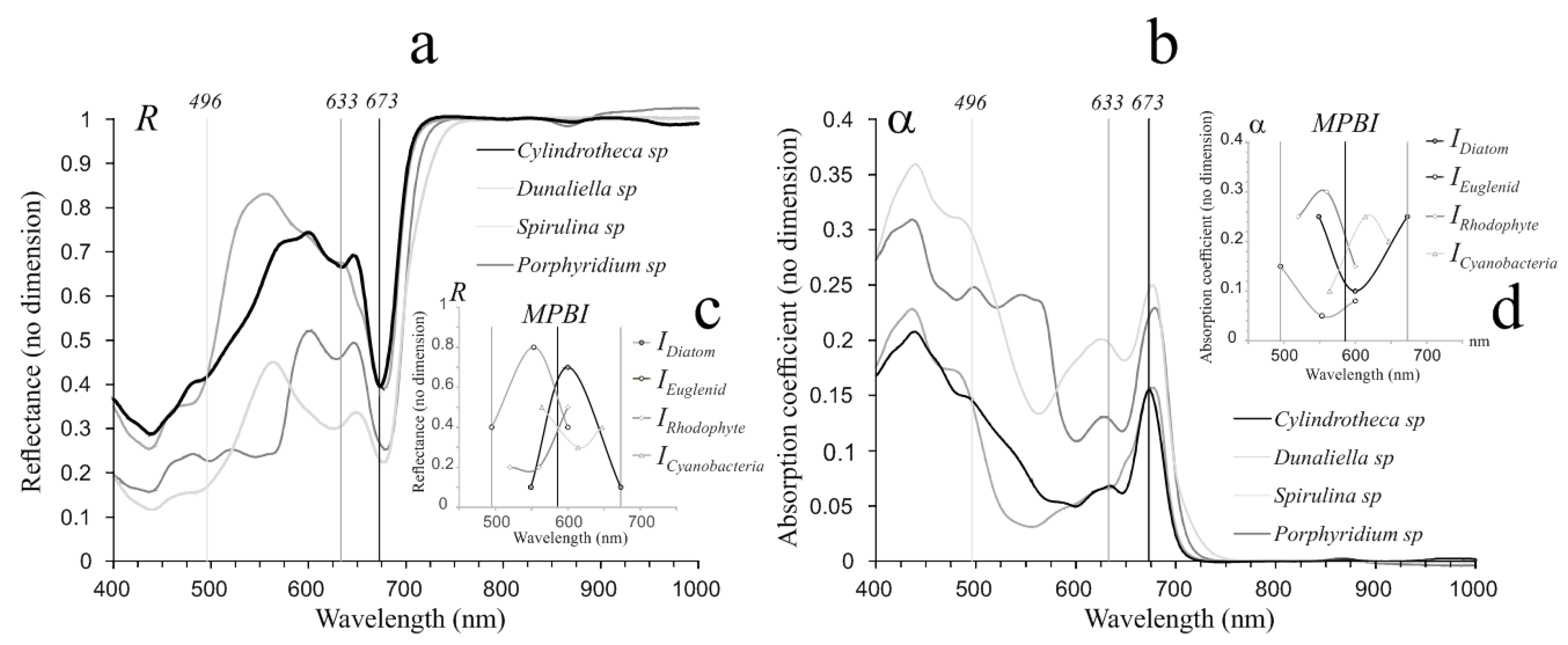
2.4.4. MPB Groups Identification by R and α Spectral Indices
3. Results
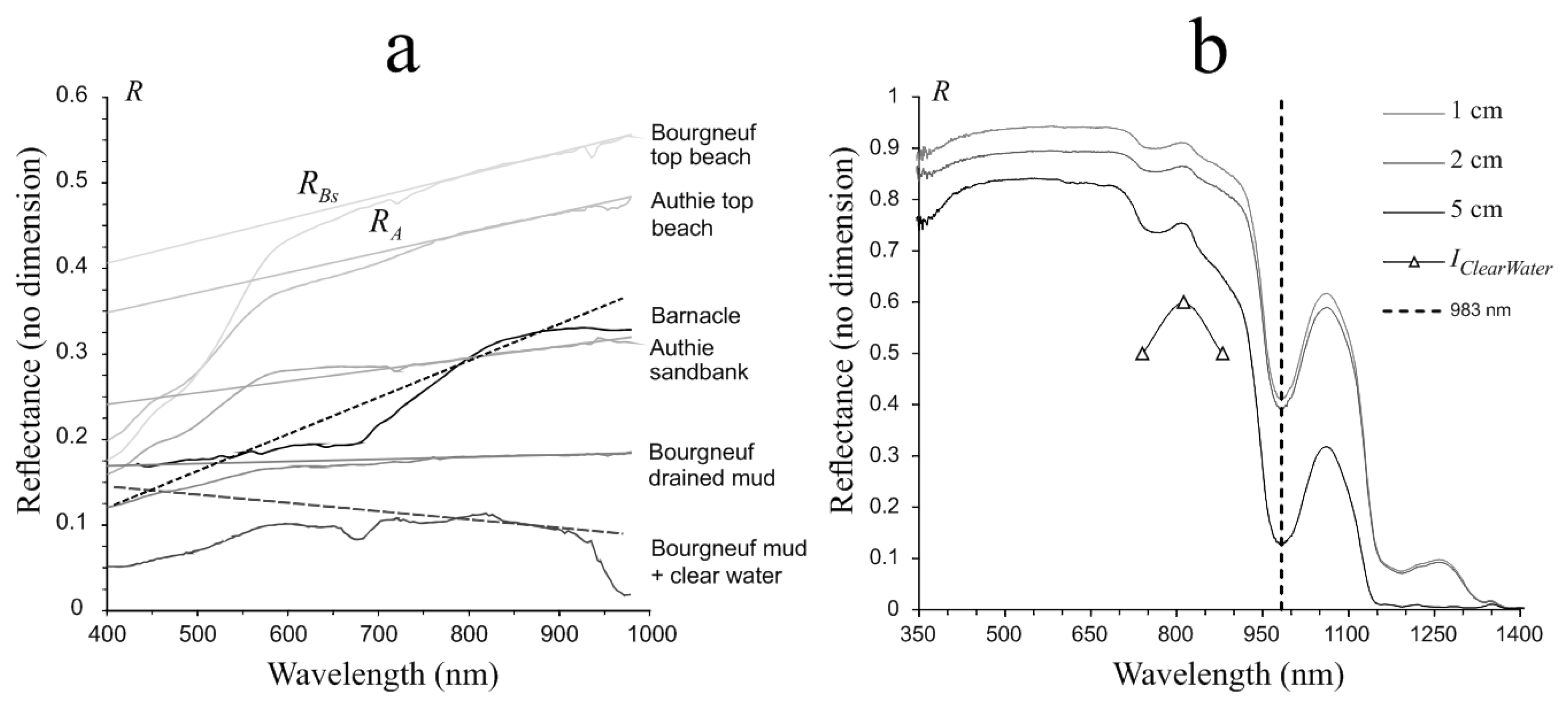
3.1. Spectra Analyses
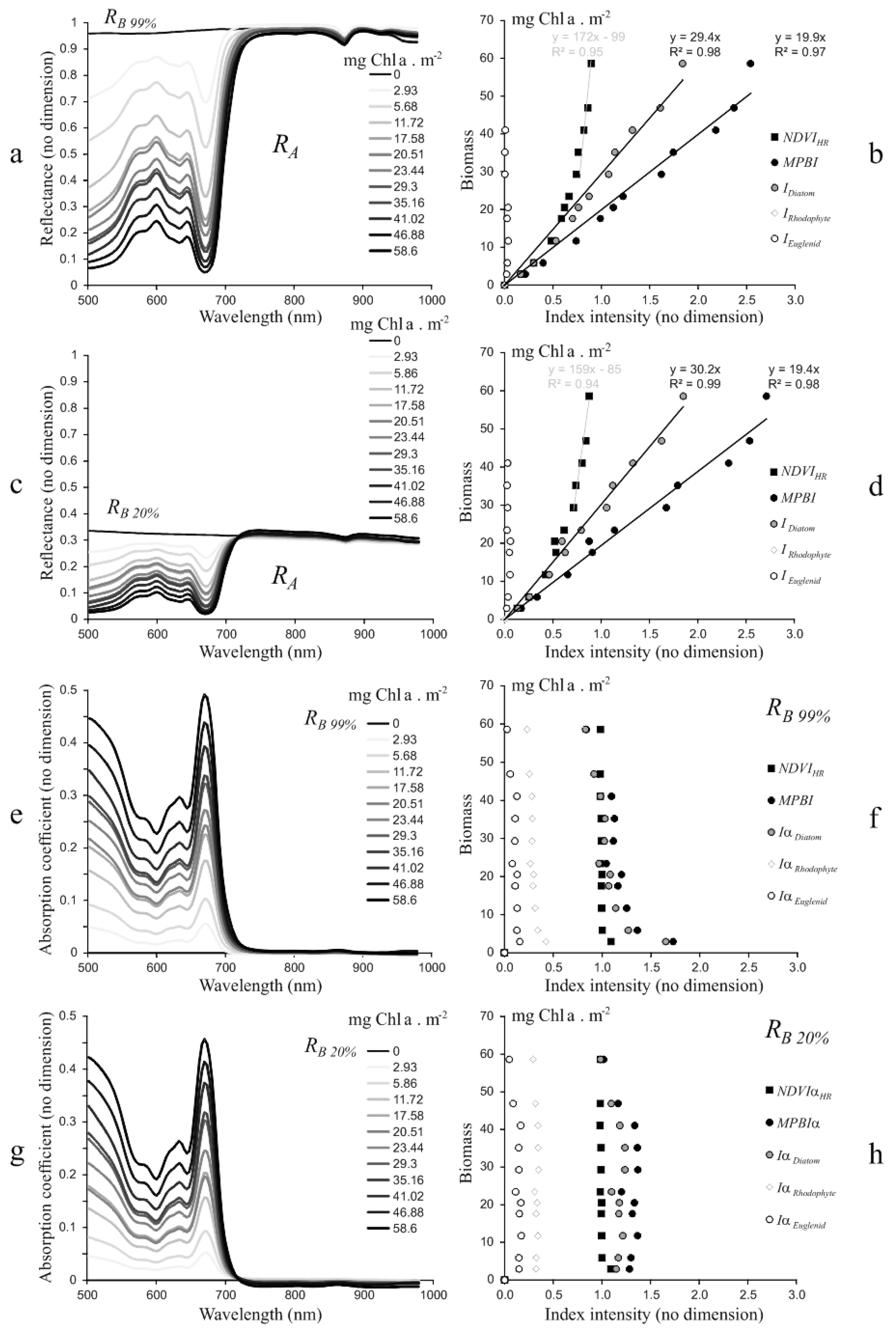
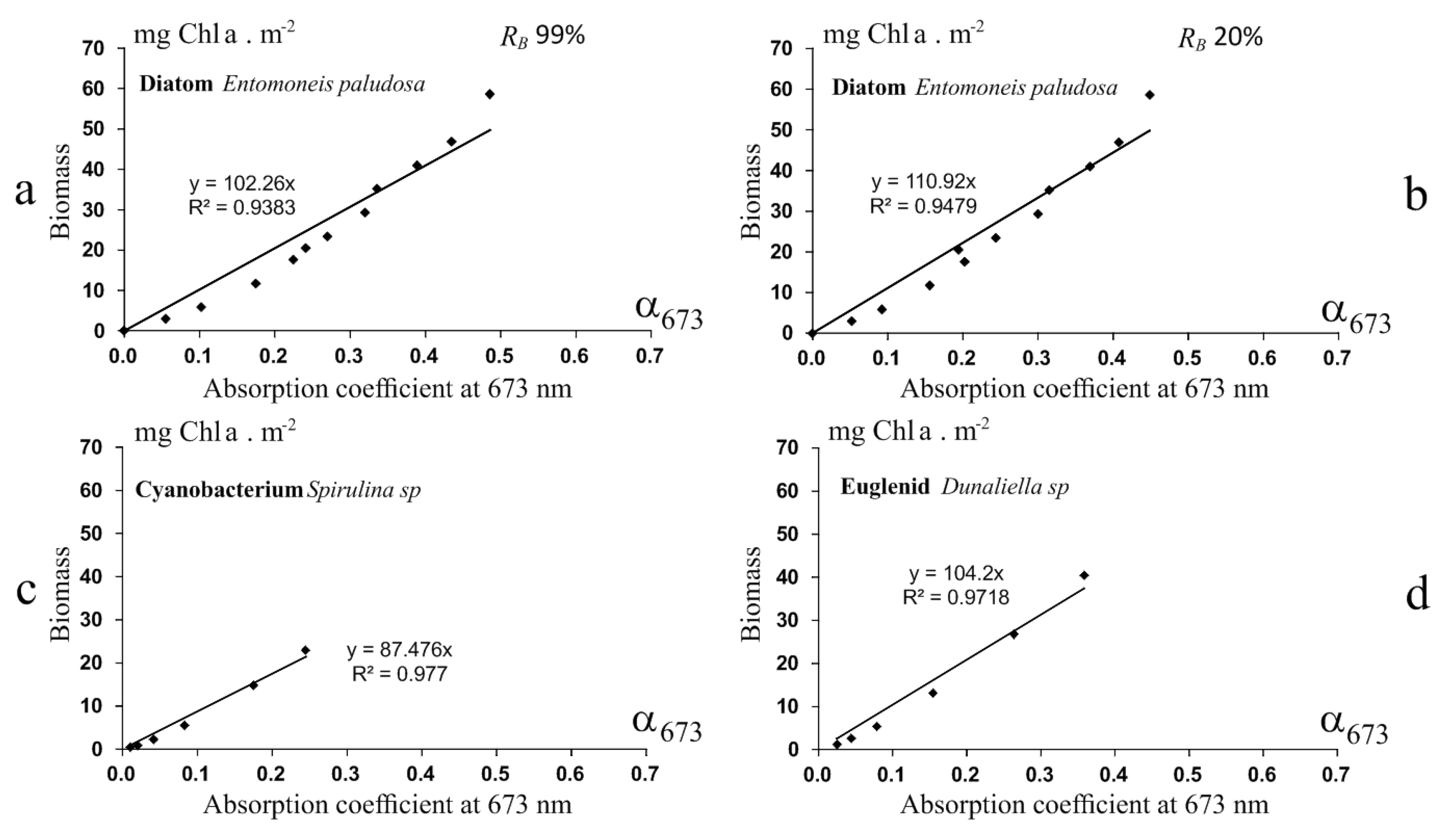
3.1.1. Biofilm Algal Composition and Biomass Estimation
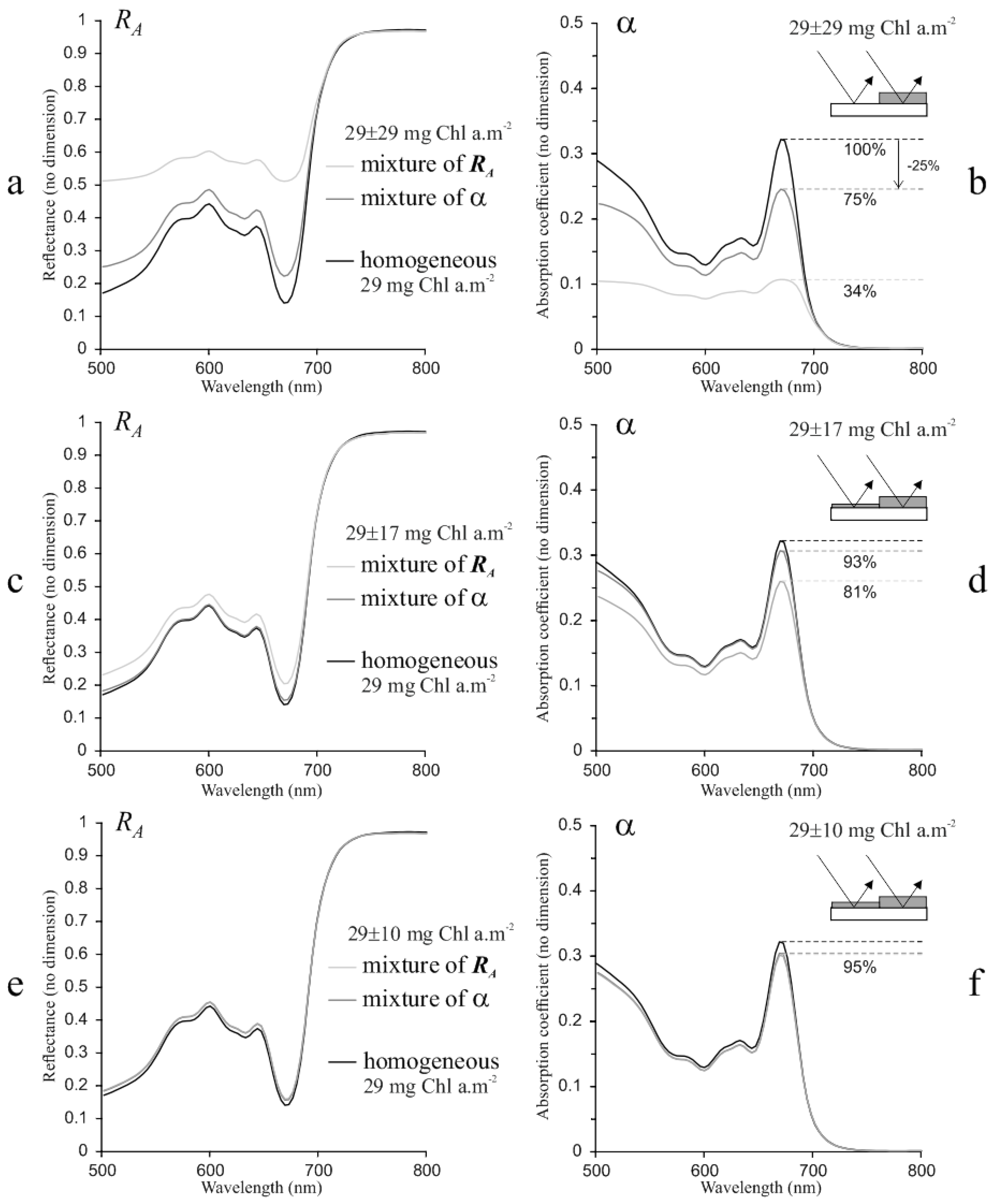
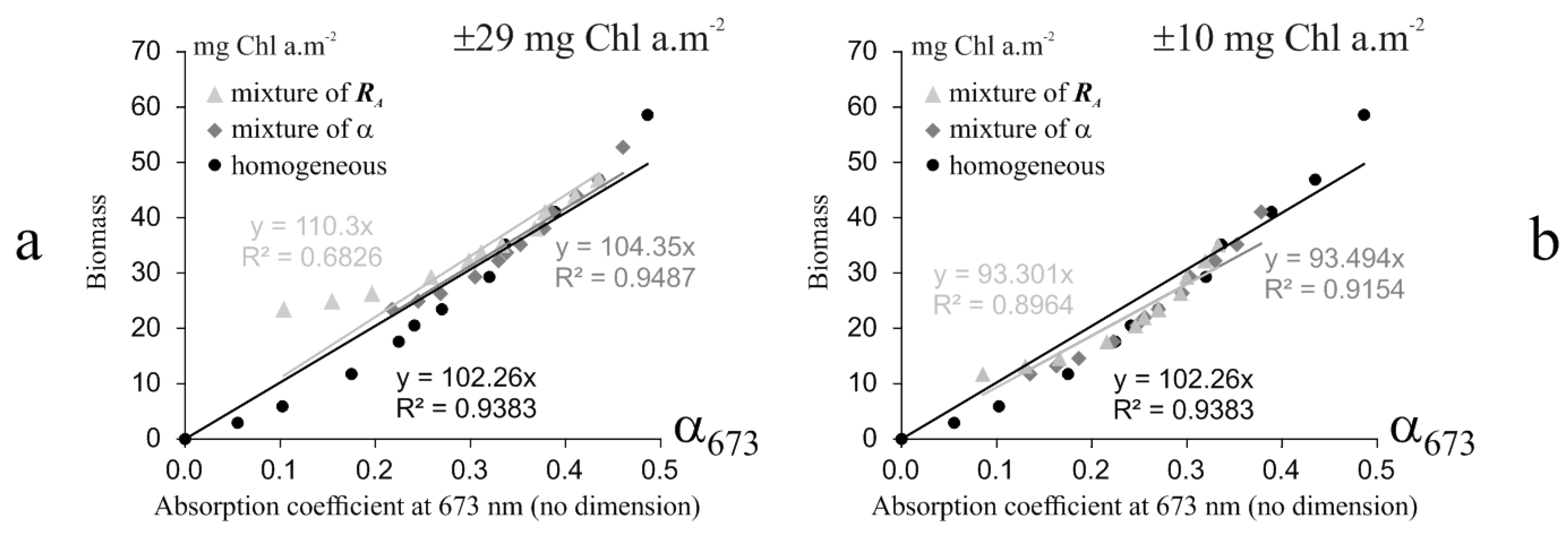
3.1.2. Subpixel Mixing Effects
3.2. Images Analyses
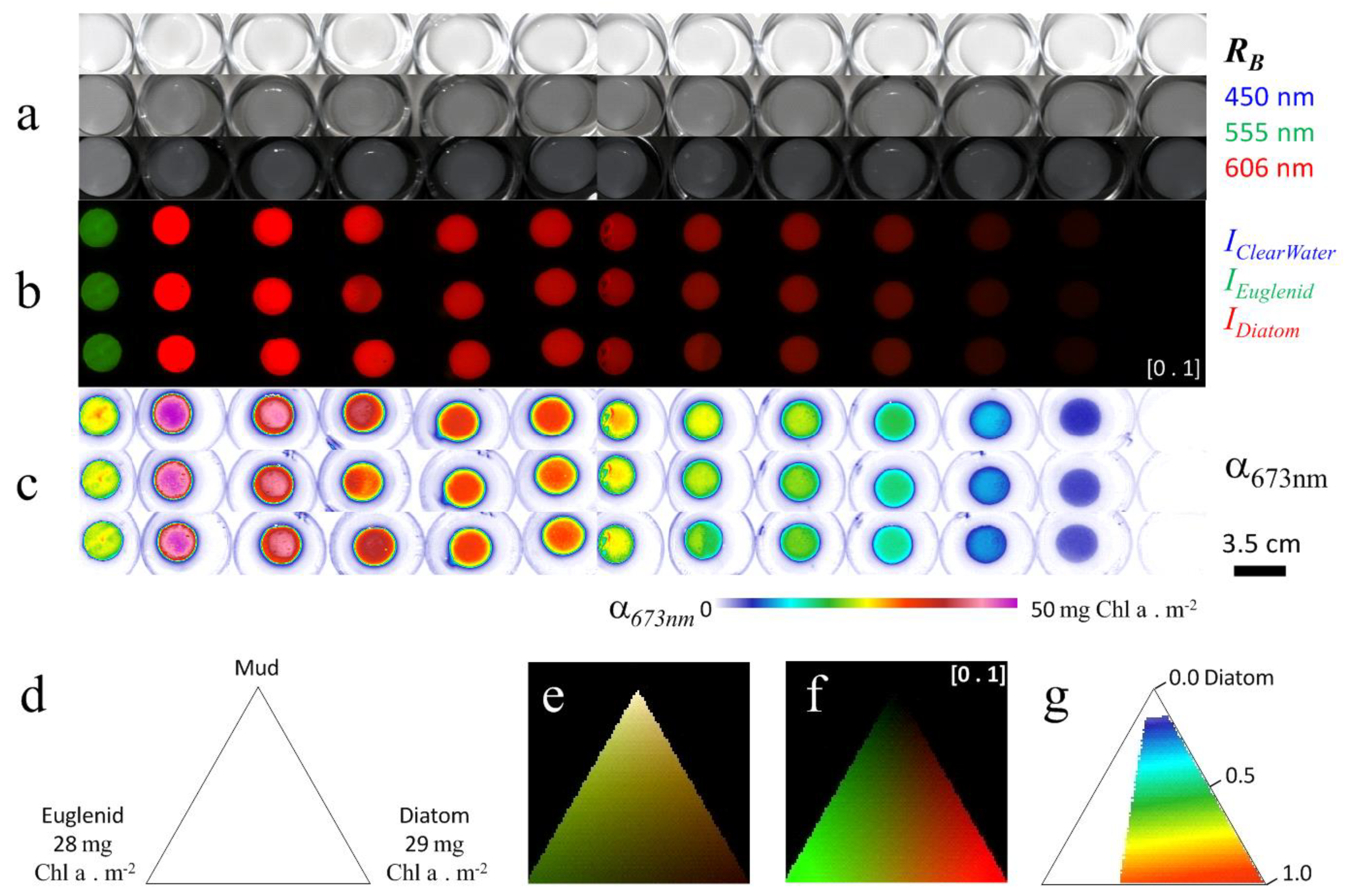
3.2.1. Laboratory Experimentations
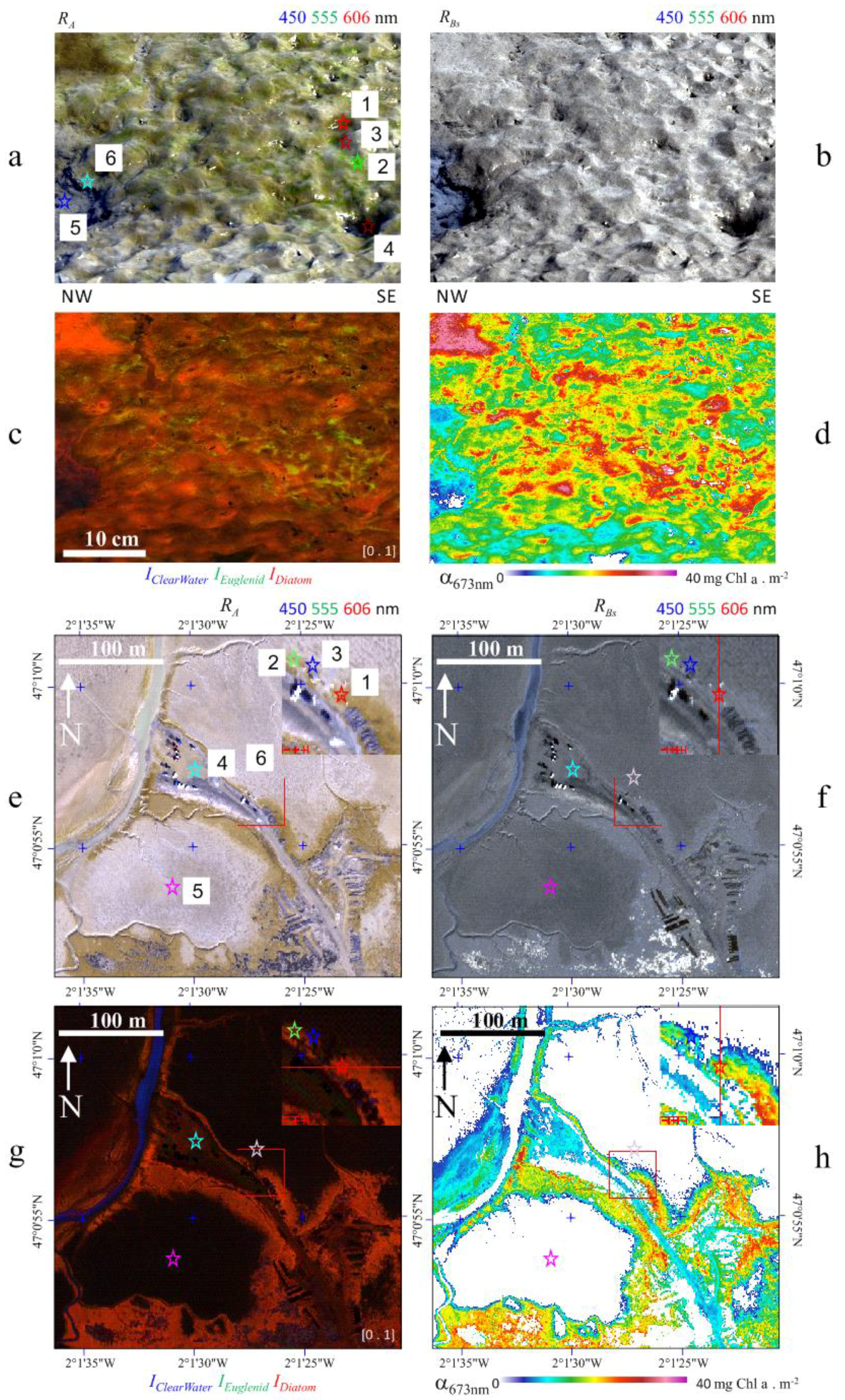
3.2.2. Ground Field Image
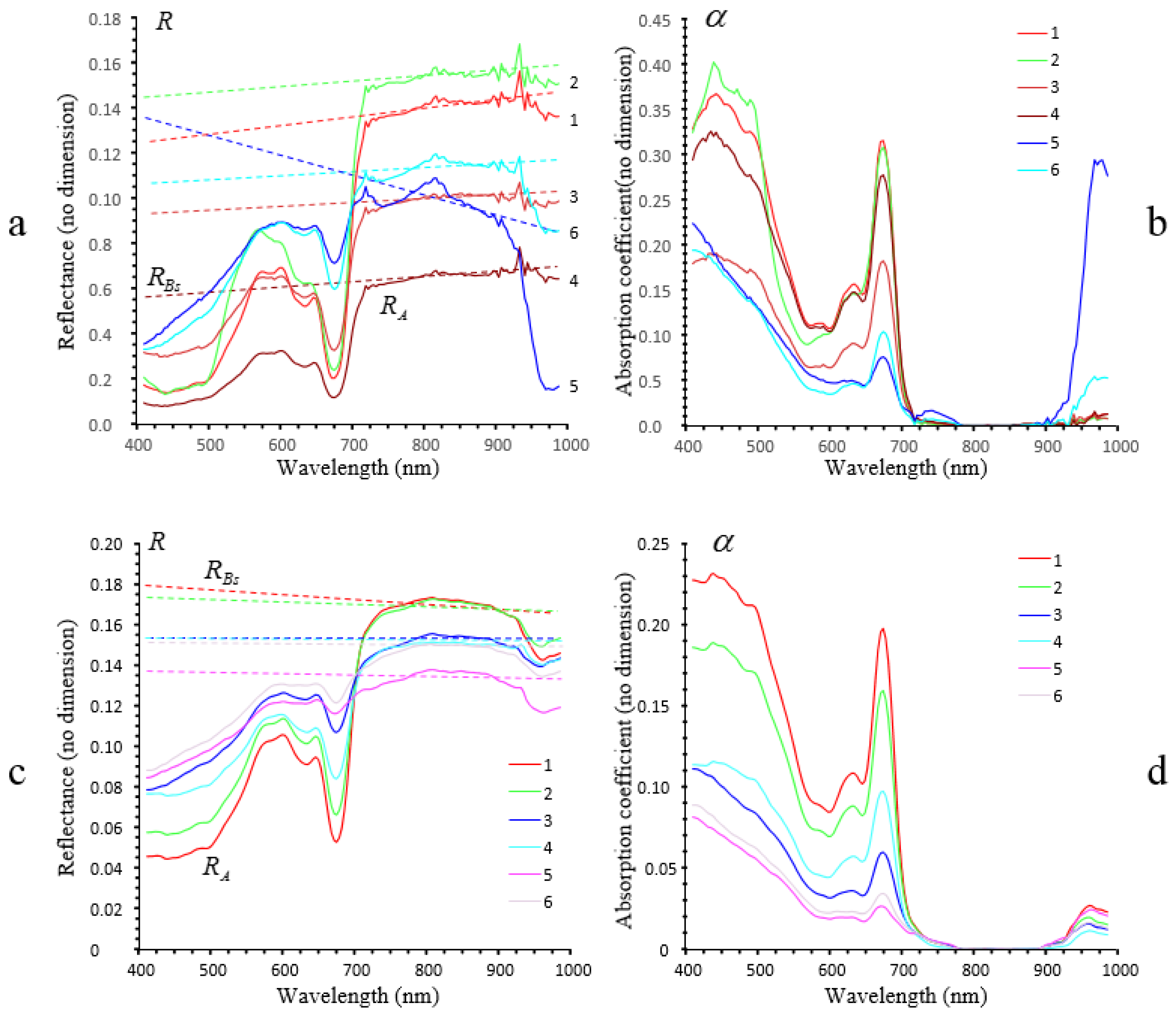
3.2.3. Airborne Imaging: Mudflat Case Study
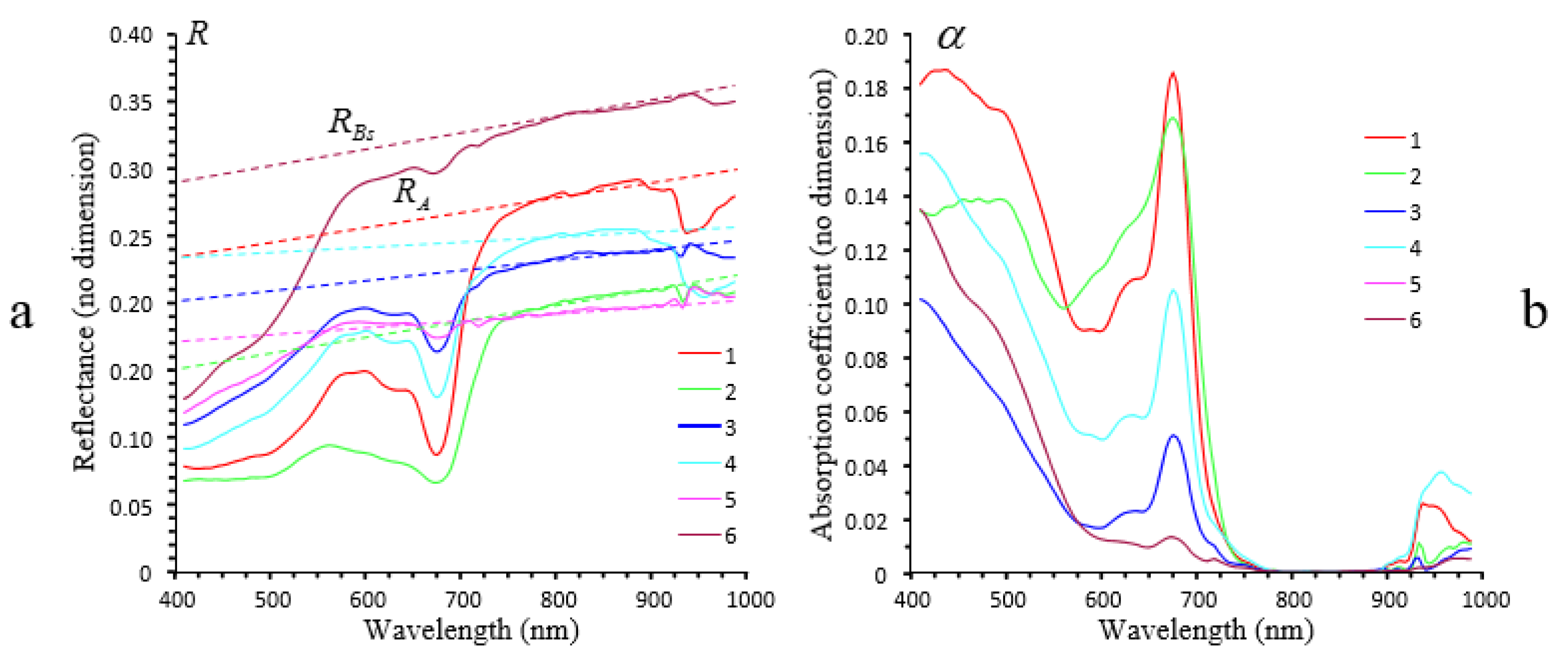
3.2.4. Airborne Imaging: Sandy Case Study
4. Discussion
4.1. A Simplified Optical Model of MPB Relying on Background
4.2. Identification of the Main MPB Groups
4.3. Sub-Pixel Mixing
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kazemipour, F.; Méléder, V.; Launeau, P. Optical properties of microphytobenthic biofilms (MPBOM): Biomass retrieval implication. J. Quant. Spectrosc. Radiat. Transf. 2011, 112, 131–142. [Google Scholar] [CrossRef]
- Kazemipour, F.; Launeau, P.; Méléder, V. Microphytobenthos biomass mapping using the optical model of diatom biofilms: Application to hyperspectral images of Bourgneuf Bay. Remote Sens. Environ. 2012, 127, 1–13. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Smith, D.J. Predicting Epipelic Diatom Exopolymer Concentration in Intertidal Sediments from Sediment Chlorophyll a. Microb. Ecol. 1998, 35, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Heip, C.H.R; Goosen, N.K.; Herman, P.M.J.; Kromkamp, J.; Middelburg, J.J.; Soetaert, K. Production and consumption of biological particles in temperate tidal estuaries. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 1–149. [Google Scholar]
- MacIntyre, H.; Geider, R.; Miller, D. Microphytobenthos: The ecological role of the “secret garden” of unvegetated, shallow water marine habitats. I. Distribution, abundance and primary production. Estuaries 1996, 19, 186–201. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Murray, S.A.; Chomerat, N.; Horiguchi, T. Marine Benthic Dinoflagellates: Unveiling Their Worldwide Biodiversity; Schweizerbart Science Publishers: Stuttgart, Germany, 2014; 276p. [Google Scholar]
- Barillé, L.; Le Bris, A.; Méléder, V.; Launeau, P.; Robin, M.; Louvrou, I.; Ribeiro, L. Photosynthetic epibionts and endobionts of pacific oyster shells from oyster reefs in rocky versus mudflat shores. PLoS ONE 2017, 12, e0185187. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.W. Application of pigment method to oceanography. In Phytoplankton Pigments in Oceanography. Monographs on Oceanographic Methodology; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 127–166. [Google Scholar]
- Consalvey, M.; Jesus, B.; Perkins, R.G.; Brotas, V.; Underwood, G.J.C.; Paterson, D.M. Monitoring migration and measuring biomass in benthic biofilms: The effects of dark/far-red adaptation and vertical migration on fluorescence measurements. Photosynth. Res. 2004, 81, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Pinckney, J.; Zingmark, R.G. Biomass and Production of Benthic Microalgal Communities in Estuarine Habitats. Estuaries 1993, 16, 887–897. [Google Scholar] [CrossRef]
- Paterson, D.M.; Black, K.S. Water flow, sediment dynamics and benthic biology. Adv. Ecol. Res. 1999, 29, 155–193. [Google Scholar]
- De Jonge, V.N.; Van Beuselom, J.E.E. Contribution of resuspended microphytobenthos to total phytoplankton in the EMS estuary and its possible role for grazers. Neth. J. Sea Res. 1992, 30, 91–105. [Google Scholar] [CrossRef]
- Hernández Fariñas, T.; Ribeiro, L.; Soudant, D.; Belin, C.; Lampert, L.; Bacher, C.; Barillé, L. Contribution of benthic microalgae to the temporal variation in phytoplanktonic assemblages in a macrotidal system. J. Phycol. 2017, 5, 1020–1034. [Google Scholar] [CrossRef] [PubMed]
- Decottignies, P.; Beninger, P.G.; Rincé, Y.; Riera, P. Trophic interactions between two introduced suspension-feeders, Crepidula fornicata and Crassostrea gigas, are influenced by seasonal effects and qualitative selection capacity. J. Exp. Mar. Biol. Ecol. 2007, 42, 231–241. [Google Scholar] [CrossRef]
- Riera, P. Trophic plasticity of the gastropod Hydrobia ulvae within an intertidal bay (Roscoff, France): A stable isotope evidence. J. Sea Res. 2010, 63, 78–83. [Google Scholar] [CrossRef]
- Cartaxana, P.; Jesus, B.; Brotas, V. Pheophorbide and pheophytin a-like pigments as useful markers for intertidal microphytobenthos grazing by Hydrobia ulvae. Estuar. Coast. Shelf Sci. 2003, 58, 293–297. [Google Scholar] [CrossRef]
- Kuwae, T.; Beninger, P.G.; Decottignies, P.; Mathot, K.J.; Loud, D.; Elner, R.W. Biofilm grazing in a higher vertebrate: The Western Sandpiper Caliders mauri. Ecology 2008, 89, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Forster, R.M.; Kromkamp, J.C. Estimating benthic primary production: Scaling up from point measurements to the whole estuary2006. In Functioning of Microphytobenthos in Estuaries; Kromkamp, J.C., de Brouwer, J.F.C., Blanchard, G.F., Forster, R.M., Creach, V., Eds.; Royal Netherlands Academy of Arts and Sciences: Amsterdam, the Netherlands, 2006; pp. 109–120. [Google Scholar]
- Ubertini, M.; Lefebvre, S.; Gangnery, A.; Grangeré, K.; Gendre, R.L.; Orvain, F. Spatial variability of benthic-pelagic coupling in an estuary ecosystem: Consequences for microphytobenthos resuspension phenomenon. PLoS ONE 2012, 7, e44155. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Benyoucef, I.; Blandin, E.; Lerouxel, A.; Jesus, B.; Rosa, P.; Meleder, V.; Launeau, P.; Barillé, L. Microphytobenthos interannual variations in a north-European estuary (Loire estuary, France) detected by visible-infrared multispectral remote sensing. Estuar. Coast. Shelf Sci. 2014, 136, 43–52. [Google Scholar] [CrossRef]
- Brito, A.C.; Benyoucef, I.; Jesus, B.; Brotas, V.; Gernez, P.; Mendes, C.R.; Launeau, P.; Dias, P.; Barillé, L. Seasonality of microphytobenthos revealed by remote-sensing in a South European estuary. Cont. Shelf Res. 2013, 66, 83–91. [Google Scholar] [CrossRef]
- Méléder, V.; Barillé, L.; Launeau, P.; Carrère, V.; Rincé, Y. Spectrometric constraint in analysis of benthic diatom biomass using monospecific cultures. Remote Sens. Environ. 2003, 88, 386–400. [Google Scholar] [CrossRef]
- Van der Wal, D.; Wielemaker-van den Dool, A.; Herman, P.M. Spatial synchrony in intertidal benthic algal biomass in temperate coastal and estuarine ecosystems. Ecosystems 2010, 13, 338–351. [Google Scholar] [CrossRef]
- Combe, J.P.; Launeau, P.; Carrere, V.; Despan, D.; Meleder, V.; Barillé, L.; Sotin, C. Mapping microphytobenthos biomass by non-linear inversion of visible-infrared hyperspectral images. Remote Sens. Environ. 2005, 98, 371–387. [Google Scholar] [CrossRef]
- Jesus, B.; Rosa, P.; Mouget, J.-L.; Meleder, V.; Launeau, P.; Barillé, L. Spectral-radiometric analysis of taxonomically mixed microphytobenthic biofilms. Remote Sens. Environ. 2014, 140, 196–205. [Google Scholar] [CrossRef]
- Chennu, A.; Färber, P.; Volkenborn, P.; Al-Najjar, M.A.A.; Janssen, F.; de Beer, D.; Polerecky, L. Hyperspectral imaging of the microscale distribution and dynamics of microphytobenthos in intertidal sediments. Limnol. Oceanogr. Methods 2013, 11, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Nogami, S.; Ohnuki, S.; Ohya, Y. Hyperspectral imaging techniques for the characterization of haematococcus pluvialis (chlorophyceae). J. Phycol. 2014, 50, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Méléder, V.; Laviale, M.; Jesus, B.; Mouget, J.-L.; Lavaud, J.; Kazemipour, F.; Launeau, P.; Barillé, L. In vivo estimation of pigment composition and optical absorption cross-section by spectroradiometry in four aquatic photosynthetic micro-organisms. J. Photochem. Photobiol. B 2013, 129, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méléder, V.; Launeau, P.; Barillé, L.; Combe, J.P.; Carrère, V.; Jesus, B.; Verpoorter, C. Hyperspectral imaging for mapping microphytobenthos in coastal areas. In Geomatic Solutions for Coastal Environments; Maanan, M., Robin, M., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2010; pp. 71–139. [Google Scholar]
- Saburova, M.A.; Polikarpov, I.G.; Burkovsky, I.V. Spatial structure of an intertidal sandflat microphytobenthic community as related to different spatial scales. Mar. Ecol. Prog. Ser. 1995, 129, 229–239. [Google Scholar] [CrossRef]
- Spilmont, N.; Seuront, L.; Meziane, T.; Welsh, D.T. There’s more to the picture than meets the eye: Sampling microphytobenthos in a heterogeneous environment. Estuar. Coast. Shelf Sci. 2011, 95, 470–476. [Google Scholar] [CrossRef]
- Richter, R.; Schläpfer, D. Atmospheric /Topographic Correction for Airborne Imagery; DLR Report DLR-IB 565-02/14, 240; DLR: Wessling, Germany, 2014. [Google Scholar]
- Green, A.A.; Berman, M.; Switzer, P.; Craig, M.D. A Transformation for Ordering Multispectral Data in Terms of Images Quality with Implications for Noise Removal. IEEE Trans. Geosci. Remote Sens. 1988, 26, 65–74. [Google Scholar] [CrossRef]
- Boardman, J.W.; Kruse, F.A. Automated Spectral Analysis: A Geologic Example using AVIRIS Data, North Grapevine Mountains, Nevada. In Proceedings, ERIM Tenth Thematic Conference on Geologic Remote Sensing; Environmental Research Institute of Michigan: Ann Arbor, MI, USA, 1994; pp. I-407–I-418. [Google Scholar]
- Decottignies, P.; Beninger, P.G.; Rincé, Y.; Robbins, R.J.; Riera, P. Exploitation of natural food sources by two sympatric, invasive suspension-feeders: Crassostrea gigas and Crepidula fornicata. Mar. Ecol. Prog. Ser. 2007, 334, 179–192. [Google Scholar] [CrossRef]
- Dobroniak, C.; Anthony, E.J. Short-term morphological expression of dune sand recycling on a macrotidal, wave-exposed estuarine shoreline. J. Coast. Res. 2002, 36, 240–248. [Google Scholar] [CrossRef]
- Marion, C.; Anthony, E.J.; Trentesaux, A. Short-term estuarine mudflat and salt-marsh sedimentation: High-resolution data from ultrasonic altimetry, rod surface-elevation table, and filter traps. Estuar. Coast. Shelf Sci. 2009, 83, 475–484. [Google Scholar] [CrossRef]
- Pye, K.; French, P. Erosion and Accretion Processes on British salt Marshes. In Introduction: Saltmarsh Processes and Morphology; Cambridge Environmental Research Consultants: Cambridge, UK, 1993; Volume 1. [Google Scholar]
- Deloffre, J.; Verney, R.; Lafite, R.; Lesueur, P.; Lesourd, S.; Cundy, A.B. Sedimentation on intertifal mudflats in the lower part of macrotifal estuaries: Sedimentation rythms and their preservation. Mar. Geol. 2007, 241, 19–32. [Google Scholar] [CrossRef]
- Carrère, V.; Spilmont, N.; Davoult, D. Comparison of simple techniques for estimating chlorophyll a concentration in the intertidal zone using high spectral-resolution field-spectrometer data. Mar. Ecol. Prog. Ser. 2004, 274, 31–40. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. PROSPECT: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Khashan, M.; El-Naggar, A. A new method of finding the optical constants of a solid from the reflectance and transmittance spectrograms of its slab. Opt. Commun. 2000, 174, 445–453. [Google Scholar] [CrossRef]
- Astachov, L.; Nevo, Z.; Brosh, T.; Vago, R. The structural, compositional and mechanical features of the calcite shell of the barnacle Tetraclita rufotincta. J. Struct. Biol. 2011, 175, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, G.; Sakshaug, E. Biooptical characteristics of PSII and PSI in 33 species (13 pigment groups) of marine phytoplankton, and the relevance for pulse-amplitude-modulated and fast-repetition-rate fluorometry1. J. Phycol. 2007, 43, 1236–1251. [Google Scholar] [CrossRef]
- Kazemipour, F. Caractérisation Hyperspectrale des Biofilms Microphytobentiques: Cartographie de la Biomasse de la Micro à la Macro Échelle. Ph.D. Thesis, Université de Nantes, Nantes, France, 2011; p. 204. [Google Scholar]
- Launeau, P.; Girardeau, J.; Sotin, C.; Tubia, J. Comparison between field measurements and airborne visible and infrared mapping spectrometry (AVIRIS and HyMap) of the Ronda peridotite massif (south-west Spain). Int. J. Remote Sens. 2004, 25, 2773–2792. [Google Scholar] [CrossRef]
- Carrère, V.; Conel, J.E. Recovery of atmospheric water vapor total column abundance from imaging spectrometer data around 940 nm-sensitivity analysis and application to Airborne Visible/Infrared Imaging Spectrometer (AVIRIS) data. Remote Sens. Environ. 1993, 44, 179–204. [Google Scholar] [CrossRef]
- Roberts, D.A.; Gardner, M.; Church, R.; Ustin, S.; Scheer, G.; Green, R.O. Mapping chaparral in the Santa Monica Mountains using multiple endmember spectral mixture models. Remote Sens. Environ. 1998, 65, 267–279. [Google Scholar] [CrossRef]
- Barillé, L.; Mouget, J.-L.; Méléder, V.; Rosa, P.; Jesus, B. Spectral response of benthic diatoms with different sediment backgrounds. Remote Sens. Environ. 2011, 115, 1034–1042. [Google Scholar] [CrossRef]
- Paterson, D.M.; Wiltshire, K.H.; Miles, A.; Blackburn, J.; Davison, I.; Yates, M.G.; McGrorty, S.; Eastwood, J.A. Microbiological mediation of spectral reflectance from intertidal cohesive sediments. Limnol. Oceanogr. Methods 1998, 43, 1207–1221. [Google Scholar] [CrossRef]
- Grinham, A.R.; Carruthers, T.J.B.; Fisher, P.L.; Udy, J.W.; Dennison, W.C. Accurately measuring the abundance of benthic microalgae in spatially variable habitats. Limnol. Oceanogr. Methods 2007, 5, 119–125. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reflectance R | Absorption Coefficient α |
|---|---|
| MPB threshold | If (NDVIHR > 0.1 and MPBI > NDVIHR) |
| Diatoms | MPBI > IDiatom > NDVIHR > 0 (IEuglenid, ICyanobacteria, IRhodophyte) |
| Euglena and green microalgae | MPBI > IEuglenid > NDVIHR > IDiatom ≥ ICyanobacteria ≥ 0 (IRhodophyte) |
| Cyanobacteria | MPBI > IEuglenid ≥ NDVIHR > ICyanobacteria ≥ 0 (IDiatom, IRhodophyte) |
| Rhodophyte and red microalgae | IDiatom > MPBI > IRhodophyte ≥ NDVIHR > 0 (ICyanobacteria, IEuglenid) |
| MPB threshold | If (NDVIHR > 0.1 and MPBI > NDVIHR) with NDVIαHR = 1 |
| Diatoms | MPBIα > IαDiatom > IαEuglenid > IαCyanobacteria ≥ 0 (IαRrodophyte) |
| Euglena and green microalgae | MPBIα ≈ IαEuglenid > IαDiatom > IαCyanobacteria ≥ 0 (IαRhodophyte) |
| Cyanobacteria | MPBIα ≈ IαEuglenid > IαCyanobacteria> IαDiatom ≥ 0 (IαRhodophyte) |
| Rhodophyte and red microalgae | IαDiatom > MPBIα > IαRhodophyte ≥ 0 (IαEuglenid, IαCyanobacteria) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Launeau, P.; Méléder, V.; Verpoorter, C.; Barillé, L.; Kazemipour-Ricci, F.; Giraud, M.; Jesus, B.; Le Menn, E. Microphytobenthos Biomass and Diversity Mapping at Different Spatial Scales with a Hyperspectral Optical Model. Remote Sens. 2018, 10, 716. https://doi.org/10.3390/rs10050716
Launeau P, Méléder V, Verpoorter C, Barillé L, Kazemipour-Ricci F, Giraud M, Jesus B, Le Menn E. Microphytobenthos Biomass and Diversity Mapping at Different Spatial Scales with a Hyperspectral Optical Model. Remote Sensing. 2018; 10(5):716. https://doi.org/10.3390/rs10050716
Chicago/Turabian StyleLauneau, Patrick, Vona Méléder, Charles Verpoorter, Laurent Barillé, Farzaneh Kazemipour-Ricci, Manuel Giraud, Bruno Jesus, and Erwan Le Menn. 2018. "Microphytobenthos Biomass and Diversity Mapping at Different Spatial Scales with a Hyperspectral Optical Model" Remote Sensing 10, no. 5: 716. https://doi.org/10.3390/rs10050716