Tree Species (Genera) Identification with GF-1 Time-Series in A Forested Landscape, Northeast China


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data from In-Situ Forest Inventory
2.3. Remotely Sensed Data and Pre-Processing
2.4. Temporal Texture Feature of Forest
2.5. Phenological Metrics Calculation
2.6. Forest Mapping on Landscape and Accuracy Assessment
3. Results
3.1. Forest Species Identification Based on NDVI Time-Series Data
3.2. Phenoloigcal Patterns and Forest Species Identification with Phenological Metrics
3.3. Forest Mapping with Temporal Texture Features
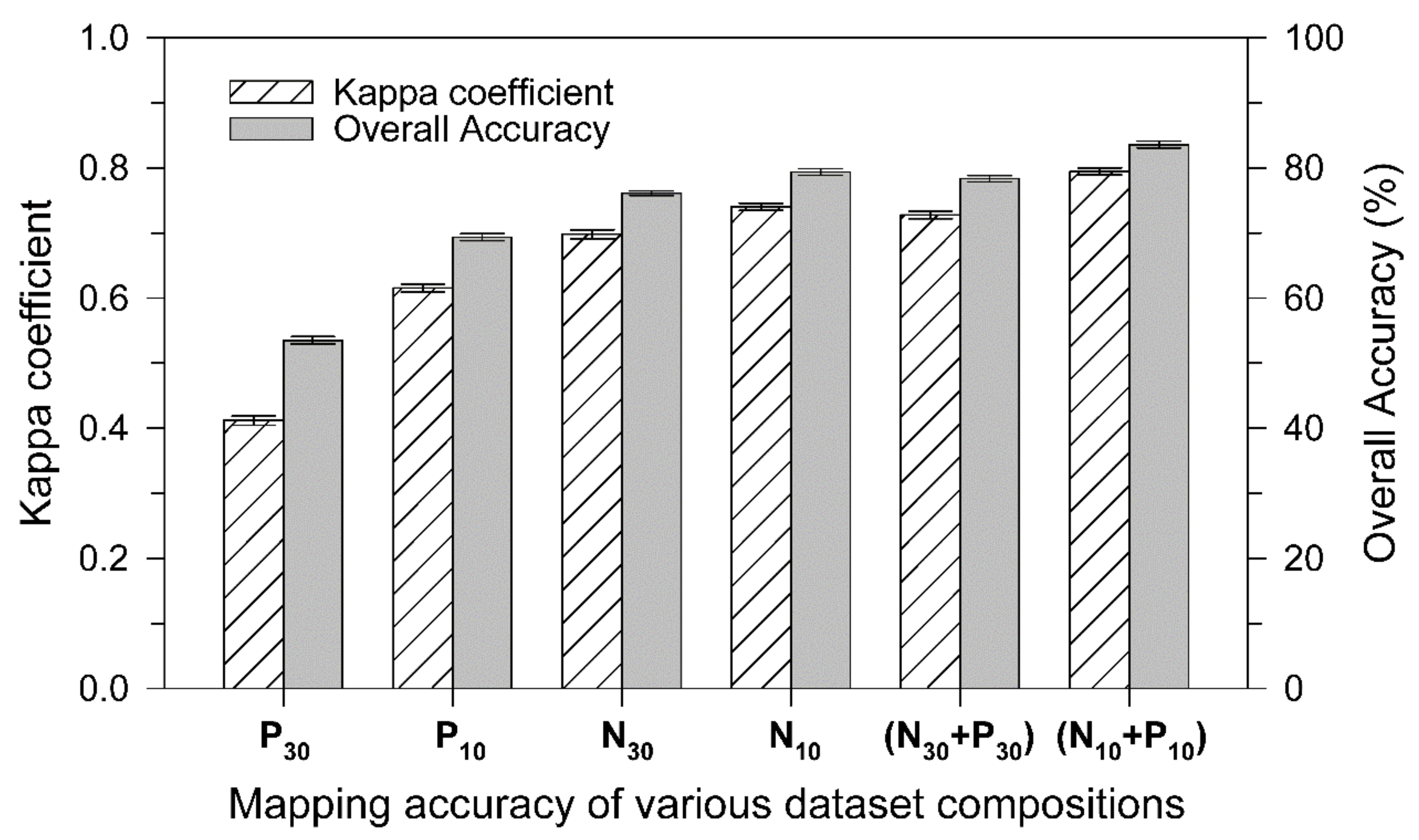
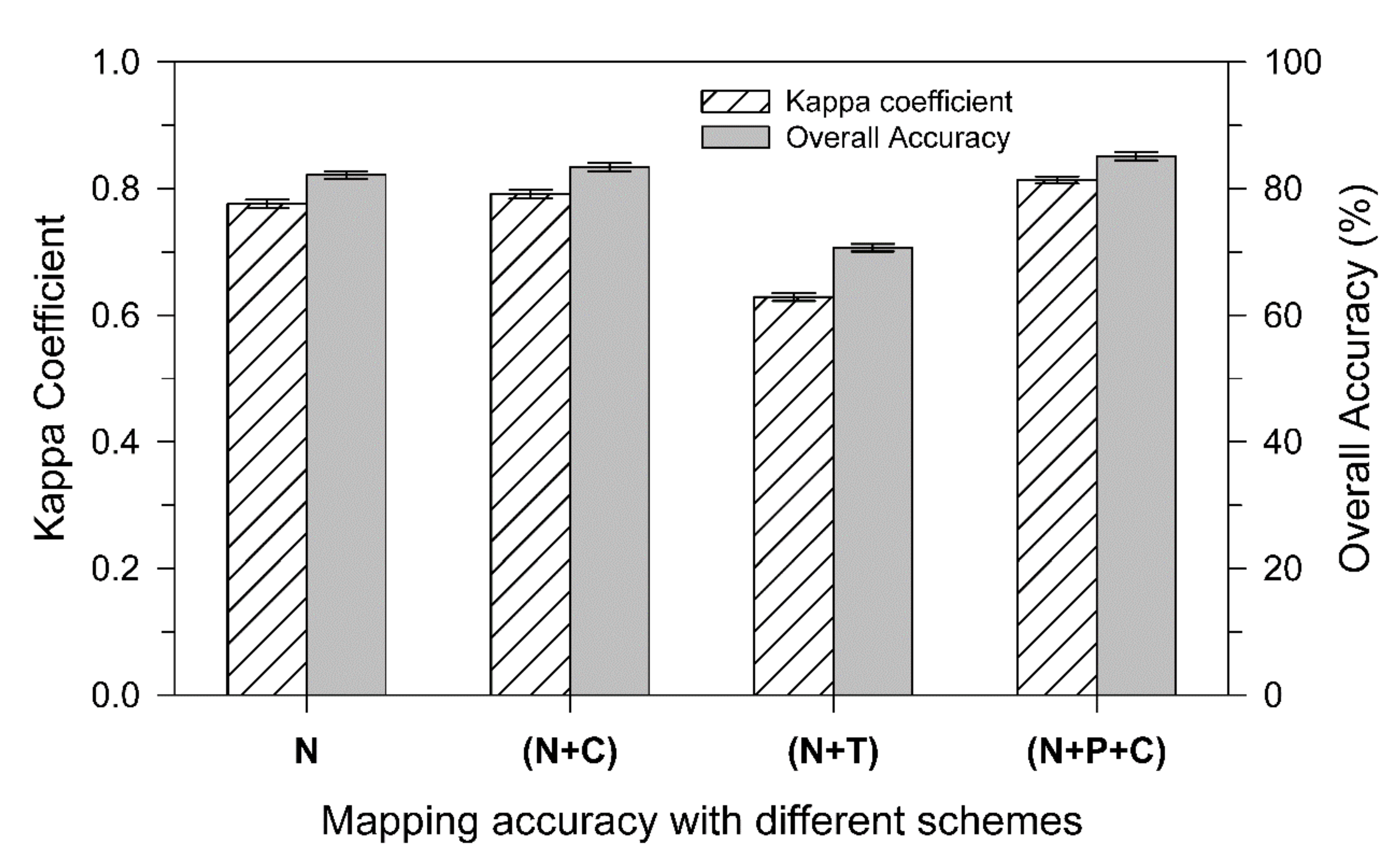
3.4. Forest Mapping on Landscape with Different Datasets
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sannier, C.; McRoberts, R.E.; Fichet, L. Suitability of global forest change data to report forest cover estimates at national level in Gabon. Remote Sens. Environ. 2016, 173, 326–338. [Google Scholar] [CrossRef]
- Westoby, M. Selective forces exerted by vertebrate herbivores on plants. Trends Ecol. Evol. 1989, 4, 115–117. [Google Scholar] [CrossRef]
- Heinzel, J.; Koch, B. Investigating multiple data sources for tree species classification in temperate forest and use for single tree delineation. Int. J. Appl. Earth Obs. 2012, 18, 101–110. [Google Scholar] [CrossRef]
- Saatchi, S.; Buermann, W.; Ter Steege, H.; Mori, S.; Smith, T.B. Modeling distribution of Amazonian tree species and diversity using remote sensing measurements. Remote Sens. Environ. 2008, 112, 2000–2017. [Google Scholar] [CrossRef]
- Tang, L.N.; Shao, G.F. Drone remote sensing for forestry research and practices. J. For. Res. 2015, 26, 791–797. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Clark, D.B. Hyperspectral discrimination of tropical rain forest tree species at leaf to crown scales. Remote Sens. Environ. 2005, 96, 375–398. [Google Scholar] [CrossRef]
- Wu, H.; Li, Z.L. Scale issues in remote sensing, a review on analysis, processing and modeling. Sensors 2009, 9, 1768–1793. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Straub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- Gessner, U.; Machwitz, M.; Conrad, C.; Dechab, S. Estimating the fractional cover of growth forms and bare surface in savannas. A multi-resolution approach based on regression tree ensembles. Remote Sens. Environ. 2013, 129, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Latorre, A.P.; Cabezudo, B. Use of monocharacteristic growth forms and phenological phases to describe and differentiate plant communities in Mediterranean-type ecosystems. Plant Ecol. 2002, 161, 231–249. [Google Scholar] [CrossRef]
- Vilhar, U.; Skudnik, M.; Simončič, P. Phenological phases of trees on the intensive forest monitoring plots in slovenia. Acta Silvae Et Ligni 2013, 100, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.; Dos Santos, J.A.; Alberton, B.; Morellato, L.P.C.; Ricardo, D.S.T. Plant species identification with phenological visual rhythms. In Proceedings of the IEEE International Conference on E-Science, Beijing, China, 22–25 October 2013. [Google Scholar] [CrossRef]
- Hill, R.A.; Wilson, A.K.; George, M.; Hinsley, S.A. Mapping tree species in temperate deciduous woodland using time-series multi-spectral data. Appl. Veg. Sci. 2010, 13, 86–99. [Google Scholar] [CrossRef]
- Jia, L.G.; Zhang, B.; Wei, H.D. The spectral characteristic variation analysis of three typical desert plants in growing season. Spectrosc. Spect. Anal. 2018, 38, 2881–2887. [Google Scholar] [CrossRef]
- Hoare, D.; Frost, P. Phenological description of natural vegetation in southern Africa using remotely-sensed vegetation data. Appl. Veg. Sci. 2004, 7, 19–28. [Google Scholar] [CrossRef]
- Tian, J.Q.; Zhu, X.L.; Wu, J.; Shen, M.G.; Chen, J. Coarse-resolution satellite images overestimate urbanization effects on vegetation spring phenology. Remote Sens. 2020, 12, 117. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.M.; Boles, S.; Liu, J.Y.; Zhuang, D.F.; Liu, M.L. Characterization of forest types in Northeastern China, using multi-temporal SPOT-4 VEGETATION sensor data. Remote Sens. Environ. 2002, 82, 335–348. [Google Scholar] [CrossRef]
- Kempeneers, P.; Sedano, F.; Seebach, L.M.; Strobl, P.; San-Miguel-Ayanz, J. Data fusion of different spatial resolution remote sensing images applied to forest-type mapping. IEEE T. Geocsi. Remote 2011, 49, 4977–4986. [Google Scholar] [CrossRef]
- Grabska, E.; Hostert, P.; Pflugmacher, D.; Ostapowicz, K. Forest stand species mapping using the Sentinel-2 time series. Remote Sens. 2019, 11, 1197. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Liu, D.S. Accurate mapping of forest types using dense seasonal Landsat time-series. ISPRS J. Photogramm. Remote Sens. 2014, 96, 1–11. [Google Scholar] [CrossRef]
- Puzzolo, V.; Denatale, F.; Gianne, F. Forest species discrimination in an Alpine mountain area using a fuzzy classification of multi-temporal SPOT (HRV) data. IEEE Int. Geosci. Remote Sens. Symp. 2003. [Google Scholar] [CrossRef]
- Persson, M.; Lindberg, E.; Reese, H. Tree species classification with multi-temporal Sentinel-2 data. Remote Sens. 2018, 10, 1794. [Google Scholar] [CrossRef] [Green Version]
- Wessel, M.; Brandmeier, M.; Tiede, D. Evaluation of different machine learning algorithms for scalable classification of tree types and tree species based on Sentinel-2 data. Remote Sens. 2018, 10, 1419. [Google Scholar] [CrossRef] [Green Version]
- Achard, F.; Estreguil, C. Forest classification of Southeast Asia using NOAA AVHRR data. Remote Sens. Environ. 1995, 54, 198–208. [Google Scholar] [CrossRef]
- Yui, X.F.; Zhuang, D.F.; Chen, H.; Hou, X.Y. Forest classification based on MODIS time series and vegetation phenology. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Anchorage, AK, USA, 20–24 September 2004; 2004. [Google Scholar] [CrossRef]
- Chang, B.H.; Wang, J.T.; Luo, Y.L.; Wang, Y.H.; Wang, Y.M. Cultivated land extraction based on GF-1/WFV remote sensing in Shenwu irrigation area of Hetao Irrigation District. Trans. Chin. Soc. Agric. Eng. 2017, 23, 188–195. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Wang, L.; Zhao, X.N.; Qu, X.N.; Xu, X.; Wang, R. Extraction of crops in oasis based on GF-1/WFV time series. Arid Zone Res. 2019, 36, 781–789. [Google Scholar] [CrossRef]
- Roth, K.L.; Roberts, D.A.; Dennison, P.E.; Peterson, S.H.; Alonzo, M. The impact of spatial resolution on the classification of plant species and functional types within imaging spectrometer data. Remote Sens. Environ. 2015, 171, 45–57. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.; Richardson, A.D.; Jensen, O.; O’Keefe, J.; Zhang, G.; Nemani, R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Change Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Ganguly, S.; Friedl, M.A.; Tan, B.; Zhang, X.Y.; Verma, M. Land surface phenology from MODIS: Characterization of the collection 5 global land cover dynamics product. Remote Sens. Environ. 2010, 114, 1805–1816. [Google Scholar] [CrossRef] [Green Version]
- Michez, A.; Piegay, H.; Jonathan, L.; Claessens, H.; Lejeune, P. Mapping of riparian invasive species with supervised classification of Unmanned Aerial System (UAS) imagery. Int. J. Appl. Earth Obs. 2015, 44, 88–94. [Google Scholar] [CrossRef]
- Kong, J.X.; Zhang, Z.C.; Zhang, J. Classification and identification of plant species based on multi-source remote sensing data: Research progress and prospect. Biodiv. Sci. 2019, 27, 796–812. [Google Scholar] [CrossRef]
- Liu, W.J.; Zeng, Y.N.; Zhang, M. Mapping rice paddy distribution by using time series HJ blend data and phenological parameters. J. Remote Sens. 2018, 22, 381–391. [Google Scholar] [CrossRef]
- Ota, T.; Mizoue, N.; Yoshida, S. Influence of using texture information in remote sensed data on the accuracy of forest type classification at different levels of spatial resolution. J. For. Res. 2011, 16, 432–437. [Google Scholar] [CrossRef]
- Li, M.Y.; Xing, Y.Q.; Liu, M.S.; Wang, Z.; Yao, S.T.; Zeng, X.J.; Xie, J. Identification of forest type with Landsat-8 image based on SVM. J. Cent. S. Univ. For. Technol. 2017, 37, 52–58. [Google Scholar] [CrossRef]
- Xu, K.J.; Tian, Q.J.; Yue, J.B.; Tang, S.F. Forest tree species identification and its response to spatial scale based on multispectral and multi-resolution remotely sensed data. Chin. J. Appl. Ecol. 2018, 29, 3986–3994. [Google Scholar] [CrossRef]
- Ke, S.F.; Li, B.; Yang, Y.H.; Ma, C.G.; Jia, Y.S. The evaluation of carbon footprint from the operation of forest farm and carbon storage by forest resources based on the Wangyedian forest farm in Chifeng of Inner Mongolia. For. Econ. 2013, 35, 93–101. [Google Scholar] [CrossRef]
- Gong, P.; Pu, R.L.; Yu, B. Conifer species recognition with seasonal hyperspectral data. J. Remote Sens. 1998, 2, 211–217. [Google Scholar] [CrossRef]
- Zheng, B.; Myint, S.W.; Thenkabail, P.S.; Aggarwal, R.M. A support vector machine to identify irrigated crop types using time-series Landsat NDVI data. Int. J. Appl. Earth Obs. 2015, 34, 103–112. [Google Scholar] [CrossRef]
- Estoque, R.C.; Murayama, Y.; Akiyama, C.M. Pixel-based and object-based classifications using high- and medium-spatial-resolution imageries in the urban and suburban landscapes. Geocarto Int. 2015, 30, 1113–1129. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Tian, Q.; Yang, Y.; Yue, J.; Tang, S. How up-scaling of remote-sensing images affects land-cover classification by comparison with multiscale satellite images. Int. J. Remote Sens. 2019, 40, 2784–2810. [Google Scholar] [CrossRef]
- Yang, Y.; Tao, B.; Ren, W.; Zourarakis, D.P.; Masri, B.E.; Sun, Z.; Tian, Q. An improved approach considering intraclass variability for mapping winter wheat using multitemporal MODIS EVI images. Remote Sens. 2019, 11, 1191. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.J.; Zeng, H.D.; Zhu, X.B.; Tian, Q.J. Evaluation of five commonly used atmospheric correction algorithms for multi-temporal aboveground forest carbon storage estimation. Spectrosc. Spect. Anal. 2017, 37, 3493–3498. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Birky, A.K. NDVI and a simple model of deciduous forest seasonal dynamics. Ecol. Model. 2001, 143, 43–58. [Google Scholar] [CrossRef]
- Hird, J.N.; Mcdermid, G.J. Noise reduction of NDVI time series: An empirical comparison of selected techniques. Remote Sens. Environ. 2009, 113, 248–258. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Wang, Y.F.; Xue, Z.H.; Chen, J.; Chen, G.Z. Spatio-temporal analysis of phenology in Yangtze river delta based on MODIS NDVI time series from 2001 to 2015. Front. Earth Sci-Prc. 2019, 13, 92–110. [Google Scholar] [CrossRef]
- Li, R.; Zhang, X.; Liu, B.; Zhang, B. Review on methods of remote sensing time-series data reconstruction. J. Remote Sens. 2009, 13, 335–341. [Google Scholar] [CrossRef]
- Zhao, G.; Maclean, A.L. A comparison of canonical discriminant analysis and principal component analysis for spectral transformation. Photogramm. Eng. Rem. Sens. 2000, 66, 841–847. [Google Scholar] [CrossRef]
- Alessandri, A.; Navarra, A. On the coupling between vegetation and rainfall inter-annual anomalies: Possible contributions to seasonal rainfall predictability over land areas. Geophys. Res. Lett. 2008, 35, L02718. [Google Scholar] [CrossRef]
- Chang, C.T.; Wang, H.C.; Huang, C. Retrieving multi-scale climatic variations from high dimensional time-series MODIS green vegetation cover in a tropical/subtropical mountains island. J. Mt. Sci. 2014, 11, 407–420. [Google Scholar] [CrossRef]
- Coburn, C.A.; Roberts, A.C.B. A multiscale texture analysis procedure for improved forest stand classification. Int. J. Remote Sens. 2004, 25, 4287–4308. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Datta, A.; Chakladar, N.D.; Pal, S.K.; Mukhopadhyay, S.; Sen, R. Detection of tool condition from the turned surface images using an accurate grey level co-occurrence technique. Precis. Eng. 2012, 36, 458–466. [Google Scholar] [CrossRef]
- Franklin, S.E.; Hall, R.J.; Moskal, L.M.; Maudie, A.J.; Lavigne, M.B. Incorporating texture into classification of forest species composition from airborne multispectral images. Int. J. Remote Sens. 2000, 21, 61–79. [Google Scholar] [CrossRef]
- Clausi, D.A. An analysis of co-occurrence texture statistics as a function of grey level quantization. Can. J. Remote Sens. 2002, 28, 45–62. [Google Scholar] [CrossRef]
- Gallardo-Cruz, J.A.; Meave, J.A.; González, E.J.; Lebrija-Trejos, E.E.; Romero-Romero, M.A.; Pérez-García, E.A.; Gallardo-Cruz, R.; Hernández-Stefanoni, J.L.; Martorell, C. Predicting tropical dry forest successional attributes from space, is the key hidden in image texture. PLoS ONE 2012, 7, e30506. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.X.; Gertner, G.; Anderson, A.B. Up-scaling methods based on variability–weighting and simulation for inferring spatial information across scales. Int. J. Remote Sens. 2004, 25, 4961–4979. [Google Scholar] [CrossRef]
- Kayitakire, F.; Hamel, C.; Defourny, P. Retrieving forest structure variables based on image texture analysis and IKONOS-2 imagery. Remote Sens. Environ. 2006, 102, 390–401. [Google Scholar] [CrossRef]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I. Textural features for image classification. IEEE Trans. Syst. Man. Cybern. 1973, 3, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Niel, T.G.V.; Mcvicar, T.R.; Datt, B. On the relationship between training sample size and data dimensionality: Monte Carlo analysis of broadband multi-temporal classification. Remote Sens. Environ. 2005, 98, 468–480. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, Z.J.; Yan, C.Y. A experimental study on imaging spectrometer data feature selection and wheat type identification. J. Remote Sens. 2006, 10, 249–255. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O. Spectral discrimination of papyrus vegetation (Cyperus papyrus L.) in swamp wetlands using field spectrometry. ISPRS J. Photogramm. Remote Sens. 2009, 64, 612–620. [Google Scholar] [CrossRef]
- Tottrup, C.; Rasmussen, M.S.; Eklundh, L. Mapping fractional forest cover across the highlands of mainland Southeast Asia using MODIS data and regression tree modelling. Int. J. Remote Sens. 2007, 28, 23–46. [Google Scholar] [CrossRef]
- Seaquist, J.W.; Hickler, T.; Eklundh, L. Disentangling the effects of climate and people on Sahel vegetation dynamics. Biogeosciences 2009, 6, 469–477. [Google Scholar] [CrossRef]
- Jönnson, P.; Eklundh, L. TIMESAT-a program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.T.; Wang, H.C.; Huang, C. Impact of vegetation onset time on the net primary productivity in a mountainous island in Pacific Asia. Environ. Res. Lett. 2013, 8, 045030. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.T.; Guan, H.D.; Shen, M.G.; Liang, W.; Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob. Change Biol. 2014, 21, 1–14. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jönsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Jiao, F.S.; Liu, H.Y.; Xu, X.J.; Gong, H.B.; Lin, Z.S. Trend evolution of vegetation phenology in China during the period of 1981–2016. Remote Sens. 2020, 12, 572. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.H.; Feng, X.Z.; Du, J.K.; Lin, G.F. SPIN-2 panchromatic and SPOT-4 multi-spectral image fusion based on support vector machine. J. Remote Sens. 2003, 7, 407–411. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Fusion of hyperspectral and LIDAR remote sensing data for classification of complex forest areas. IEEE Trans. Geosci. Remote. 2008, 46, 1416–1427. [Google Scholar] [CrossRef] [Green Version]
- Bruzzone, L.; Persello, C. A novel context-sensitive semi-supervised SVM classifier robust to mislabeled training samples. IEEE Trans. Geosci. Remote. 2009, 47, 2142–2154. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Lunetta, R.S. Comparison of support vector machine, neural network, and CART algorithms for the land-cover classification using limited training data points. ISPRS J. Photogramm. 2012, 70, 78–87. [Google Scholar] [CrossRef]
- Kumar, P.; Gupta, D.K.; Mishra, V.N.; Prasad, R. Comparison of support vector machine, artificial neural network, and spectral angle mapper algorithms for crop classification using LISS IV data. Int. J. Remote Sens. 2015, 36, 1604–1617. [Google Scholar] [CrossRef]
- Raczko, E.; Zagajewski, B. Comparison of support vector machine, random forest and neural network classifiers for tree species classification on airborne hyperspectral APEX images. Eur. J. Remote Sens. 2017, 50, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Thanh-Noi, P.; Kappas, M. Comparison of random forest, k-nearest neighbor, and support vector machine classifiers for land cover classification using Sentinel-2 imagery. Sensors 2018, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Kuemmerle, T.; Radeloff, V.C.; Perzanowski, K.; Hostert, P. Cross-border comparison of land cover and landscape pattern in eastern Europe using a hybrid classification technique. Remote Sens. Environ. 2006, 103, 449–464. [Google Scholar] [CrossRef]
- Sothe, C.; de Almeida, C.M.; Liesenberg, V.; Schimalski, M.B. Evaluating Sentinel-2 and Landsat-8 data to map sucessional forest stages in a subtropical forest in southern Brazil. Remote Sens. 2017, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.L.F.; van de Rwel, F.J.M. Accuracy assessment of satellite derived land-gover data: A review. Photogramm. Eng. Rem. S. 1994, 60, 419–426. [Google Scholar] [CrossRef]
- Madonsela, S.; Cho, M.A.; Mathieu, R.; Mutanga, O.; Ramoelo, A.; Kaszta, Ż.; van de Kerchove, R.; Wolff, E. Multi-phenology WorldView-2 imagery improves remote sensing of savannah tree species. Int. J. Appl. Earth Obs. 2017, 58, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.L.; Chen, Y.L.; Lu, D.S.; Li, G.Y.; Chen, R.X. Classification of land cover, forest, and tree species classes with ZiYuan-3 multispectral and stereo data. Remote Sens. 2019, 11, 164. [Google Scholar] [CrossRef] [Green Version]
- Mannel, S.; Price, M. Comparing classification results of multi-seasonal TM against AVIRIS imagery – seasonality more important than number of bands. Photogramm. Fernerkun. 2012, 2012, 603–612. [Google Scholar] [CrossRef]
- Immitzer, M.; Vuolo, F.; Atzberger, C. First experience with Sentinel-2 data for crop and tree species classifications in Central Europe. Remote Sens. 2016, 8, 166. [Google Scholar] [CrossRef]
- Vuolo, F.; Neuwirth, M.; Immitzer, M.; Atzberger, C.; Ng, W.T. How much does multi-temporal Sentinel-2 data improve crop type classification? Int. J. Appl. Earth Obs. 2018, 72, 122–130. [Google Scholar] [CrossRef]
- Zhang, J.K.; Rivard, B.; Sánchez-Azofeifa, A.; Castro-Esau, K. Intra- and inter- class spectral variability of tropical tree species at La Selva, Costa Rica, Implications for species identification using HYDICE imagery. Remote Sens. Environ. 2006, 105, 129–141. [Google Scholar] [CrossRef]
- Chakravorty, R.; Gauri, S.K.; Chakravorty, S. A modified principal component analysis-based utility theory approach for optimization of correlated responses of EDM process. Int. J. Technol. Manag. 2012, 4, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Moulin, S.; Kergoat, L.; Viovy, N.; Dedieu, G. Global-scale assessment of vegetation phenology using NOAA/AVHRR satellite measurements. J. Clim. 1997, 10, 1154–1170. [Google Scholar] [CrossRef]
- Ahl, D.E.; Gower, S.T.; Burrows, S.N.; Shabanov, N.V.; Myneni, R.B.; Knyazikhin, Y. Monitoring spring canopy phenology of a deciduous broadleaf forest using MODIS. Remote Sens. Environ. 2006, 104, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B. Sensitivity of vegetation phenology detection to the temporal resolution of satellite data. Int. J. Remote Sens. 2009, 30, 2061–2074. [Google Scholar] [CrossRef]
- Luo, L.L.; Wang, X.X.; Nong, J.H.; Liang, Z.J.; Tang, G.Y. Remote sensing forest classification with texture based on ICA and SVM. Comput. Eng. Appl. 2012, 48, 227–229. [Google Scholar] [CrossRef]
- Pu, R.L.; Landry, S. A comparative analysis of high spatial resolution IKONOS and WorldView-2 imagery for mapping urban tree species. Remote Sens. Environ. 2012, 124, 516–533. [Google Scholar] [CrossRef]
- Kushwaha, S.P.S.; Kuntz, S.; Oesten, G. Applications of image texture in forest classification. Int. J. Remote Sens. 1994, 15, 2273–2284. [Google Scholar] [CrossRef]
- Wulder, M.A.; LeDrew, E.F.; Franklin, S.E.; Lavigne, M.B. Aerial image texture information in the estimation of northern deciduous and mixed wood forest leaf area index (LAI). Remote Sens. Environ. 1998, 64, 64–76. [Google Scholar] [CrossRef]
- Pietsch, S.A.; Hasenauer, H.; Thornton, P.E. BGC-model parameters for tree species growing in central European forests. For. Ecol. Manag. 2005, 211, 264–295. [Google Scholar] [CrossRef]
- Liu, J.; Liu, S.; Loveland, T.R.; Tieszen, L.L. Integrating remotely sensed land cover observations and a biogeochemical model for estimating forest ecosystem carbon dynamics. Ecol. Model. 2008, 219, 361–372. [Google Scholar] [CrossRef]
- Keller, A.B.; Reed, S.C.; Townsend, A.R.; Cleveland, C.C. Effects of canopy tree species on belowground biogeochemistry in a lowland wet tropical forest. Soil Biol. Biochem. 2013, 58, 61–69. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Acquisition Date | Number | Acquisition Date | Number | Acquisition Date |
|---|---|---|---|---|---|
| 1 | 20170110 | 13 | 20170507 | 25 | 20170921 |
| 2 | 20170118 | 14 | 20170520 | 26 | 20170930 |
| 3 | 20170202 | 15 | 20170529 | 27 | 20171014 |
| 4 | 20170211 | 16 | 20170610 | 28 | 20161019 |
| 5 | 20170219 | 17 | 20170626 | 29 | 20161023 |
| 6 | 20170228 | 18 | 20170708 | 30 | 20171103 |
| 7 | 20170308 | 19 | 20170717 | 31 | 20171116 |
| 8 | 20170312 | 20 | 20170810 | 32 | 20171124 |
| 9 | 20170325 | 21 | 20170822 | 33 | 20171202 |
| 10 | 20170409 | 22 | 20170830 | 34 | 20171215 |
| 11 | 20160422 | 23 | 20170907 | 35 | 20171223 |
| 12 | 20170430 | 24 | 20170912 | 36 | 20171231 |
| Pair-Comparisons | CON | ENT | SM | COR |
|---|---|---|---|---|
| Pt - Lg | 1.9895 | 1.6694 | 1.7974 | 1.4946 |
| Pt - Qm | 2.0000 | 1.9698 | 1.9848 | 1.9299 |
| Pt - Bp | 1.9995 | 1.8105 | 1.9046 | 1.7102 |
| Pt - Pd | 2.0000 | 1.9968 | 1.9998 | 1.9707 |
| Lg - Qm | 2.0000 | 1.9489 | 1.9737 | 1.9305 |
| Lg - Bp | 1.9989 | 1.5586 | 1.6454 | 1.4655 |
| Lg - Pd | 2.0000 | 1.9860 | 1.9997 | 1.9529 |
| Qm - Bp | 1.9991 | 1.9622 | 1.9830 | 1.9504 |
| Qm - Pd | 2.0000 | 1.9962 | 1.9999 | 1.9875 |
| Bp - Pd | 2.0000 | 1.9974 | 2.0000 | 1.9778 |
| Schemes | N | NPCA | NPCA+P | NPCA+P+C | ||||
|---|---|---|---|---|---|---|---|---|
| Accuracy | PA | UA | PA | UA | PA | UA | PA | UA |
| Pt | 99.18 | 92.02 | 96.72 | 93.65 | 100 | 91.67 | 100 | 92.4 |
| Lg | 77.95 | 76.21 | 78.87 | 77.70 | 78.70 | 79.74 | 80.43 | 81.86 |
| Qm | 80.49 | 83.76 | 85.15 | 85.57 | 84.24 | 89.01 | 84.98 | 89.23 |
| Bp | 78.88 | 70.66 | 77.83 | 75.21 | 82.46 | 77.37 | 81.94 | 80.17 |
| Pd | 52.91 | 71.09 | 70.05 | 77.98 | 70.77 | 79.31 | 77.32 | 81.52 |
| Overall accuracy | 79.40 | 82.18 | 83.62 | 85.13 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, K.; Tian, Q.; Zhang, Z.; Yue, J.; Chang, C.-T. Tree Species (Genera) Identification with GF-1 Time-Series in A Forested Landscape, Northeast China. Remote Sens. 2020, 12, 1554. https://doi.org/10.3390/rs12101554
Xu K, Tian Q, Zhang Z, Yue J, Chang C-T. Tree Species (Genera) Identification with GF-1 Time-Series in A Forested Landscape, Northeast China. Remote Sensing. 2020; 12(10):1554. https://doi.org/10.3390/rs12101554
Chicago/Turabian StyleXu, Kaijian, Qingjiu Tian, Zhaoying Zhang, Jibo Yue, and Chung-Te Chang. 2020. "Tree Species (Genera) Identification with GF-1 Time-Series in A Forested Landscape, Northeast China" Remote Sensing 12, no. 10: 1554. https://doi.org/10.3390/rs12101554