
One-Class Classification of Natural Vegetation Using Remote Sensing: A Review

Abstract
1. Introduction
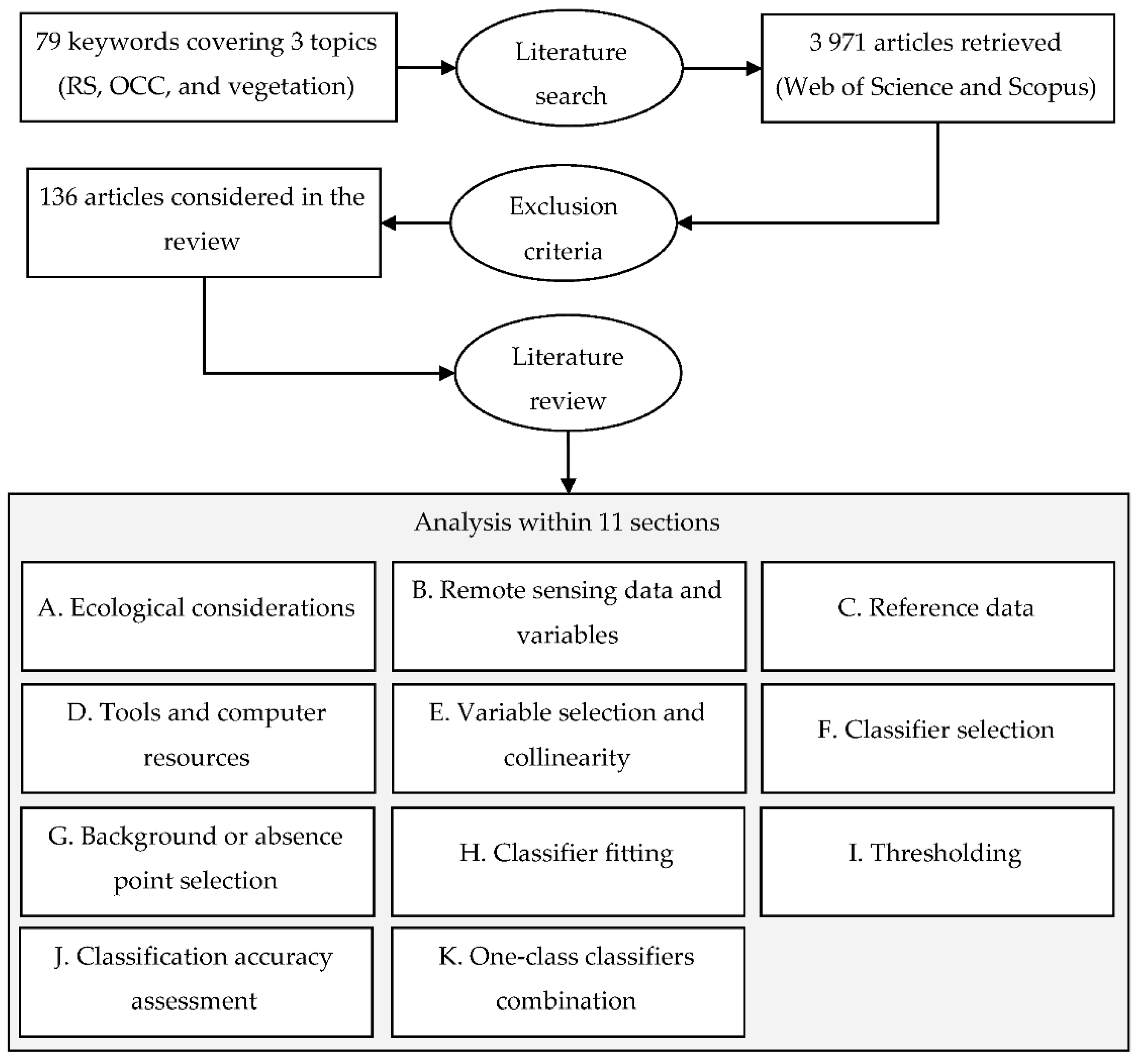
2. Literature Search and Review
3. A Wide Range of RS Data for Multiple Ecological Considerations
3.1. Identifying Potential Restoration or Invasion Areas
3.2. From Plant Species to Land Cover
3.3. Site Extent and Spatial Scale
3.4. Long- or Short-Term Vegetation Monitoring
3.5. The Importance of Spatio-Temporal Resolutions
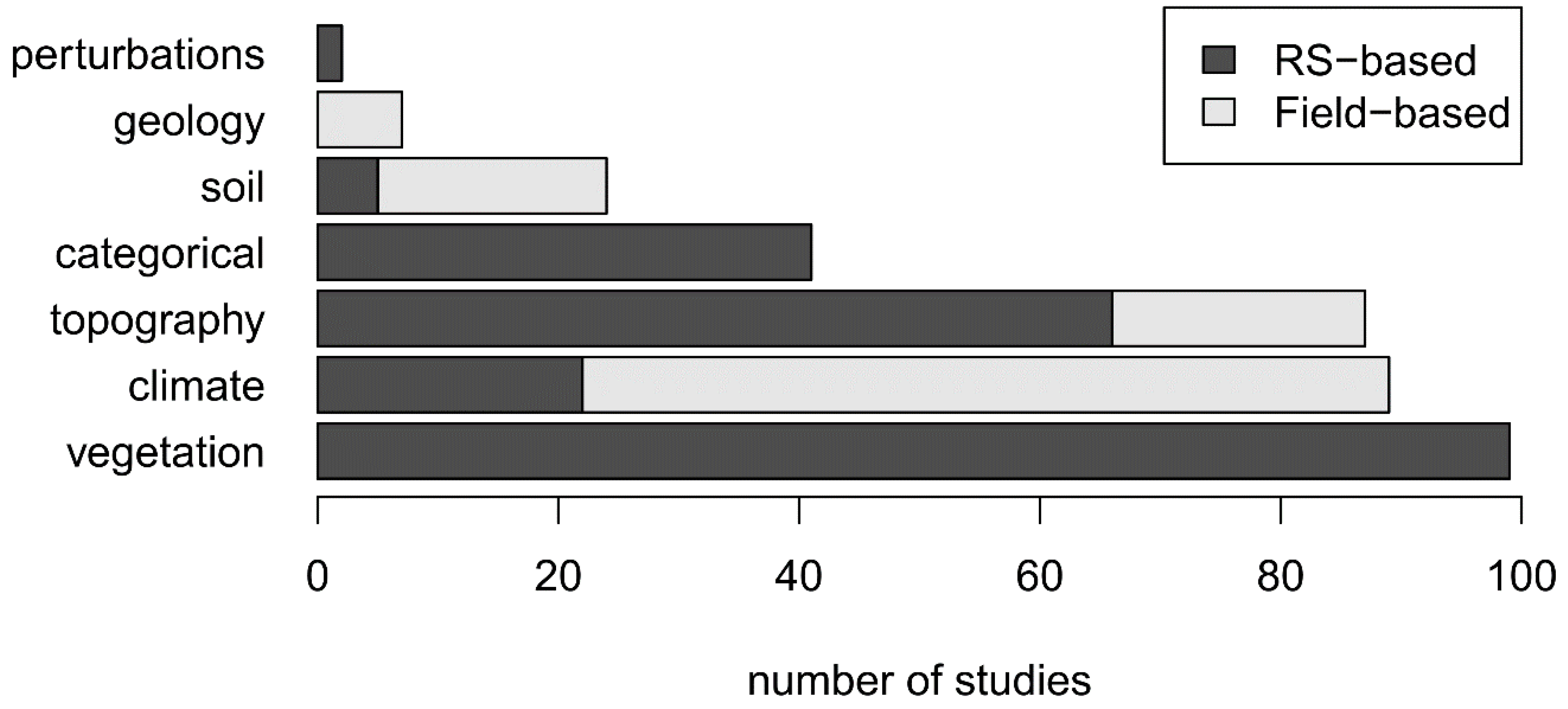
3.6. Underused RS-Based Environmental Variables
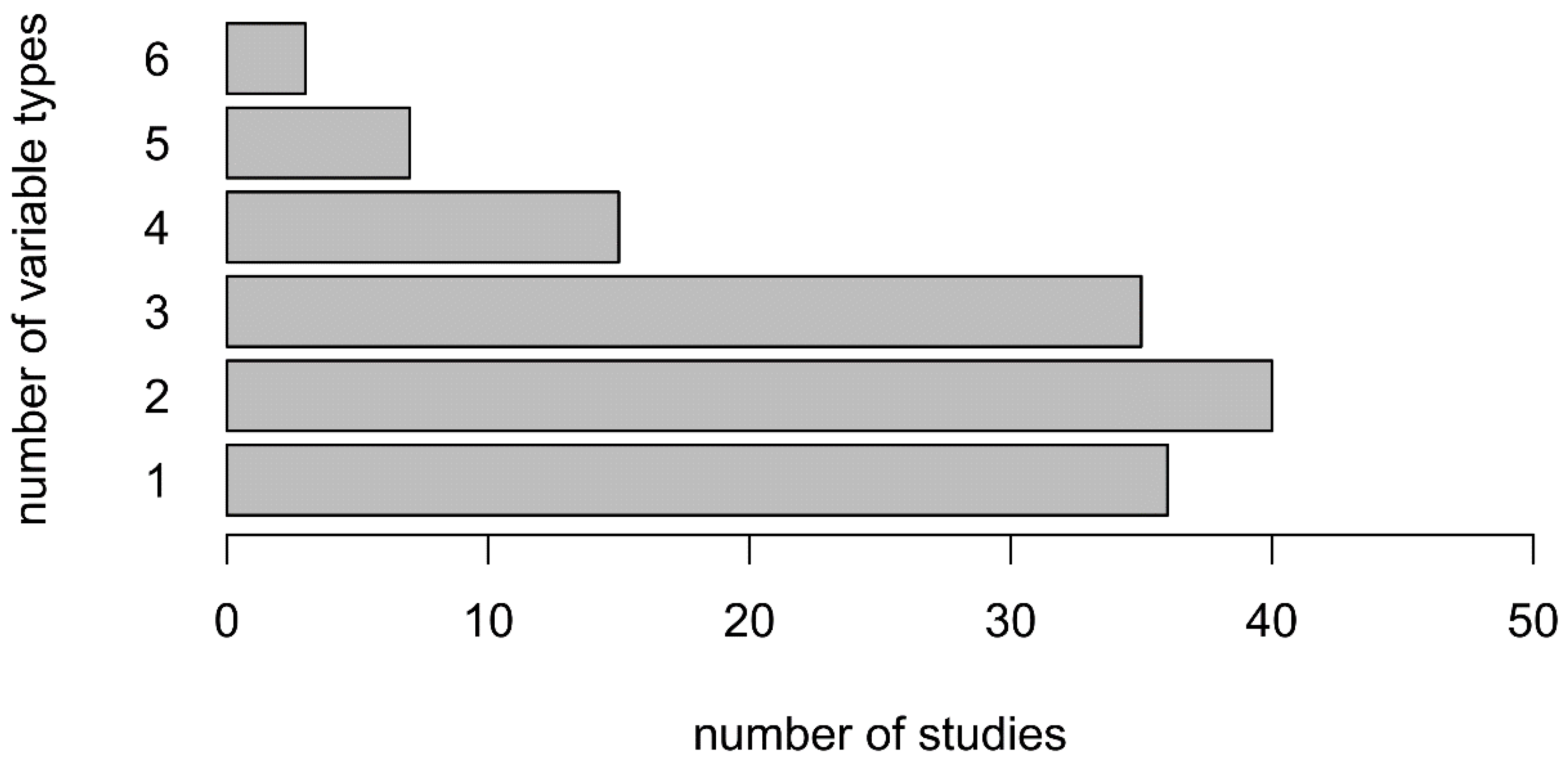
3.7. Combining Variables Improves OCC Performance
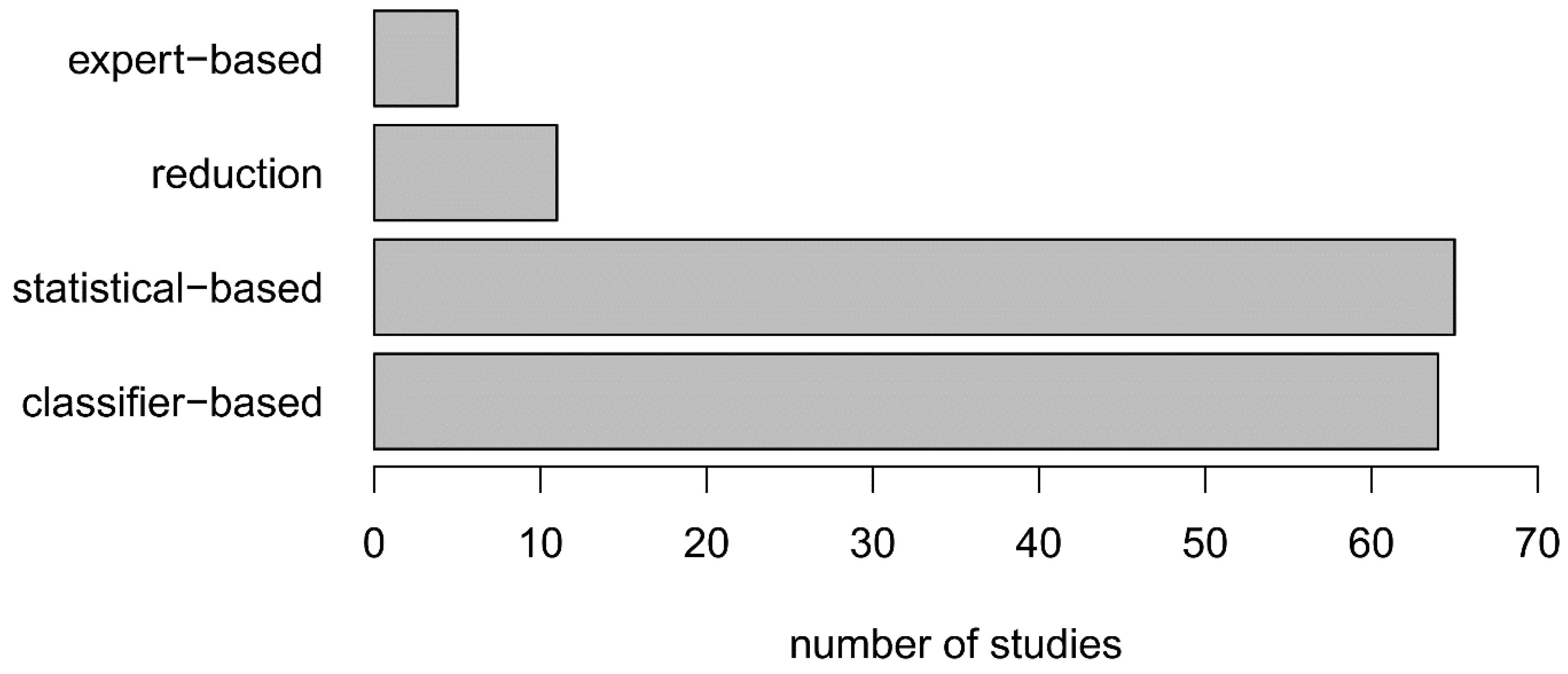
3.8. Data Quality Influences OCC Performance
- Subsampling: Several authors corrected for spatial sampling bias by subsampling reference data in densely plotted areas [118].
- UAV image analysis: Reference data can be derived automatically from very high spatial resolution UAV images [75]. This approach is interesting since pure pixels can be extracted from reference polygons, and reference data can be collected from sites that are difficult to access in the field.
4. A Wide Range of Tools and Settings for OCCs
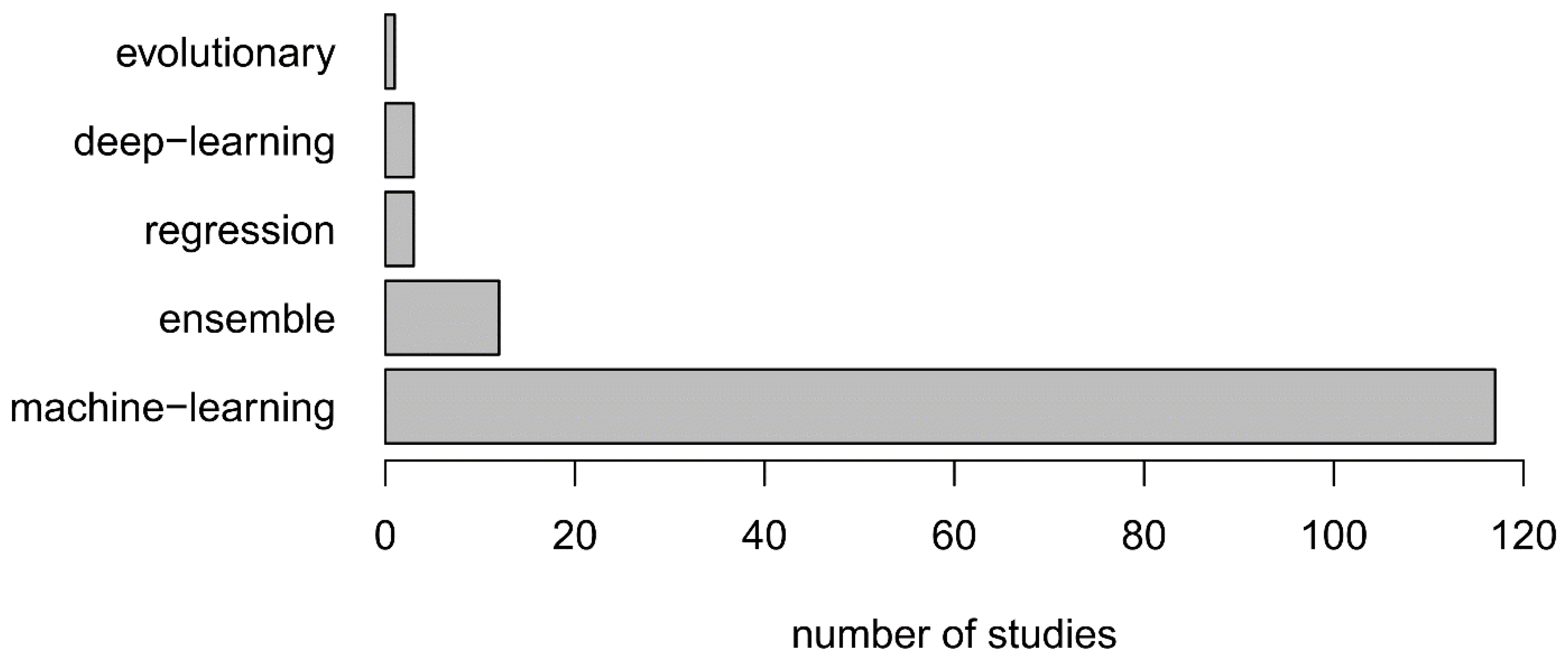
4.1. Classifier Selection
4.2. Tools and Computer Resources
4.3. Variable Selection
4.4. Background vs. (Pseudo-)Absence Points
- Confusion over the use of background and pseudo-absence points still occurs (Table 6): 42% of studies based on “presence-only” classifiers mentioned the use of pseudo-absence or absence points. Conversely, and to a lesser extent, 7% of the studies based on presence/absence classifiers used background points. This confusion occurred mainly in “ensemble classifiers” that combined presence-only and presence/absence OCC.
- The number of randomly generated background points varied greatly, from 200 points [45] to 50,000 points [115]. Most often (n = 49), the number of background points was unspecified and thus probably defaulted to 10,000. Conversely, many studies explicitly mentioned this default value (n = 26). A few studies used less than 10,000 points (to decrease calculation times [85]) (n = 9) or more than 10,000 (n = 4).
- Spatial bias was rarely assumed when generating background points (n = 10).
- Background points were usually selected over the entire landscape (n = 29), although most studies (n = 51) did not specify this. Seven studies selected background points only in natural areas.
- The selection of background points rarely excluded the areas with field vegetation plots (n = 6).
4.5. Classifier Fitting
4.6. Thresholding
- Maximum F-score [50];
4.7. Assessing Classification Accuracy
4.8. Combining One-Class Classifiers
5. Conclusions and Recommendations
- Mapping of potential and actual vegetation areas: Using spectral variables derived from high spatial resolution RS data in OCCs enables classifying potential and actual vegetation areas, which provides new insights into the quantification of species diversity, ecosystem restoration (identification of suitable areas), or the control of invasive species (identification of vulnerable areas).
- Long-term monitoring of vegetation: The use of RS databases, such as Landsat archives (available since 1972) or historical aerial photographs (available since the beginning of the XX century), enables temporal monitoring of vegetation classes over many decades.
- Generation of multiple ecological variables: A wide range of ecological variables can be derived from RS data available at the global scale, including vegetation (phenology, physiognomy, height …), topography, and LULC, as well as climate (precipitation, temperature …), soil (physical and chemical properties), and disturbances (fires, flooding …). These variables can be combined to increase OCC performance.
- Availability of open-source tools and open-access databases: Many innovative open-source tools, software, as well as archives of RS data and derived variables are freely available, which provide access to the most recent advances in OCCs by a larger user community. Future studies could focus on wider use of cloud computing and development of open-source software to pre-process RS data. For example, Sentinel data can be pre-processed with the Sentinel Application Platform (SNAP) [150], provided by the European Space Agency using the ESA RSS Cloud Toolbox service [151].
- Reduction in plotting effort: A significant advantage of OCCs, compared to traditional multi-class classifiers, is to restrict to plotting the vegetation classes of interest. Using UAV images, plots can be generated automatically using artificial intelligence, such as deep learning.
- Quantification of over-detection: Use of very high spatial resolution RS data enables collection of absence plots (e.g., impervious areas, waterbodies, or crops) to quantify more objectively over-detection (i.e., producer’s accuracy) on natural vegetation maps derived from presence-only OCC.
- Increasing the visibility and use of available RS variables: Variables derived from the RS data remain under-used in OCCs. To increase their visibility and thus their application in the ecology community, it is crucial—whenever possible—to relate RS variable (e.g., vegetation, topography, bioclimate, soil, LULC, disturbance) to essential climate or biodiversity variables [23], such as phenology, ecosystem vertical profile, or soil moisture.
- Following good classification practices: The performance of OCCs depends largely on good classification practices. In particular, it involves: (i) removing correlated variables; (ii) fitting all classification parameters and prioritizing transferability over performance; (iii) using background points and absence points for presence-only and presence/absence classifiers, respectively; (iv) limiting thresholding; (v) correcting spatial sampling biases; (vi) validating classifications statistically and spatially with independent and non-spatially autocorrelated field plots [152]; and (vii) discussing the influence of the quality of RS variables and field plots on OCC performance.
- Bridging the gap between spatial resolution and site extent: Future studies could focus on applying OCCs at national, continental, or global scales using high or very high spatial resolution RS data. This could be done by combining advances in classification algorithms (e.g., convolutional neural network) with growing databases (e.g., citizen science data [153]) and enhanced computing ability (e.g., cloud computing).
- Classifying plant communities: Although plant communities are rarely classified, mapping them is indicative of the conservation status of natural habitats [154]. To this end, developing harmonized databases such as the European vegetation archive [155] is crucial to providing field plots for OCC fitting and validation.
- Improve quality of RS-based variables: Microwave remote sensors, such as SAR or emerging global navigation satellite system reflectometry data, could be more broadly used in OCCs, since they have great potential for characterizing vegetation structure (e.g., volume) and for monitoring ecosystem disturbances (e.g., flooding, snow cover, fires, soil moisture) at higher spatio-temporal resolutions. Moreover, using climate variables with higher spatial resolution in OCCs generated from LiDAR [156] and Sentinel-3 data also appears promising.
- Classify time first, space later: Traditionally, vegetation classes were monitored over time by annual change detection, which may be due to real vegetation dynamics but also to multiple errors generated by each annual classification. Future studies could focus on “time-first, space-later” approach that examines inter-annual NDVI profile rather than each annual NDVI profile independently [62].
- Improve classifier transferability: Classifier transferability in space and time is a major challenge for vegetation mapping due to phenological variations in space and time. Using algorithms based on optimal transport, which is a robust probabilistic and geometric tool for comparing the similarity between two distributions [157], into OCCs seems promising. In addition, the AIC index could also be integrated into OCC tools more widely to fit classifier based on their transferability rather than their accuracy.
- Connect artificial intelligence to ecological expertise: Although it is interesting to use artificial intelligence (e.g., deep learning, data mining) to map vegetation, the ecological community is concerned with the “black box” issue and stresses the need to understand relationships (i.e., transparency, interpretability, and explanation) between classifier functioning and ecological processes, e.g., using videos based on RS time-series to highlight vegetation dynamics [158]. Moreover, future studies could also focus on the development of dynamic classifiers that establish a strong relationship between environmental variables and ecological processes [17].
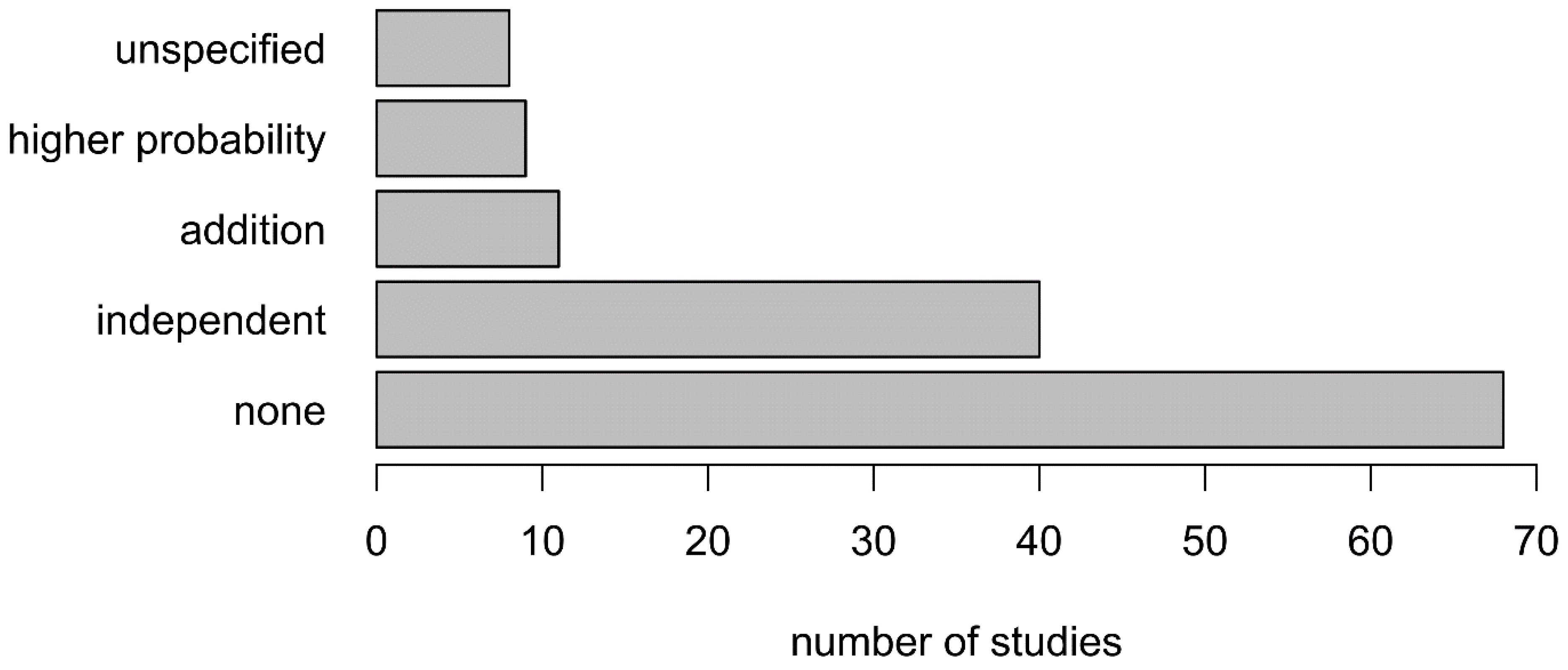
- Develop a method to combine one-class classifiers: Although most studies involve several vegetation classes, they are rarely combined in the same map given the requirement to select, for each OCC, the same variables and ratio of presence/absence points [7]. It thus seems necessary to develop a generic methodological framework to combine multiple one-class classifiers.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Pedrotti, F. Plant and Vegetation Mapping; Springer: Berlin, Germany, 2013. [Google Scholar]
- Corbane, C.; Lang, S.; Pipkins, K.; Alleaume, S.; Deshayes, M.; Millán, V.E.G.; Strasser, T.; Borre, J.V.; Toon, S.; Michael, F. Remote sensing for mapping natural habitats and their conservation status—New opportunities and challenges. Int. J. Appl. Earth Obs. Geoinf. 2015, 37, 7–16. [Google Scholar] [CrossRef]
- Gobeyn, S.; Mouton, A.M.; Cord, A.F.; Kaim, A.; Volk, M.; Goethals, P.L.M. Evolutionary Algorithms for Species Distribution Modelling: A Review in the Context of Machine Learning. Ecol. Model. 2019, 392, 179–195. [Google Scholar] [CrossRef]
- Miller, J. Species Distribution Modeling. Geogr. Compass 2010, 4, 490–509. [Google Scholar] [CrossRef]
- Álvarez-Martínez, J.M.; Jiménez-Alfaro, B.; Barquín, J.; Ondiviela, B.; Recio, M.; Silió-Calzada, A.; Juanes, J.A. Modelling the Area of Occupancy of Habitat Types with Remote Sensing. Methods Ecol. Evol. 2018, 9, 580–593. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.J.; Iglesias, T.L. Evaluating Presence-only Species Distribution Models with Discrimination Accuracy Is Uninformative for Many Applications. J. Biogeogr. 2020, 47, 167–180. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping Species Distributions with MAXENT Using a Geographically Biased Sample of Presence Data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s Parameter Configuration and Small Samples: Are We Paying Attention to Recommendations? A Systematic Review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef]
- Scherrer, D.; Mod, H.K.; Guisan, A. How to Evaluate Community Predictions without Thresholding? Methods Ecol. Evol. 2020, 11, 51–63. [Google Scholar] [CrossRef]
- Moudrý, V.; Lecours, V.; Malavasi, M.; Misiuk, B.; Gábor, L.; Gdulová, K.; Šímová, P.; Wild, J. Potential Pitfalls in Rescaling Digital Terrain Model-Derived Attributes for Ecological Studies. Ecol. Inform. 2019, 54, 100987. [Google Scholar] [CrossRef]
- Gábor, L.; Moudrý, V.; Barták, V.; Lecours, V. How Do Species and Data Characteristics Affect Species Distribution Models and When to Use Environmental Filtering? Int. J. Geogr. Inf. Sci. 2019, 1–18. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings Predict the Distribution of Species, or the Challenge of Selecting Environmental Predictors and Evaluation Statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Mod, H.K.; Scherrer, D.; Luoto, M.; Guisan, A. What We Use Is Not What We Know: Environmental Predictors in Plant Distribution Models. J. Veg. Sci. 2016, 27, 1308–1322. [Google Scholar] [CrossRef]
- Pradervand, J.-N.; Dubuis, A.; Pellissier, L.; Guisan, A.; Randin, C. Very High Resolution Environmental Predictors in Species Distribution Models: Moving beyond Topography? Prog. Phys. Geogr. Earth Environ. 2014, 38, 79–96. [Google Scholar] [CrossRef]
- Yates, K.L.; Bouchet, P.J.; Caley, M.J.; Mengersen, K.; Randin, C.F.; Parnell, S.; Fielding, A.H.; Bamford, A.J.; Ban, S.; Barbosa, A.M.; et al. Outstanding Challenges in the Transferability of Ecological Models. Trends Ecol. Evol. 2018, 33, 790–802. [Google Scholar] [CrossRef]
- Franklin, J.; Serra-Diaz, J.M.; Syphard, A.D.; Regan, H.M. Big Data for Forecasting the Impacts of Global Change on Plant Communities. Glob. Ecol. Biogeogr. 2017, 26, 6–17. [Google Scholar] [CrossRef]
- Schrodt, F.; Santos, M.J.; Bailey, J.J.; Field, R. Challenges and Opportunities for Biogeography-What Can We Still Learn from von Humboldt? J. Biogeogr. 2019, 46, 1631–1642. [Google Scholar] [CrossRef]
- Petrou, Z.I.; Manakos, I.; Stathaki, T. Remote Sensing for Biodiversity Monitoring: A Review of Methods for Biodiversity Indicator Extraction and Assessment of Progress towards International Targets. Biodivers. Conserv. 2015, 24, 2333–2363. [Google Scholar] [CrossRef]
- Duputie, A.; Zimmermann, N.E.; Chuine, I. Where Are the Wild Things? Why We Need Better Data on Species Distribution. Glob. Ecol. Biogeogr. 2014, 23, 457–467. [Google Scholar] [CrossRef]
- Morán-Ordóñez, A.; Suárez-Seoane, S.; Elith, J.; Calvo, L.; de Luis, E. Satellite Surface Reflectance Improves Habitat Distribution Mapping: A Case Study on Heath and Shrub Formations in the Cantabrian Mountains (NW Spain). Divers. Distrib. 2012, 18, 588–602. [Google Scholar] [CrossRef]
- Leitão, P.J.; Santos, M.J. Improving Models of Species Ecological Niches: A Remote Sensing Overview. Front. Ecol. Evol. 2019, 7, 9. [Google Scholar] [CrossRef]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.-N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will Remote Sensing Shape the next Generation of Species Distribution Models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef]
- Schulte to Bühne, H.; Pettorelli, N. Better Together: Integrating and Fusing Multispectral and Radar Satellite Imagery to Inform Biodiversity Monitoring, Ecological Research and Conservation Science. Methods Ecol. Evol. 2018, 9, 849–865. [Google Scholar] [CrossRef]
- Randin, C.F.; Ashcroft, M.B.; Bolliger, J.; Cavender-Bares, J.; Coops, N.C.; Dullinger, S.; Dirnböck, T.; Eckert, S.; Ellis, E.; Fernández, N.; et al. Monitoring Biodiversity in the Anthropocene Using Remote Sensing in Species Distribution Models. Remote Sens. Environ. 2020, 239, 111626. [Google Scholar] [CrossRef]
- Pettorelli, N.; Wegmann, M.; Skidmore, A.; Mücher, S.; Dawson, T.P.; Fernandez, M.; Lucas, R.; Schaepman, M.E.; Wang, T.; O’Connor, B.; et al. Framing the Concept of Satellite Remote Sensing Essential Biodiversity Variables: Challenges and Future Directions. Remote Sens. Ecol. Conserv. 2016, 2, 122–131. [Google Scholar] [CrossRef]
- Bradley, B.A.; Olsson, A.D.; Wang, O.; Dickson, B.G.; Pelech, L.; Sesnie, S.E.; Zachmann, L.J. Species Detection vs. Habitat Suitability: Are We Biasing Habitat Suitability Models with Remotely Sensed Data? Ecol. Model. 2012, 244, 57–64. [Google Scholar] [CrossRef]
- Cord, A.F.; Meentemeyer, R.K.; Leitão, P.J.; Václavík, T. Modelling Species Distributions with Remote Sensing Data: Bridging Disciplinary Perspectives. J. Biogeogr. 2013, 40, 2226–2227. [Google Scholar] [CrossRef]
- Girma, A.; de Bie, C.A.J.M.; Skidmore, A.K.; Venus, V.; Bongers, F. Hyper-Temporal SPOT-NDVI Dataset Parameterization Captures Species Distributions. Int. J. Geogr. Inf. Sci. 2016, 30, 89–107. [Google Scholar] [CrossRef]
- Wüest, R.O.; Bergamini, A.; Bollmann, K.; Baltensweiler, A. LiDAR Data as a Proxy for Light Availability Improve Distribution Modelling of Woody Species. For. Ecol. Manag. 2020, 456, 117644. [Google Scholar] [CrossRef]
- José-Silva, L.; dos Santos, R.C.; de Lima, B.M.; Lima, M.; de Oliveira-Júnior, J.F.; Teodoro, P.E.; Eisenlohr, P.V.; da Silva Junior, C.A. Improving the Validation of Ecological Niche Models with Remote Sensing Analysis. Ecol. Model. 2018, 380, 22–30. [Google Scholar] [CrossRef]
- Long, A.L.; Kettenring, K.M.; Hawkins, C.P.; Neale, C.M.U. Distribution and Drivers of a Widespread, Invasive Wetland Grass, Phragmites Australis, in Wetlands of the Great Salt Lake, Utah, USA. Wetlands 2017, 37, 45–57. [Google Scholar] [CrossRef]
- Diao, C.; Wang, L. Development of an Invasive Species Distribution Model with Fine-Resolution Remote Sensing. Int. J. Appl. Earth Obs. Geoinf. 2014, 30, 65–75. [Google Scholar] [CrossRef]
- Pouteau, R.; Meyer, J.-Y.; Larrue, S. Using Range Filling Rather than Prevalence of Invasive Plant Species for Management Prioritisation: The Case of Spathodea Campanulata in the Society Islands (South Pacific). Ecol. Indic. 2015, 54, 87–95. [Google Scholar] [CrossRef]
- Shiferaw, H.; Bewket, W.; Eckert, S. Performances of Machine Learning Algorithms for Mapping Fractional Cover of an Invasive Plant Species in a Dryland Ecosystem. Ecol. Evol. 2019, 9, 2562–2574. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Mir, A.H.; Upadhaya, K.; Iralu, V.; Roy, D.K. Abundance and Habitat-Suitability Relationship Deteriorate in Fragmented Forest Landscapes: A Case of Adinandra Griffithii Dyer, a Threatened Endemic Tree from Meghalaya in Northeast India. Ecol. Process. 2018, 7, 3. [Google Scholar] [CrossRef]
- Aguilar-Soto, V.; Melgoza-Castillo, A.; Villarreal-Guerrero, F.; Wehenkel, C.; Pinedo-Alvarez, C. Modeling the Potential Distribution of Picea Chihuahuana Martínez, an Endangered Species at the Sierra Madre Occidental, Mexico. Forests 2015, 6, 692–707. [Google Scholar] [CrossRef]
- Gonçalves, J.; Alves, P.; Pôças, I.; Marcos, B.; Sousa-Silva, R.; Lomba, Â.; Honrado, J.P. Exploring the Spatiotemporal Dynamics of Habitat Suitability to Improve Conservation Management of a Vulnerable Plant Species. Biodivers. Conserv. 2016, 25, 2867–2888. [Google Scholar] [CrossRef]
- Chen, X.; Yin, D.; Chen, J.; Cao, X. Effect of Training Strategy for Positive and Unlabelled Learning Classification: Test on Landsat Imagery. Remote Sens. Lett. 2016, 7, 1063–1072. [Google Scholar] [CrossRef]
- Deng, X.; Li, W.; Liu, X.; Guo, Q.; Newsam, S. One-Class Remote Sensing Classification: One-Class vs. Binary Classifiers. Int. J. Remote Sens. 2018, 39, 1890–1910. [Google Scholar] [CrossRef]
- Fernandez, I.C.; Morales, N.S. One-Class Land-Cover Classification Using MaxEnt: The Effect of Modelling Parameterization on Classification Accuracy. PeerJ 2019, 7, e7016. [Google Scholar] [CrossRef]
- Mack, B.; Roscher, R.; Waske, B. Can I Trust My One-Class Classification? Remote Sens. 2014, 6, 8779–8802. [Google Scholar] [CrossRef]
- Mack, B.; Waske, B. In-Depth Comparisons of MaxEnt, Biased SVM and One-Class SVM for One-Class Classification of Remote Sensing Data. Remote Sens. Lett. 2017, 8, 290–299. [Google Scholar] [CrossRef]
- Araya-López, R.A.; Lopatin, J.; Fassnacht, F.E.; Hernández, H.J. Monitoring Andean High Altitude Wetlands in Central Chile with Seasonal Optical Data: A Comparison between Worldview-2 and Sentinel-2 Imagery. ISPRS J. Photogramm. Remote Sens. 2018, 145, 213–224. [Google Scholar] [CrossRef]
- Chignell, S.M.; Luizza, M.W.; Skach, S.; Young, N.E.; Evangelista, P.H. An Integrative Modeling Approach to Mapping Wetlands and Riparian Areas in a Heterogeneous Rocky Mountain Watershed. Remote Sens. Ecol. Conserv. 2018, 4, 150–165. [Google Scholar] [CrossRef]
- Räsänen, A.; Elsakov, V.; Virtanen, T. Usability of One-Class Classification in Mapping and Detecting Changes in Bare Peat Surfaces in the Tundra. Int. J. Remote Sens. 2019, 40, 4083–4103. [Google Scholar] [CrossRef]
- Prins, E. Landsat Approaches to Map Agro-Pastoral Farming in the Wetlands of Southern Sudan. Int. J. Remote Sens. 2018, 39, 854–878. [Google Scholar] [CrossRef]
- Bradter, U.; O’Connell, J.; Kunin, W.E.; Boffey, C.W.H.; Ellis, R.J.; Benton, T.G. Classifying Grass-Dominated Habitats from Remotely Sensed Data: The Influence of Spectral Resolution, Acquisition Time and the Vegetation Classification System on Accuracy and Thematic Resolution. Sci. Total Environ. 2020, 711, 134584. [Google Scholar] [CrossRef]
- Fenske, K.; Feilhauer, H.; Förster, M.; Stellmes, M.; Waske, B. Hierarchical Classification with Subsequent Aggregation of Heathland Habitats Using an Intra-Annual RapidEye Time-Series. Int. J. Appl. Earth Obs. Geoinf. 2020, 87, 102036. [Google Scholar] [CrossRef]
- Mack, B.; Roscher, R.; Stenzel, S.; Feilhauer, H.; Schmidtlein, S.; Waske, B. Mapping Raised Bogs with an Iterative One-Class Classification Approach. ISPRS J. Photogramm. Remote Sens. 2016, 120, 53–64. [Google Scholar] [CrossRef]
- Schwager, P.; Berg, C. Global Warming Threatens Conservation Status of Alpine EU Habitat Types in the European Eastern Alps. Reg. Environ. Chang. 2019, 19, 2411–2421. [Google Scholar] [CrossRef]
- Stenzel, S.; Feilhauer, H.; Mack, B.; Metz, A.; Schmidtlein, S. Remote Sensing of Scattered Natura 2000 Habitats Using a One-Class Classifier. Int. J. Appl. Earth Obs. Geoinf. 2014, 33, 211–217. [Google Scholar] [CrossRef]
- Suárez-Seoane, S.; Jiménez-Alfaro, B.; Obeso, J.R. Habitat-Partitioning Improves Regional Distribution Models in Multi-Habitat Species: A Case Study with the European Bilberry. Biodivers. Conserv. 2020, 29, 987–1008. [Google Scholar] [CrossRef]
- Connor, T.; Hull, V.; Vina, A.; Shortridge, A.; Tang, Y.; Zhang, J.; Wang, F.; Liu, J. Effects of Grain Size and Niche Breadth on Species Distribution Modeling. Ecography 2018, 41, 1270–1282. [Google Scholar] [CrossRef]
- Tang, Y.; Winkler, J.A.; Vina, A.; Wang, F.; Zhang, J.; Zhao, Z.; Connor, T.; Yang, H.; Zhang, Y.; Zhang, X.; et al. Expanding Ensembles of Species Present-Day and Future Climatic Suitability to Consider the Limitations of Species Occurrence Data. Ecol. Indic. 2020, 110, 105891. [Google Scholar] [CrossRef]
- Anderson, C.B. Biodiversity Monitoring, Earth Observations and the Ecology of Scale. Ecol. Lett. 2018, 21, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, G.-Y.; Do, Y.; Park, H.-S.; Joo, G.-J. Relative Importance of Hydrological Variables in Predicting the Habitat Suitability of Euryale Ferox Salisb. J. Plant Ecol. 2018, 11, 169–179. [Google Scholar] [CrossRef]
- Doninck, J.V.; Jones, M.M.; Zuquim, G.; Ruokolainen, K.; Moulatlet, G.M.; Sirén, A.; Cárdenas, G.; Lehtonen, S.; Tuomisto, H. Multispectral Canopy Reflectance Improves Spatial Distribution Models of Amazonian Understory Species. Ecography 2020, 43, 128–137. [Google Scholar] [CrossRef]
- Hengl, T.; Walsh, M.G.; Sanderman, J.; Wheeler, I.; Harrison, S.P.; Prentice, I.C. Global Mapping of Potential Natural Vegetation: An Assessment of Machine Learning Algorithms for Estimating Land Potential. PeerJ 2018, 6, e5457. [Google Scholar] [CrossRef]
- Rocchini, D. Seeing the Unseen by Remote Sensing: Satellite Imagery Applied to Species Distribution Modelling. J. Veg. Sci. 2013, 24, 209–210. [Google Scholar] [CrossRef]
- Picoli, M.C.A.; Camara, G.; Sanches, I.; Simões, R.; Carvalho, A.; Maciel, A.; Coutinho, A.; Esquerdo, J.; Antunes, J.; Begotti, R.A. Big Earth Observation Time Series Analysis for Monitoring Brazilian Agriculture. ISPRS J. Photogramm. Remote Sens. 2018, 145, 328–339. [Google Scholar] [CrossRef]
- Amici, V.; Marcantonio, M.; La Porta, N.; Rocchini, D. A Multi-Temporal Approach in MaxEnt Modelling: A New Frontier for Land Use/Land Cover Change Detection. Ecol. Inform. 2017, 40, 40–49. [Google Scholar] [CrossRef]
- Rebelo, A.J.; Scheunders, P.; Esler, K.J.; Meire, P. Detecting, Mapping and Classifying Wetland Fragments at a Landscape Scale. Remote Sens. Appl. Soc. Environ. 2017, 8, 212–223. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Regos, A.; Gonçalves, J.F.; Alcaraz-Segura, D.; Honrado, J. Remotely Sensed Variables of Ecosystem Functioning Support Robust Predictions of Abundance Patterns for Rare Species. Remote Sens. 2019, 11, 2086. [Google Scholar] [CrossRef]
- Carlson, B.Z.; Georges, D.; Rabatel, A.; Randin, C.F.; Renaud, J.; Delestrade, A.; Zimmermann, N.E.; Choler, P.; Thuiller, W. Accounting for Tree Line Shift, Glacier Retreat and Primary Succession in Mountain Plant Distribution Models. Divers. Distrib. 2014, 20, 1379–1391. [Google Scholar] [CrossRef]
- Ramachandran, R.M.; Roy, P.S.; Chakravarthi, V.; Sanjay, J.; Joshi, P.K. Long-Term Land Use and Land Cover Changes (1920–2015) in Eastern Ghats, India: Pattern of Dynamics and Challenges in Plant Species Conservation. Ecol. Indic. 2018, 85, 21–36. [Google Scholar] [CrossRef]
- Keshtkar, H.; Voigt, W. Potential Impacts of Climate and Landscape Fragmentation Changes on Plant Distributions: Coupling Multi-Temporal Satellite Imagery with GIS-Based Cellular Automata Model. Ecol. Inform. 2016, 32, 145–155. [Google Scholar] [CrossRef]
- Tredennick, A.T.; Hooten, M.B.; Aldridge, C.L.; Homer, C.G.; Kleinhesselink, A.R.; Adler, P.B. Forecasting Climate Change Impacts on Plant Populations over Large Spatial Extents. Ecosphere 2016, 7, e01525. [Google Scholar] [CrossRef]
- Vacchiano, G.; Motta, R. An Improved Species Distribution Model for Scots Pine and Downy Oak under Future Climate Change in the NW Italian Alps. Ann. For. Sci. 2015, 72. [Google Scholar] [CrossRef]
- Lastiri-Hernández, M.A.; Cruz-Cárdenas, G.; Álvarez-Bernal, D.; Vázquez-Sánchez, M.; Bermúdez-Torres, K. Ecological Niche Modeling for Halophyte Species with Possible Anthropogenic Use in Agricultural Saline Soils. Environ. Model. Assess. 2020. [Google Scholar] [CrossRef]
- Malahlela, O.E.; Adjorlolo, C.; Olwoch, J.M. Mapping the Spatial Distribution of Lippia javanica (Burm. f.) Spreng Using Sentinel-2 and SRTM-Derived Topographic Data in Malaria Endemic Environment. Ecol. Model. 2019, 392, 147–158. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C. Land-Cover Classification Using MaxEnt: Can We Trust in Model Quality Metrics for Estimating Classification Accuracy? Entropy 2020, 22, 342. [Google Scholar] [CrossRef] [PubMed]
- Delalay, M.; Tiwari, V.; Ziegler, A.D.; Gopal, V.; Passy, P. Land-Use and Land-Cover Classification Using Sentinel-2 Data and Machine-Learning Algorithms: Operational Method and Its Implementation for a Mountainous Area of Nepal. J. Appl. Remote Sens. 2019, 13, 014530. [Google Scholar] [CrossRef]
- Kattenborn, T.; Lopatin, J.; Förster, M.; Braun, A.C.; Fassnacht, F.E. UAV Data as Alternative to Field Sampling to Map Woody Invasive Species Based on Combined Sentinel-1 and Sentinel-2 Data. Remote Sens. Environ. 2019, 227, 61–73. [Google Scholar] [CrossRef]
- Alexandridis, T.K.; Tamouridou, A.A.; Pantazi, X.E.; Lagopodi, A.L.; Kashefi, J.; Ovakoglou, G.; Polychronos, V.; Moshou, D. Novelty Detection Classifiers in Weed Mapping: Silybum marianum Detection on UAV Multispectral Images. Sensors 2017, 17, 2007. [Google Scholar] [CrossRef] [PubMed]
- Kattenborn, T.; Eichel, J.; Fassnacht, F.E. Convolutional Neural Networks Enable Efficient, Accurate and Fine-Grained Segmentation of Plant Species and Communities from High-Resolution UAV Imagery. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Kattenborn, T.; Fassnacht, F.E. How Canopy Shadow Affects Invasive Plant Species Classification in High Spatial Resolution Remote Sensing. Remote Sens. Ecol. Conserv. 2019, 5, 302–317. [Google Scholar] [CrossRef]
- Hengl, T.; de Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global Gridded Soil Information Based on Machine Learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed]
- Vega, G.C.; Pertierra, L.R.; Olalla-Tarraga, M.A. Data Descriptor: MERRAclim, a High-Resolution Global Dataset of Remotely Sensed Bioclimatic Variables for Ecological Modelling. Sci. Data 2017, 4, 170078. [Google Scholar] [CrossRef]
- Gascoin, S.; Grizonnet, M.; Bouchet, M.; Salgues, G.; Hagolle, O. Theia Snow Collection: High-Resolution Operational Snow Cover Maps from Sentinel-2 and Landsat-8 Data. Earth Syst. Sci. Data 2019, 11, 493–514. [Google Scholar] [CrossRef]
- Cord, A.F.; Klein, D.; Gernandt, D.S.; de la Rosa, J.A.P.; Dech, S. Remote Sensing Data Can Improve Predictions of Species Richness by Stacked Species Distribution Models: A Case Study for Mexican Pines. J. Biogeogr. 2014, 41, 736–748. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field Validation of an Invasive Species Maxent Model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef]
- Judith, C.; Schneider, J.V.; Schmidt, M.; Ortega, R.; Gaviria, J.; Zizka, G. Using High-Resolution Remote Sensing Data for Habitat Suitability Models of Bromeliaceae in the City of Merida, Venezuela. Landsc. Urban Plan. 2013, 120, 107–118. [Google Scholar] [CrossRef]
- Skowronek, S.; Asner, G.P.; Feilhauer, H. Performance of One-Class Classifiers for Invasive Species Mapping Using Airborne Imaging Spectroscopy. Ecol. Inform. 2017, 37, 66–76. [Google Scholar] [CrossRef]
- Fedrigo, M.; Stewart, S.B.; Roxburgh, S.H.; Kasel, S.; Bennett, L.T.; Vickers, H.; Nitschke, C.R. Predictive Ecosystem Mapping of South-Eastern Australian Temperate Forests Using Lidar-Derived Structural Profiles and Species Distribution Models. Remote Sens. 2019, 11, 93. [Google Scholar] [CrossRef]
- Piiroinen, R.; Fassnacht, F.E.; Heiskanen, J.; Maeda, E.; Mack, B.; Pellikka, P. Invasive Tree Species Detection in the Eastern Arc Mountains Biodiversity Hotspot Using One Class Classification. Remote Sens. Environ. 2018, 218, 119–131. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Wan, Z. New Refinements and Validation of the Collection-6 MODIS Land-Surface Temperature/Emissivity Product. Remote Sens. Environ. 2014, 140, 36–45. [Google Scholar] [CrossRef]
- Metz, M.; Rocchini, D.; Neteler, M. Surface Temperatures at the Continental Scale: Tracking Changes with Remote Sensing at Unprecedented Detail. Remote Sens. 2014, 6, 3822–3840. [Google Scholar] [CrossRef]
- Deblauwe, V.; Droissart, V.; Bose, R.; Sonké, B.; Blach-Overgaard, A.; Svenning, J.-C.; Wieringa, J.J.; Ramesh, B.R.; Stévart, T.; Couvreur, T.L.P. Remotely Sensed Temperature and Precipitation Data Improve Species Distribution Modelling in the Tropics. Glob. Ecol. Biogeogr. 2016, 25, 443–454. [Google Scholar] [CrossRef]
- Shiferaw, H.; Schaffner, U.; Bewket, W.; Alamirew, T.; Zeleke, G.; Teketay, D.; Eckert, S. Modelling the Current Fractional Cover of an Invasive Alien Plant and Drivers of Its Invasion in a Dryland Ecosystem. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Lembrechts, J.; Lenoir, J.; Roth, N.; Hattab, T.; Milbau, A.; Haider, S.; Pellissier, L.; Pauchard, A.; Backes, A.R.; Dimarco, R.D.; et al. Comparing Temperature Data Sources for Use in Species Distribution Models: From in-Situ Logging to Remote Sensing. Glob. Ecol. Biogeogr. 2019, 28, 1578–1596. [Google Scholar] [CrossRef]
- Bazzichetto, M.; Malavasi, M.; Bartak, V.; Acosta, A.T.R.; Moudry, V.; Carranza, M.L. Modeling Plant Invasion on Mediterranean Coastal Landscapes: An Integrative Approach Using Remotely Sensed Data. Landsc. Urban. Plan. 2018, 171, 98–106. [Google Scholar] [CrossRef]
- Campos, V.E.; Cappa, F.M.; Viviana, F.M.; Giannoni, S.M. Using Remotely Sensed Data to Model Suitable Habitats for Tree Species in a Desert Environment. J. Veg. Sci. 2016, 27, 200–210. [Google Scholar] [CrossRef]
- O’Neill, A.R. Evaluating High-Altitude Ramsar Wetlands in the Eastern Himalayas. Glob. Ecol. Conserv. 2019, 20, e00715. [Google Scholar] [CrossRef]
- Rahimian Boogar, A.; Salehi, H.; Pourghasemi, H.R.; Blaschke, T. Predicting Habitat Suitability and Conserving Juniperus Spp. Habitat Using SVM and Maximum Entropy Machine Learning Techniques. Water 2019, 11, 2049. [Google Scholar] [CrossRef]
- Buse, J.; Boch, S.; Hilgersd, J.; Griebeler, E.M. Conservation of Threatened Habitat Types under Future Climate Change—Lessons from Plant-Distribution Models and Current Extinction Trends in Southern Germany. J. Nat. Conserv. 2015, 27, 18–25. [Google Scholar] [CrossRef]
- McCartney, K.R.; Kumar, S.; Sing, S.E.; Ward, S.M. Using Invaded-Range Species Distribution Modeling to Estimate the Potential Distribution of Linaria Species and Their Hybrids in the US Northern Rockies. Invasive Plant Sci. Manag. 2019, 12, 97–111. [Google Scholar] [CrossRef]
- Malavasi, M.; Barták, V.; Jucker, T.; Acosta, A.T.R.; Carranza, M.L.; Bazzichetto, M. Strength in Numbers: Combining Multi-Source Remotely Sensed Data to Model Plant Invasions in Coastal Dune Ecosystems. Remote Sens. 2019, 11, 275. [Google Scholar] [CrossRef]
- Cord, A.F.; Klein, D.; Mora, F.; Dech, S. Comparing the Suitability of Classified Land Cover Data and Remote Sensing Variables for Modeling Distribution Patterns of Plants. Ecol. Model. 2014, 272, 129–140. [Google Scholar] [CrossRef]
- Duff, T.J.; Bell, T.L.; York, A. Recognising Fuzzy Vegetation Pattern: The Spatial Prediction of Floristically Defined Fuzzy Communities Using Species Distribution Modelling Methods. J. Veg. Sci. 2014, 25, 323–337. [Google Scholar] [CrossRef]
- Tuomisto, H.; Van Doninck, J.; Ruokolainen, K.; Moulatlet, G.M.; Figueiredo, F.O.G.; Siren, A.; Cardenas, G.; Lehtonen, S.; Zuquim, G. Discovering Floristic and Geoecological Gradients across Amazonia. J. Biogeogr. 2019, 46, 1734–1748. [Google Scholar] [CrossRef]
- Baumbach, L.; Niamir, A.; Hickler, T.; Yousefpour, R. Regional Adaptation of European Beech (Fagus sylvatica) to Drought in Central European Conditions Considering Environmental Suitability and Economic Implications. Reg. Environ. Chang. 2019, 19, 1159–1174. [Google Scholar] [CrossRef]
- Mudereri, B.T.; Abdel-Rahman, E.M.; Dube, T.; Landmann, T.; Khan, Z.; Kimathi, E.; Owino, R.; Niassy, S. Multi-Source Spatial Data-Based Invasion Risk Modeling of Striga (Striga asiatica) in Zimbabwe. GIScience Remote Sens. 2020, 57, 553–571. [Google Scholar] [CrossRef]
- Truong, T.T.A.; Hardy, G.E.S.J.; Andrew, M.E. Contemporary Remotely Sensed Data Products Refine Invasive Plants Risk Mapping in Data Poor Regions. Front. Plant Sci. 2017, 8, 770. [Google Scholar] [CrossRef] [PubMed]
- Bloom, T.D.S.; Flower, A.; Medler, M.; DeChaine, E.G. The Compounding Consequences of Wildfire and Climate Change for a High-Elevation Wildflower (Saxifraga austromontana). J. Biogeogr. 2018, 45, 2755–2765. [Google Scholar] [CrossRef]
- Niittynen, P.; Luoto, M. The Importance of Snow in Species Distribution Models of Arctic Vegetation. Ecography 2018, 41, 1024–1037. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using Species Distribution Models at Local Scale to Guide the Search of Poorly Known Species: Review, Methodological Issues and Future Directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Pottier, J.; Malenovský, Z.; Psomas, A.; Homolová, L.; Schaepman, M.E.; Choler, P.; Thuiller, W.; Guisan, A.; Zimmermann, N.E. Modelling Plant Species Distribution in Alpine Grasslands Using Airborne Imaging Spectroscopy. Biol. Lett. 2014, 10, 20140347. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Saintilan, N.; Yang, X.; Hunter, S.; Mawer, D. MODIS NDVI Based Metrics Improve Habitat Suitability Modelling in Fragmented Patchy Floodplains. Remote Sens. Appl. Soc. Environ. 2015, 1, 85–97. [Google Scholar] [CrossRef]
- Halmy, M.W.A.; Fawzy, M.; Ahmed, D.A.; Saeed, N.M.; Awad, M.A. Monitoring and Predicting the Potential Distribution of Alien Plant Species in Arid Ecosystem Using Remotely-Sensed Data. Remote Sens. Appl. Soc. Environ. 2019, 13, 69–84. [Google Scholar] [CrossRef]
- Moudrý, V.; Lecours, V.; Gdulová, K.; Gábor, L.; Moudrá, L.; Kropáček, J.; Wild, J. On the Use of Global DEMs in Ecological Modelling and the Accuracy of New Bare-Earth DEMs. Ecol. Model. 2018, 383, 3–9. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Pérez Chaves, P.; Ruokolainen, K.; Tuomisto, H. Using Remote Sensing to Model Tree Species Distribution in Peruvian Lowland Amazonia. Biotropica 2018, 50, 758–767. [Google Scholar] [CrossRef]
- Richard, K.; Abdel-Rahman, E.M.; Mohamed, S.A.; Ekesi, S.; Borgemeister, C.; Landmann, T. Importance of Remotely-Sensed Vegetation Variables for Predicting the Spatial Distribution of African Citrus Triozid (Trioza erytreae) in Kenya. ISPRS Int. J. Geo-Inf. 2018, 7, 429. [Google Scholar] [CrossRef]
- Tomlinson, S.; Lewandrowski, W.; Elliott, C.P.; Miller, B.P.; Turner, S.R. High-resolution Distribution Modeling of a Threatened Short-range Endemic Plant Informed by Edaphic Factors. Ecol. Evol. 2020, 10, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Title, P.O.; Bemmels, J.B. ENVIREM: An Expanded Set of Bioclimatic and Topographic Variables Increases Flexibility and Improves Performance of Ecological Niche Modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Srivastava, V.; Griess, V.C.; Padalia, H. Mapping Invasion Potential Using Ensemble Modelling. A Case Study on Yushania Maling in the Darjeeling Himalayas. Ecol. Model. 2018, 385, 35–44. [Google Scholar] [CrossRef]
- Kattenborn, T.; Eichel, J.; Wiser, S.; Burrows, L.; Fassnacht, F.E.; Schmidtlein, S. Convolutional Neural Networks Accurately Predict Cover Fractions of Plant Species and Communities in Unmanned Aerial Vehicle Imagery. Remote Sens. Ecol. Conserv. 2020, 6, 472–486. [Google Scholar] [CrossRef]
- Wagner, F.H.; Sanchez, A.; Tarabalka, Y.; Lotte, R.G.; Ferreira, M.P.; Aidar, M.P.M.; Gloor, E.; Phillips, O.L.; Aragão, L.E.O.C. Using the U-Net Convolutional Network to Map Forest Types and Disturbance in the Atlantic Rainforest with Very High Resolution Images. Remote Sens. Ecol. Conserv. 2019, 5, 360–375. [Google Scholar] [CrossRef]
- Rocchini, D.; Petras, V.; Petrasova, A.; Horning, N.; Furtkevicova, L.; Neteler, M.; Leutner, B.; Wegmann, M. Open Data and Open Source for Remote Sensing Training in Ecology. Ecol. Inform. 2017, 40, 57–61. [Google Scholar] [CrossRef]
- Guo, H.; Liu, Z.; Jiang, H.; Wang, C.; Liu, J.; Liang, D. Big Earth Data: A New Challenge and Opportunity for Digital Earth’s Development. Int. J. Digit. Earth 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Package ‘Dismo’. Circles 2017, 9, 1–68. [Google Scholar]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM Eval: An R Package for Conducting Spatially Independent Evaluations and Estimating Optimal Model Complexity for Maxent Ecological Niche Models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Evans, S.J.; Murphy, M.A.; Ram, K. SpatialEco. Package Version 1.1-0. 2018. Available online: https://cran.rproject.org/web/packages/spatialEco/index.html (accessed on 15 March 2021).
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A Platform for Ensemble Forecasting of Species Distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the Black Box: An Open-source Release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Adepoju, K.A.; Adelabu, S.A. Improving Accuracy Evaluation of Landsat-8 OLI Using Image Composite and Multisource Data with Google Earth Engine. Remote Sens. Lett. 2020, 11, 107–116. [Google Scholar] [CrossRef]
- Iannella, M.; De Simone, W.; D’Alessandro, P.; Console, G.; Biondi, M. Investigating the Current and Future Co-Occurrence of Ambrosia Artemisiifolia and Ophraella Communa in Europe through Ecological Modelling and Remote Sensing Data Analysis. Int. J. Environ. Res. Public Health 2019, 16, 3416. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Varia, J.; Mathew, S. Overview of Amazon Web Services; Amazon: Seattle, WA, USA, 2014; pp. 1–22. [Google Scholar]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Maxent Is Not a Presence-Absence Method: A Comment on Thibaud et al. Methods Ecol. Evol. 2014, 5, 1192–1197. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample Selection Bias and Presence-only Distribution Models: Implications for Background and Pseudo-absence Data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Vollering, J.; Halvorsen, R.; Auestad, I.; Rydgren, K. Bunching up the Background Betters Bias in Species Distribution Models. Ecography 2019, 42, 1717–1727. [Google Scholar] [CrossRef]
- Skowronek, S.; Van De Kerchove, R.; Rombouts, B.; Aerts, R.; Ewald, M.; Warrie, J.; Schiefer, F.; Garzon-Lopez, C.; Hattab, T.; Honnay, O.; et al. Transferability of Species Distribution Models for the Detection of an Invasive Alien Bryophyte Using Imaging Spectroscopy Data. Int. J. Appl. Earth Obs. Geoinf. 2018, 68, 61–72. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Gong, H.; Lin, Z.; Lv, S. Appling the One-Class Classification Method of Maxent to Detect an Invasive Plant Spartina Alterniflora with Time-Series Analysis. Remote Sens. 2017, 9, 1120. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E.; Kellner, J.R.; Wright, S.J. Operational Tree Species Mapping in a Diverse Tropical Forest with Airborne Imaging Spectroscopy. PLoS ONE 2015, 10, e0118403. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, W.; Liu, X.; Lu, X.; Li, T.; Guo, Q. An Ensemble of Classifiers Based on Positive and Unlabeled Data in One-Class Remote Sensing Classification. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 11, 572–584. [Google Scholar] [CrossRef]
- Vollering, J.; Halvorsen, R.; Mazzoni, S. The MIAmaxent R Package: Variable Transformation and Model Selection for Species Distribution Models. Ecol. Evol. 2019, 9, 12051–12068. [Google Scholar] [CrossRef]
- Yu, J.; Kang, S. Clustering-Based Proxy Measure for Optimizing One-Class Classifiers. Pattern Recognit. Lett. 2019, 117, 37–44. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting Thresholds for the Prediction of Species Occurrence with Presence-only Data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Cefalì, M.E.; Ballesteros, E.; Riera, J.L.; Chappuis, E.; Terradas, M.; Mariani, S.; Cebrian, E. The Optimal Sampling Design for Littoral Habitats Modelling: A Case Study from the North-Western Mediterranean. PLoS ONE 2018, 13, e0197234. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, S.; Fassnacht, F.E.; Mack, B.; Schmidtlein, S. Identification of High Nature Value Grassland with Remote Sensing and Minimal Field Data. Ecol. Indic. 2017, 74, 28–38. [Google Scholar] [CrossRef]
- Del Río, S.; Álvarez-Esteban, R.; Cano, E.; Pinto-Gomes, C.; Penas, Á. Potential Impacts of Climate Change on Habitat Suitability of Fagus sylvatica L. Forests in Spain. Plant Biosyst. 2018, 152, 1205–1213. [Google Scholar] [CrossRef]
- Ghareghan, F.; Ghanbarian, G.; Pourghasemi, H.R.; Safaeian, R. Prediction of Habitat Suitability of Morina persica L. Species Using Artificial Intelligence Techniques. Ecol. Indic. 2020, 112, 106096. [Google Scholar] [CrossRef]
- Fernandes, R.F.; Scherrer, D.; Guisan, A. Effects of Simulated Observation Errors on the Performance of Species Distribution Models. Divers. Distrib. 2019, 25, 400–413. [Google Scholar] [CrossRef]
- Tang, Y.; Winkler, J.A.; Vina, A.; Liu, J.; Zhang, Y.; Zhang, X.; Li, X.; Wang, F.; Zhang, J.; Zhao, Z. Uncertainty of Future Projections of Species Distributions in Mountainous Regions. PLoS ONE 2018, 13, e0189496. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Asner, G.P. Single-Species Detection with Airborne Imaging Spectroscopy Data: A Comparison of Support Vector Techniques. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 2501–2512. [Google Scholar] [CrossRef]
- European Space Agency Sentinel Application Platform (SNAP). Available online: https://step.esa.int/main/toolboxes/snap/ (accessed on 15 March 2021).
- European Space Agency ESA RSS Cloud Toolbox Service. Available online: https://eogrid.esrin.esa.int/cloudtoolbox/ (accessed on 15 March 2021).
- Valavi, R.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. BlockCV: An r Package for Generating Spatially or Environmentally Separated Folds for k-Fold Cross-Validation of Species Distribution Models. Methods Ecol. Evol. 2019, 10, 225–232. [Google Scholar] [CrossRef]
- Johnston, A.; Moran, N.; Musgrove, A.; Fink, D.; Baillie, S.R. Estimating Species Distributions from Spatially Biased Citizen Science Data. Ecol. Model. 2020, 422, 108927. [Google Scholar] [CrossRef]
- Delbosc, P.; Lagrange, I.; Rozo, C.; Bensettiti, F.; Bouzillé, J.-B.; Evans, D.; Lalanne, A.; Rapinel, S.; Bioret, F. Assessing the Conservation Status of Coastal Habitats under Article 17 of the EU Habitats Directive. Biol. Conserv. 2021, 254, 108935. [Google Scholar] [CrossRef]
- Chytrý, M.; Hennekens, S.M.; Jiménez-Alfaro, B.; Knollová, I.; Dengler, J.; Jansen, F.; Landucci, F.; Schaminée, J.H.; Aćić, S.; Agrillo, E. European Vegetation Archive (EVA): An Integrated Database of European Vegetation Plots. Appl. Veg. Sci. 2016, 19, 173–180. [Google Scholar] [CrossRef]
- Lembrechts, O.; Nijs, I.; Lenoir, J. Incorporating Microclimate into Species Distribution Models. Ecography 2019, 42, 1267–1279. [Google Scholar] [CrossRef]
- Courty, N.; Flamary, R.; Tuia, D.; Rakotomamonjy, A. Optimal Transport for Domain Adaptation. IEEE Trans. Pattern Anal. Mach. Intell. 2016. [Google Scholar] [CrossRef] [PubMed]
- Roscher, R.; Bohn, B.; Duarte, M.F.; Garcke, J. Explainable Machine Learning for Scientific Insights and Discoveries. IEEE Access 2020, 8, 42200–42216. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topics | |||
|---|---|---|---|
| Remote Sensing | One-Class Classifier | Natural Vegetations | |
| --------------------------------------- AND --------------------------------------------- | |||
 | remote * sens * | one-class classifier | plant * |
| remote-sens * | one class classifier | vegetation | |
| Earth observation | one-class model * | land cover | |
| satellite | one class model * | land-cover | |
| imagery | OCSVM | land-use | |
| orthophoto | OC-SVM | land use | |
| UAV | OCC | LULC | |
| drone | biased-SVM | landscape | |
| unmanned aerial | biased SVM | habitat | |
| aerial image * | GLM | alliance | |
| aircraft * | Generalized Linear Model | phytosociology | |
| airborne | AUC | ecosystems | |
| air-borne | TSS | Natura 2000 | |
| spaceborne | AIC | N2000 | |
| space-borne | SDM | EUNIS | |
| hyperspectral | S-SDM | functional traits | |
| hyper-spectral | Species distribution model * | dryland | |
| multispectral | Species classification | wetland | |
| multi-spectral | Maxent | peat * | |
| SAR | bog | ||
| radar | scrub * | ||
| LiDAR | shrub * | ||
| NDVI | heath * | ||
| MODIS | grass * | ||
| TRMM | meadow | ||
| Landsat * | tree * | ||
| Sentinel * | forest | ||
| Rapideye | wood * | ||
| ALOS * | phenology | ||
| RADARSAT * | biomass | ||
| SPOT * | biotope | ||
| SRTM | biome | ||
| Land Surface Temperature | conservation | ||
| ASTER | environment | ||
| Google Earth * | |||
| GEE | |||
| reflectance | |||
| Topic | Attribute | Categories |
|---|---|---|
| A. Ecological considerations | A1. Area of occupancy | Potential area; actual area |
| A2. Hierarchical level | Land cover; plant community; plant species | |
| A3. Number of classes | One class; multi-class | |
| A4. Site extent | Local (<1000 km2); regional (1000–100,000 km2); national; continental; global | |
| A5. Temporal monitoring | No; yes | |
| B. Remote sensing data and variables | B1. Sensor type | Multispectral; hyperspectral; SAR; LiDAR |
| B2. Platform type | Unmanned airborne; manned airborne; satellite | |
| B3. Spatial resolution | very high spatial resolution (<5 m); high spatial resolution (6–30 m); moderate spatial resolution (31–250 m); low spatial resolution (>251 m) | |
| B4. Acquisition frequency | Single; multi-temporal; annual time-series | |
| B5. Type of remote-sensing-based variable | Climate; soil; topography; vegetation; categorical; disturbance | |
| C. Reference data | C1. Reference type | Vegetation database; field collection; image collection |
| C2. Reference quantity | Very small (<10); small (10–50); moderate (50–100); large (100–1000); very large (>1000) | |
| D. Tools and computer Resources | D1. Software license | Open-source; commercial |
| D2. Computer location | Local computing; cloud computing | |
| E. Variable selection and collinearity | E1. Initial variable selection | Expert; literature; data mining |
| E2. Consideration of correlated variables | Classifier-based; expert-based; statistical-based; reduction | |
| F. Classifier selection | F1. Classifier type | Machine learning; ensemble classifier; deep learning; evolutionary algorithm |
| G. Background point selection | G1. Background point quantity | Small (<10,000); default (10,000); large (>10,000); unspecified; not applicable |
| G2. Consideration of spatial sampling | No; yes; not applicable | |
| G3. Area for background point selection | All landscapes; distribution area; natural areas; artificial areas; unspecified; not applicable | |
| G4. Consideration of area of occurrence | No; yes; not applicable | |
| H. Classifier fitting | H1. Type of classifier fitting | None; best performance; best transferability |
| H2. Type of absence data | Background; pseudo-absence; true absence; unspecified | |
| I. Thresholding | I1. Thresholding stage | Never; fitting/validation; categorical mapping |
| I2. Thresholding method | Default; statistical; expert-based; unspecified | |
| J. Classification accuracy assessment | J1. Consideration of spatial autocorrelation | No; yes |
| J2. Independent validation plots | No; yes | |
| J3. Accuracy assessment criterion | Best performance; spatial uncertainty; best transferability; expert-based | |
| K. One-class classifiers combination | K1. Classification combination method | Independent; higher probability; categorical map addition; unspecified |
| Area of Occupancy | Number of Studies |
|---|---|
| Potential area | 65 |
| Actual area | 66 |
| Potential and actual areas | 5 |
| Hierarchical Level | Number of Studies |
|---|---|
| Plant species | 102 |
| Plant community | 19 |
| Land cover | 15 |
| Acquisition Frequency | Number of Studies |
|---|---|
| Single | 67 |
| Multi-temporal | 34 |
| Annual time-series | 35 |
| Types of Points | ||||
|---|---|---|---|---|
| Type of Classifier | Background | Absence | Pseudo-Absence | Unspecified |
| Presence only | 44% | 15% | 28% | 13% |
| Presence/absence | 7% | 67% | 26% | 0% |
| Type of Classifier Fitting | Number of Studies |
|---|---|
| None | 89 |
| Best performance | 30 |
| Best transferability | 17 |
| Thresholding Stage | Number of Studies |
|---|---|
| None | 33 |
| Fitting/validation | 47 |
| Categorical mapping | 56 |
| Use of Spatially Uncorrelated Validation Plots | Use of Independent Validation Plots | |
|---|---|---|
| Yes | No | |
| Yes | 43 | 7 |
| No | 38 | 48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapinel, S.; Hubert-Moy, L. One-Class Classification of Natural Vegetation Using Remote Sensing: A Review. Remote Sens. 2021, 13, 1892. https://doi.org/10.3390/rs13101892
Rapinel S, Hubert-Moy L. One-Class Classification of Natural Vegetation Using Remote Sensing: A Review. Remote Sensing. 2021; 13(10):1892. https://doi.org/10.3390/rs13101892
Chicago/Turabian StyleRapinel, Sébastien, and Laurence Hubert-Moy. 2021. "One-Class Classification of Natural Vegetation Using Remote Sensing: A Review" Remote Sensing 13, no. 10: 1892. https://doi.org/10.3390/rs13101892
APA StyleRapinel, S., & Hubert-Moy, L. (2021). One-Class Classification of Natural Vegetation Using Remote Sensing: A Review. Remote Sensing, 13(10), 1892. https://doi.org/10.3390/rs13101892