
Linking Remotely Sensed Carbon and Water Use Efficiencies with In Situ Soil Properties

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Selection
2.2. Soil Physical and Chemical Analysis
2.3. Satellite Data
2.4. Statistical Analysis
3. Results
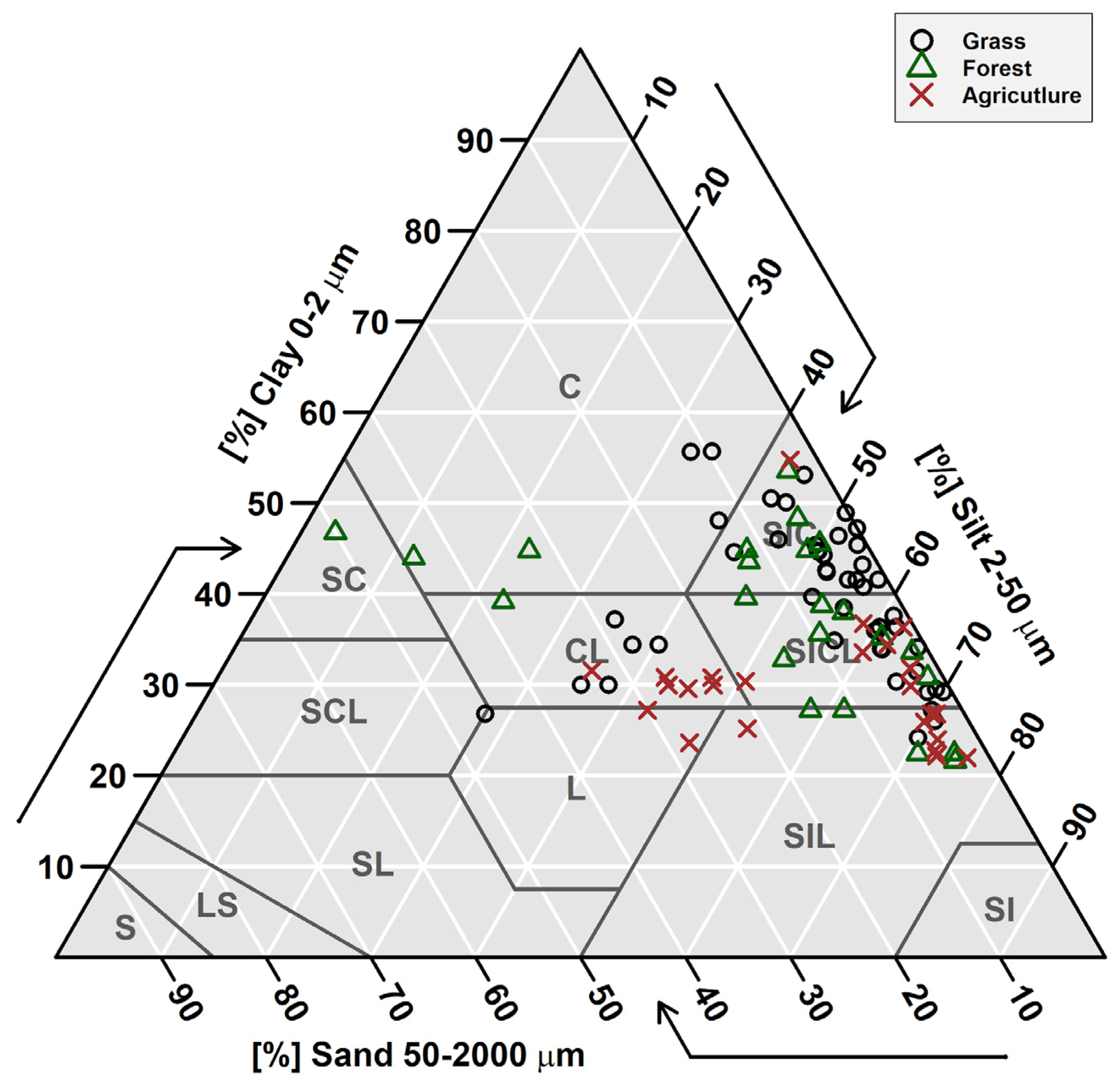
3.1. Soil Data
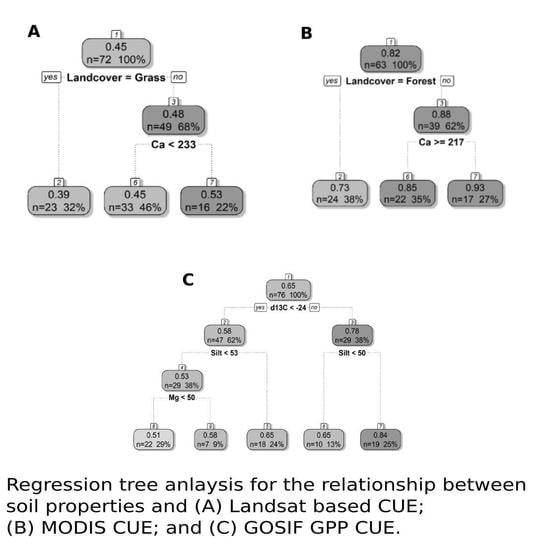
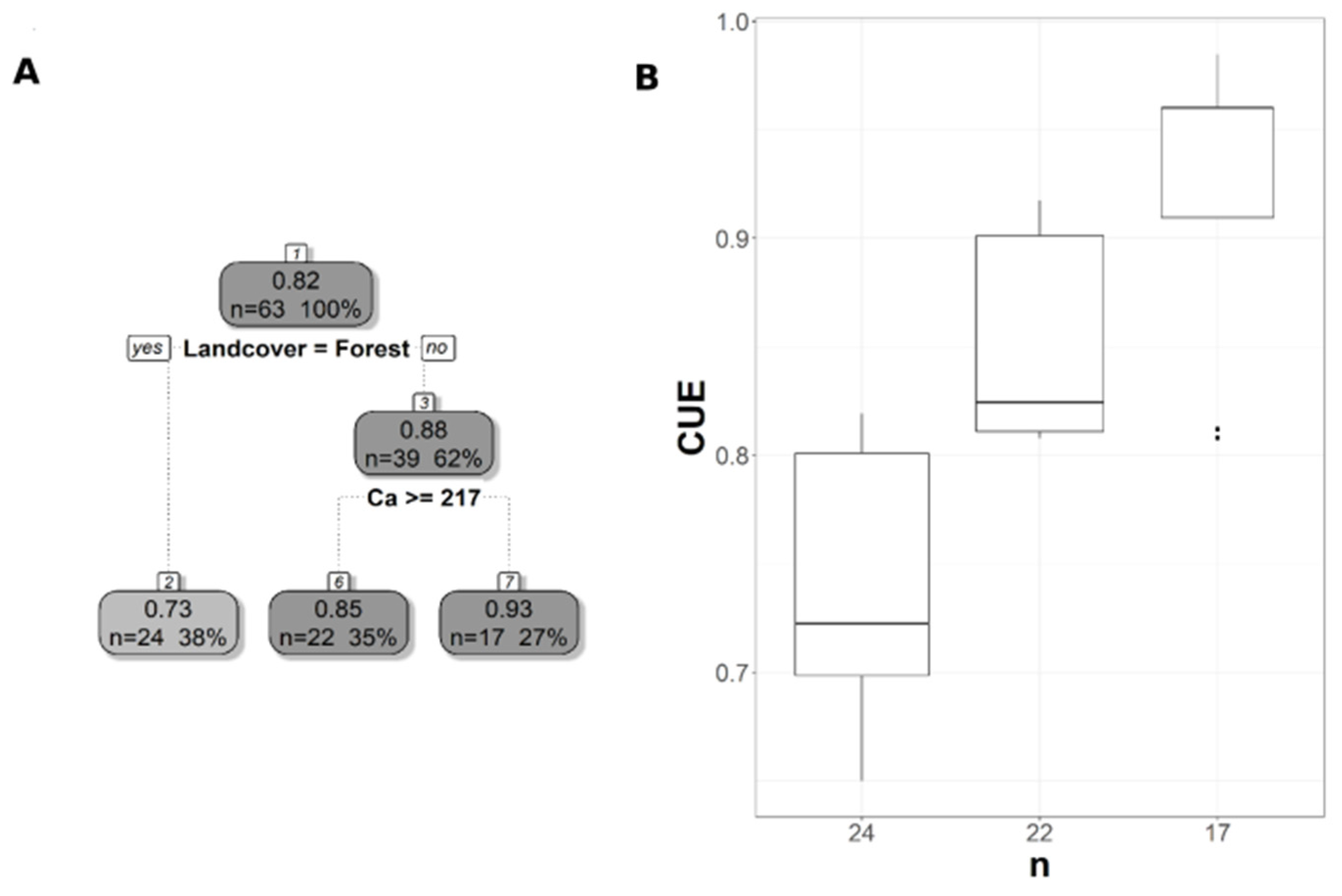
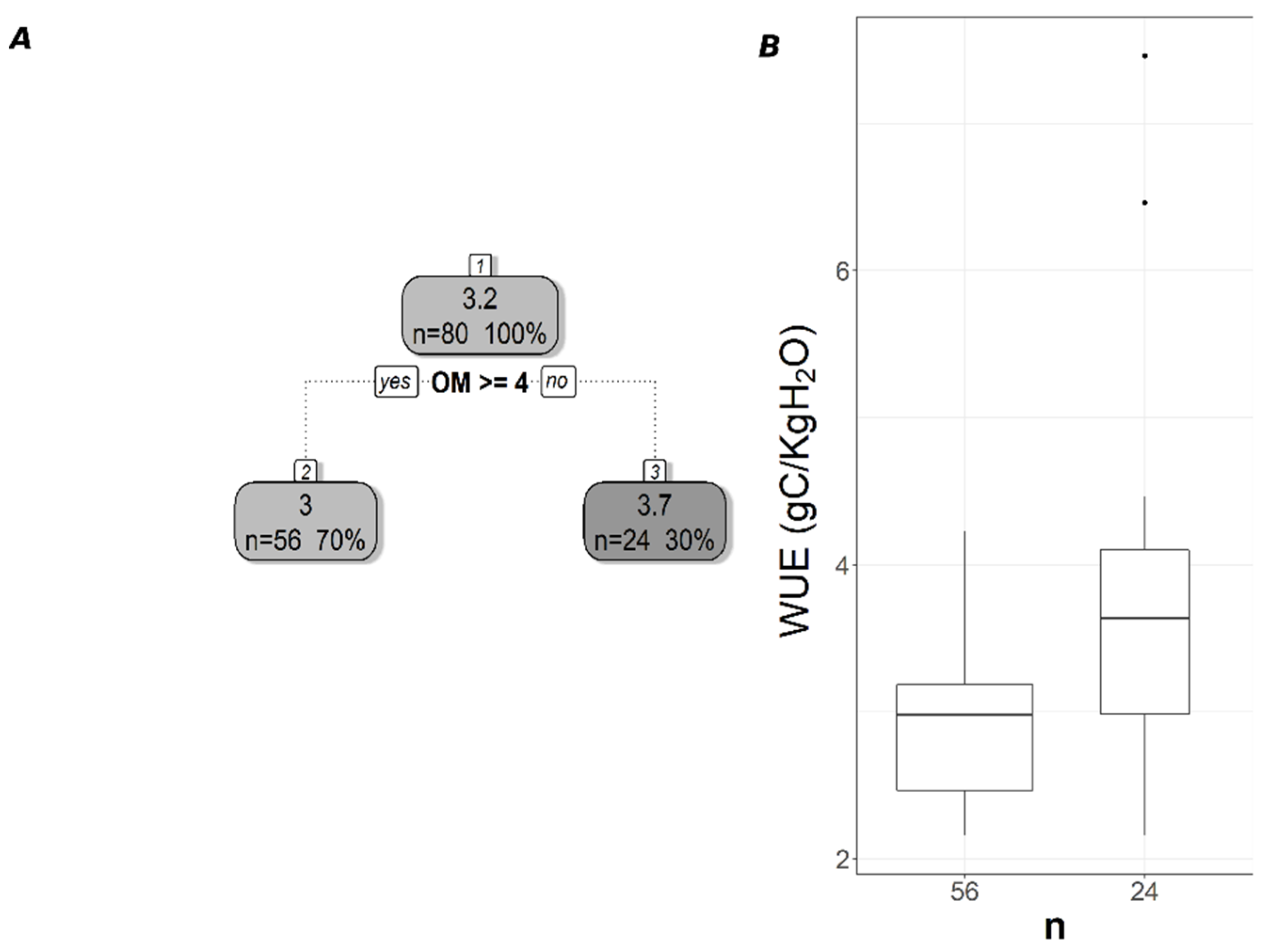
3.2. Soil Variables Versus CUE and WUE
3.3. Impacts of Different Productivity Measures and Spatial Resolution on the Relationship Satellite-Derived CUE and, WUE Withsoil Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapin, F.S.; Bloom, A.J.; Field, C.B.; Waring, R.H. Plant Responses to Multiple Environmental FactorsPhysiological Ecology Provides Tools for Studying How Interacting Environmental Resources Control Plant Growth. BioScience 1987, 37, 49–57. [Google Scholar] [CrossRef]
- Fisher, J.B.; Badgley, G.; Blyth, E. Global Nutrient Limitation in Terrestrial Vegetation. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, K. Co-Limitation of Plant Primary Productivity by Nitrogen and Phosphorus in a Species-Rich Wooded Meadow on Calcareous Soils. Acta Oecologica 2005, 28, 345–356. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Kelliher, F.M.; Korner, C.; Lloyd, J.; Leuning, R. Relationships Among Maximum Stomatal Conductance, Ecosystem Surface Conductance, Carbon Assimilation Rate, and Plant Nitrogen Nutrition: A Global Ecology Scaling Exercise. Annu. Rev. Ecol. Syst. 1994, 25, 629–660. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Sardans, J.; Luyssaert, S.; Campioli, M.; Iii, F.S.C.; Ciais, P.; Malhi, Y.; Obersteiner, M.; et al. Nutrient Availability as the Key Regulator of Global Forest Carbon Balance. Nat. Clim. Chang. 2014, 4, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Law, R.M.; Pak, B. A Global Model of Carbon, Nitrogen and Phosphorus Cycles for the Terrestrial Biosphere. Biogeosciences 2010, 7, 2261–2282. [Google Scholar] [CrossRef] [Green Version]
- Terrer, C.; Phillips, R.P.; Hungate, B.A.; Rosende, J.; Pett-Ridge, J.; Craig, M.E.; van Groenigen, K.J.; Keenan, T.F.; Sulman, B.N.; Stocker, B.D.; et al. A Trade-off between Plant and Soil Carbon Storage under Elevated CO2. Nature 2021, 591, 599–603. [Google Scholar] [CrossRef]
- De Vries, W.; Solberg, S.; Dobbertin, M.; Sterba, H.; Laubhann, D.; van Oijen, M.; Evans, C.; Gundersen, P.; Kros, J.; Wamelink, G.W.W.; et al. The Impact of Nitrogen Deposition on Carbon Sequestration by European Forests and Heathlands. For. Ecol. Manag. 2009, 258, 1814–1823. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Luyssaert, S.; Campioli, M.; Sardans, J.; Estiarte, M.; Peñuelas, J. Spatial Variability and Controls over Biomass Stocks, Carbon Fluxes, and Resource-Use Efficiencies across Forest Ecosystems. Trees 2014, 28, 597–611. [Google Scholar] [CrossRef]
- Hungate, B.A.; Dukes, J.S.; Shaw, M.R.; Luo, Y.; Field, C.B. Nitrogen and Climate Change. Science 2003, 302, 1512–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrer, C.; Jackson, R.B.; Prentice, I.C.; Keenan, T.F.; Kaiser, C.; Vicca, S.; Fisher, J.B.; Reich, P.B.; Stocker, B.D.; Hungate, B.A.; et al. Nitrogen and Phosphorus Constrain the CO2 Fertilization of Global Plant Biomass. Nat. Clim. Chang. 2019, 9, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial Phosphorus Limitation: Mechanisms, Implications, and Nitrogen–Phosphorus Interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Ågren, G.I.; Wetterstedt, J.Å.M.; Billberger, M.F.K. Nutrient Limitation on Terrestrial Plant Growth—Modeling the Interaction between Nitrogen and Phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef]
- Vicca, S.; Luyssaert, S.; Peñuelas, J.; Campioli, M.; Chapin, F.S.; Ciais, P.; Heinemeyer, A.; Högberg, P.; Kutsch, W.L.; Law, B.E.; et al. Fertile Forests Produce Biomass More Efficiently. Ecol. Lett. 2012, 15, 520–526. [Google Scholar] [CrossRef]
- Lanning, M.; Wang, L.; Scanlon, T.M.; Vadeboncoeur, M.A.; Adams, M.B.; Epstein, H.E.; Druckenbrod, D. Intensified Vegetation Water Use under Acid Deposition. Sci. Adv. 2019, 5, eaav5168. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Vitousek, P.M.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Zhou, G.; Zou, X.; Bai, E.; Scanlon, T.M.; Hou, E.; et al. Plant Acclimation to Long-Term High Nitrogen Deposition in an N-Rich Tropical Forest. Proc. Natl. Acad. Sci. USA 2018, 115, 5187–5192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripullone, F.; Lauteri, M.; Grassi, G.; Amato, M.; Borghetti, M. Variation in Nitrogen Supply Changes Water-Use Efficiency of Pseudotsuga Menziesii and Populus × Euroamericana; a Comparison of Three Approaches to Determine Water-Use Efficiency. Tree Physiol. 2004, 24, 671–679. [Google Scholar] [CrossRef]
- Fisher, J.B.; Sitch, S.; Malhi, Y.; Fisher, R.A.; Huntingford, C.; Tan, S.-Y. Carbon Cost of Plant Nitrogen Acquisition: A Mechanistic, Globally Applicable Model of Plant Nitrogen Uptake, Retranslocation, and Fixation. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.; Fisher, J.B.; Phillips, R.P.; Powers, J.S.; Brzostek, E.R. Modeling the Carbon Cost of Plant Nitrogen and Phosphorus Uptake Across Temperate and Tropical Forests. Front. For. Glob. Chang. 2020, 3, 43. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Fisher, J.B.; Phillips, R.P. Modeling the Carbon Cost of Plant Nitrogen Acquisition: Mycorrhizal Trade-Offs and Multipath Resistance Uptake Improve Predictions of Retranslocation. J. Geophys. Res. Biogeosciences 2014, 119, 1684–1697. [Google Scholar] [CrossRef]
- Burke, I.C.; Yonker, C.M.; Parton, W.J.; Cole, C.V.; Flach, K.; Schimel, D.S. Texture, Climate, and Cultivation Effects on Soil Organic Matter Content in U.S. Grassland Soils. Soil Sci. Soc. Am. J. 1989, 53, 800–805. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of Soil Organic Matter as an Ecosystem Property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Neina, D. The Role of Soil PH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, e5794869. [Google Scholar] [CrossRef]
- Aciego Pietri, J.C.; Brookes, P.C. Nitrogen Mineralisation along a PH Gradient of a Silty Loam UK Soil. Soil Biol. Biochem. 2008, 40, 797–802. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Farrington, H. Nutrient Limitation and Soil Development: Experimental Test of a Biogeochemical Theory. Biogeochemistry 1997, 37, 63–75. [Google Scholar] [CrossRef]
- Buendía, C.; Arens, S.; Hickler, T.; Higgins, S.I.; Porada, P.; Kleidon, A. On the Potential Vegetation Feedbacks That Enhance Phosphorus Availability—Insights from a Process-Based Model Linking Geological and Ecological Timescales. Biogeosciences 2014, 11, 3661–3683. [Google Scholar] [CrossRef] [Green Version]
- Vadeboncoeur, M.A. Meta-Analysis of Fertilization Experiments Indicates Multiple Limiting Nutrients in Northeastern Deciduous Forests. Can. J. For. Res. 2010, 40, 1766–1780. [Google Scholar] [CrossRef] [Green Version]
- Jobbágy, E.G.; Jackson, R.B. The Distribution of Soil Nutrients with Depth: Global Patterns and the Imprint of Plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Stark, J.M. Causes of Soil Nutrient Heterogeneity at Different Scales. In Exploitation of Environmental Heterogeneity by Plants; Caldwell, M.M., Pearcy, R.W., Eds.; Physiological Ecology; Academic Press: Boston, MA, USA, 1994; pp. 255–284. ISBN 978-0-12-155070-7. [Google Scholar]
- Keller, A.B.; Brzostek, E.R.; Craig, M.E.; Fisher, J.B.; Phillips, R.P. Root-Derived Inputs Are Major Contributors to Soil Carbon in Temperate Forests, but Vary by Mycorrhizal Type. Ecol. Lett. 2021, 24, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Sousa, D.; Fisher, J.B.; Galvan, F.R.; Pavlick, R.P.; Cordell, S.; Giambelluca, T.W.; Giardina, C.P.; Gilbert, G.S.; Imran-Narahari, F.; Litton, C.M.; et al. Tree Canopies Reflect Mycorrhizal Composition. Geophys. Res. Lett. 2021, 48, e2021GL092764. [Google Scholar] [CrossRef]
- Köhler, I.H.; Macdonald, A.J.; Schnyder, H. Last-Century Increases in Intrinsic Water-Use Efficiency of Grassland Communities Have Occurred over a Wide Range of Vegetation Composition, Nutrient Inputs, and Soil PH1. Plant Physiol. 2016, 170, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Yu, G. Spatial Variations and Controls of Carbon Use Efficiency in China’s Terrestrial Ecosystems. Sci. Rep. 2019, 9, 19516. [Google Scholar] [CrossRef]
- Maxwell, T.M.; Silva, L.C.R.; Horwath, W.R. Integrating Effects of Species Composition and Soil Properties to Predict Shifts in Montane Forest Carbon–Water Relations. Proc. Natl. Acad. Sci. USA 2018, 115, E4219–E4226. [Google Scholar] [CrossRef] [Green Version]
- DeLUCIA, E.H.; Drake, J.E.; Thomas, R.B.; Gonzalez-Meler, M. Forest Carbon Use Efficiency: Is Respiration a Constant Fraction of Gross Primary Production? Glob. Chang. Biol. 2007, 13, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Jassal, R.S.; Black, T.A.; Spittlehouse, D.L.; Brümmer, C.; Nesic, Z. Evapotranspiration and Water Use Efficiency in Different-Aged Pacific Northwest Douglas-Fir Stands. Agric. For. Meteorol. 2009, 149, 1168–1178. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Saifullah; Ahmad, M. Improving Agricultural Water Use Efficiency by Nutrient Management in Crop Plants. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in Forest Water-Use Efficiency as Atmospheric Carbon Dioxide Concentrations Rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Tang, X.; Li, H.; Desai, A.R.; Nagy, Z.; Luo, J.; Kolb, T.E.; Olioso, A.; Xu, X.; Yao, L.; Kutsch, W.; et al. How Is Water-Use Efficiency of Terrestrial Ecosystems Distributed and Changing on Earth? Sci. Rep. 2014, 4, 7483. [Google Scholar] [CrossRef]
- Robinson, N.P.; Allred, B.W.; Smith, W.K.; Jones, M.O.; Moreno, A.; Erickson, T.A.; Naugle, D.E.; Running, S.W. Terrestrial Primary Production for the Conterminous United States Derived from Landsat 30 m and MODIS 250 m. Remote Sens. Ecol. Conserv. 2018, 4, 264–280. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Margolis, H.A.; Cescatti, A.; Richardson, A.D.; Arain, M.A.; Arneth, A.; Bernhofer, C.; Bonal, D.; Chen, J.; et al. Global Patterns of Land-Atmosphere Fluxes of Carbon Dioxide, Latent Heat, and Sensible Heat Derived from Eddy Covariance, Satellite, and Meteorological Observations. J. Geophys. Res. Biogeosciences 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.B.; Lee, B.; Purdy, A.J.; Halverson, G.H.; Dohlen, M.B.; Cawse-Nicholson, K.; Wang, A.; Anderson, R.G.; Aragon, B.; Arain, M.A.; et al. ECOSTRESS: NASA’s Next Generation Mission to Measure Evapotranspiration From the International Space Station. Water Resour. Res. 2020, 56, 26058. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J.; Fisher, J.B.; Baldocchi, D.D. ECOSTRESS Estimates Gross Primary Production with Fine Spatial Resolution for Different Times of Day from the International Space Station. Remote Sens. Environ. 2021, 258, 112360. [Google Scholar] [CrossRef]
- Zhang, Y.; Guanter, L.; Berry, J.A.; van der Tol, C.; Yang, X.; Tang, J.; Zhang, F. Model-Based Analysis of the Relationship between Sun-Induced Chlorophyll Fluorescence and Gross Primary Production for Remote Sensing Applications. Remote Sens. Environ. 2016, 187, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Magney, T.S.; Bowling, D.R.; Logan, B.A.; Grossmann, K.; Stutz, J.; Blanken, P.D.; Burns, S.P.; Cheng, R.; Garcia, M.A.; Kӧhler, P.; et al. Mechanistic Evidence for Tracking the Seasonality of Photosynthesis with Solar-Induced Fluorescence. Proc. Natl. Acad. Sci. USA 2019, 116, 11640–11645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xiao, J.; He, B.; Arain, M.A.; Beringer, J.; Desai, A.R.; Emmel, C.; Hollinger, D.Y.; Krasnova, A.; Mammarella, I.; et al. Solar-Induced Chlorophyll Fluorescence Is Strongly Correlated with Terrestrial Photosynthesis for a Wide Variety of Biomes: First Global Analysis Based on OCO-2 and Flux Tower Observations. Glob. Chang. Biol. 2018, 24, 3990–4008. [Google Scholar] [CrossRef]
- He, Y.; Piao, S.; Li, X.; Chen, A.; Qin, D. Global Patterns of Vegetation Carbon Use Efficiency and Their Climate Drivers Deduced from MODIS Satellite Data and Process-Based Models. Agric. For. Meteorol. 2018, 256–257, 150–158. [Google Scholar] [CrossRef]
- Kwon, Y.; Larsen, C.P.S. Effects of Forest Type and Environmental Factors on Forest Carbon Use Efficiency Assessed Using MODIS and FIA Data across the Eastern USA. Int. J. Remote Sens. 2013, 34, 8425–8448. [Google Scholar] [CrossRef]
- Xue, B.-L.; Guo, Q.; Otto, A.; Xiao, J.; Tao, S.; Li, L. Global Patterns, Trends, and Drivers of Water Use Efficiency from 2000 to 2013. Ecosphere 2015, 6, art174. [Google Scholar] [CrossRef]
- Huang, L.; He, B.; Han, L.; Liu, J.; Wang, H.; Chen, Z. A Global Examination of the Response of Ecosystem Water-Use Efficiency to Drought Based on MODIS Data. Sci. Total Environ. 2017, 601–602, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, J.; Liu, S.; Rentch, J.S.; Sun, P.; Lu, C. Global Gross Primary Productivity and Water Use Efficiency Changes under Drought Stress. Environ. Res. Lett. 2017, 12, 014016. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.B.; Perakalapudi, N.V.; Turner, B.L.; Schimel, D.S.; Cusack, D.F. Competing Effects of Soil Fertility and Toxicity on Tropical Greening. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Inatomi, M. Water-Use Efficiency of the Terrestrial Biosphere: A Model Analysis Focusing on Interactions between the Global Carbon and Water Cycles. J. Hydrometeorol. 2011, 13, 681–694. [Google Scholar] [CrossRef]
- Xiao, J.; Sun, G.; Chen, J.; Chen, H.; Chen, S.; Dong, G.; Gao, S.; Guo, H.; Guo, J.; Han, S.; et al. Carbon Fluxes, Evapotranspiration, and Water Use Efficiency of Terrestrial Ecosystems in China. Agric. For. Meteorol. 2013, 182–183, 76–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, M.; Chen, H.; Adams, J. Global Pattern of NPP to GPP Ratio Derived from MODIS Data: Effects of Ecosystem Type, Geographical Location and Climate. Glob. Ecol. Biogeogr. 2009, 18, 280–290. [Google Scholar] [CrossRef]
- Kim, D.; Lee, M.-I.; Jeong, S.-J.; Im, J.; Cha, D.H.; Lee, S. Intercomparison of Terrestrial Carbon Fluxes and Carbon Use Efficiency Simulated by CMIP5 Earth System Models. Biogeosciences Discuss. 2016, 2016, 1–50. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated World Map of the Köppen-Geiger Climate Classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Schoeneberger, P.J.; Wysocki, D.A.; Benham, E.C. Field Book for Describing and Sampling Soils, Version 3.0 NRCS Soils. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054184 (accessed on 20 May 2021).
- Soil Survey Laboratory Methods Manual. NRCS Soils. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054247 (accessed on 18 February 2020).
- Hoogsteen, M.J.J.; Lantinga, E.A.; Bakker, E.J.; Groot, J.C.J.; Tittonell, P.A. Estimating Soil Organic Carbon through Loss on Ignition: Effects of Ignition Conditions and Structural Water Loss. Eur. J. Soil Sci. 2015, 66, 320–328. [Google Scholar] [CrossRef]
- Sikora, F.J. A Buffer That Mimics the SMP Buffer for Determining Lime Requirement of Soil. Soil Sci. Soc. Am. J. 2006, 70, 474–486. [Google Scholar] [CrossRef]
- Soil Analysis Handbook of Reference Methods. Available online: https://www.routledge.com/Soil-Analysis-Handbook-of-Reference-Methods/Jones-Jr/p/book/9780849302053 (accessed on 24 April 2021).
- Hosseini Bai, S.; Xu, C.-Y.; Xu, Z.; Blumfield, T.J.; Zhao, H.; Wallace, H.; Reverchon, F.; Van Zwieten, L. Soil and Foliar Nutrient and Nitrogen Isotope Composition (Δ15N) at 5 Years after Poultry Litter and Green Waste Biochar Amendment in a Macadamia Orchard. Environ. Sci. Pollut. Res. 2015, 22, 3803–3809. [Google Scholar] [CrossRef]
- Malone, E.T.; Abbott, B.W.; Klaar, M.J.; Kidd, C.; Sebilo, M.; Milner, A.M.; Pinay, G. Decline in Ecosystem Δ13C and Mid-Successional Nitrogen Loss in a Two-Century Postglacial Chronosequence. Ecosystems 2018, 21, 1659–1675. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, J. Mapping Photosynthesis Solely from Solar-Induced Chlorophyll Fluorescence: A Global, Fine-Resolution Dataset of Gross Primary Production Derived from OCO-2. Remote Sens. 2019, 11, 2563. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, J. A Global, 0.05-Degree Product of Solar-Induced Chlorophyll Fluorescence Derived from OCO-2, MODIS, and Reanalysis Data. Remote Sens. 2019, 11, 517. [Google Scholar] [CrossRef] [Green Version]
- Lukens, W.E.; Stinchcomb, G.E.; Nordt, L.C.; Kahle, D.J.; Driese, S.G.; Tubbs, J.D. Recursive Partitioning Improves Paleosol Proxies for Rainfall. Am. J. Sci. 2019, 319, 819–845. [Google Scholar] [CrossRef]
- Tittonell, P.; Shepherd, K.; Vanlauwe, B.; Giller, K. Unravelling the Effects of Soil and Crop Management on Maize Productivity in Smallholder Agricultural Systems of Western Kenya—An Application of Classification and Regression Tree Analysis. Agric. Ecosyst. Environ. 2008, 123, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological Interpretations of Nitrogen Isotope Ratios of Terrestrial Plants and Soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Lin, A.; Wang, L.; Xia, Y.; Zou, L. Evaluation of MODIS Gross Primary Production across Multiple Biomes in China Using Eddy Covariance Flux Data. Remote Sens. 2016, 8, 395. [Google Scholar] [CrossRef] [Green Version]
- Acton, P.; Fox, J.; Campbell, E.; Rowe, H.; Wilkinson, M. Carbon Isotopes for Estimating Soil Decomposition and Physical Mixing in Well-Drained Forest Soils: CARBON ISOTOPES FOR FOREST SOILS. J. Geophys. Res. Biogeosci. 2013, 118, 1532–1545. [Google Scholar] [CrossRef]
- Moreno-Gutiérrez, C.; Dawson, T.E.; Nicolás, E.; Querejeta, J.I. Isotopes Reveal Contrasting Water Use Strategies among Coexisting Plant Species in a Mediterranean Ecosystem. New Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.E.; Kauwe, M.G.D.; Lin, Y.-S.; Knauer, J.; Duursma, R.A.; Williams, C.A.; Arneth, A.; Clement, R.; Isaac, P.; Limousin, J.-M.; et al. How Do Leaf and Ecosystem Measures of Water-Use Efficiency Compare? New Phytol. 2017, 216, 758–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Wang, W.; Zeng, W.; Zeng, H. Nitrogen Deposition Enhances Carbon Sequestration by Plantations in Northern China. PLoS ONE 2014, 9, e87975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, N.; Chen, S.; Wilske, B.; Sun, G.; Chen, J. Evapotranspiration and Soil Water Relationships in a Range of Disturbed and Undisturbed Ecosystems in the Semi-Arid Inner Mongolia, China. J. Plant Ecol. 2011, 4, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Groffman, P.M.; Fisk, M.C.; Driscoll, C.T.; Likens, G.E.; Fahey, T.J.; Eagar, C.; Pardo, L.H. Calcium Additions and Microbial Nitrogen Cycle Processes in a Northern Hardwood Forest. Ecosystems 2006, 9, 1289–1305. [Google Scholar] [CrossRef]
- Minick, K.J.; Fisk, M.C.; Groffman, P.M. Soil Ca Alters Processes Contributing to C and N Retention in the Oa/A Horizon of a Northern Hardwood Forest. Biogeochemistry 2017, 132, 343–357. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wimmer, R. Tansley Review No. 104, Calcium Physiology and Terrestrial Ecosystem Processes. New Phytol. 1999, 142, 373–417. [Google Scholar] [CrossRef]
- Gransee, A.; Führs, H. Magnesium Mobility in Soils as a Challenge for Soil and Plant Analysis, Magnesium Fertilization and Root Uptake under Adverse Growth Conditions. Plant Soil 2013, 368, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J. Potassium: A Neglected Nutrient in Global Change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Hauer-Jákli, M.; Tränkner, M. Critical Leaf Magnesium Thresholds and the Impact of Magnesium on Plant Growth and Photo-Oxidative Defense: A Systematic Review and Meta-Analysis From 70 Years of Research. Front. Plant Sci. 2019, 10, 766. [Google Scholar] [CrossRef] [PubMed]
- Hedwall, P.-O.; Bergh, J.; Brunet, J. Phosphorus and Nitrogen Co-Limitation of Forest Ground Vegetation under Elevated Anthropogenic Nitrogen Deposition. Oecologia 2017, 185, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooley, S.S.; Fisher, J.B.; Halverson, G.H.; Goldmisth, G.R. Convergence in Water Use Efficiency within Plant Functional Types across Contrasting Climates. Nat. Plants. in press.
- Fisher, J.B.; Tu, K.P.; Baldocchi, D.D. Global Estimates of the Land–Atmosphere Water Flux Based on Monthly AVHRR and ISLSCP-II Data, Validated at 16 FLUXNET Sites. Remote Sens. Environ. 2008, 112, 901–919. [Google Scholar] [CrossRef]
- Ryu, Y.; Baldocchi, D.D.; Kobayashi, H.; van Ingen, C.; Li, J.; Black, T.A.; Beringer, J.; van Gorsel, E.; Knohl, A.; Law, B.E.; et al. Integration of MODIS Land and Atmosphere Products with a Coupled-Process Model to Estimate Gross Primary Productivity and Evapotranspiration from 1 Km to Global Scales. Glob. Biogeochem. Cycles 2011, 25. [Google Scholar] [CrossRef] [Green Version]
- El Masri, B.; Schwalm, C.; Huntzinger, D.N.; Mao, J.; Shi, X.; Peng, C.; Fisher, J.B.; Jain, A.K.; Tian, H.; Poulter, B.; et al. Carbon and Water Use Efficiencies: A Comparative Analysis of Ten Terrestrial Ecosystem Models under Changing Climate. Sci. Rep. 2019, 9, 14680. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
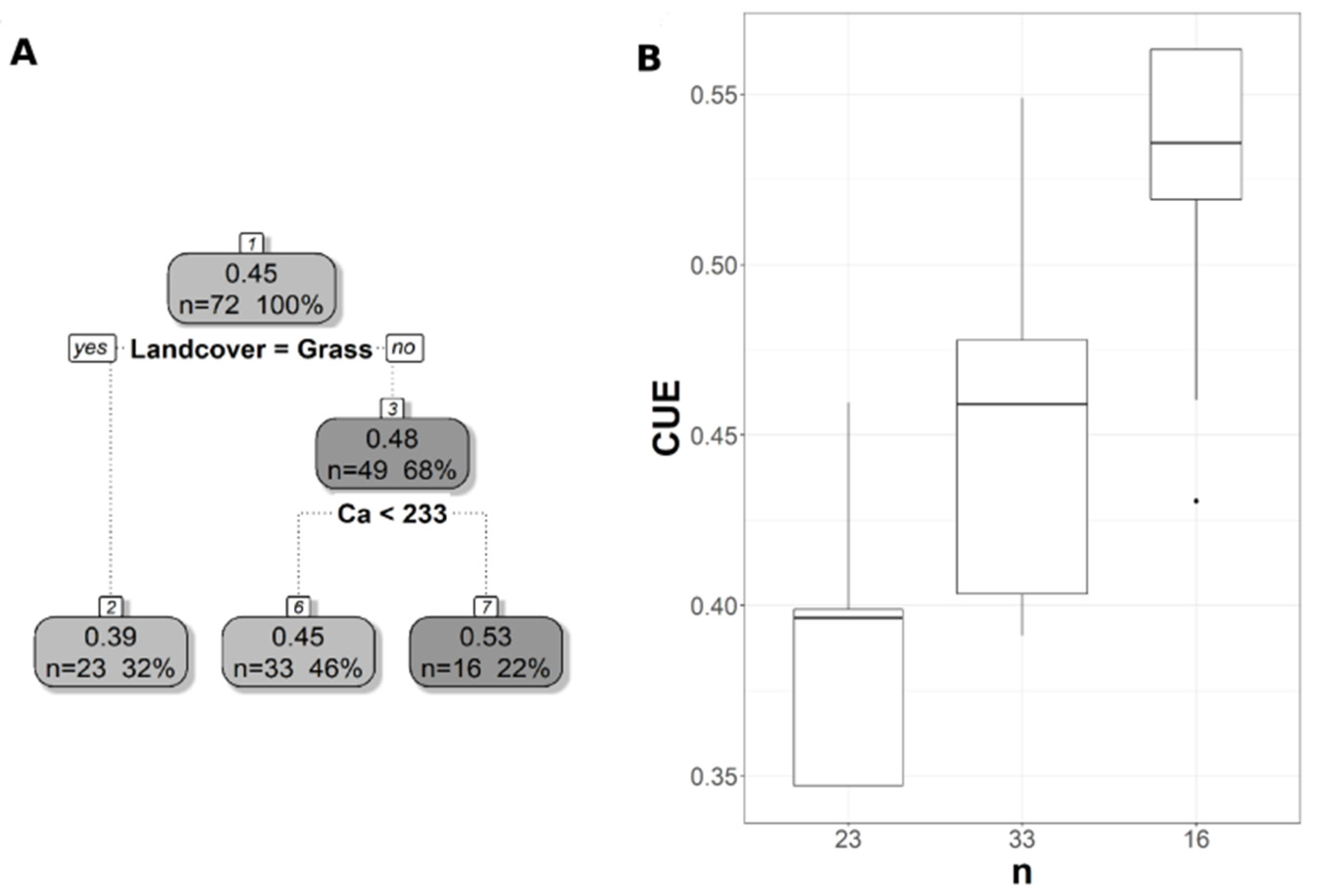{kind=link}
{kind=link}
{kind=link}
| Site ID | Latitude | Longitude | Land Cover |
|---|---|---|---|
| DRJ | 37.40N | 88.34W | Agriculture |
| PTN | 37.40N | 87.70W | Agriculture |
| HWY | 37.68N | 86.98W | Agriculture |
| COR | 37.22N | 83.89W | Agriculture |
| LWF | 37.20N | 83.92W | Agriculture |
| UKS | 37.97N | 84.53W | Agriculture |
| ARB | 36.60N | 88.33W | Grassland |
| NFM | 36.63N | 88.33W | Grassland |
| FFG | 37.75N | 86.62W | Grassland |
| HLF | 37.88N | 84.46W | Grassland |
| HLP | 38.13N | 86.71W | Grassland |
| SPF | 38.13N | 84.50W | Grassland |
| BCF | 36.60N | 88.23W | Grassland |
| HBS | 36.76N | 88.12W | Deciduous Forest |
| CR | 36.93N | 88.46W | Deciduous Forest |
| HEA | 37.41N | 88.19W | Deciduous Forest |
| FFF | 37.75N | 86.63W | Deciduous Forest |
| RFS | 37.47N | 83.17W | Deciduous Forest |
| MMF | 36.69N | 88.15W | Deciduous Forest |
| Site ID | Horizon | Top of Subsoil (cm) | Soil Order | BD | LOI | pH | N (%) | δ13C (‰) | δ15N (‰) | C(%) | C:N | P (g/m2) | K (g/m2) | Ca (g/m2) | Mg (g/m2) | Zn (g/m2) | Fe (mg/kg) | Land Cover |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DRJ | A | 25 | Alfisol | 1.26 | 5.78 | 6.00 | 0.12 | −21.8 | 7.5 | 1.46 | 11.86 | 31.22 | 36.04 | 474.62 | 16.03 | 2.20 | 196.50 | AG |
| B | 1.34 | 4.75 | 5.75 | 0.05 | −24.1 | 7.5 | 0.33 | 6.68 | 5.21 | 21.88 | 437.97 | 32.62 | 0.68 | 135.25 | ||||
| PTN | A | 20 | Alfisol | 1.24 | 5.16 | 6.70 | 0.10 | −27.1 | 4.7 | 1.21 | 12.34 | 3.36 | 11.32 | 389.78 | 17.88 | 0.47 | 167.50 | AG |
| B | 1.40 | 3.24 | 5.27 | 0.04 | −22.9 | 8.1 | 0.33 | 8.29 | 1.20 | 17.90 | 197.72 | 28.13 | 0.03 | 110.33 | ||||
| HWY | A | 12 | Alfisol | 1.06 | 5.54 | - | 0.15 | −22.6 | 6.0 | 1.53 | 10.00 | 1.79 | 8.74 | 284.81 | 31.94 | 0.89 | 120.00 | AG |
| B | 1.26 | 3.53 | 5.30 | 0.04 | −23.0 | 7.5 | 0.34 | 7.85 | 0.31 | 7.96 | 171.36 | 21.88 | 0.10 | 58.00 | ||||
| COR | A | 30 | Alfisol | 1.15 | 3.75 | 5.77 | 0.07 | −24.8 | 7.7 | 0.67 | 9.84 | 93.70 | 26.60 | 180.08 | 17.86 | 0.35 | 275.33 | AG |
| B | 1.00 | 8.61 | 6.30 | 0.24 | −27.3 | 3.3 | 2.66 | 11.03 | 2.80 | 48.98 | 278.19 | 39.68 | 0.45 | 104.00 | ||||
| LWF | A | 26 | Alfisol | 1.43 | 5.17 | 5.13 | 0.09 | −25.8 | 7.1 | 0.84 | 9.86 | 128.37 | 18.49 | 161.81 | 14.05 | 0.32 | 317.00 | AG |
| B | 1.57 | 4.81 | 5.00 | 0.03 | −23.9 | 7.4 | 0.18 | 6.35 | 2.73 | 12.96 | 138.31 | 7.62 | 0.09 | 50.67 | ||||
| UKS | A | 22 | Alfisol | 1.16 | 6.92 | 5.55 | 0.11 | −23.7 | 7.7 | 1.03 | 8.02 | 21.13 | 79.52 | 311.54 | 32.11 | 0.65 | 151.50 | AG |
| B | 1.22 | 6.18 | 5.63 | 0.08 | −23.5 | 7.8 | 0.56 | 5.83 | 11.46 | 48.95 | 315.21 | 32.36 | 0.33 | 133.00 | ||||
| ARB | A | 24 | Alfisol | 1.23 | 4.97 | 5.70 | 0.13 | −21.3 | 7.2 | 1.19 | 9.05 | 2.35 | 10.54 | 194.84 | 32.09 | 0.70 | 149.33 | GR |
| B | 1.27 | 3.73 | 4.77 | 0.04 | −22.5 | 7.2 | 0.30 | 7.40 | 0.56 | 13.90 | 184.42 | 67.96 | 0.21 | 74.67 | ||||
| NFM | A | 4 | Alfisol | 0.83 | 7.74 | 5.30 | 0.29 | −20.7 | 6.3 | 2.57 | 8.86 | 10.54 | 70.50 | 310.14 | 45.62 | 1.97 | 203.00 | GR |
| B | 1.32 | 3.45 | 5.40 | 0.06 | −21.6 | 8.1 | 0.54 | 8.61 | 1.77 | 34.91 | 204.36 | 40.85 | 0.68 | 70.25 | ||||
| FFG | A | 23 | Alfisol | 1.21 | 4.63 | 5.75 | 0.08 | −25.0 | 5.6 | 0.76 | 9.88 | 2.19 | 9.30 | 233.70 | 12.75 | 0.11 | 107.75 | GR |
| B | 1.30 | 4.42 | 5.60 | 0.06 | −24.5 | 8.8 | 0.64 | 9.43 | 0.84 | 9.75 | 263.29 | 12.55 | 0.08 | 101.00 | ||||
| HLF | A | 23 | Alfisol | 1.09 | 9.42 | 5.85 | 0.26 | −25.1 | 6.3 | 2.48 | 9.58 | 48.53 | 75.21 | 357.72 | 41.92 | 0.85 | 203.00 | GR |
| B | 1.33 | 6.68 | 5.28 | 0.06 | −22.8 | 7.9 | 0.41 | 6.52 | 19.17 | 44.27 | 289.10 | 23.73 | 0.10 | 168.25 | ||||
| HLP | A | 22 | Alfisol | 1.24 | 5.93 | 5.55 | 0.13 | −23.2 | 7.2 | 1.03 | 6.79 | 7.96 | 19.67 | 301.06 | 18.05 | 0.47 | 135.00 | GR |
| B | 1.41 | 4.34 | 5.95 | 0.08 | −23 | 8.0 | 0.56 | 6.13 | 1.20 | 17.43 | 382.13 | 9.89 | 0.10 | 94.75 | ||||
| SPF | A | 29 | Alfisol | 1.41 | 8.73 | 6.90 | 0.20 | −24.1 | 6.5 | 1.76 | 8.93 | 65.27 | 18.01 | 726.50 | 33.66 | 19.00 | 221.33 | GR |
| B | 1.42 | 7.18 | 6.33 | 0.10 | −22.9 | 7.9 | 0.76 | 7.27 | 74.09 | 30.97 | 596.44 | 33.59 | 1.55 | 284.00 | ||||
| BCF | A | 14 | Alfisol | 1.43 | 4.97 | 5.90 | 0.11 | −25.8 | 6.8 | 1.20 | 11.15 | 30.66 | 64.56 | 268.39 | 36.09 | 1.00 | 270.50 | GR |
| B | 1.48 | 4.56 | 5.08 | 0.04 | −24.9 | 7.6 | 0.39 | 8.54 | 2.13 | 43.71 | 169.00 | 80.84 | 0.08 | 113.75 | ||||
| HBS | A | 7 | fUltisol | 0.84 | 25.56 | - | 0.35 | −27.9 | 0.1 | 5.90 | 16.57 | 8.46 | 16.36 | 124.97 | 22.30 | 0.73 | 173.00 | FR |
| B | 1.31 | 3.33 | 3.93 | 0.04 | −25.9 | 7.6 | 0.50 | 12.24 | 1.65 | 16.95 | 42.90 | 42.87 | 0.13 | 94.75 | ||||
| CR | A | 21 | Ultisol | 0.86 | 7.65 | 4.30 | 0.09 | −26.7 | 8.0 | 0.94 | 10.82 | - | - | - | - | - | - | FR |
| B | 1.19 | 4.10 | 4.20 | 0.03 | −25.5 | 6.4 | 0.26 | 9.03 | 3.11 | 14.74 | 22.22 | 63.69 | 0.69 | 214.00 | ||||
| HEA | A | 16 | Alfisol | 1.35 | 2.52 | 4.40 | 0.04 | −26.5 | 5.9 | 0.62 | 13.95 | 3.47 | 13.90 | 37.32 | 17.49 | 0.16 | 91.00 | FR |
| B | 1.24 | 3.47 | 3.98 | 0.02 | −25.4 | 8.3 | 0.27 | 10.57 | 0.78 | 12.69 | 42.56 | 37.07 | 0.15 | 67.75 | ||||
| FFF | A | 7 | Alfisol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | FR |
| B | 1.30 | 4.64 | 7.13 | 0.08 | −25.5 | 3.4 | 0.80 | 9.71 | 0.95 | 41.58 | 805.61 | 33.82 | 0.21 | 73.25 | ||||
| RFS | A | 17 | Ultisol | 1.33 | 6.45 | 5.73 | 0.14 | −27.0 | 6.2 | 1.65 | 12.52 | 1.53 | 23.13 | 231.98 | 26.83 | 0.29 | 52.33 | FR |
| B | 1.24 | 5.09 | 5.68 | 0.08 | −26.4 | 6.3 | 0.84 | 10.53 | 0.92 | 23.09 | 107.91 | 24.71 | 0.09 | 104.00 | ||||
| MMF | A | 11 | Alfisol | 1.20 | 3.31 | 4.60 | 0.06 | −26.1 | - | 0.71 | 12.30 | 2.58 | 18.38 | 27.85 | 14.18 | 0.22 | 116.50 | FR |
| B | 1.40 | 4.89 | 4.00 | 0.04 | −24.3 | - | 0.34 | 8.41 | 0.50 | 35.47 | 89.42 | 108.67 | 0.11 | 102.25 |
| CART | CARTnLC | CARTnSO | CARTnSH | CARTnSD | ||
|---|---|---|---|---|---|---|
| WUE | r2 | 0.31 | 0.39 | 0.15 | 0.31 | 0.31 |
| MAE | 0.17 | 0.17 | 0.17 | 0.17 | 0.17 | |
| WUESIF | r2 | 0.19 | 0.19 | 0.19 | 0.19 | 0.19 |
| MAE | 1.4 | 0.64 | 0.64 | 0.64 | 0.64 | |
| CUE30 | r2 | 0.62 | 0.52 | 0.62 | 0.64 | 0.59 |
| MAE | 0.04 | 0.03 | 0.04 | 0.04 | 0.04 | |
| CUE250 | r2 | 0.78 | 0.61 | 0.78 | 0.79 | 0.78 |
| MAE | 0.06 | 0.07 | 0.06 | 0.06 | 0.06 | |
| CUESIF | r2 | 0.45 | 0.39 | 0.4 | 0.4 | 0.4 |
| MAE | 0.1 | 0.11 | 0.11 | 0.1 | 0.1 |
| Data | GPP Algorithm | References |
|---|---|---|
| MODIS 250 m | Light use efficiency model using MODIS 250 m derived leaf area index and the fraction of photosynthetically active radiation | [40] |
| Landsat 30 m | Light use efficiency model using Landsat 30 m derived leaf area index and the fraction of photosynthetically active radiation | [40] |
| GOSIF GPP | OCO-2 SIF vs. GPP universal relationships | [66,67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Masri, B.; Stinchcomb, G.E.; Cetin, H.; Ferguson, B.; Kim, S.L.; Xiao, J.; Fisher, J.B. Linking Remotely Sensed Carbon and Water Use Efficiencies with In Situ Soil Properties. Remote Sens. 2021, 13, 2593. https://doi.org/10.3390/rs13132593
El Masri B, Stinchcomb GE, Cetin H, Ferguson B, Kim SL, Xiao J, Fisher JB. Linking Remotely Sensed Carbon and Water Use Efficiencies with In Situ Soil Properties. Remote Sensing. 2021; 13(13):2593. https://doi.org/10.3390/rs13132593
Chicago/Turabian StyleEl Masri, Bassil, Gary E. Stinchcomb, Haluk Cetin, Benedict Ferguson, Sora L. Kim, Jingfeng Xiao, and Joshua B. Fisher. 2021. "Linking Remotely Sensed Carbon and Water Use Efficiencies with In Situ Soil Properties" Remote Sensing 13, no. 13: 2593. https://doi.org/10.3390/rs13132593
APA StyleEl Masri, B., Stinchcomb, G. E., Cetin, H., Ferguson, B., Kim, S. L., Xiao, J., & Fisher, J. B. (2021). Linking Remotely Sensed Carbon and Water Use Efficiencies with In Situ Soil Properties. Remote Sensing, 13(13), 2593. https://doi.org/10.3390/rs13132593