Improvement of Remote Sensing-Based Assessment of Defoliation of Pinus spp. Caused by Thaumetopoea pityocampa Denis and Schiffermüller and Related Environmental Drivers in Southeastern Spain

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Outbreak Area
2.2. Defoliation Data Selection and Quality Control
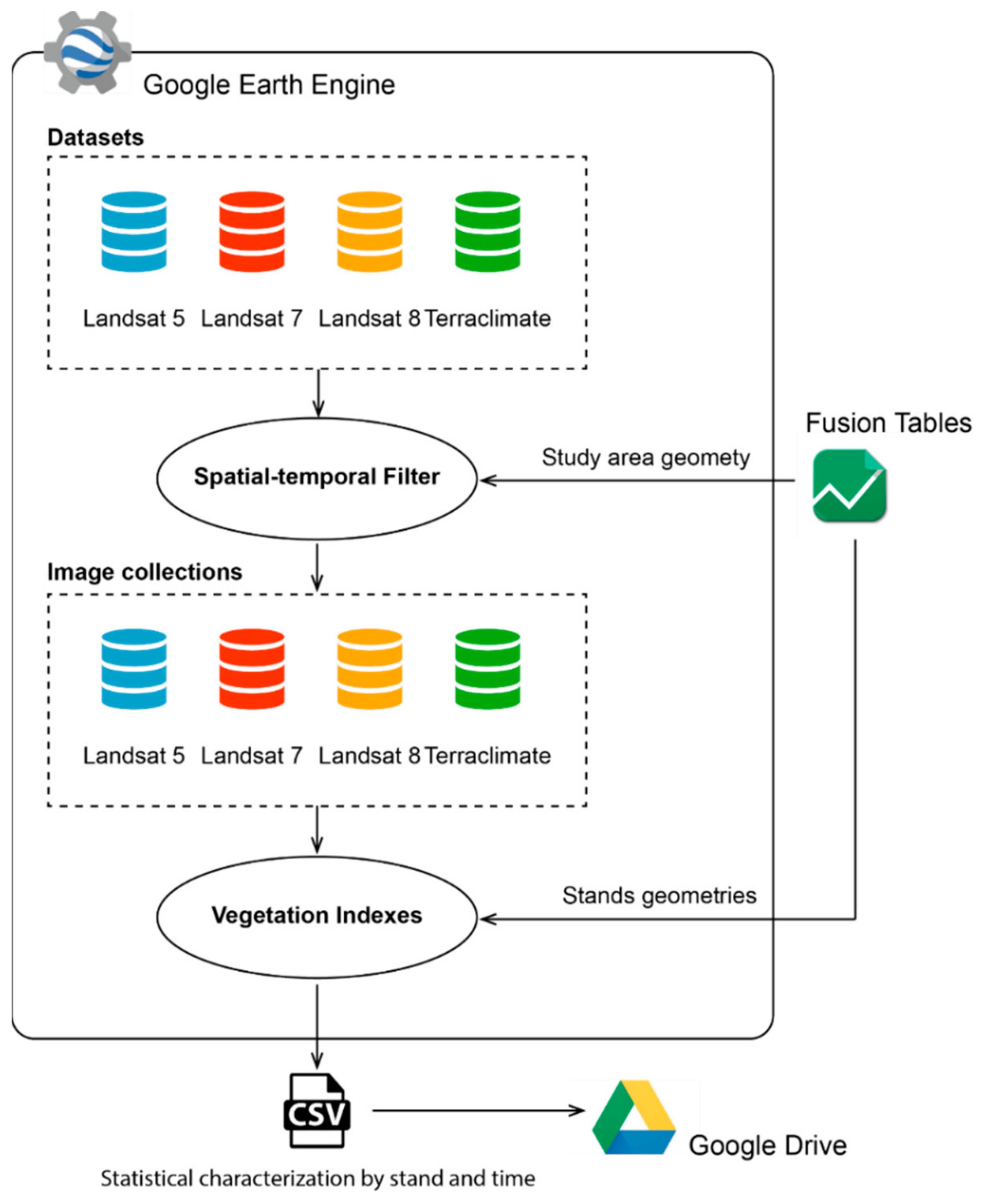
2.3. Datasets and Image Processing
2.4. Environmental Variables
2.5. Statistical Analysis
3. Results
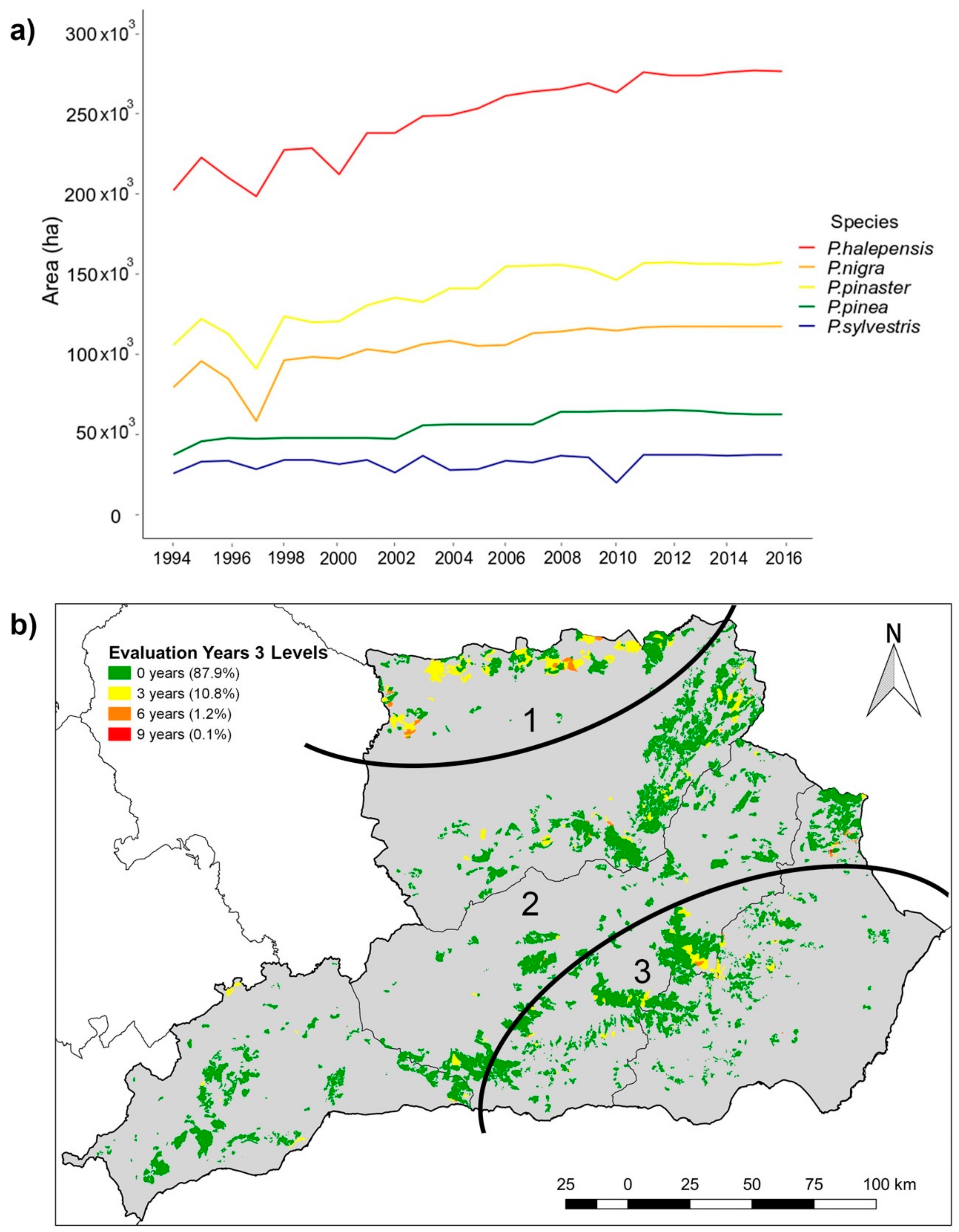
3.1. Defoliation of Individual Species
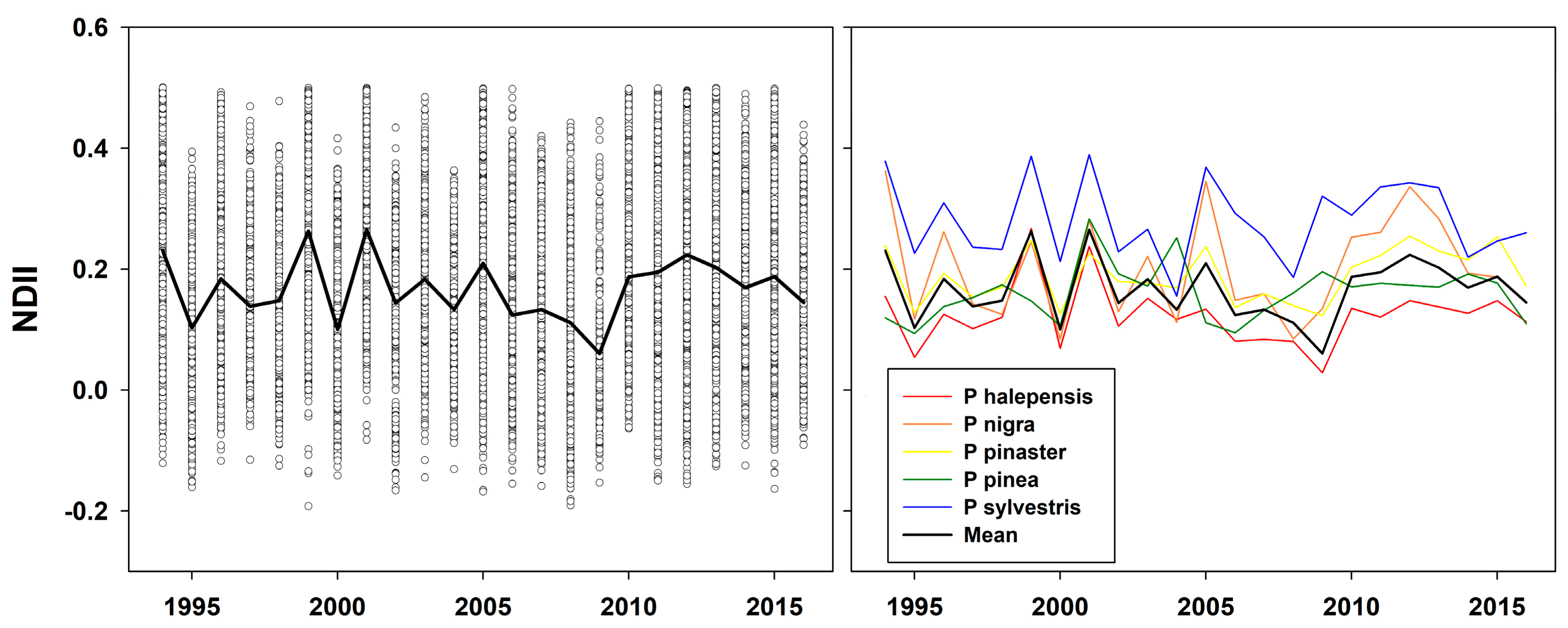
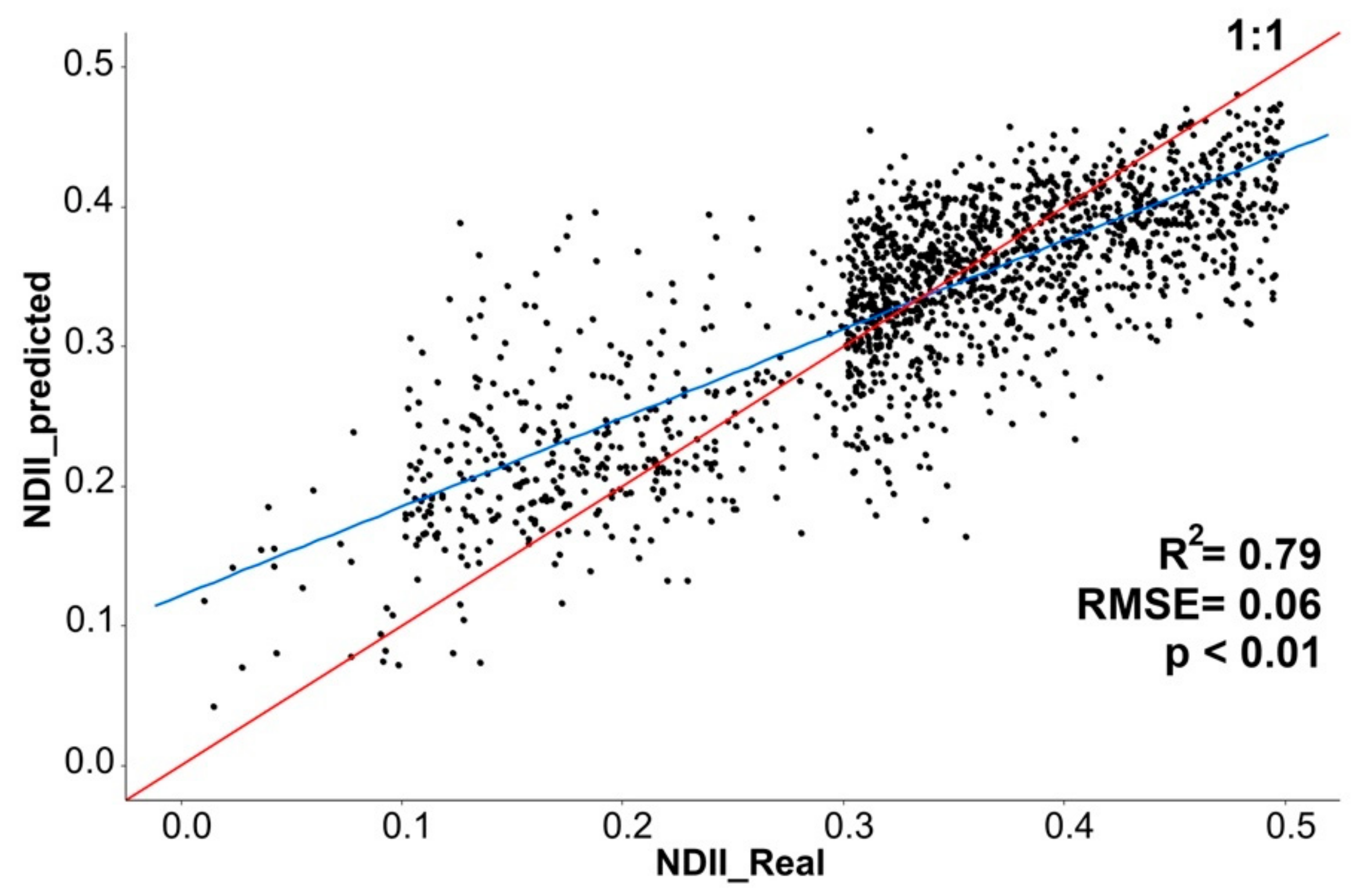
3.2. Vegetation Indexes
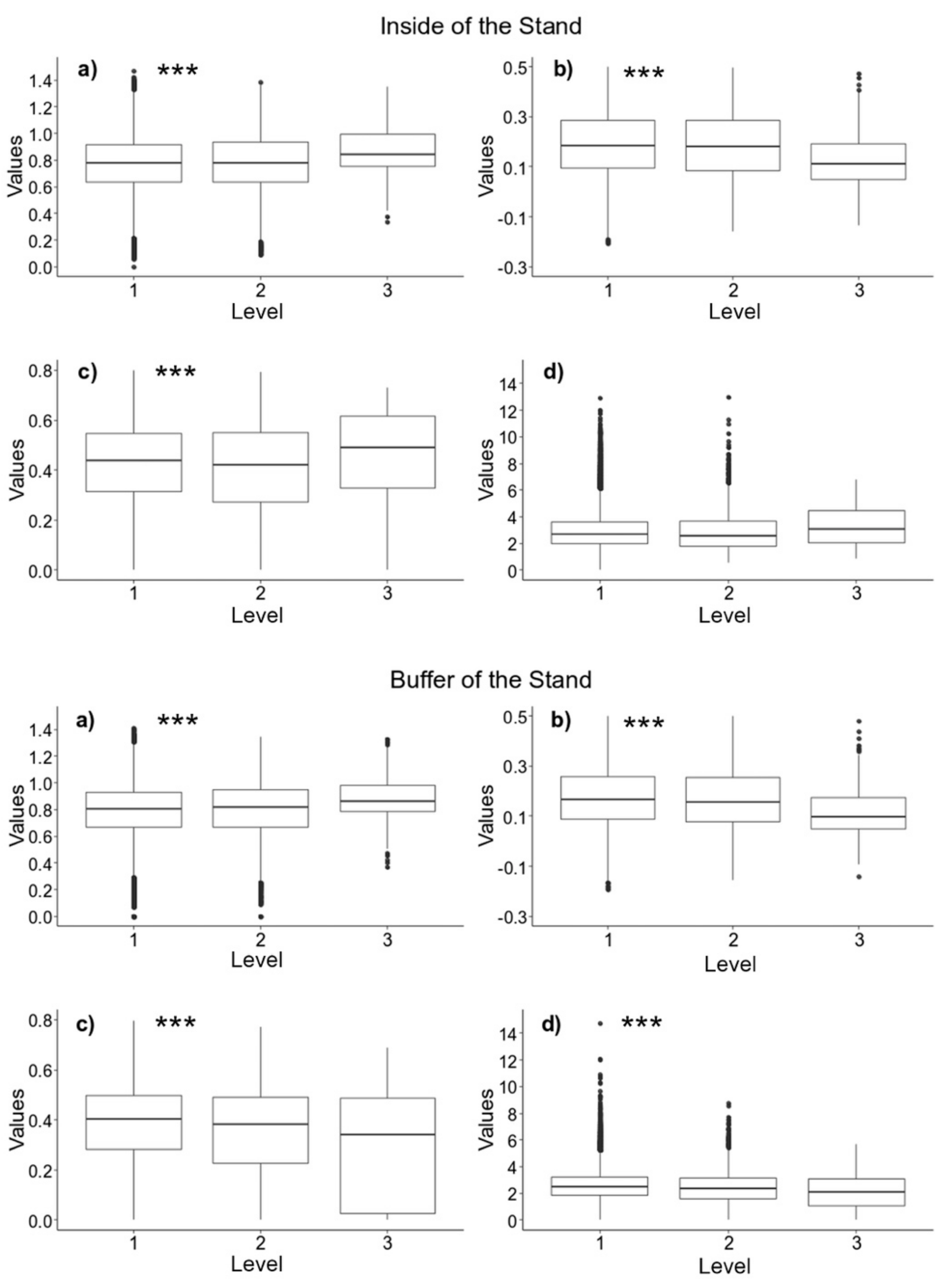
3.3. Synchronization and Defoliation Patterns
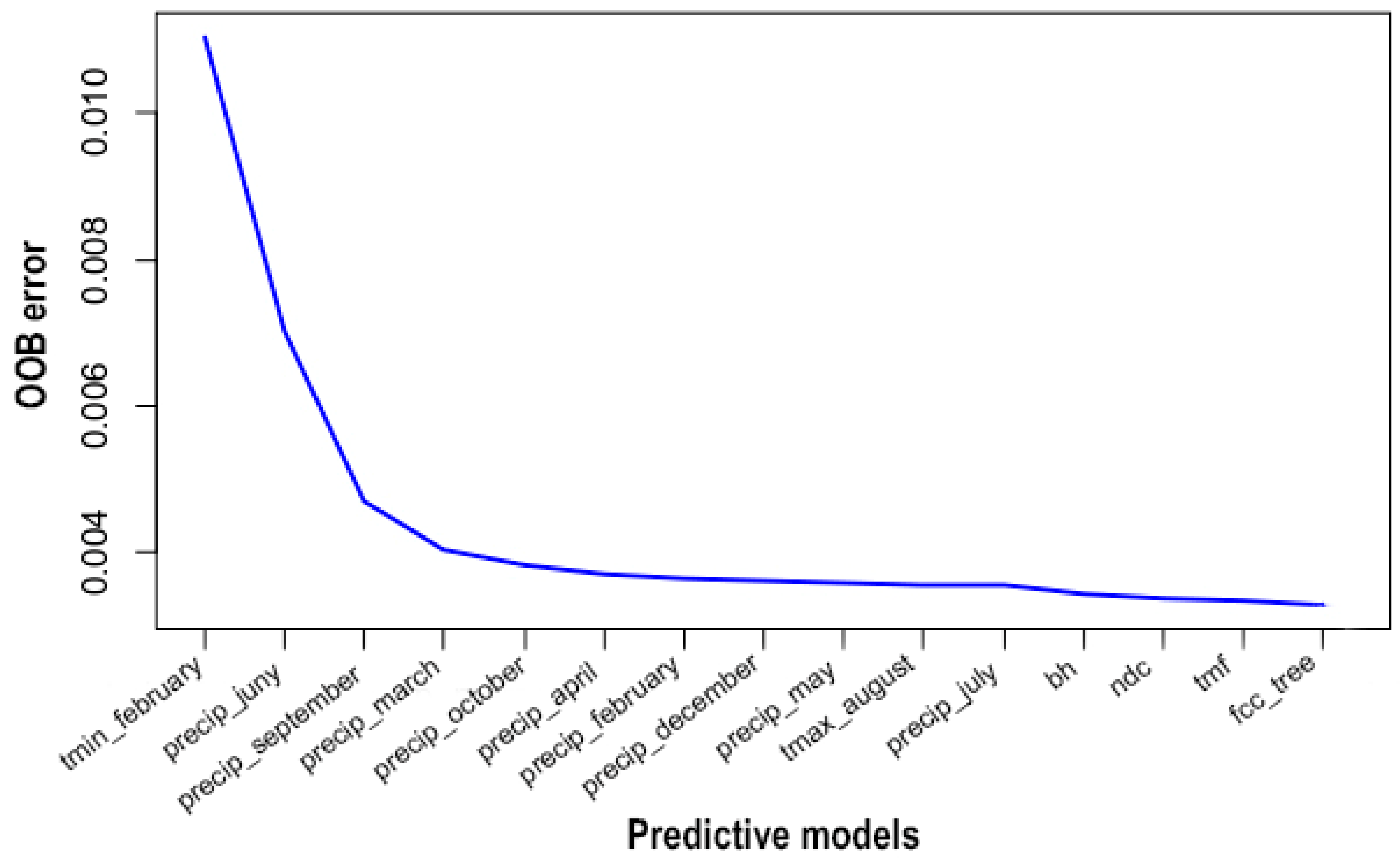
3.4. Environmental Predictors
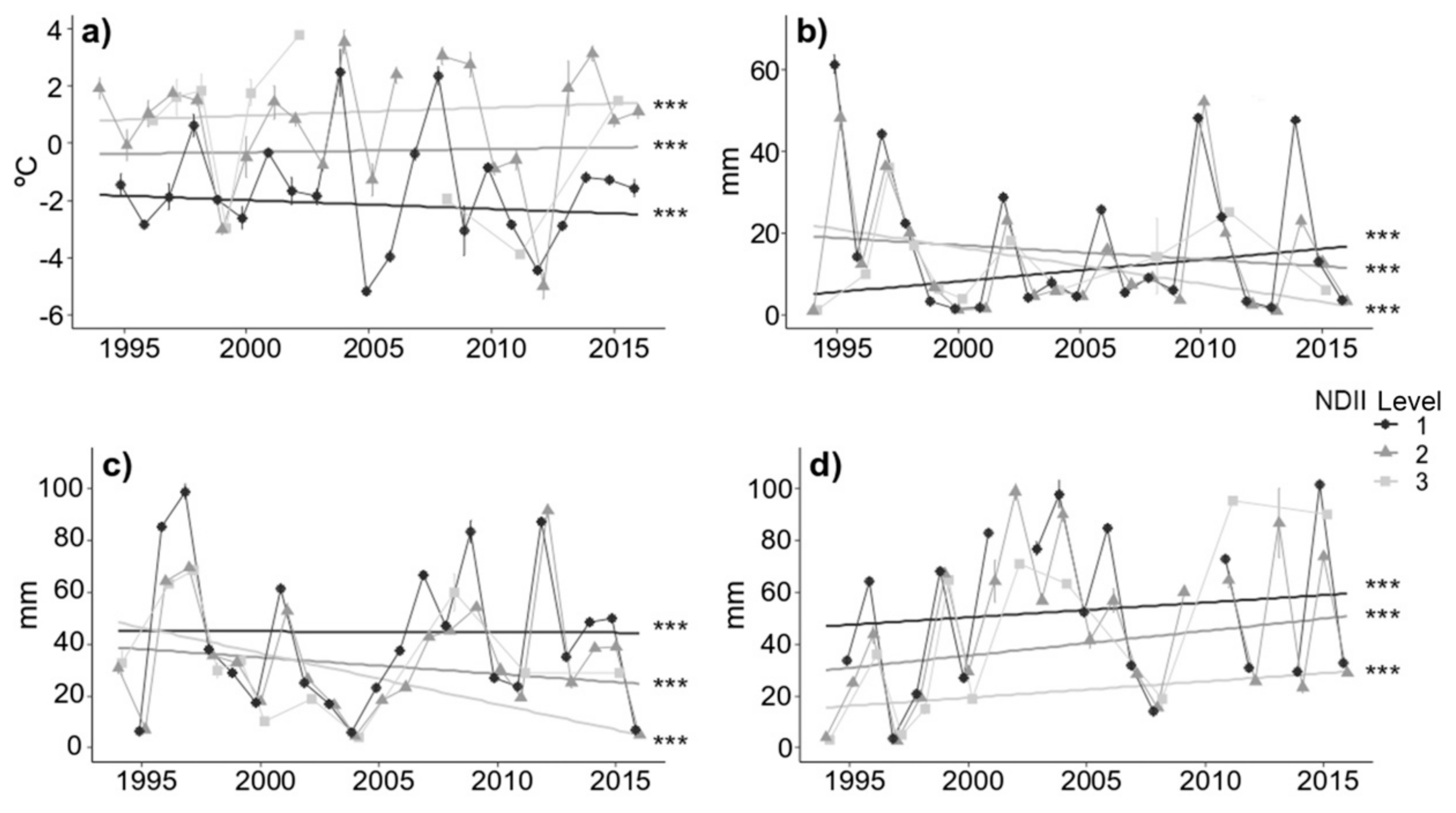
3.5. Temporal Trends
4. Discussion
4.1. Defoliation of Individual Species
4.2. Vegetation Indexes
4.3. Environmental Variables Associated with the PPM
4.4. Temporal Trends
4.5. Limitations and Uncertainties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pemán García, J.; Iriarte Goñi, I.; Lario Leza, F.J. La Restauración Forestal de España: 75 Años de una Ilusión; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente: Madrid, Spain, 2017; ISBN 978-84-491-1495-3.
- Raffa, K.F.; Aukema, B.; Bentz, B.J.; Carroll, A.; Erbilgin, N.; Herms, D.A.; Hicke, J.A.; Hofstetter, R.W.; Katovich, S.; Lindgren, B.S.; et al. A Literal Use of “Forest Health” Safeguards against Misuse and Misapplication. J. For. 2009, 107, 276–277. [Google Scholar]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol. Invasions 2010, 12, 389–405. [Google Scholar] [CrossRef]
- Netherer, S.; Schopf, A. Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manag. 2010, 259, 831–838. [Google Scholar] [CrossRef]
- Lakatos, F.; Mirtchev, S.; Mehmeti, A.; Shabanaj, H. Handbook of the Major Forest Pests in South East Europe; Food and Agriculture Organization of the Unitednations: Pristina, Serbia, 2014; p. 117. [Google Scholar]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangüesa-Barreda, G.; Camarero, J.J.; García-Martín, A.; Hernández, R.; de la Riva, J. Remote-sensing and tree-ring based characterization of forest defoliation and growth loss due to the Mediterranean pine processionary moth. For. Ecol. Manag. 2014, 320, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Kapeller, S.; Schroeder, H.; Schueler, S. Modelling the spatial population dynamics of the green oak leaf roller (Tortrix viridana) using density dependent competitive interactions: Effects of herbivore mortality and varying host-plant quality. Ecol. Model. 2011, 222, 1293–1302. [Google Scholar] [CrossRef]
- Li, S.; Daudin, J.J.; Piou, D.; Robinet, C.; Jactel, H. Periodicity and synchrony of pine processionary moth outbreaks in France. For. Ecol. Manag. 2015, 354, 309–317. [Google Scholar] [CrossRef]
- Dudley, T.L.; Bean, D.W. Tamarisk biocontrol, endangered species risk and resolution of conflict through riparian restoration. BioControl 2012, 57, 331–347. [Google Scholar] [CrossRef]
- Kautz, M.; Meddens, A.J.H.; Hall, R.J.; Arneth, A. Biotic disturbances in Northern Hemisphere forests—A synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr. 2017, 26, 533–552. [Google Scholar] [CrossRef]
- Meng, R.; Dennison, P.E.; Zhao, F.; Shendryk, I.; Rickert, A.; Hanavan, R.P.; Cook, B.D.; Serbin, S.P. Mapping canopy defoliation by herbivorous insects at the individual tree level using bi-temporal airborne imaging spectroscopy and LiDAR measurements. Remote Sens. Environ. 2018, 215, 170–183. [Google Scholar] [CrossRef]
- Kirilenko, A.P.; Sedjo, R.A. Climate change impacts on forestry. Proc. Natl. Acad. Sci. USA 2007, 104, 19697–19702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hódar, J.A.; Zamora, R.; Cayuela, L. Climate change and the incidence of a forest pest in Mediterranean ecosystems: Can the North Atlantic Oscillation be used as a predictor? Clim. Chang. 2012, 113, 699–711. [Google Scholar] [CrossRef]
- Roques, A.; Rousselet, J.; Avcı, M.; Avtzis, D.N.; Basso, A.; Battisti, A.; Jamaa, M.L.B.; Bensidi, A.; Berardi, L.; Berretima, W.; et al. Climate Warming and Past and Present Distribution of the Processionary Moths Thaumetopoea spp. in Europe, Asia Minor and North Africa. In Processionary Moths and Climate Change: An Update; Springer: Dordrecht, The Netherland, 2015; pp. 81–161. ISBN 978-94-017-9339-1. [Google Scholar]
- Montoya; del Pino, H.P. Informaciones Técnicas Departamento de Medio Ambiente del Gobierno de Aragón. 1998. Available online: http://www.caib.es/sites/sanitatforestal/f/23622 (accessed on 29 April 2019).
- Démolin, G. Bioecologia de la procesionaria del pino Thaumetopoea pityocampa Schiff. Incidencia de los factores climáticos. Bol. Serv. Plagas For. 1969, 23, 9–24. [Google Scholar]
- Consejeria de Medio Ambiente y Ordenación del Territorio Plan de Lucha Integrada Contra la Procesionaria del Pino. 2003, p. 43. Available online: http://www.juntadeandalucia.es/medioambiente/site/portalweb/menuitem.7e1cf46ddf59bb227a9ebe205510e1ca/?vgnextoid=1815e6f1563d6510VgnVCM1000001325e50aRCRD&vgnextchannel=d9cfe6f1563d6510VgnVCM1000001325e50aRCRD (accessed on 29 April 2019).
- De Beurs, K.M.; Townsend, P.A. Estimating the effect of gypsy moth defoliation using MODIS. Remote Sens. Environ. 2008, 112, 3983–3990. [Google Scholar]
- Pause, M.; Schweitzer, C.; Rosenthal, M.; Keuck, V.; Bumberger, J.; Dietrich, P.; Heurich, M.; Jung, A.; Lausch, A. In Situ/Remote Sensing Integration to Assess Forest Health—A Review. Remote Sens. 2016, 8, 471. [Google Scholar] [CrossRef]
- Rullan-Silva, C.D.; Olthoff, A.E.; de la Mata, J.A.D.; Pajares-Alonso, J.A. Remote Monitoring of Forest Insect Defoliation—A Review. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, X.; Zhang, N.; Hassan, M.A.; Zhao, L. Assessing the Defoliation of Pine Forests in a Long Time-Series and Spatiotemporal Prediction of the Defoliation Using Landsat Data. Remote Sens. 2018, 10, 360. [Google Scholar] [CrossRef]
- Belgiu, M.; Drăguţ, L. Random forest in remote sensing: A review of applications and future directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Leckie, D.G.; Carroll, A.L. Surveying mountain pine beetle damage of forests: A review of remote sensing opportunities. For. Ecol. Manag. 2006, 221, 27–41. [Google Scholar] [CrossRef]
- Goodwin, N.R.; Coops, N.C.; Wulder, M.A.; Gillanders, S.; Schroeder, T.A.; Nelson, T. Estimation of insect infestation dynamics using a temporal sequence of Landsat data. Remote Sens. Environ. 2008, 112, 3680–3689. [Google Scholar] [CrossRef]
- Wang, C.; Lu, Z.; Haithcoat, T.L. Using Landsat images to detect oak decline in the Mark Twain National Forest, Ozark Highlands. For. Ecol. Manag. 2007, 240, 70–78. [Google Scholar] [CrossRef]
- Navarro, R.M.; Blanco, P.; Fernández, P. Aplicación de las imágenes IRS-WiFS al análisis y evaluación de daños producidos por la procesionaria del pino (Thaumatopoea pytocampa Den. & Schiff.) en los pinares de Andalucía oriental. Mapping 2000, 66, 26–36. [Google Scholar]
- Robinet, C.; Baier, P.; Pennerstorfer, J.; Schopf, A.; Roques, A. Modelling the effects of climate change on the potential feeding activity of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in France. Glob. Ecol. Biogeogr. 2007, 16, 460–471. [Google Scholar]
- Hódar, J.A.; Castro, J.; Zamora, R. Pine processionary caterpillar Thaumetopoea pityocampa as a new threat for relict Mediterranean Scots pine forests under climatic warming. Biol. Conserv. 2003, 110, 123–129. [Google Scholar] [CrossRef]
- Toïgo, M.; Barraquand, F.; Barnagaud, J.Y.; Piou, D.; Jactel, H. Geographical variation in climatic drivers of the pine processionary moth population dynamics. For. Ecol. Manag. 2017, 404, 141–155. [Google Scholar] [CrossRef]
- Wulder, M.A.; White, J.C.; Coops, N.C.; Butson, C.R. Multi-temporal analysis of high spatial resolution imagery for disturbance monitoring. Remote Sens. Environ. 2008, 112, 2729–2740. [Google Scholar] [CrossRef]
- Woodcock, C.E.; Allen, R.; Anderson, M.; Belward, A.; Bindschadler, R.; Cohen, W.; Gao, F.; Goward, S.N.; Helder, D.; Helmer, E.; et al. Free access to Landsat imagery. Science 2008, 320, 1011. [Google Scholar] [CrossRef]
- Wang, P.; Wang, J.; Chen, Y.; Ni, G. Rapid processing of remote sensing images based on cloud computing. Future Gener. Comput. Syst. 2013, 29, 1963–1968. [Google Scholar] [CrossRef]
- Parks, S.; Holsinger, L.; Voss, M.; Loehman, R.; Robinson, N.; Parks, S.A.; Holsinger, L.M.; Voss, M.A.; Loehman, R.A.; Robinson, N.P. Mean Composite Fire Severity Metrics Computed with Google Earth Engine Offer Improved Accuracy and Expanded Mapping Potential. Remote Sens. 2018, 10, 879. [Google Scholar] [CrossRef]
- Lee, J.; Cardille, J.; Coe, M.; Lee, J.; Cardille, J.A.; Coe, M.T. BULC-U: Sharpening Resolution and Improving Accuracy of Land-Use/Land-Cover Classifications in Google Earth Engine. Remote Sens. 2018, 10, 1455. [Google Scholar] [CrossRef]
- Liu, C.C.; Shieh, M.C.; Ke, M.S.; Wang, K.H.; Liu, C.C.; Shieh, M.C.; Ke, M.S.; Wang, K.H. Flood Prevention and Emergency Response System Powered by Google Earth Engine. Remote Sens. 2018, 10, 1283. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Woodward, B.D.; Evangelista, P.H.; Vorster, A.G. Mapping Progression and Severity of a Southern Colorado Spruce Beetle Outbreak Using Calibrated Image Composites. Forests 2018, 9, 336. [Google Scholar] [CrossRef]
- Pastick, N.J.; Jorgenson, M.T.; Goetz, S.J.; Jones, B.M.; Wylie, B.K.; Minsley, B.J.; Genet, H.; Knight, J.F.; Swanson, D.K.; Jorgenson, J.C. Spatiotemporal remote sensing of ecosystem change and causation across Alaska. Glob. Chang. Biol. 2019, 25, 1171–1189. [Google Scholar] [CrossRef] [PubMed]
- USGS. Landsat 4–7 Surface Reflectance (LEDAPS) Product Guide; USGS: Sioux Falls, SD, USA, 2018.
- Zhu, Z.; Woodcock, C.E. Object-based cloud and cloud shadow detection in Landsat imagery. Remote Sens. Environ. 2012, 118, 83–94. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, S.; Woodcock, C.E. Improvement and expansion of the Fmask algorithm: Cloud, cloud shadow, and snow detection for Landsats 4–7, 8, and Sentinel 2 images. Remote Sens. Environ. 2015, 159, 269–277. [Google Scholar] [CrossRef]
- Rock, B.N.; Vogelmann, J.E.; Williams, D.L.; Vogelmann, A.F.; Hoshizaki, T. Remote Detection of Forest Damage. BioScience 1986, 36, 439–445. [Google Scholar] [CrossRef]
- Hunt, E.R.; Rock, B.N. Detection of changes in leaf water content using Near- and Middle-Infrared reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar]
- Sriwongsitanon, N.; Gao, H.; Savenije, H.; Maekan, E.; Saengsawang, S.; Thianpopirug, S. The Normalized Difference Infrared Index (NDII) as a proxy for soil moisture storage in hydrological modelling. Hydrol. Earth Syst. Sci. Discuss. 2015, 12, 8419–8457. [Google Scholar] [CrossRef]
- Xue, J.; Su, B. Significant Remote Sensing Vegetation Indices: A Review of Developments and Applications. Available online: https://www.hindawi.com/journals/js/2017/1353691/ (accessed on 15 September 2018).
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, J.F. Biometry: The Principles and Practice of Statistics in Biological Research. Available online: https://openlibrary.org/books/OL1087142M/Biometry (accessed on 29 April 2019).
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Univ. Ariz. 1983, 43, 69–678. [Google Scholar]
- De la Cruz, A.C.; Gil, P.M.; Fernández-Cancio, Á.; Minaya, M.; Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Grau, J.M. Defoliation triggered by climate induced effects in Spanish ICP Forests monitoring plots. For. Ecol. Manag. 2014, 331, 245–255. [Google Scholar] [CrossRef]
- Cristianini, N.; Shawe-Taylor, J. An Introduction to Support Vector Machines and Other Kernel-Based Learning Methods; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar] [CrossRef]
- Chirici, G.; Mura, M.; McInerney, D.; Py, N.; Tomppo, E.O.; Waser, L.T.; Travaglini, D.; McRoberts, R.E. A meta-analysis and review of the literature on the k-Nearest Neighbors technique for forestry applications that use remotely sensed data. Remote Sens. Environ. 2016, 176, 282–294. [Google Scholar] [CrossRef]
- Crookston, N.L.; Rehfeldt, G.E.; Ferguson, D.E.; Warwell, M. FVS and global Warming: A prospectus for future development. In Proceedings of the Third Forest Vegetation Simulator Conference, Fort Colline, CO, USA, 13–15 February 2017; pp. 7–16. [Google Scholar]
- Crookston, N.L.; Finley, A.O. yaImpute: An R Package for KNN Imputation. J. Stat. Softw. 2008, 23, 1–16. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Genuer, R.; Poggi, J.M.; Tuleau-Malot, C. Package “VSURF” Variable Selection Using Random Forests. R J. R Found. Stat. Comput. 2015, 7, 19–33. [Google Scholar] [CrossRef]
- Blanco, E.; Casado, M.A.; Costa, M.; Escribano, R.; García, M.; Gévora, M.; Gómez, A.; Gómez, F.; Moreno, J.C.; Morla, C.; et al. Los Bosques Ibéricos. Planeta: Barcelona, Spain, 1997. [Google Scholar]
- Kendall, B.E.; Ellner, S.P.; McCauley, E.; Wood, S.N.; Briggs, C.J.; Murdoch, W.W.; Turchin, P. Population Cycles in the Pine Looper Moth: Dynamical Tests of Mechanistic Hypotheses. Ecol. Monogr. 2005, 75, 259–276. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of Geographic Range in the Pine Processionary Moth Caused by Increased Winter Temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S.A. Comparison of time series tasseled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]
- White, J.C.; Coops, N.C.; Hilker, T.; Wulder, M.A.; Carroll, A.L. Detecting mountain pine beetle red attack damage with EO-1 Hyperion moisture indices. Int. J. Remote Sens. 2007, 28, 2111–2121. [Google Scholar] [CrossRef]
- Schultz, M.; Clevers, J.G.P.W.; Carter, S.; Verbesselt, J.; Avitabile, V.; Quang, H.V.; Herold, M. Performance of vegetation indices from Landsat time series in deforestation monitoring. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 318–327. [Google Scholar] [CrossRef]
- Zhu, Z.; Woodcock, C.E.; Olofsson, P. Continuous monitoring of forest disturbance using all available Landsat imagery. Remote Sens. Environ. 2012, 122, 75–91. [Google Scholar] [CrossRef]
- Jactel, H.; Barbaro, L.; Battisti, A.; Bosc, A.; Branco, M.; Brockerhoff, E.; Castagneyrol, B.; Dulaurent, A.M.; Hódar, J.A.; Jacquet, J.S.; et al. Insect—Tree Interactions in Thaumetopoea pityocampa. In Processionary Moths and Climate Change: An Update; Roques, A., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 265–310. ISBN 978-94-017-9340-7. [Google Scholar]
- White, T.C.R. The abundance of invertebrate herbivores in relation to the availability of nitrogen in stressed food plants. Oecologia 1984, 63, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Caldeira, M.C.; Burban, C.; Kerdelhué, C.; Branco, M. Shifted phenology in the pine processionary moth affects the outcome of tree-insect interaction. Bull. Entomol. Res. 2019, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shestakova, T.A.; Camarero, J.J.; Ferrio, J.P.; Knorre, A.A.; Gutiérrez, E.; Voltas, J. Increasing drought effects on five European pines modulate Δ13C-growth coupling along a Mediterranean altitudinal gradient. Funct. Ecol. 2017, 31, 1359–1370. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Tomas-Burguera, M.; Beguería, S.; Reig, F.; Latorre, B.; Peña-Gallardo, M.; Luna, M.Y.; Morata, A.; González-Hidalgo, J.C. A High Resolution Dataset of Drought Indices for Spain. Data 2017, 2, 22. [Google Scholar] [CrossRef]
- Otsu, K.; Pla, M.; Vayreda, J.; Brotons, L. Calibrating the Severity of Forest Defoliation by Pine Processionary Moth with Landsat and UAV Imagery. Sensors 2018, 18, 3278. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | R12 | MS |
|---|---|---|---|
| P. halepensis | 977 | 0.556 | 0.467 |
| P. nigra | 419 | 0.804 | 0.533 |
| P. pinaster | 480 | 0.467 | 0.268 |
| P. pinea | 227 | 0.532 | 0.234 |
| P. sylvestris | 200 | 0.612 | 0.268 |
| Pinus sp. | 3147 | 0.515 | 0.384 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Romero, J.; Navarro-Cerrillo, R.M.; Palacios-Rodriguez, G.; Acosta, C.; Mesas-Carrascosa, F.J. Improvement of Remote Sensing-Based Assessment of Defoliation of Pinus spp. Caused by Thaumetopoea pityocampa Denis and Schiffermüller and Related Environmental Drivers in Southeastern Spain. Remote Sens. 2019, 11, 1736. https://doi.org/10.3390/rs11141736
Pérez-Romero J, Navarro-Cerrillo RM, Palacios-Rodriguez G, Acosta C, Mesas-Carrascosa FJ. Improvement of Remote Sensing-Based Assessment of Defoliation of Pinus spp. Caused by Thaumetopoea pityocampa Denis and Schiffermüller and Related Environmental Drivers in Southeastern Spain. Remote Sensing. 2019; 11(14):1736. https://doi.org/10.3390/rs11141736
Chicago/Turabian StylePérez-Romero, Javier, Rafael María Navarro-Cerrillo, Guillermo Palacios-Rodriguez, Cristina Acosta, and Francisco Javier Mesas-Carrascosa. 2019. "Improvement of Remote Sensing-Based Assessment of Defoliation of Pinus spp. Caused by Thaumetopoea pityocampa Denis and Schiffermüller and Related Environmental Drivers in Southeastern Spain" Remote Sensing 11, no. 14: 1736. https://doi.org/10.3390/rs11141736
APA StylePérez-Romero, J., Navarro-Cerrillo, R. M., Palacios-Rodriguez, G., Acosta, C., & Mesas-Carrascosa, F. J. (2019). Improvement of Remote Sensing-Based Assessment of Defoliation of Pinus spp. Caused by Thaumetopoea pityocampa Denis and Schiffermüller and Related Environmental Drivers in Southeastern Spain. Remote Sensing, 11(14), 1736. https://doi.org/10.3390/rs11141736