Geographic Object-Based Image Analysis Framework for Mapping Vegetation Physiognomic Types at Fine Scales in Neotropical Savannas

 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.1.1. Study Site 1: Taquara Watershed
2.1.2. Study Site 2: Western Bahia
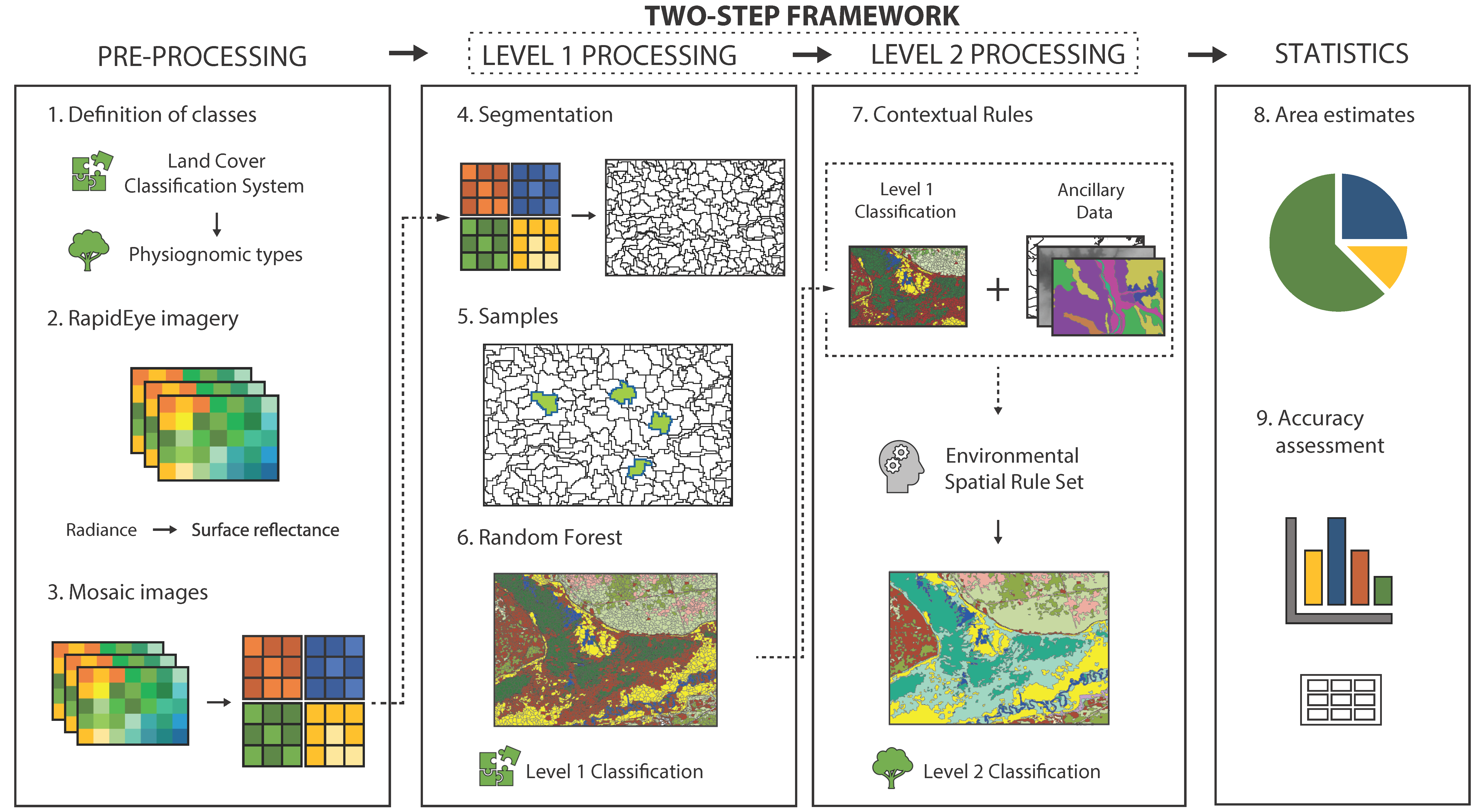
2.2. Methods Overview
2.2.1. Definition of Classes
2.2.2. Imagery Acquisition and Pre-Processing
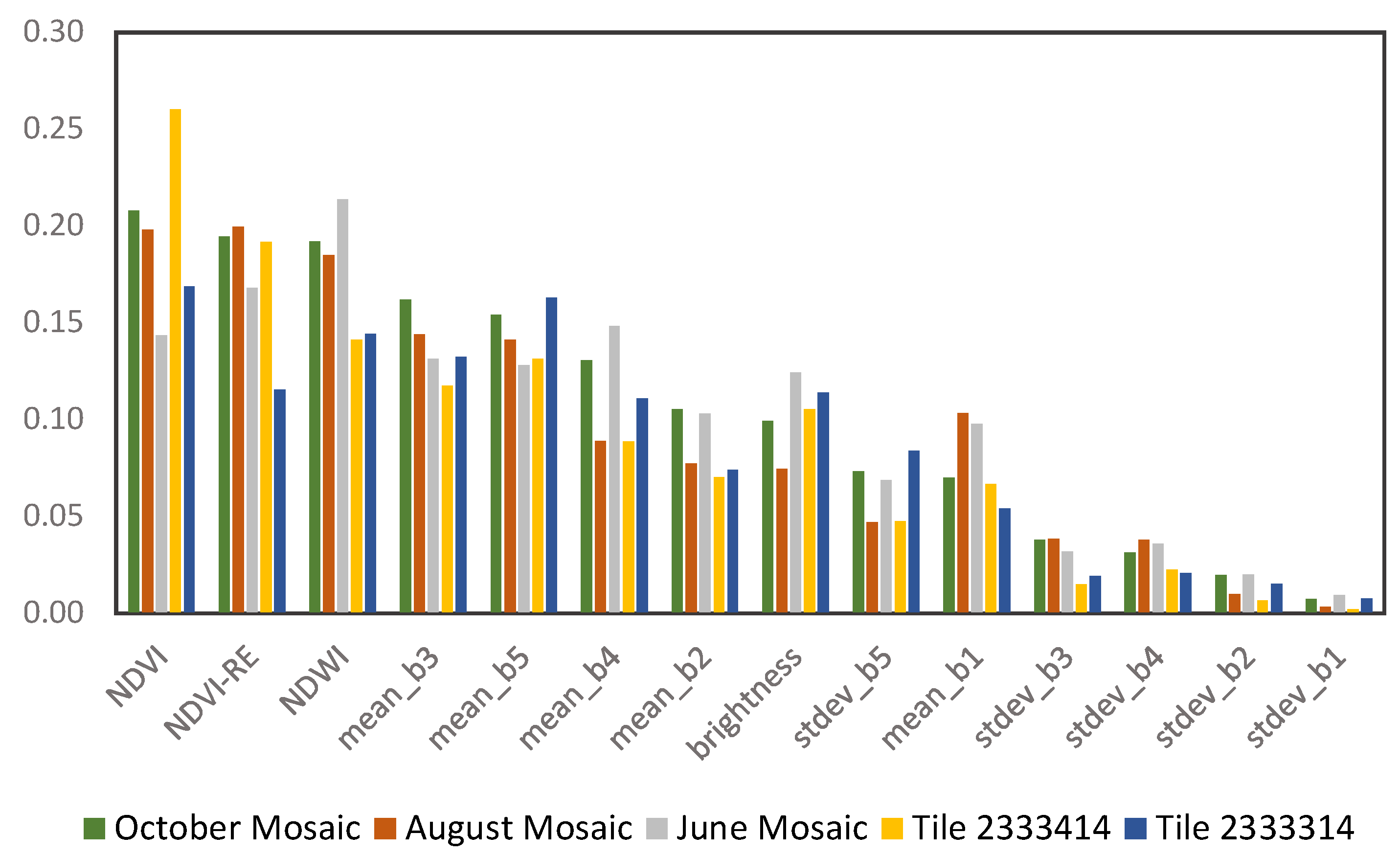
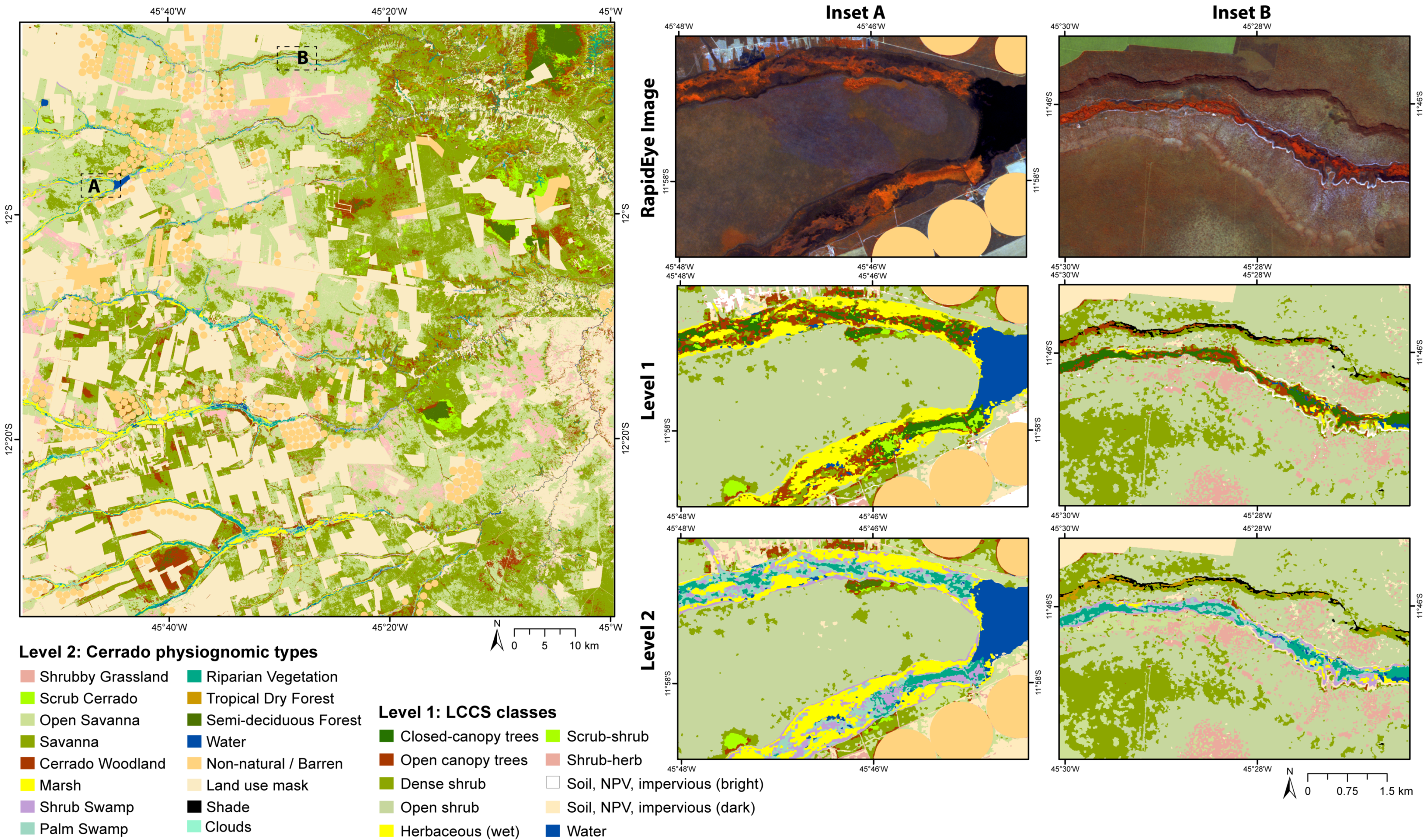
2.2.3. Level 1 Classification: Major Land Cover Types
2.2.4. Level 2 Classification: Physiognomic Types
2.2.5. Accuracy Assessment Procedures
3. Results
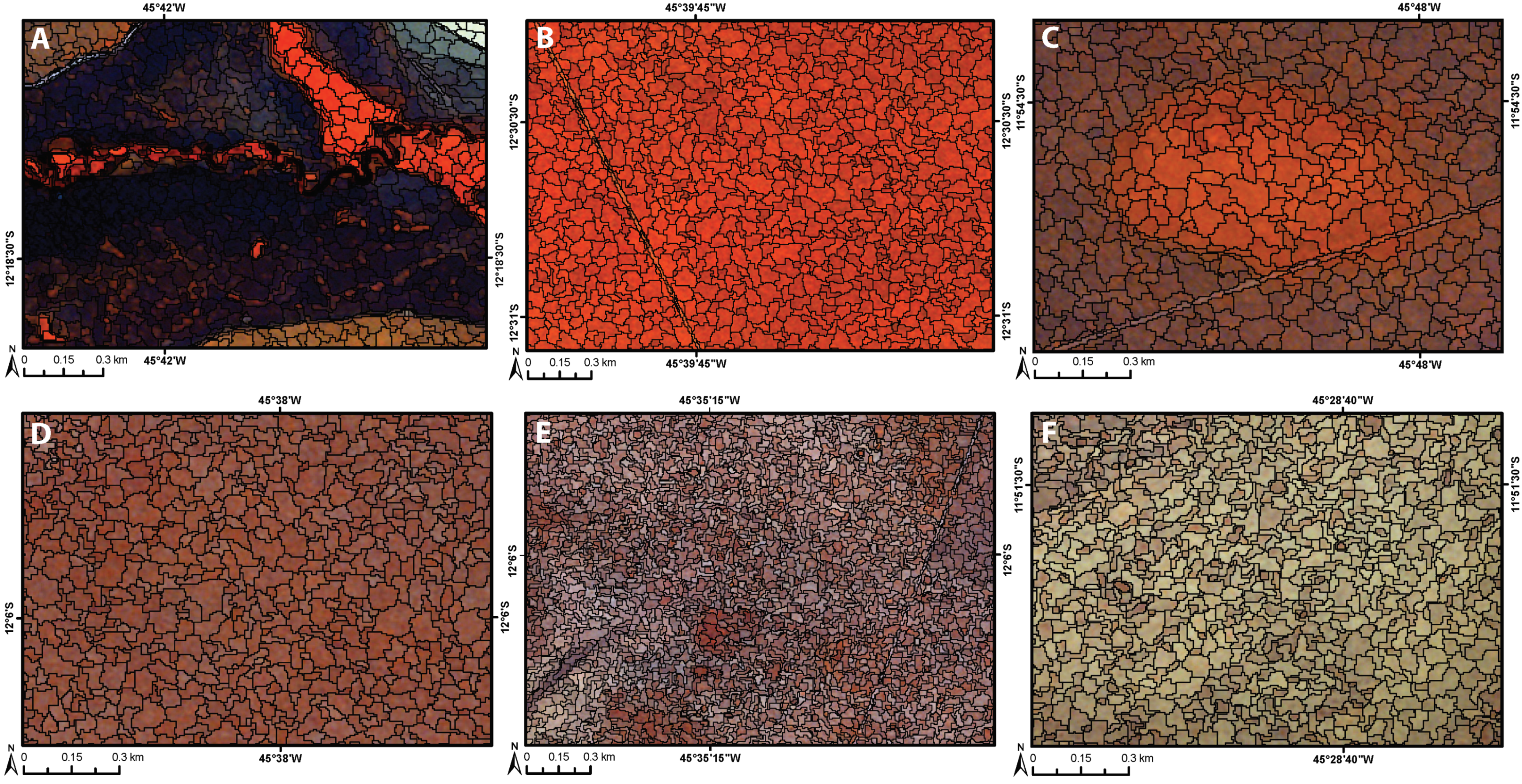
3.1. Segmentation Results
3.2. Accuracy Assessment
3.3. Landscape Composition: Area Assessments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bond, W.J.; Parr, C.L. Beyond the forest edge: Ecology, diversity and conservation of the grassy biomes. Biol. Conserv. 2010, 143, 2395–2404. [Google Scholar] [CrossRef]
- Eiten, G. The cerrado vegetation of Brazil. Bot. Rev. 1972, 38, 201–341. [Google Scholar] [CrossRef]
- Huntley, B.J.; Walker, B.H. Ecology of Tropical Savannas; Springer: Berlin/Heidelberg, Germany, 1982; Volume 42, ISBN 3-540-11885-3. [Google Scholar]
- Parr, C.L.; Lehmann, C.E.R.; Bond, W.J.; Hoffmann, W.A.; Andersen, A.N. Tropical grassy biomes: Misunderstood, neglected, and under threat. Trends Ecol. Evol. 2014, 29, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Bastin, J.-F.; Berrahmouni, N.; Grainger, A.; Maniatis, D.; Mollicone, D.; Moore, R.; Patriarca, C.; Picard, N.; Sparrow, B.; Abraham, E.M.; et al. The extent of forest in dryland biomes. Science 2017, 356, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Symeonakis, E.; Higginbottom, T.P.; Petroulaki, K.; Rabe, A. Optimisation of Savannah Land Cover Characterisation with Optical and SAR Data. Remote Sens. 2018, 10, 499. [Google Scholar] [CrossRef] [Green Version]
- Cord, A.; Conrad, C.; Schmidt, M.; Dech, S. Standardized FAO-LCCS land cover mapping in heterogeneous tree savannas of West Africa. J. Arid Environ. 2010, 74, 1083–1091. [Google Scholar] [CrossRef]
- Herold, M.; Mayaux, P.; Woodcock, C.E.; Baccini, A.; Schmullius, C. Some challenges in global land cover mapping: An assessment of agreement and accuracy in existing 1 km datasets. Remote Sens. Environ. 2008, 112, 2538–2556. [Google Scholar] [CrossRef]
- Scholes, R.J.; Archer, S.R. Tree-Grass Interactions in Savannas. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Eiten, G. Brazilian “Savannas”. In Ecology of Tropical Savannas; Springer: Berlin/Heidelberg, Germany, 1982; Volume 42, pp. 25–47. ISBN 3-540-11885-3. [Google Scholar]
- Mendonça, R.; Felfili, J.; Walter, B.; Silva-Junior, M.; Rezende, A.; Filgueiras, T.; Nogueira, P.; Fagg, C. Vascular flora of the Cerrado biome: Checklist with 12,356 species. In Cerrado: Ecologia e Flora; Embrapa Cerrados: Brasilia, DF, Brazil, 2008; Volume 2, pp. 422–442. [Google Scholar]
- Felfili, M.C.; Felfili, J.M. Diversidade alfa e beta no Cerrado Sensu Strictu da Chapada Pratinha, Brasil. Acta Bot. Bras. 2001, 15, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.F.; Farinas, M.R.; Felfili, J.M.; Klink, C.A. Spatial heterogeneity, land use and conservation in the cerrado region of Brazil. J. Biogeogr. 2006, 33, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.; José, J.S.; Meir, P.; Miranda, H.S.; Montes, R.A. Productivity and carbon fluxes of tropical savannas. J. Biogeogr. 2006, 33, 387–400. [Google Scholar] [CrossRef]
- Coulter, L.L.; Stow, D.A.; Tsai, Y.-H.; Ibanez, N.; Shih, H.; Kerr, A.; Benza, M.; Weeks, J.R.; Mensah, F. Classification and assessment of land cover and land use change in southern Ghana using dense stacks of Landsat 7 ETM+ imagery. Remote Sens. Environ. 2016, 184, 396–409. [Google Scholar] [CrossRef]
- Mayes, M.T.; Mustard, J.F.; Melillo, J.M. Forest cover change in Miombo Woodlands: Modeling land cover of African dry tropical forests with linear spectral mixture analysis. Remote Sens. Environ. 2015, 165, 203–215. [Google Scholar] [CrossRef]
- Müller, H.; Rufin, P.; Griffiths, P.; Barros Siqueira, A.J.; Hostert, P. Mining dense Landsat time series for separating cropland and pasture in a heterogeneous Brazilian savanna landscape. Remote Sens. Environ. 2015, 156, 490–499. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, S.N.; de Júnior, O.A.C.; Gomes, R.A.T.; Guimarães, R.F.; de Martins, É.S. Detecção de mudança do uso e cobertura da terra usando o método de pós-classificação na fronteira agrícola do oeste da Bahia sobre o Grupo Urucuia durante o período 1988–2011. Rev. Bras. Cartogr. 2014, 66, 1157–1176. [Google Scholar]
- Beuchle, R.; Grecchi, R.C.; Shimabukuro, Y.E.; Seliger, R.; Eva, H.D.; Sano, E.; Achard, F. Land cover changes in the Brazilian Cerrado and Caatinga biomes from 1990 to 2010 based on a systematic remote sensing sampling approach. Appl. Geogr. 2015, 58, 116–127. [Google Scholar] [CrossRef]
- de Oliveira, S.N.; Abílio de Carvalho Júnior, O.; Trancoso Gomes, R.A.; Fontes Guimarães, R.; McManus, C.M. Deforestation analysis in protected areas and scenario simulation for structural corridors in the agricultural frontier of Western Bahia, Brazil. Land Use Policy 2017, 61, 40–52. [Google Scholar] [CrossRef]
- Johansen, K.; Phinn, S.; Taylor, M. Mapping woody vegetation clearing in Queensland, Australia from Landsat imagery using the Google Earth Engine. Remote Sens. Appl. Soc. Environ. 2015, 1, 36–49. [Google Scholar] [CrossRef]
- Trancoso, R.; Sano, E.E.; Meneses, P.R. The spectral changes of deforestation in the Brazilian tropical savanna. Environ. Monit. Assess. 2014, 187, 4145. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Yoshioka, H.; Huete, A.; Sano, E.E. Seasonal landscape and spectral vegetation index dynamics in the Brazilian Cerrado: An analysis within the Large-Scale Biosphere–Atmosphere Experiment in Amazônia (LBA). Remote Sens. Environ. 2003, 87, 534–550. [Google Scholar] [CrossRef]
- Jin, C.; Xiao, X.; Merbold, L.; Arneth, A.; Veenendaal, E.; Kutsch, W.L. Phenology and gross primary production of two dominant savanna woodland ecosystems in Southern Africa. Remote Sens. Environ. 2013, 135, 189–201. [Google Scholar] [CrossRef]
- Schwieder, M.; Leitão, P.J.; da Cunha Bustamante, M.M.; Ferreira, L.G.; Rabe, A.; Hostert, P. Mapping Brazilian savanna vegetation gradients with Landsat time series. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 361–370. [Google Scholar] [CrossRef]
- Whiteside, T.G.; Boggs, G.S.; Maier, S.W. Comparing object-based and pixel-based classifications for mapping savannas. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 884–893. [Google Scholar] [CrossRef]
- Sano, E.E.; Ferreira, L.G.; Asner, G.P.; Steinke, E.T. Spatial and temporal probabilities of obtaining cloud-free Landsat images over the Brazilian tropical savanna. Int. J. Remote Sens. 2007, 28, 2739–2752. [Google Scholar] [CrossRef]
- Cochrane, M.A. Fire science for rainforests. Nature 2003, 421, 913–919. [Google Scholar] [CrossRef]
- Ferreira, M.E.; Ferreira, L.G.; Sano, E.E.; Shimabukuro, Y.E. Spectral linear mixture modelling approaches for land cover mapping of tropical savanna areas in Brazil. Int. J. Remote Sens. 2007, 28, 413–429. [Google Scholar] [CrossRef]
- Sano, E.E.; Rosa, R.; Brito, J.L.S.; Ferreira, L.G. Land cover mapping of the tropical savanna region in Brazil. Environ. Monit. Assess. 2010, 166, 113–124. [Google Scholar] [CrossRef]
- Hill, M.J.; Zhou, Q.; Sun, Q.; Schaaf, C.B.; Palace, M. Relationships between vegetation indices, fractional cover retrievals and the structure and composition of Brazilian Cerrado natural vegetation. Int. J. Remote Sens. 2017, 38, 874–905. [Google Scholar] [CrossRef]
- Ratana, P.; Huete, A.R.; Ferreira, L. Analysis of Cerrado Physiognomies and Conversion in the MODIS Seasonal-Temporal Domain. Earth Interact. 2005, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.G.; Huete, A.R. Assessing the seasonal dynamics of the Brazilian Cerrado vegetation through the use of spectral vegetation indices. Int. J. Remote Sens. 2004, 25, 1837–1860. [Google Scholar] [CrossRef]
- Franklin, J. Land cover stratification using Landsat Thematic Mapper data in Sahelian and Sudanian woodland and wooded grassland. J. Arid Environ. 1991, 20, 141–163. [Google Scholar] [CrossRef]
- Franklin, J.; Davis, F.W.; Lefebvre, P. Thematic mapper analysis of tree cover in semiarid woodlands using a model of canopy shadowing. Remote Sens. Environ. 1991, 36, 189–202. [Google Scholar] [CrossRef]
- Nagendra, H.; Rocchini, D. High resolution satellite imagery for tropical biodiversity studies: The devil is in the detail. Biodivers. Conserv. 2008, 17, 3431–3442. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Walter, B.M.T. As principais fitofisionomias do bioma Cerrado. In Cerrado: Ecologia e Flora; Embrapa Cerrados: Brasilia, DF, Brazil, 2008; pp. 151–199. [Google Scholar]
- Arroyo, L.A.; Johansen, K.; Armston, J.; Phinn, S. Integration of LiDAR and QuickBird imagery for mapping riparian biophysical parameters and land cover types in Australian tropical savannas. For. Ecol. Manag. 2010, 259, 598–606. [Google Scholar] [CrossRef]
- Gibbes, C.; Adhikari, S.; Rostant, L.; Southworth, J.; Qiu, Y. Application of Object Based Classification and High Resolution Satellite Imagery for Savanna Ecosystem Analysis. Remote Sens. 2010, 2, 2748–2772. [Google Scholar] [CrossRef] [Green Version]
- Nagendra, H.; Lucas, R.; Honrado, J.P.; Jongman, R.H.G.; Tarantino, C.; Adamo, M.; Mairota, P. Remote sensing for conservation monitoring: Assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Girolamo-Neto, C.D.; Fonseca, L.M.G.; Körting, S.; Soares, A.R. Mapping Brazilian Savanna Physiognomies using WorldView-2 Imagery and Geographic Object Based Image Analysis. In Proceedings of the GEOBIA 2018-From Pixels to Ecosystems and Global Sustainability, Montpellier, France, 18–22 June 2018. [Google Scholar]
- Blaschke, T.; Hay, G.J.; Kelly, M.; Lang, S.; Hofmann, P.; Addink, E.; Queiroz Feitosa, R.; van der Meer, F.; van der Werff, H.; van Coillie, F.; et al. Geographic Object-Based Image Analysis—Towards a new paradigm. ISPRS J. Photogramm. Remote Sens. 2014, 87, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Hay, G.J.; Niemann, K.O.; McLean, G.F. An object-specific image-texture analysis of H-resolution forest imagery. Remote Sens. Environ. 1996, 55, 108–122. [Google Scholar] [CrossRef]
- Lang, S. Object-based image analysis for remote sensing applications: Modeling reality—Dealing with complexity. In Object-Based Image Analysis: Spatial Concepts for Knowledge-Driven Remote Sensing Applications; Blaschke, T., Lang, S., Hay, G.J., Eds.; Lecture Notes in Geoinformation and Cartography; Springer: Berlin/Heidelberg, Germany, 2008; pp. 3–27. ISBN 978-3-540-77058-9. [Google Scholar]
- Blaschke, T. Object based image analysis for remote sensing.pdf. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef] [Green Version]
- Hay, G.J.; Castilla, G. Geographic Object-Based Image Analysis (GEOBIA): A new name for a new discipline. In Object-Based Image Analysis: Spatial Concepts for Knowledge-Driven Remote Sensing Applications; Blaschke, T., Lang, S., Hay, G.J., Eds.; Lecture Notes in Geoinformation and Cartography; Springer: Berlin/Heidelberg, Germany, 2008; pp. 75–89. ISBN 978-3-540-77058-9. [Google Scholar]
- Boggs, G.S. Assessment of SPOT 5 and QuickBird remotely sensed imagery for mapping tree cover in savannas. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 217–224. [Google Scholar] [CrossRef]
- Kaszta, Ż.; Van De Kerchove, R.; Ramoelo, A.; Cho, M.A.; Madonsela, S.; Mathieu, R.; Wolff, E. Seasonal Separation of African Savanna Components Using Worldview-2 Imagery: A Comparison of Pixel- and Object-Based Approaches and Selected Classification Algorithms. Remote Sens. 2016, 8, 763. [Google Scholar] [CrossRef] [Green Version]
- Girolamo-Neto, C.D.; Fonseca, L.M.G.; Körting, T.S. Assessment of texture features for Brazilian Savanna classification: A case study in Brasília National Park. Rev. Bras. Cartogr. 2017, 69, 891–901. [Google Scholar]
- Orozco Filho, J. Avaliação do Uso da Abordagem Orientada-Objeto com Imagens de Alta Resolução RapidEye na Classificação das Fitofisionomias do Cerrado; Universidade de Brasilia: Brasilia, Brazil, 2017. [Google Scholar]
- Teixeira, L.R.; Nunes, G.M.; Finger, Z.; Siqueira, A.J.B. Potencialidades da Classificação Orientada a Objetos em Imagens SPOT5 no Mapeamento de Fitofisionomias do Cerrado. Rev. Espac. 2015, 36, 1. [Google Scholar]
- Gomes, L.; Miranda, H.S.; Silvério, D.V.; Bustamante, M.M.C. Effects and behaviour of experimental fires in grasslands, savannas, and forests of the Brazilian Cerrado. For. Ecol. Manag. 2020, 458, 117804. [Google Scholar] [CrossRef]
- Dave, R.; Saint-Laurent, C.; Murray, L.; Antunes Daldegan, G.; Brouwer, R.; de Mattos Scaramuzza, C.A.; Raes, L.; Simonit, S.; Catapan, M.; García Contreras, G.; et al. Second Bonn Challenge Progress Report: Application of the Barometer in 2018; IUCN, International Union for Conservation of Nature: Gland, Switzerland, 2019; ISBN 978-2-8317-1980-1. [Google Scholar]
- Brannstrom, C.; Jepson, W.; Filippi, A.M.; Redo, D.; Xu, Z.; Ganesh, S. Land change in the Brazilian Savanna (Cerrado), 1986–2002: Comparative analysis and implications for land-use policy. Land Use Policy 2008, 25, 579–595. [Google Scholar] [CrossRef]
- Grimm, A.M. Interannual climate variability in South America: Impacts on seasonal precipitation, extreme events, and possible effects of climate change. Stoch. Environ. Res. Risk Assess. 2011, 25, 537–554. [Google Scholar] [CrossRef]
- Cole, M. The Savannas: Biogeography and Geobotany; Academic Press: London, UK, 1986. [Google Scholar]
- Oliveira-Filho, A.T.; Ratter, J.A. Vegetation physiognomies and woody flora of the Cerrado biome. In The Cerrados of Brazil: Ecology and Natural History of A Neotropical Savanna; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- Bridgewater, S.; Ratter, J.A.; Felipe Ribeiro, J. Biogeographic patterns, β-diversity and dominance in the cerrado biome of Brazil. Biodivers. Conserv. 2004, 13, 2295–2317. [Google Scholar] [CrossRef]
- Roberts, D.A.; Keller, M.; Soares, J.V. Studies of land-cover, land-use, and biophysical properties of vegetation in the Large Scale Biosphere Atmosphere experiment in Amazônia. Remote Sens. Environ. 2003, 87, 377–388. [Google Scholar] [CrossRef]
- Silva, A.D.; Bergamini, L.L. Biodiversidade. In Reserva Ecológica do IBGE; Compromisso com a biodiversidade; IBGE, Coordenacao de Recursos Naturais e Estudos Ambientais: Brasilia, DF, Brazil; Volume 2, in press.
- Pereira, B.A.S.; Furtado, P.P. Vegetação da Bacia do Córrego Taquara: Coberturas Naturais e Antrópicas. In Reserva Ecológica do IBGE: Biodiversidade Terrestre; IBGE, Coordenacao de Recursos Naturais e Estudos Ambientais: Rio de Janeiro, Brazil, 2011; Volume 1. [Google Scholar]
- Nou, E.; Costa, N. Diagnóstico da Qualidade Ambiental da Bacia do Rio São Francisco: Sub-Bacias do Oeste Baiano e Sobradinho; IBGE, Primeira Divisão de Geociências do Nordeste: Rio de Janeiro, Brazil, 1994; ISBN 85-240-0502-5.
- Silva, J.M.; Bates, J.M. Biogeographic Patterns and Conservation in the South American Cerrado: A Tropical Savanna Hotspot. BioScience 2002, 52, 225. [Google Scholar] [CrossRef]
- Santana, O.A.; de Júnior, O.A.C.; Gomes, R.A.T.; dos Cardoso, W.S.; de Martins, É.S.; Passo, D.P.; Guimarães, R.F. Distribuição de espécies vegetais nativas em distintos macroambientes na Região do oeste da Bahia. Rev. Espaço E Geogr. 2010, 13, 181–223. [Google Scholar]
- Furley, P.A.; Ratter, J.A. Soil Resources and Plant Communities of the Central Brazilian Cerrado and Their Development. J. Biogeogr. 1988, 15, 97–108. [Google Scholar] [CrossRef]
- Coutinho, L.M. O conceito de Cerrado. Rev. Bras. Bot. 1978, 11, 17–23. [Google Scholar]
- Eiten, G. Delimitation of the cerrado concept. Vegetatio 1978, 36, 169–178. [Google Scholar] [CrossRef]
- Di Gregorio, A.; Jansen, L.J.M. Land Cover Classification System (LCCS): Classification Concepts and User Manual; Food and Agriculture Organization: Rome, Italy, 2000. [Google Scholar]
- Radoux, J.; Bogaert, P.; Radoux, J.; Bogaert, P. Good Practices for Object-Based Accuracy Assessment. Remote Sens. 2017, 9, 646. [Google Scholar] [CrossRef] [Green Version]
- Baatz, M.; Schäpe, A. Multiresolution Segmentation: An Optimization Approach for High Quality Multi-Scale Image Segmentation; Wichmann-Verlag: Heidelberg, Germany, 2000; pp. 12–23. [Google Scholar]
- Schuster, C.; Förster, M.; Kleinschmit, B. Testing the red edge channel for improving land-use classifications based on high-resolution multi-spectral satellite data. Int. J. Remote Sens. 2012, 33, 5583–5599. [Google Scholar] [CrossRef]
- Myint, S.W.; Gober, P.; Brazel, A.; Grossman-Clarke, S.; Weng, Q. Per-pixel vs. object-based classification of urban land cover extraction using high spatial resolution imagery. Remote Sens. Environ. 2011, 115, 1145–1161. [Google Scholar] [CrossRef]
- Rouse, J.W.; Hass, H.R.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, Goddard Space Flight Center, NASA SP-351, NASA, Washington, DC, USA, 10–14 December 1973. [Google Scholar]
- McFeeters, S.K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M. Reserva Ecologica do IBGE: Biodiversidade Terrestre; IBGE, Coordenacao de Recursos Naturais e Estudos Ambientais: Rio de Janeiro, Brazil, 2011; Volume 1.
- Crippen, R.; Buckley, S.; Agram, P.; Belz, E.; Gurrola, E.; Hensley, S.; Kobrick, M.; Lavalle, M.; Martin, J.; Neumann, M.; et al. NASADEM Global Elevation Model: Methods and Progress. ISPRS Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, XLI-B4, 125–128. [Google Scholar] [CrossRef]
- Embrapa. Reunião Técnica de Levantamento de Solos. In Serviço Nacional de Levantamento e Conservação de Solos; Empresa Brasileira de Pesquisa Agropecuária: Rio de Janeiro, Brazil, 1979; p. 83. [Google Scholar]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Olofsson, P.; Foody, G.M.; Herold, M.; Stehman, S.V.; Woodcock, C.E.; Wulder, M.A. Good practices for estimating area and assessing accuracy of land change. Remote Sens. Environ. 2014, 148, 42–57. [Google Scholar] [CrossRef]
- Powell, R.L.; Matzke, N.; de Souza, C.; Clark, M.; Numata, I.; Hess, L.L.; Roberts, D.A. Sources of error in accuracy assessment of thematic land-cover maps in the Brazilian Amazon. Remote Sens. Environ. 2004, 90, 221–234. [Google Scholar] [CrossRef]
- Richards, J.A. Classifier performance and map accuracy. Remote Sens. Environ. 1996, 57, 161–166. [Google Scholar] [CrossRef]
- Stehman, S.V. Selecting and interpreting measures of thematic classification accuracy. Remote Sens. Environ. 1997, 62, 77–89. [Google Scholar] [CrossRef]
- MacLean, M.G.; Congalton, D.R.G. Map accuracy assessment issues when using an object-oriented approach. In Proceedings of the American Society for Photogrammetry and Remote Sensing 2012 Annual Conference, Sacramento, CA, USA, 19–23 March 2012; pp. 1–5. [Google Scholar]
- Radoux, J.; Bogaert, P. Accounting for the area of polygon sampling units for the prediction of primary accuracy assessment indices. Remote Sens. Environ. 2014, 142, 9–19. [Google Scholar] [CrossRef]
- Stehman, S.V.; Wickham, J.D.; Fattorini, L.; Wade, T.D.; Baffetta, F.; Smith, J.H. Estimating accuracy of land-cover composition from two-stage cluster sampling. Remote Sens. Environ. 2009, 113, 1236–1249. [Google Scholar] [CrossRef]
- Reynolds, J.; Wesson, K.; Desbiez, A.L.J.; Ochoa-Quintero, J.M.; Leimgruber, P. Using Remote Sensing and Random Forest to Assess the Conservation Status of Critical Cerrado Habitats in Mato Grosso do Sul, Brazil. Land 2016, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- IBGE. Manuais Técnicos de Geociências. In Instituto Brasileiro de Geografia e Estatística. Manual Técnico da Vegetação Brasileira: Sistema Fitogeográfico, Inventário das Formações Florestais e Campestres, Técnicas e Manejo de Coleções Botânicas, Procedimentos para Mapeamentos; IBGE—Diretoria de Geociências: Rio de Janeiro, Brazil, 2012. [Google Scholar]
- Jansen, L.J.M.; Gregorio, A.D. Parametric land cover and land-use classifications as tools for environmental change detection. Agric. Ecosyst. Environ. 2002, 91, 89–100. [Google Scholar] [CrossRef]
- Kosmidou, V.; Petrou, Z.; Bunce, R.G.H.; Mücher, C.A.; Jongman, R.H.G.; Bogers, M.M.B.; Lucas, R.M.; Tomaselli, V.; Blonda, P.; Padoa-Schioppa, E.; et al. Harmonization of the Land Cover Classification System (LCCS) with the General Habitat Categories (GHC) classification system. Ecol. Indic. 2014, 36, 290–300. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.J.; Radabaugh, K.R.; Moyer, R.P.; Muller-Karger, F.E. Enabling efficient, large-scale high-spatial resolution wetland mapping using satellites. Remote Sens. Environ. 2018, 208, 189–201. [Google Scholar] [CrossRef]
- Smith, J.H.; Wickham, J.D.; Stehman, S.V.; Yang, L. Impacts of Patch Size and Land-Cover Heterogeneity on Thematic Image Classification Accuracy. Photogramm. Eng. Remote Sens. 2002, 68, 65–70. [Google Scholar]
- Silva, L.R.; Sano, E.E. Análise das imagens do satélite RapidEye para discriminação da cobertura vegetal do bioma Cerrado. Rev. Bras. Cartogr. 2016, 68, 1269–1283. [Google Scholar]
- Baldeck, C.A.; Colgan, M.S.; Féret, J.-B.; Levick, S.R.; Martin, R.E.; Asner, G.P. Landscape-scale variation in plant community composition of an African savanna from airborne species mapping. Ecol. Appl. 2014, 24, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Baldeck, C.A.; Asner, G.P. Estimating Vegetation Beta Diversity from Airborne Imaging Spectroscopy and Unsupervised Clustering. Remote Sens. 2013, 5, 2057–2071. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, N.R.; Asner, G.P.; Smit, I.P.J.; Riddel, E.S. Multiple Scales of Control on the Structure and Spatial Distribution of Woody Vegetation in African Savanna Watersheds. PLoS ONE 2015, 10, e0145192. [Google Scholar] [CrossRef]
- Cho, M.A.; Mathieu, R.; Asner, G.P.; Naidoo, L.; van Aardt, J.; Ramoelo, A.; Debba, P.; Wessels, K.; Main, R.; Smit, I.P.J.; et al. Mapping tree species composition in South African savannas using an integrated airborne spectral and LiDAR system. Remote Sens. Environ. 2012, 125, 214–226. [Google Scholar] [CrossRef]
- Colgan, M.S.; Baldeck, C.A.; Féret, J.-B.; Asner, G.P. Mapping Savanna Tree Species at Ecosystem Scales Using Support Vector Machine Classification and BRDF Correction on Airborne Hyperspectral and LiDAR Data. Remote Sens. 2012, 4, 3462–3480. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, L.; Cho, M.A.; Mathieu, R.; Asner, G. Classification of savanna tree species, in the Greater Kruger National Park region, by integrating hyperspectral and LiDAR data in a Random Forest data mining environment. ISPRS J. Photogramm. Remote Sens. 2012, 69, 167–179. [Google Scholar] [CrossRef]
- Asner, G.P.; Levick, S.R.; Kennedy-Bowdoin, T.; Knapp, D.E.; Emerson, R.; Jacobson, J.; Colgan, M.S.; Martin, R.E. Large-scale impacts of herbivores on the structural diversity of African savannas. Proc. Natl. Acad. Sci. USA 2009, 106, 4947–4952. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecosystems | Physiognomic Types (English; Portuguese) | Description |
|---|---|---|
| Forest | Riparian Forest; mata riparia, mata de galeria, mata ciliar | Closed-canopy semi-deciduous and evergreen trees following rivers and streams. This class includes gallery forests with a variety of soil moisture regimes |
| Seasonally Dry Tropical Forest; mata seca (semi-decidual, decidual, sempre-verde), floresta estacional | Closed-canopy semi-deciduous, deciduous, and/or evergreen trees across nutrient-rich environments on interfluves. This class is associated with mountainous terrain, such as cliffs | |
| Semi-deciduous Forest; mata semi-decidual; cerradão; mata seca | Closed-canopy semi-deciduous trees with dense layer of xeromorphic shrubs located across flat interfluvial terrain. This class contains tropical dry forest and/or sclerophyll forest, which can occur in different successional stages due to recent deforestation or fire activity | |
| Savanna | Cerrado Woodland; cerrado denso | Open canopy semi-deciduous trees over an open herbaceous layer and dense layer of xeromorphic shrubs |
| Savanna; cerrado tipico | High density of xeromorphic shrubs over an herbaceous layer with scattered to medium density of trees; may contain elements of transition to caatinga vegetation | |
| Open Savanna; cerrado ralo; campo sujo | Low density of xeromorphic shrubs and sub-shrubs over a closed herbaceous layer, which may contain scattered trees throughout the landscape | |
| Grassland | Grassland; campo limpo, campo limpo com murundus | Treeless herbaceous layer |
| Non-natural Shrubby Grassland; campo sujo, campo sujo degradado, capoeira | Sparse xeromorphic shrubs over an open herbaceous layer with strong presence of exposed soil. This class may contain degraded areas, such as abandoned pastures and agricultural areas | |
| Scrub Cerrado; campo sujo denso, campo cerrado, scrub | High density of xeromorphic shrubs and sub-shrubs, with occasional scattered deciduous trees and no presence of herbaceous layer or soil. It may contain elements of transition to caatinga vegetation | |
| Wetlands | Shrub Swamp; vereda, scrub de vereda | High to low density of shrubs and sub-shrubs, usually clustered, over a seasonally flooded herbaceous layer |
| Palm Swamp; vereda | High to low density of palm trees (most commonly Mauritia flexuosa), either clustered throughout a seasonally flooded herbaceous layer, or aligned along a water course | |
| Marsh; brejo, campo limpo umido | Seasonally flooded herbaceous layer composed mainly of grass species. This class usually surrounds riparian forests and contains palm and shrub swamps |
| Selected Features/Statistics | Description |
|---|---|
| Brightness | Sum of mean values of all layers (spectral bands) divided by the total number of spectral bands |
| Mean Value | Mean (reflectance) value of each spectral band within an image object |
| Standard Deviation Value | Standard deviation (reflectance) value of each spectral band within an image object |
| Customized Attributes | |
| NDVI | (NIR–Red)/(NIR+Red) |
| NDVI-RE | (RedEdge–Red)/(RedEdge+Red) |
| NDWI | (Green–NIR)/(Green+NIR) |
| Level 1 Classes | Spatial Rules | Level 2 Classes |
|---|---|---|
| Closed Canopy | 1. Within hydromorphic soils | Riparian Forest |
| 2. Within steep slopes (>20%) and not adjacent to perennial streams and water (relative border to ‘streams’ = 0) ** | Seasonally Dry Tropical Forest ** | |
| 3. Adjacent to streams and water (relative border to ‘streams’ > 0) | Riparian Vegetation | |
| 4. Within high elevation (>670 m) and flat terrain (slope < 8%) ** | Semi-Deciduous Forest ** | |
| Open Canopy | 1. Within hydromorphic soils | Palm Swamp |
| 2. Within steep slopes (>20%) ** | Seasonally Dry Tropical Forest ** | |
| 3. All other conditions | Cerrado Woodland | |
| Dense Shrub | 1. Within hydromorphic soils | Shrub Swamp |
| 2. All other conditions | Savanna | |
| Open Shrub | 1. Within hydromorphic soils | Shrub Swamp |
| 2. All other conditions | Open Savanna | |
| Scrub–Shrub | 1. Within hydromorphic soils ** | Shrub Swamp ** |
| 2. All other conditions ** | Scrub Cerrado ** | |
| Herbaceous (wet) | 1. Within hydromorphic soils | Marsh |
| 2. Within steep slopes (>20%) ** | Shade ** | |
| 3. All other conditions | Non-Natural/Barren | |
| Herbaceous (dry) | Grassland * | |
| Herbaceous | Invasive Forbs and Shrubs * | |
| Water | 1. Isolated small objects (size < 60 pixels) not within hydromorphic soils ** | Shade ** |
| 2. Within steep slopes (>20%) | Shade ** | |
| 2. All other conditions ** | Water ** | |
| Shrub–Herbaceous | Semi-Natural Shrubby Grassland ** | |
| Soil, NPV ***, impervious surfaces (bright) | Non-Natural/Barren | |
| Soil, NPV ***, impervious surfaces (dark) |
| Closed Canopy (125) | Dense Shrub (90) | Herbaceous (Wet) (50) | Open Canopy (125) | Open Shrub (100) | Scrub–Shrub (59) | Shrub–Herb (50) | Soil, NPV, Impervious (Bright) (30) | Soil, NPV, Impervious (Dark) (46) | Water (50) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Closed Canopy | 17.8 | 0.1 | 0 | 2.0 | 0 | 0.5 | 0 | 0 | 0 | 0 |
| Dense Shrub | 0 | 15.8 | 0.1 | 0.5 | 2.3 | 0.2 | 0.4 | 0 | 0 | 0 |
| Herbaceous (wet) | 0 | 0 | 12.6 | 0.2 | 0.5 | 0 | 0 | 0 | 0 | 0 |
| Open Canopy | 0.7 | 1.7 | 0.5 | 13.9 | 0 | 0.3 | 0 | 0 | 0 | 0 |
| Open Shrub | 0 | 1.5 | 1.8 | 0 | 13.9 | 0.3 | 0.6 | 0 | 0 | 0 |
| Scrub–Shrub | 0.7 | 0 | 0 | 0 | 0.4 | 7.6 | 0.2 | 0 | 0 | 0 |
| Shrub–Herb | 0 | 0 | 0 | 0 | 0.7 | 0 | 6.8 | 0.1 | 0 | 0 |
| Soil, NPV, Impervious (bright) | 0 | 0 | 0 | 0 | 0 | 0 | 0.3 | 2.8 | 1.9 | 0 |
| Soil, NPV, Impervious (dark) | 0 | 0 | 0 | 0 | 0 | 0 | 0.9 | 0.1 | 9.6 | 0 |
| Water | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 7.7 |
| Overall Accuracy (%) | 84.4 | |||||||||
| Producer’s Accuracy (%) | 92.5% | 82.4% | 81.3% | 83.6% | 77.7% | 86.1% | 74.4% | 95.4% | 83.6% | 100% |
| User’s Accuracy (%) | 87.3% | 81.7% | 94.5% | 80.8% | 77.1% | 85.2% | 89.7% | 56.6% | 91.0% | 93.3% |
| Cerrado Woodland (50) | Marsh (50) | Non-Natural/Barren (76) | Open Savanna (74) | Palm Swamp (50) | Riparian Forest (50) | Savanna (75) | Scrub Cerrado (50) | Semi-Deciduous Forest (50) | Shrubby Grassland (50) | Shrub Swamp (50) | Tropical Dry Forest (50) | Water (50) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cerrado Woodland | 6.2 | 0 | 0 | 0 | 0 | 0.2 | 0.7 | 0.3 | 0.1 | 0 | 0 | 0 | 0 |
| Marsh | 0 | 12.6 | 0 | 0 | 0.2 | 0 | 0 | 0 | 0 | 0 | 0.5 | 0 | 0 |
| Non-Natural/Barren | 0 | 0 | 14.3 | 0 | 0 | 0 | 0 | 0 | 0 | 1.2 | 0 | 0 | 0 |
| Open Savanna | 0 | 0 | 0 | 11.7 | 0 | 0 | 1.3 | 0.3 | 0 | 0.6 | 0 | 0 | 0 |
| Palm Swamp | 0.1 | 0.5 | 0 | 0 | 5.0 | 0.4 | 0 | 0 | 0 | 0 | 0.7 | 0 | 0 |
| Riparian Forest | 0.2 | 0 | 0 | 0 | 0 | 5.9 | 0 | 0 | 0 | 0 | 0.3 | 0 | 0 |
| Savanna | 0 | 0 | 0 | 1.5 | 0 | 0 | 15.2 | 0 | 0.3 | 0.4 | 0 | 0 | 0 |
| Scrub Cerrado | 0 | 0 | 0 | 0.4 | 0 | 0 | 0.0 | 6.7 | 0.6 | 0.2 | 0 | 0 | 0 |
| Semi-Deciduous Forest | 0.4 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0.2 | 8.5 | 0 | 0 | 0 | 0 |
| Shrubby Grassland | 0 | 0 | 0.1 | 0.7 | 0 | 0 | 0 | 0 | 0 | 6.8 | 0 | 0 | 0 |
| Shrub Swamp | 0 | 1.9 | 0 | 0 | 0.2 | 0.1 | 0.1 | 0 | 0 | 0 | 5.0 | 0 | 0 |
| Tropical Dry Forest | 0.3 | 0 | 0 | 0 | 0 | 0 | 0.4 | 0 | 0 | 0 | 0 | 7.1 | 0 |
| Water | 0 | 0.6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7.7 |
| Overall Accuracy (%) | 87.6 | ||||||||||||
| Producer’s Accuracy (%) | 85.8% | 81.3% | 99.5% | 81.8% | 92.4% | 89.4% | 85.5% | 89.5% | 89.4% | 74.4% | 77.3% | 100% | 100% |
| User’s Accuracy (%) | 81.7% | 94.5% | 92.5% | 84.4% | 75% | 92.3% | 87.5% | 84.2% | 92.9% | 89.7% | 69.2% | 91.4% | 93.3% |
| Total (%) | September 13th Image | September 16th Image | June Mosaic | August Mosaic | October Mosaic | |
|---|---|---|---|---|---|---|
| Cerrado Woodland | 4.9 | 4.2 | 3.2 | 7.0 | 1.7 | 5.9 |
| Marsh | 1.2 | 0.2 | 0.0 | 0.3 | 1.4 | 2.9 |
| Non-Natural/Barren | 26.6 | 14.5 | 31.6 | 11.5 | 33.2 | 43.9 |
| Open Savanna | 26.2 | 35.9 | 32.1 | 22.9 | 37.2 | 15.5 |
| Palm Swamp | 0.8 | 0.1 | 0.1 | 0.2 | 0.7 | 2.0 |
| Riparian Forest | 1.1 | 0.4 | 0.4 | 1.3 | 0.8 | 1.4 |
| Savanna | 26.2 | 39.4 | 14.6 | 40.0 | 11.1 | 20.9 |
| Scrub Cerrado | 1.9 | 0.0 | 0.0 | 4.4 | 1.2 | 0.0 |
| Semi-Deciduous Forest | 1.5 | 0.5 | 0.1 | 3.4 | 0.4 | 0.2 |
| Shade | 0.1 | 0.0 | 0.0 | 0.3 | 0.1 | 0.0 |
| Semi-Natural Shrubby Grassland | 8.0 | 4.2 | 17.1 | 6.3 | 10.4 | 6.6 |
| Shrub Swamp | 0.6 | 0.1 | 0.1 | 0.5 | 1.3 | 0.2 |
| Seasonally Dry Tropical Forest | 0.6 | 0.2 | 0.4 | 1.5 | 0.1 | 0.0 |
| Water | 0.3 | 0.5 | 0.3 | 0.2 | 0.3 | 0.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, F.F.; Roberts, D.A.; Hess, L.L.; W. Davis, F.; Caylor, K.K.; Antunes Daldegan, G. Geographic Object-Based Image Analysis Framework for Mapping Vegetation Physiognomic Types at Fine Scales in Neotropical Savannas. Remote Sens. 2020, 12, 1721. https://doi.org/10.3390/rs12111721
Ribeiro FF, Roberts DA, Hess LL, W. Davis F, Caylor KK, Antunes Daldegan G. Geographic Object-Based Image Analysis Framework for Mapping Vegetation Physiognomic Types at Fine Scales in Neotropical Savannas. Remote Sensing. 2020; 12(11):1721. https://doi.org/10.3390/rs12111721
Chicago/Turabian StyleRibeiro, Fernanda F., Dar A. Roberts, Laura L. Hess, Frank W. Davis, Kelly K. Caylor, and Gabriel Antunes Daldegan. 2020. "Geographic Object-Based Image Analysis Framework for Mapping Vegetation Physiognomic Types at Fine Scales in Neotropical Savannas" Remote Sensing 12, no. 11: 1721. https://doi.org/10.3390/rs12111721