Monitoring Grass Phenology and Hydrological Dynamics of an Oak–Grass Savanna Ecosystem Using Sentinel-2 and Terrestrial Photography

 and
and
Abstract
:
1. Introduction
2. Material and Methods
2.1. Study Site and Datasets
- Air temperature, recorded using an HMP45A probe (Vaisala OyJ); daily minimum (Tmin), maximum (Tmax) and mean temperature (Tmed) values were computed from half-hourly records.
- Incoming and outgoing solar radiation (Rad), measured with a four-way radiometer NR-01 (Campbell Sci. Inc); daily cumulative radiation fluxes were computed from half-hourly records.
- Vapor pressure deficit (VPD) was derived from atmospheric pressure and relative humidity (HMP45A probe (Vaisala OyJ); daily values were computed from half-hourly records.
- Precipitation (R) was measured with a weighing-type recording rain-gauge ARG100 (Campbell Sci. Inc); daily values were computed from the aggregation of 30 min cumulative values.
- Volumetric soil moisture (SM) was measured at two depths (10 and 30 cm) with an ENVIROSCAN (Campbell Sci. Inc) probe at 10 min intervals. Daily values were computed from these data and averaged between both depths.
2.2. Phenological Parameters From Terrestrial Photography
- GCC: Green Chromatic Coordinate index;
- R: digital level in red;
- G: digital level in green;
- B: digital level in blue.
- v(t): value of the function at time t;
- vmin: minimum value of the amplitude;
- vmax: maximum value of the amplitude;
- x and y: parameters that control the shape of the curve. x1 and x2 control the left and right inflexion points, respectively, and y1 and y2 represent the rate of change at time t.
2.3. Selection of Satellite Vegetation Indices
2.4. Selection of Abiotic Variables
2.5. Satellite VI and Abiotic Variable Relationship
2.6. Statistical Analysis
3. Results
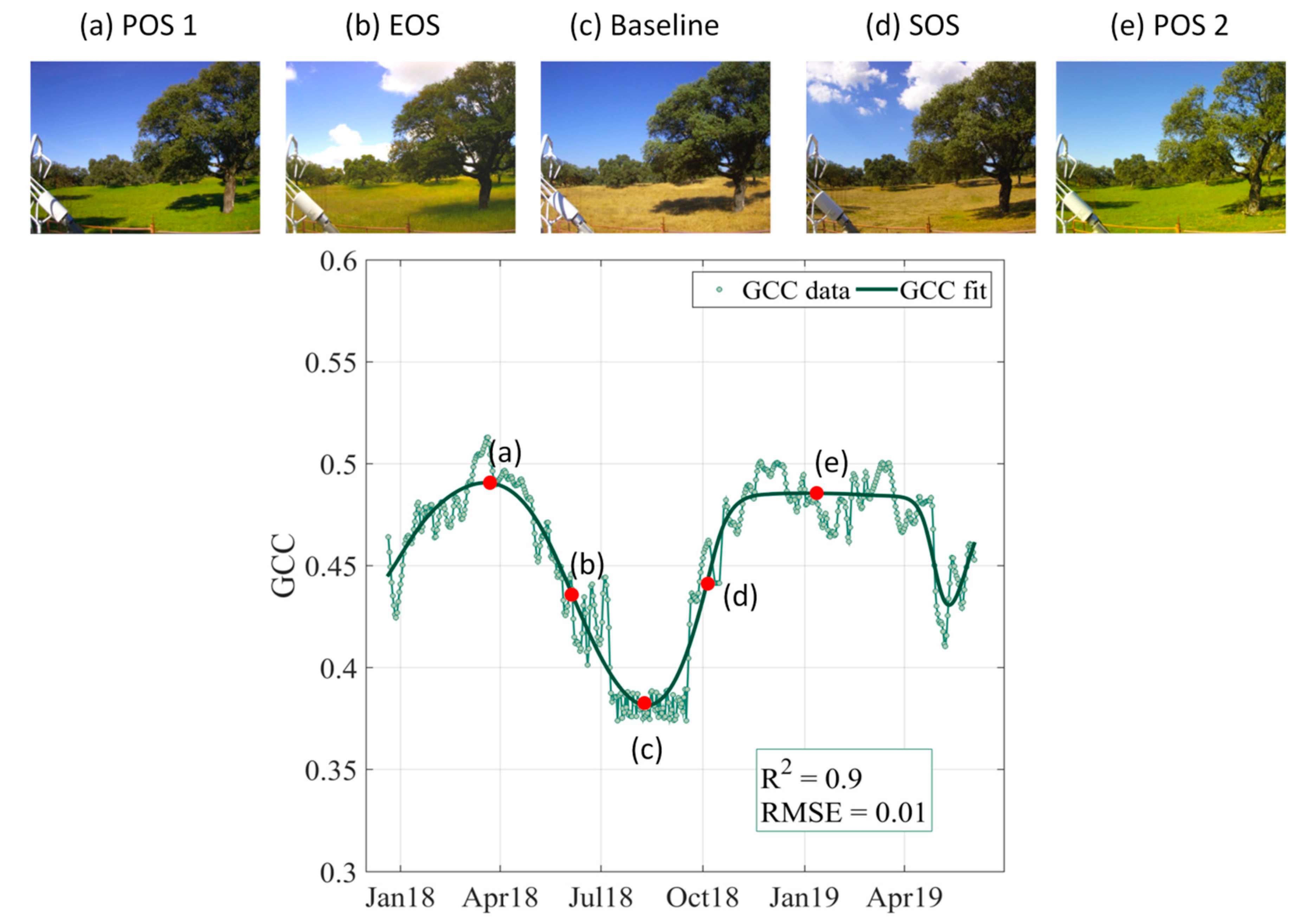
3.1. Deriving Phenological Parameters From Terrestrial Photography
3.2. Terrestrial Camera vs. Satellite-derived Indices
3.2.1. Satellite and Ground-based Indices Comparison
3.2.2. Satellite-derived Phenology
3.3. Analysis of Abiotic Variables and Greenness Dynamics
Grassland Phenology and Soil Moisture Relationship
3.4. Relationships Between Soil Moisture and NDVI
4. Discussion
4.1. Capability of the Terrestrial Photography to Provide Phenological Parameters
4.2. Comparison of Ground and Satellite-based Indices
4.3. Analysis of the Dynamics of Abiotic Variables, Greenness and Phenology
4.4. Relationship Between SM and NDVI
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olea, L.; Miguel-ayanz, A.S. The Spanish dehesa, a traditional Mediterranean silvopastoral system. In Proceedings of the 21st General Meeting of the European Grassland Federation, Badajoz, Spain, 3–6 April 2006; pp. 1–15. [Google Scholar]
- Scholes, R.J.; Archer, S.R. Tree-Grass Interactions in Savannas. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Eagleson, P.S.; Segarra, R.I. Water-Limited Equilibrium of Savanna Vegetation Systems. Water Resour. Res. 1985, 21, 1483–1493. [Google Scholar] [CrossRef] [Green Version]
- Eagleson, P.S.; Tellers, T.E. Ecological optimality in water-limited natural soil-vegetation systems: 2. Tests and applications. Water Resour. Res. 1982, 18, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Moreno, G.; Gonzalez-Bornay, G.; Pulido, F.; Lopez-Diaz, M.L.; Bertomeu, M.; Juárez, E.; Diaz, M. Exploring the causes of high biodiversity of Iberian dehesas: The importance of wood pastures and marginal habitats. Agrofor. Syst. 2016, 90, 87–105. [Google Scholar] [CrossRef]
- Turner, N.C. Drought resistance and adaptation to water deficits in crop plants. Stress Physiol. Crop Plants 1979, 343–372. Available online: https://ci.nii.ac.jp/naid/10006285376/en/ (accessed on 25 December 2019).
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 37–42. [Google Scholar] [CrossRef]
- IPCC 2001. Climate change 2001: Impacts, adaptation, and vulnerability. Contribution of Working Group II to the third assessment report of the Intergovernmental Panel on Climate Change (IPCC). In Choice Reviews Online (Vol. 39); McCarthy, J.J., Canziani, O.F., Leary, N.A., Dokken, D.J., White, K.S., Eds.; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar] [CrossRef]
- Parmesan, C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Glob. Chang. Biol. 2007, 13, 1860–1872. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef]
- Menzel, A. Phenology: Its importance to the global change community—An editorial comment. Clim. Chang. 2002, 54, 379–385. [Google Scholar] [CrossRef]
- Rafferty, N.E.; CaraDonna, P.J.; Burkle, L.A.; Iller, A.M.; Bronstein, J.L. Phenological overlap of interacting species in a changing climate: An assessment of available approaches. Ecol. Evol. 2013, 3. [Google Scholar] [CrossRef]
- Fisher, J.I.; Richardson, A.D.; Mustard, J.F. Phenology model from surface meteorology does not capture satellite-based greenup estimations. Glob. Chang. Biol. 2007, 13, 707–721. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, A.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Jin, Z.; Zhuang, Q.; He, J.S.; Luo, T.; Shi, Y. Phenology shift from 1989 to 2008 on the Tibetan Plateau: An analysis with a process-based soil physical model and remote sensing data. Clim. Chang. 2013, 119, 435–449. [Google Scholar] [CrossRef]
- Viña, A.; Gitelson, A.A.; Rundquist, D.C.; Keydan, G.; Leavitt, B.; Schepers, J. Monitoring maize (Zea mays L.) phenology with remote sensing. Agron. J. 2004, 96, 1139–1147. [Google Scholar] [CrossRef]
- Xiao, X.; Hagen, S.; Zhang, Q.; Keller, M.; Moore, B. Detecting leaf phenology of seasonally moist tropical forests in South America with multi-temporal MODIS images. Remote Sens. Environ. 2006, 103, 465–473. [Google Scholar] [CrossRef]
- Duchemin, B.; Hadria, R.; Erraki, S.; Boulet, G.; Maisongrande, P.; Chehbouni, A.; Escadafal, R.; Ezzahar, J.; Hoedjes, J.C.B.; Kharrou, M.H.; et al. Monitoring wheat phenology and irrigation in Central Morocco: On the use of relationships between evapotranspiration, crops coefficients, leaf area index and remotely-sensed vegetation indices. Agric. Water Manag. 2006, 79, 1–27. [Google Scholar] [CrossRef]
- Peña-Barragán, J.M.; Ngugi, M.K.; Plant, R.E.; Six, J. Object-based crop identification using multiple vegetation indices, textural features and crop phenology. Remote Sens. Environ. 2011, 115, 1301–1316. [Google Scholar] [CrossRef]
- Lamb, D.W.; Weedon, M.M.; Bramley, R.G.V. Using remote sensing to predict grape phenolics and colour at harvest in a Cabernet Sauvignon vineyard: Timing observations against vine phenology and optimising image resolution. Aust. J. Grape Wine 2004, 10, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Poenaru, V.; Badea, A.; Dana Negula, I.; Moise, C. Monitoring Vegetation Phenology in the Braila Plain Using Sentinel 2 Data. Sci. Pap. Ser. E Land R 2017, 6, 175–180. Available online: https://scihub.copernicus.eu/ (accessed on 25 December 2019).
- Lange, M.; Dechant, B.; Rebmann, C.; Vohland, M.; Cuntz, M.; Doktor, D. Validating MODIS and sentinel-2 NDVI products at a temperate deciduous forest site using two independent ground-based sensors. Sensors 2017, 17, 1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jönsson, P.; Cai, Z.; Melaas, E.; Friedl, M.A.; Eklundh, L. A method for robust estimation of vegetation seasonality from Landsat and Sentinel-2 time series data. Remote Sens. 2018, 10, 635. [Google Scholar] [CrossRef] [Green Version]
- Frampton, W.J.; Dash, J.; Watmough, G.; Milton, E.J. Evaluating the capabilities of Sentinel-2 for quantitative estimation of biophysical variables in vegetation. ISPRS J. Photogramm Remote Sens. 2013, 82, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-surface remote sensing of spatial and temporal variation in canopy phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Motohka, T.; Nasahara, K.N.; Oguma, H.; Tsuchida, S. Applicability of Green-Red Vegetation Index for remote sensing of vegetation phenology. Remote Sens. 2010, 2, 2369–2387. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.B.; Hultine, K.R.; Steltzer, H.; Denny, E.G.; Denslow, M.W.; Granados, J.; Richardson, A.D. Using phenocams to monitor our changing earth: Toward a global phenocam network. Front. Ecol. Environ. 2016, 14, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.E.; Beringer, J.; Evans, B.; Hutley, L.B.; Tapper, N.J. Tree-grass phenology information improves light use efficiency modelling of gross primary productivity for an Australian tropical savanna. Biogeosciences 2017, 14, 111–129. [Google Scholar] [CrossRef] [Green Version]
- Knox, S.H.; Dronova, I.; Sturtevant, C.; Oikawa, P.Y.; Matthes, J.H.; Verfaillie, J.; Baldocchi, D. Using digital camera and Landsat imagery with eddy covariance data to model gross primary production in restored wetlands. Agric. For. Meteorol. 2017, 237–238. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, R.; Herrero, J.; Polo, M.J. Subgrid parameterization of snow distribution at a Mediterranean site using terrestrial photography. Hydrol. Earth Syst. Sc. 2017, 21, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Polo, M.J.; Herrero, J.; Pimentel, R.; Pérez-Palazón, M.J. The Guadalfeo Monitoring Network (Sierra Nevada, Spain): 14 years of measurements to understand the complexity of snow dynamics in semiarid regions. Earth Syst. Sci. Data 2019, 11, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Migliavacca, M.; Galvagno, M.; Cremonesec, E.; Rossini, M.; Meroni, M.; Sonnentage, O.; Cogliati, S.; Mancaf, G.; Diotri, F.; Busetto, L.; et al. Using digital repeat photography and eddy covariance data to model grassland phenology and photosynthetic CO2 uptake. Agric. For. Meteorol. 2011, 151, 1325–1337. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, C.; Peng, D.; Wang, S.; Gonsamo, A.; Fang, B.; Yuan, W. Improved modeling of gross primary production from a better representation of photosynthetic components in vegetation canopy. Agric. For. Meteorol. 2017, 233, 222–234. [Google Scholar] [CrossRef]
- Moore, C.E.; Brown, T.; Keenan, T.F.; Duursma, R.A.; Van Dijk, A.I.J.M.; Beringer, J.; Liddell, M.J. Reviews and syntheses: Australian vegetation phenology: New insights from satellite remote sensing and digital repeat photography. Biogeosciences 2016, 13, 5085–5102. [Google Scholar] [CrossRef] [Green Version]
- Norrant, C.; Douguédroit, A. Monthly and daily precipitation trends in the Mediterranean (1950–2000). Theor. Appl. Climatol. 2006, 83, 89–106. [Google Scholar] [CrossRef]
- Cortesi, N.; González-Hidalgo, J.C.; Brunetti, M.; Martin-Vide, J. Daily precipitation concentration across Europe 1971–2010. Nat. Hazards Earth Syst. Sci. 2012, 12, 2799–2810. [Google Scholar] [CrossRef] [Green Version]
- Solomon, S.; Quin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.; Tignort, M.; Miller, H. Climate Change: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2017. [Google Scholar]
- Kovats, R.S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.F. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1267–1326. [Google Scholar]
- Gómez-Giráldez, P.J.; Aguilar, C.; Polo, M.J. Natural vegetation covers as indicators of the soil water content in a semiarid mountainous watershed. Ecol. Indic. 2014, 46, 524–535. [Google Scholar] [CrossRef]
- Andreu, A.; Kustas, W.P.; Polo, M.J.; Carrara, A.; González-Dugo, M.P. Modeling surface energy fluxes over a dehesa (oak savanna) ecosystem using a thermal based two-source energy balance model (TSEB) I. Remote Sens. 2018, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- García-moreno, A.; Fernández-Rebollo, P.; Muñoz, M.; Carbonero, M. Gestión de los pastos en la dehesa. Instituto de Investigación y Formación Agraria y Pesquera (IFAPA). 2016 Dep. Legal CO-614-2016. Available online: https://www.juntadeandalucia.es/agriculturaypesca/ifapa/servifapa/contenidoAlf?id=1e33ce7b-9a13-4d73-8832-f367be91c551§or=69cc80a0-9a2d-11df-accb-b374239e8181§orf=69cc80a0-9a2d-11df-accb-b374239e8181&l=material (accessed on 25 December 2019).
- Schaap, M.G.; Leij, F.J.; Van Genuchten, M.T. Rosetta: A computer program for estimating soil hydraulic parameters with hierarchical pedotransfer functions. J. Hydrol. 2001, 251, 163–176. [Google Scholar] [CrossRef]
- Agencial Estatal de Meteorología (Ministerio de Medio Ambiente y Medio Rural y Marino); Instituto de Meteorologia de Portugal. Atlas climático de España y Portugal. 2011. Available online: http://www.aemet.es/es/serviciosclimaticos/datosclimatologicos/atlas_climatico (accessed on 25 December 2019).
- Gillespie, A.R.; Kahle, A.B.; Walker, R.E. Color enhancement of highly correlated images. II. Channel ratio and “chromaticity” transformation techniques. Remote Sens. 1987, 22, 343–365. [Google Scholar] [CrossRef]
- Vrieling, A.; Meroni, M.; Darvishzadeh, R.; Skidmore, A.K.; Wang, T.; Zurita-Milla, R.; Oosterbeek, K.; O’Connor, B.; Paganini, M. Vegetation phenology from Sentinel-2 and field cameras for a Dutch barrier island. Remote Sens. Environ. 2018, 215, 517–529. [Google Scholar] [CrossRef]
- Zhang, X.; Jayavelu, S.; Liu, L.; Friedl, M.A.; Henebry, G.M.; Liu, Y.; Schaaf, C.-B.; Richardson, A.D.; Gray, J. Evaluation of land surface phenology from VIIRS data using time series of PhenoCam imagery. Agric. For. Meteorol. 2018, 256–257, 137–149. [Google Scholar] [CrossRef]
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT - A program for analyzing time-series of satellite sensor data. Comput. Geoscis. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green leaf phenology at Landsat resolution: Scaling from the field to the satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F. Cross-Scalar Satellite Phenology from Ground, Landsat, and MODIS Data. Remote Sens. Environ. 2007, 109, 261–273. [Google Scholar] [CrossRef]
- Eklundh, L.; Jönson, P. TIMESAT: A Software Package for Time-Series Processing and Assessment of Vegetation Dynamics. In Remote Sensing Time Series. Remote Sensing and Digital Image Processing; Kuenzer, C., Dech, S., Wagner, W., Eds.; Springer: Cham, Switzerland, 2015; p. 22. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans Geosci Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, W.D. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, NASA SP-351, Washington, DC, USA, 10–14 December 1973; pp. 309–317. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index. Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreria, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Dash, J.; Curran, P.J. 2004. The MERIS terrestrial chlorophyll index. Int. J. Remote Sens. 2004, 25, 5403–5413. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least-Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Chen, J.; Jönson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Jackson, J.E. A User’s Guide to Principal Components; John Wiley and Sons: New York, NY, USA, 1991; p. 592. [Google Scholar]
- Luo, Y.; El-Madany, T.S.; Filippa, G.; Ma, X.; Ahrens, B.; Carrara, A.; Migliavacca, M. Using near-infrared-enabled digital repeat photography to track structural and physiological phenology in Mediterranean tree-grass ecosystems. Remote Sens. 2018, 10, 1293. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Frolking, S. Intercomparison of phenological transition dates derived from the PhenoCam Dataset V1.0 and MODIS satellite remote sensing. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Huete, A.R.; Liu, H.Q.; van Leeuwen, W.J.D. Use of vegetation indices in forested regions: Issues of linearity and saturation. Int. Geosci. Remote Sens. Symp. (IGARSS) 1997, 4, 1966–1968. [Google Scholar] [CrossRef]
- Gu, Y.; Wylie, B.K.; Howard, D.M.; Phuyal, K.P.; Ji, L. NDVI saturation adjustment: A new approach for improving cropland performance estimates in the Greater Platte River Basin, USA. Ecol. Indic. 2013, 30, 1–6. [Google Scholar] [CrossRef]
- Glenn, E.P.; Huete, A.R.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices and plant physiological processes: What vegetation indices can and cannot tell us about the landscape. Sensors 2008, 8, 2136. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Galiano, V.F.; Dash, J.; Atkinson, P.M. Intercomparison of satellite sensor land surface phenology and ground phenology in Europe. Geophys. Res. 2015, 42, 2253–2260. [Google Scholar] [CrossRef] [Green Version]
- Sibanda, M.; Mutanga, O.; Rouget, M. Testing the capabilities of the new WorldView-3 space-borne sensor’s red-edge spectral band in discriminating and mapping complex grassland management treatments. Int. J. Remote Sens. 2017, 38, 1–22. [Google Scholar] [CrossRef]
- Browning, D.M.; Karl, J.W.; Morin, D.; Richardson, A.D.; Tweedie, C.E. Phenocams bridge the gap between field and satellite observations in an arid grassland ecosystem. Remote Sens. 2017, 9, 1071. [Google Scholar] [CrossRef] [Green Version]
- Bolton, D.K.; Friedl, M.A. Forecasting crop yield using remotely sensed vegetation indices and crop phenology metrics. Agric. For. Meteorol. 2007, 173, 74–84. [Google Scholar] [CrossRef]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite remote sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using PhenoCam imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef] [Green Version]
- Jolly, W.M.; Running, S.W. Effects of precipitation and soil water potential on drought deciduous phenology in the Kalahari. Glob. Chang. Biol. 2004, 10, 303–308. [Google Scholar] [CrossRef]
- Allen, R.G. FAO Irrigation and Drainage Paper Crop. Irrig. Drain. 1998, 300, 300. [Google Scholar] [CrossRef]
- Jones, M.O.; Kimball, J.S.; Nemani, R.R. Asynchronous Amazon. forest canopy phenology indicates adaptation to both water and light availability. Environ. Res. 2014, 9. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Sentinel-2 Formulation | Spatial Resolution |
|---|---|---|
| EVI | 2.5(B8 − B4)/(B8 + 6B4 − 7.5B2 + 1) | 10 m |
| EVI2 | 2.5(B8 − B4)/(B8 + 2.4B4 + 1) | 10 m |
| GCCs | B3/(B2 + B3 + B4) | 10 m |
| GNDVI | (B8 − B3)/(B8 + B3) | 10 m |
| IRECI | (B7 − B4)/(B5/B6) | 20 m |
| MTCI | (B6 − B5)/(B5 − B4) | 20 m |
| NDVI | (B8 − B4)/(B8 + B4) | 10 m |
| S2REP | 705 + 35(((B7 + B4)/2) − B5)/(B6 − B5)) | 20 m |
| SAVI | 1.5(B8 − B4)/(B8 + B4 + 0.5) | 10 m |
| Variable | r (GCC) |
|---|---|
| EVI | 0.72 * |
| EVI2 | 0.77 * |
| GCCs | 0.79 * |
| GNDVI | 0.82 * |
| IRECI | 0.71 * |
| MTCI | 0.52 * |
| NDVI | 0.83 * |
| S2REP | −0.44 * |
| SAVI | 0.78 * |
| POS 1 (raw data) | POS 1 | EOS | SOS | POS 2 | POS 2 (raw data) | |
|---|---|---|---|---|---|---|
| EVI | −8 (3.7) | 1 (0.5) | 6 (2.9) | 31 (14.8) | 30 (14.3) | 36 (17.2) |
| EVI2 | 20 (9.3) | 28 (13.3) | −2 (9) | 25 (11.9) | 23 (11) | 23 (11) |
| GCCs | 18 (8.7) | 28 (13.3) | −13 (6.2) | 12 (5.7) | 3 (1.4) | −3 (1.4) |
| GNDVI | 13 (6) | 22 (10.5) | −6 (2.9) | 9 (4.3) | 11 (5.2) | 6 (2.9) |
| IRECI | 26 (12.4) | 31 (14.8) | −5 (2.4) | 34 (16.2) | 30 (14.3) | 37 (17.7) |
| NDVI | 4 (1.9) | 9 (4.3) | 1 (0.5) | 9 (4.3) | 9 (4.3) | 2 (0.9) |
| SAVI | 17 (8.2) | 26 (12.4) | −1 (0.5) | 21 (10) | 21 (10) | 23 (11) |
| Variable | r (GCCc) |
|---|---|
| SM | 0.75 * |
| VPD | −0.68 * |
| R | 0.17 * |
| Rad | −0.56 * |
| Tmin | −0.68 * |
| Tmed | −0.72 * |
| Tmax | −0.69 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Giráldez, P.J.; Pérez-Palazón, M.J.; Polo, M.J.; González-Dugo, M.P. Monitoring Grass Phenology and Hydrological Dynamics of an Oak–Grass Savanna Ecosystem Using Sentinel-2 and Terrestrial Photography. Remote Sens. 2020, 12, 600. https://doi.org/10.3390/rs12040600
Gómez-Giráldez PJ, Pérez-Palazón MJ, Polo MJ, González-Dugo MP. Monitoring Grass Phenology and Hydrological Dynamics of an Oak–Grass Savanna Ecosystem Using Sentinel-2 and Terrestrial Photography. Remote Sensing. 2020; 12(4):600. https://doi.org/10.3390/rs12040600
Chicago/Turabian StyleGómez-Giráldez, Pedro J., María J. Pérez-Palazón, María J. Polo, and María P. González-Dugo. 2020. "Monitoring Grass Phenology and Hydrological Dynamics of an Oak–Grass Savanna Ecosystem Using Sentinel-2 and Terrestrial Photography" Remote Sensing 12, no. 4: 600. https://doi.org/10.3390/rs12040600