
Disentangling LiDAR Contribution in Modelling Species–Habitat Structure Relationships in Terrestrial Ecosystems Worldwide. A Systematic Review and Future Directions

Abstract
:1. Introduction
2. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LiDAR Traits | Description | Examples of LiDAR Explanatory Variables |
|---|---|---|
| CS: canopy height | Includes metrics used to obtain canopy height measures (e.g., maximum, mode, 95th percentile, etc.). | Mean height [36,37]; Mean outer canopy height [38]; 25th and 95th percentiles [39,40]; Maximum height [37,41]. |
| CS: canopy cover | Includes metrics describing canopy horizontal structure (e.g., cover, gaps, density, roughness, etc.). | Canopy cover: Number of first returns above a height/total number of first returns [42]; Intensity sum of all non-ground points divided by intensity sum of all points [43]; Canopy gaps and ruggedness [44]; Fractional cover [45]; LiDAR penetration ratio (LPI), as the ratio between terrain points and total points [22]. |
| CS: canopy vertical distribution | Includes metrics describing canopy vertical variation (heterogeneity, complexity, layering, etc.). | Standard deviation of vegetation height [46]; Proportion of LiDAR returns at different layers [47]; Gini coefficient [48]; Simpson index [39]; Vertical gap index, measured as the total distance between individual canopy strata divided by maximum canopy height [43]. |
| Understory structure and shrubland | Describes height, cover, volume and/or contribution of the strata below tree canopy and shrub structure in non-forest habitats. | Index of foliage height diversity, such as the Shannon diversity index for the returns between 0.5 m and 3 m [49]; Density of understory cover: ratio of understory returns to the total number of understory and ground returns [33]; Shrub and snags cover: vegetation returns between 1 and 2.5 m [50]; Understory penetration ratio [36]. |
| Topography | GIS variables commonly derived from LiDAR data for describing surface topography (elevation, slope, aspect, terrain roughness) and derived variables such as soil moisture, solar radiation, etc. | Rugosity [51]; Slope, aspect, curvature [52]; Topographic wetness index [30,53]; Depth-to-water Index [26]; Potential incoming solar radiation [30]. |
3. Results
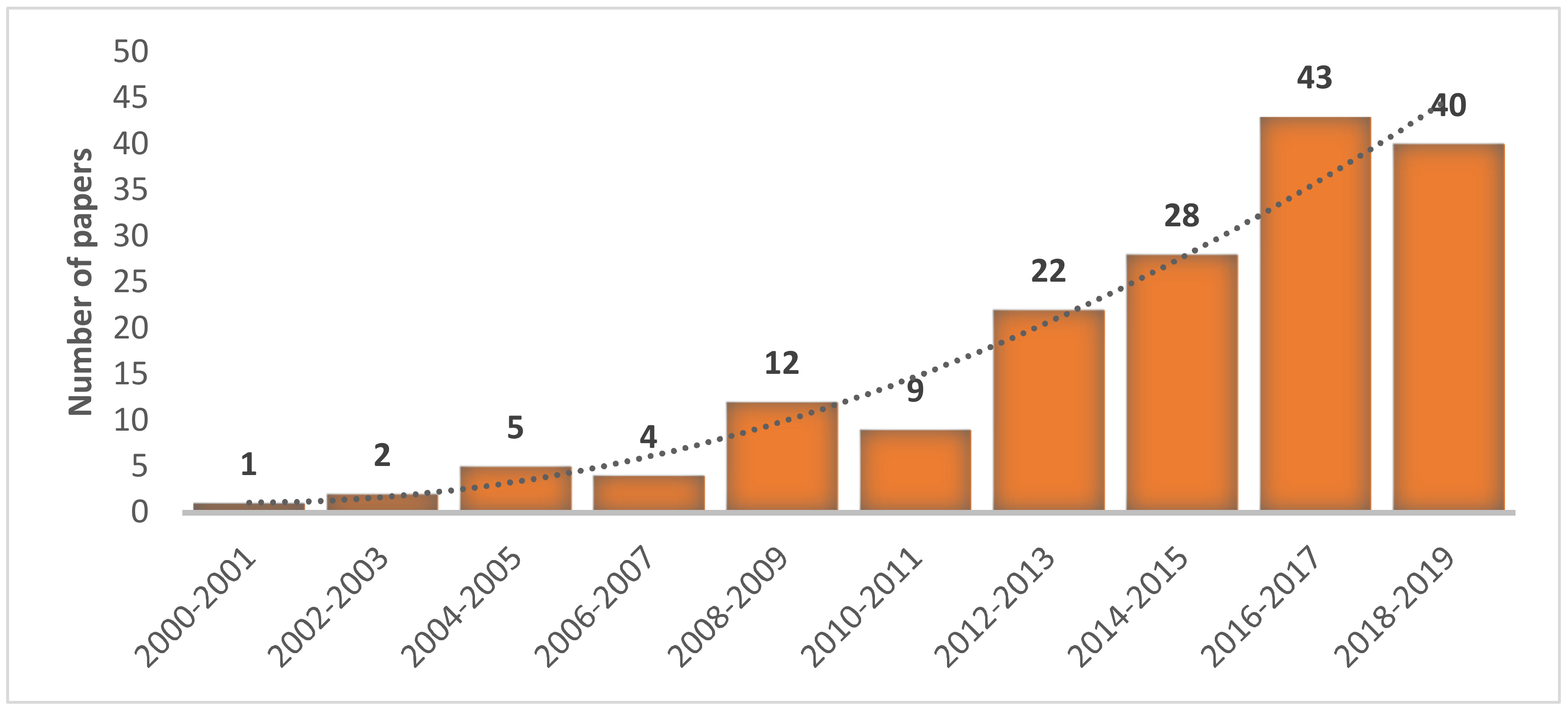
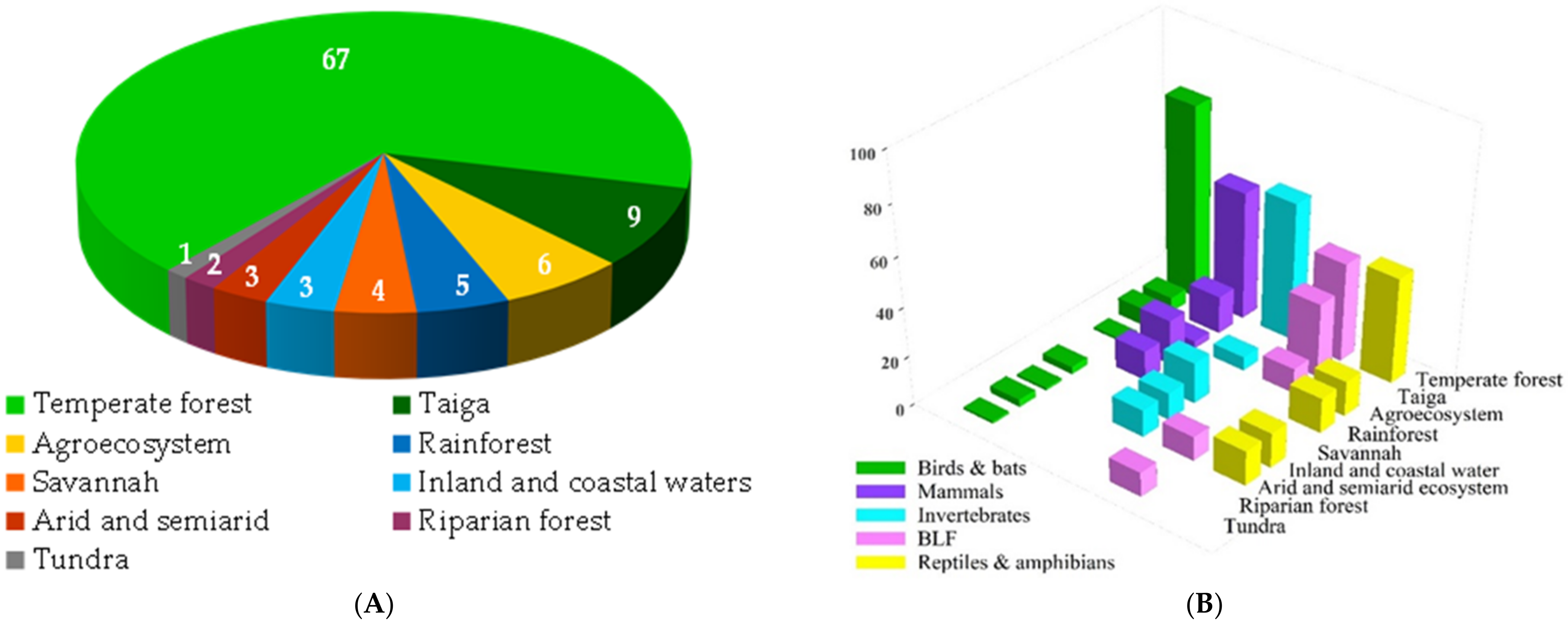
3.1. Geographic Distribution, Taxonomic Groups and Ecosystems Surveyed
3.2. LiDAR Traits: Use versus Performance
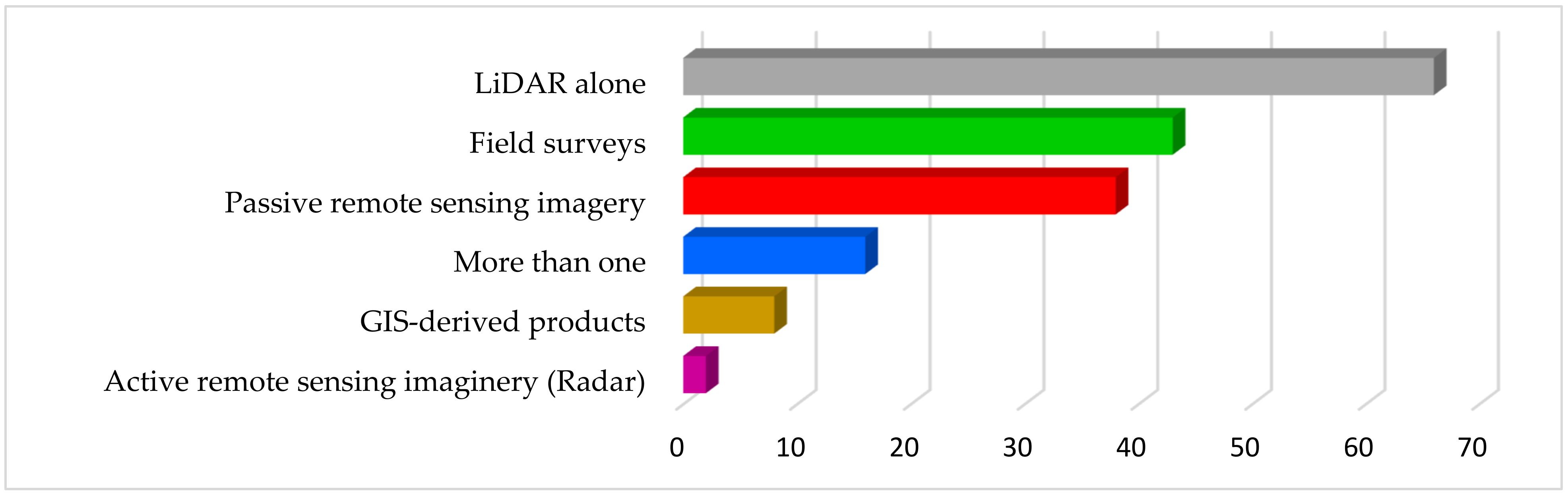
3.3. LiDAR Characteristics
3.4. Use of Multi-Temporal LiDAR and Combination of LiDAR Data with Other Sources
4. Discussion
From Local to Global Analyses Characterizing Terrestrial Ecosystem Structure for Biodiversity with LiDAR Data. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nat. Cell Biol. 2017, 546, 73–81. [Google Scholar] [CrossRef]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodi-Versity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- MacArthur, R.H.; MacArthur, J.W. On Bird Species Diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.L.; Marcot, B.; Mannan, W. Wildlife-Habitat Relationships: Concepts and Applications; Island Press: Washington, DC, USA, 2012. [Google Scholar]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar Remote Sensing for Ecosystem Studies: Lidar, an emerging remote sensing technology that directly measures the three-dimensional distribution of plant canopies, can accurately estimate vegetation structural attributes and should be of particular interest to forest, landscape, and global ecologists. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Vierling, K.T.; Vierling, L.A.; Gould, W.; Martinuzzi, S.; Clawges, R.M. Lidar: Shedding new light on habitat characterization and modeling. Front. Ecol. Environ. 2008, 6, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Roll, U.; Geffen, E.; Yom-Tov, Y. Linking vertebrate species richness to tree canopy height on a global scale. Glob. Ecol. Biogeogr. 2015, 24, 814–825. [Google Scholar] [CrossRef]
- De Thoisy, B.; Fayad, I.; Clément, L.; Barrioz, S.; Poirier, E.; Gond, V. Predators, Prey and Habitat Structure: Can Key Conservation Areas and Early Signs of Population Collapse Be Detected in Neotropical Forests? PLoS ONE 2016, 11, e0165362. [Google Scholar] [CrossRef] [PubMed]
- Dubayah, R.; Blair, J.B.; Goetz, S.; Fatoyinbo, L.; Hansen, M.; Healey, S.; Hofton, M.; Hurtt, G.; Kellner, J.; Luthcke, S.; et al. The Global Ecosystem Dynamics Investigation: High-resolution laser ranging of the Earth’s forests and topography. Sci. Remote Sens. 2020, 1, 100002. [Google Scholar] [CrossRef]
- Müller, J.; Brandl, R.; Brändle, M.; Förster, B.; De Araujo, B.C.; Gossner, M.M.; Ladas, A.; Wagner, M.; Maraun, M.; Schall, P.; et al. LiDAR-derived canopy structure supports the more-individuals hypothesis for arthropod diversity in temperate forests. Oikos 2017, 127, 814–824. [Google Scholar] [CrossRef]
- Jenkins, J.M.A.; Lesmeister, D.B.; Wiens, J.D.; Kane, J.T.; Kane, V.R.; Verschuyl, J. Three-dimensional partitioning of resources by congeneric forest predators with recent sympatry. Sci. Rep. 2019, 9, 6036. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santin, L.; Rudge, M.L.; Bartolo, R.E.; Erskine, P.D. Identifying Species and Monitoring Understorey from UAS-Derived Data: A Literature Review and Future Directions. Drones 2019, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, N.; Perroy, R.; Eiben, J.; Klasner, F. High resolution habitat suitability modelling for an endemic restricted-range Hawaiian insect (Nysius wekiuicola, Hemiptera: Lygaeidae). J. Insect Conserv. 2017, 21, 87–96. [Google Scholar] [CrossRef]
- Maltamo, M.; Næsset, E.; Vauhkonen, J. (Eds.) Forestry Applications of Airborne Laser Scanning; Springer: Berlin/Heidelberg, Germany, 2014; Volume 27, p. 460. [Google Scholar]
- Davies, A.B.; Asner, G. Advances in animal ecology from 3D-LiDAR ecosystem mapping. Trends Ecol. Evol. 2014, 29, 681–691. [Google Scholar] [CrossRef]
- Bakx, T.; Koma, Z.; Seijmonsbergen, H.; Kissling, W.D. Use and categorization of Light Detection and Ranging vegetation metrics in avian diversity and species distribution research. Divers. Distrib. 2019, 25, 1045–1059. [Google Scholar] [CrossRef] [Green Version]
- Dennis, R.L. Just how important are structural elements as habitat components? Indications from a declining lycaenid butterfly with priority conservation status. J. Insect Conserv. 2004, 8, 37–45. [Google Scholar] [CrossRef]
- Jaime-González, C.; Acebes, P.; Mateos, A.; Mezquida, E.T. Bridging gaps: On the performance of airborne LiDAR to model wood mouse-habitat structure relationships in pine forests. PLoS ONE 2017, 12, e0182451. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.B.; Oram, F.; Ancrenaz, M.; Asner, G.P. Combining behavioural and LiDAR data to reveal relationships between canopy structure and orangutan nest site selection in disturbed forests. Biol. Conserv. 2019, 232, 97–107. [Google Scholar] [CrossRef]
- Bombi, P.; Gnetti, V.; D’Andrea, E.; De Cinti, B.; Taglianti, A.V.; Bologna, M.A.; Matteucci, G. Identifying priority sites for insect conservation in forest ecosystems at high resolution: The potential of LiDAR data. J. Insect Conserv. 2019, 23, 689–698. [Google Scholar] [CrossRef]
- Kerfoot, W.C.; Hobmeier, M.M.; Regis, R.; Raman, V.K.; Brooks, C.N.; Shuchman, R.; Sayers, M.; Yousef, F.; Reif, M. Lidar (light detection and ranging) and benthic invertebrate investigations: Migrating tailings threaten Buffalo Reef in Lake Superior. J. Great Lakes Res. 2019, 45, 872–887. [Google Scholar] [CrossRef]
- Evans, L.J.; Davies, A.B.; Goossens, B.; Asner, G.P. Riparian vegetation structure and the hunting behavior of adult estuarine crocodiles. PLoS ONE 2017, 12, e0184804. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.W.; Calhoun, D.L.; Barichivich, W.J.; Walls, S.C. Identifying Small Depressional Wetlands and Using a Topographic Position Index to Infer Hydroperiod Regimes for Pond-Breeding Amphibians. Wetlands 2017, 37, 325–338. [Google Scholar] [CrossRef]
- Bartels, S.F.; Caners, R.T.; Ogilvie, J.; White, B.; Macdonald, S.E. Relating Bryophyte Assemblages to a Remotely Sensed Depth-to-Water Index in Boreal Forests. Front. Plant Sci. 2018, 9, 858. [Google Scholar] [CrossRef] [PubMed]
- Moeslund, J.E.; Zlinszky, A.; Ejrnæs, R.; Brunbjerg, A.K.; Bøcher, P.K.; Svenning, J.; Normand, S.; Ejrnaes, R. Light detection and ranging explains diversity of plants, fungi, lichens, and bryophytes across multiple habitats and large geographic extent. Ecol. Appl. 2019, 29, e01907. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.B.; Baldeck, C.A.; Asner, G.P. Termite mounds alter the spatial distribution of African savanna tree species. J. Biogeogr. 2015, 43, 301–313. [Google Scholar] [CrossRef]
- Boelman, N.; Holbrook, J.; Greaves, H.; Krause, J.; Chmura, H.; Magney, T.; Perez, J.; Eitel, J.; Gough, L.; Vierling, K.; et al. Airborne laser scanning and spectral remote sensing give a bird’s eye perspective on arctic tundra breeding habitat at multiple spatial scales. Remote Sens. Environ. 2016, 184, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Caballero, E.; Román, J.R.; Chamizo, S.; Ramos, B.R.; Cantón, Y. Biocrust landscape-scale spatial distribution is strongly controlled by terrain attributes: Topographic thresholds for colonization in a semiarid badland system. Earth Surf. Process. Landf. 2019, 44, 2771–2779. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Fahrig, L.; Förster, B.; Heurich, M.; Hothorn, T.; Scheipl, F.; Thorn, S.; Müller, J. An experimental test of the habitat-amount hypothesis for saproxylic beetles in a forested region. Ecology 2017, 98, 1613–1622. [Google Scholar] [CrossRef]
- Renner, S.C.; Suarez-Rubio, M.; Kaiser, S.; Nieschulze, J.; Kalko, E.K.; Tschapka, M.; Jung, K. Divergent response to forest structure of two mobile vertebrate groups. For. Ecol. Manag. 2018, 415-416, 129–138. [Google Scholar] [CrossRef]
- Lone, K.; Loe, L.E.; Gobakken, T.; Linnell, J.D.C.; Odden, J.; Remmen, J.; Mysterud, A. Living and dying in a multi-predator landscape of fear: Roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans. Oikos 2014, 123, 641–651. [Google Scholar] [CrossRef]
- Swift, C.E.; Vierling, K.T.; Hudak, A.T.; Vierling, L.A. Relationships among Vegetation Structure, Canopy Composition, and Avian Richness Patterns across an Aspen-Conifer Forest Gradient. Can. J. Remote Sens. 2017, 37, 231–243. [Google Scholar] [CrossRef]
- Sumnall, M.; Peduzzi, A.; Fox, T.R.; Wynne, R.H.; Thomas, V.A.; Cook, B. Assessing the transferability of statistical predictive models for leaf area index between two airborne discrete return LiDAR sensor designs within multiple intensely managed Loblolly pine forest locations in the south-eastern USA. Remote Sens. Environ. 2016, 176, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Levick, S.R.; Heidrich, L.; Magdon, P.; Leutner, B.F.; Wöllauer, S.; Serebryanyk, A.; Nauss, T.; Krzystek, P.; Gossner, M.M.; et al. Radar vision in the mapping of forest biodiversity from space. Nat. Commun. 2019, 10, 4757. [Google Scholar] [CrossRef] [PubMed]
- Thers, H.; Brunbjerg, A.K.; Læssøe, T.; Ejrnæs, R.; Bøcher, P.K.; Svenning, J.-C. Lidar-derived variables as a proxy for fungal species richness and composition in temperate Northern Europe. Remote Sens. Environ. 2017, 200, 102–113. [Google Scholar] [CrossRef]
- Fotis, A.T.; Patel, S.; Chavez, A.S. Habitat-based isolating barriers are not strong in the speciation of ecologically divergent squirrels (Tamiasciurus douglasii and T. hudsonicus). Behav. Ecol. Sociobiol. 2020, 74, 1–14. [Google Scholar] [CrossRef]
- Glad, A.; Reineking, B.; Montadert, M.; Depraz, A.; Monnet, J. Assessing the performance of object-oriented Li DAR predictors for forest bird habitat suitability modeling. Remote Sens. Ecol. Conserv. 2019, 6, 5–19. [Google Scholar] [CrossRef]
- Blomdahl, E.M.; Thompson, C.M.; Kane, J.R.; Kane, V.R.; Churchill, D.; Moskal, L.M.; Lutz, J.A. Forest structure predictive of fisher (Pekania pennanti) dens exists in recently burned forest in Yosemite, California, USA. For. Ecol. Manag. 2019, 444, 174–186. [Google Scholar] [CrossRef]
- Kellner, K.F. Local-scale Habitat Components Driving Bird Abundance in Eastern Deciduous Forests. Am. Midl. Nat. 2018, 180, 52–65. [Google Scholar] [CrossRef]
- Schooler, S.L.; Zald, H. Lidar Prediction of Small Mammal Diversity in Wisconsin, USA. Remote Sens. 2019, 11, 2222. [Google Scholar] [CrossRef] [Green Version]
- Khanaposhtani, M.G.; Gasc, A.; Francomano, D.; Villanueva-Rivera, L.J.; Jung, J.; Mossman, M.J.; Pijanowski, B.C. Effects of highways on bird distribution and soundscape diversity around Aldo Leopold’s shack in Baraboo, Wisconsin, USA. Landsc. Urban Plan. 2019, 192, 103666. [Google Scholar] [CrossRef]
- Carr, A.; Zeale, M.; Weatherall, A.; Froidevaux, J.S.; Jones, G. Ground-based and LiDAR-derived measurements reveal scale-dependent selection of roost characteristics by the rare tree-dwelling bat Barbastella barbastellus. For. Ecol. Manag. 2018, 417, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Ewald, M.; Dupke, C.; Heurich, M.; Muller, J.P.; Reineking, B. LiDAR Remote Sensing of Forest Structure and GPS Telemetry Data Provide Insights on Winter Habitat Selection of European Roe Deer. Forests 2014, 5, 1374–1390. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Gutiérrez, J.; WallisDeVries, M.F.; Marshall, L.; Zelfde, M.V.; Villalobos-Arámbula, A.R.; Boekelo, B.; Bartholomeus, H.; Franzén, M.; Biesmeijer, J.C. Butterflies show different functional and species diversity in relationship to vegetation structure and land use. Glob. Ecol. Biogeogr. 2017, 26, 1126–1137. [Google Scholar] [CrossRef] [Green Version]
- Bernheim, M.; Shanas, U.; Bar-Massada, A. Soil Type Impacts Macrohabitat Selection and Spatiotemporal Activity Patterns of Testudo graeca in an Eastern Mediterranean Ecosystem. Copeia 2019, 107, 332–340. [Google Scholar] [CrossRef]
- Mononen, L.; Auvinen, A.-P.; Packalen, P.; Virkkala, R.; Valbuena, R.; Bohlin, I.; Valkama, J.; Vihervaara, P. Usability of citizen science observations together with airborne laser scanning data in determining the habitat preferences of forest birds. For. Ecol. Manag. 2018, 430, 498–508. [Google Scholar] [CrossRef]
- Lindberg, E.; Roberge, J.-M.; Johansson, T.; Hjältén, J. Can Airborne Laser Scanning (ALS) and Forest Estimates Derived from Satellite Images Be Used to Predict Abundance and Species Richness of Birds and Beetles in Boreal Forest? Remote Sens. 2015, 7, 4233–4252. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, T.J.; Vierling, K.T.; Vogeler, J.; Lonneker, J.; Aycrigg, J. Incorporating Shrub and Snag Specific LiDAR Data into GAP Wildlife Models. J. Fish Wildl. Manag. 2015, 6, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Culver, M.; Gibeaut, J.C.; Shaver, D.J.; Tissot, P.; Starek, M. Using Lidar Data to Assess the Relationship Between Beach Geomorphology and Kemp’s Ridley (Lepidochelys kempii) Nest Site Selection Along Padre Island, TX, United States. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Work, T.T.; Onge, B.S.; Jacobs, J. Response of female beetles to LIDAR derived topographic variables in Eastern boreal mixedwood forests (Coleoptera, Carabidae). ZooKeys 2011, 147, 623–639. [Google Scholar] [CrossRef] [Green Version]
- Svendsen, J.K.; Sell, H.; Bocher, P.K.; Svenning, J. Habitat and nest site preferences of Red-backed Shrike (Lanius collurio) in western Denmark. Ornis Fenn. 2015, 92, 63–75. [Google Scholar]
- Huber, N.; Kéry, M.; Pasinelli, G. Occupancy dynamics of the Wood Warbler Phylloscopus sibilatrix assessed with habitat and remote sensing data. Ibis 2017, 159, 623–637. [Google Scholar] [CrossRef]
- Kortmann, M.; Heurich, M.; Latifi, H.; Rösner, S.; Seidl, R.; Müller, J.; Thorn, S. Forest structure following natural disturbances and early succession provides habitat for two avian flagship species, capercaillie (Tetrao urogallus) and hazel grouse (Tetrastes bonasia). Biol. Conserv. 2018, 226, 81–91. [Google Scholar] [CrossRef]
- Taillie, P.J.; Moorman, C.E.; Smart, L.; Pacifici, K. Bird community shifts associated with saltwater exposure in coastal forests at the leading edge of rising sea level. PLoS ONE 2019, 14, e0216540. [Google Scholar] [CrossRef] [Green Version]
- Paulson, E.L.; Martin, A.P. Discerning invasion history in an ephemerally connected system: Landscape genetics of Procambarus clarkii in Ash Meadows, Nevada. Biol. Invasions 2013, 16, 1719–1734. [Google Scholar] [CrossRef]
- Hogan, S.; Reidenbach, M. Quantifying and mapping intertidal oyster reefs utilizing LiDAR-based remote sensing. Mar. Ecol. Prog. Ser. 2019, 630, 83–99. [Google Scholar] [CrossRef]
- Müller, J.; Bae, S.; Röder, J.; Chao, A.; Didham, R.K. Airborne LiDAR reveals context dependence in the effects of canopy architecture on arthropod diversity. For. Ecol. Manag. 2014, 312, 129–137. [Google Scholar] [CrossRef]
- Zellweger, F.; Baltensweiler, A.; Ginzler, C.; Roth, T.; Braunisch, V.; Bugmann, H.; Bollmann, K. Environmental predictors of species richness in forest landscapes: Abiotic factors versus vegetation structure. J. Biogeogr. 2016, 43, 1080–1090. [Google Scholar] [CrossRef]
- Cooper, W.J.; McShea, W.J.; Luther, D.A.; Forrester, T. Incorporating local habitat heterogeneity and productivity measures when modelling vertebrate richness. Environ. Conserv. 2019, 47, 7–14. [Google Scholar] [CrossRef]
- Zellweger, F.; Roth, T.; Bugmann, H.; Bollmann, K. Beta diversity of plants, birds and butterflies is closely associated with climate and habitat structure. Glob. Ecol. Biogeogr. 2017, 26, 898–906. [Google Scholar] [CrossRef]
- Palminteri, S.; Powell, G.V.; Asner, G.; Peres, C. LiDAR measurements of canopy structure predict spatial distribution of a tropical mature forest primate. Remote Sens. Environ. 2012, 127, 98–105. [Google Scholar] [CrossRef]
- McLean, K.A.; Trainor, A.M.; Asner, G.P.; Crofoot, M.C.; Hopkins, M.E.; Campbell, C.J.; Martin, R.E.; Knapp, D.E.; Jansen, P. Movement patterns of three arboreal primates in a Neotropical moist forest explained by LiDAR-estimated canopy structure. Landsc. Ecol. 2016, 31, 1849–1862. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Cheyne, S.M.; Smith, D.E. How conspecific primates use their habitats: Surviving in an anthropogenically-disturbed forest in Central Kalimantan, Indonesia. Ecol. Indic. 2018, 87, 167–177. [Google Scholar] [CrossRef]
- Seaman, D.J.; Bernard, H.; Ancrenaz, M.; Coomes, D.; Swinfield, T.; Milodowski, D.T.; Humle, T.; Struebig, M.J. Densities of Bornean orang-utans ( Pongo pygmaeus morio ) in heavily degraded forest and oil palm plantations in Sabah, Borneo. Am. J. Primatol. 2019, 81, e23030. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.B.; Tambling, C.; Kerley, G.I.H.; Asner, G.P. Effects of Vegetation Structure on the Location of Lion Kill Sites in African Thicket. PLoS ONE 2016, 11, e0149098. [Google Scholar] [CrossRef] [Green Version]
- Loarie, S.R.; Tambling, C.; Asner, G. Lion hunting behaviour and vegetation structure in an African savanna. Anim. Behav. 2013, 85, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Owen-Smith, N.; Loarie, S.R.; Davies, A.; Le Roux, E.; Levick, S.R. Habitat differences do not explain population declines of sable antelope in an African savanna. J. Zool. 2015, 297, 225–234. [Google Scholar] [CrossRef]
- Davies, A.B.; Tambling, C.; Kerley, G.I.; Asner, G.P. Limited spatial response to direct predation risk by African herbivores following predator reintroduction. Ecol. Evol. 2016, 6, 5728–5748. [Google Scholar] [CrossRef]
- Levick, S.R.; Asner, G.; Chadwick, O.A.; Khomo, L.M.; Rogers, K.H.; Hartshorn, A.; Kennedy-Bowdoin, T.; Knapp, D.E. Regional insight into savanna hydrogeomorphology from termite mounds. Nat. Commun. 2010, 1, 65. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.B.; Levick, S.; Asner, G.; Robertson, M.P.; van Rensburg, B.; Parr, C. Spatial variability and abiotic determinants of termite mounds throughout a savanna catchment. Ecography 2014, 37, 852–862. [Google Scholar] [CrossRef]
- Davenport, I.; Bradbury, R.B.; Anderson, G.Q.A.; Hayman, G.R.F.; Krebs, J.R.; Mason, D.C.; Wilson, J.D.; Veck, N.J. Improving bird population models using airborne remote sensing. Int. J. Remote Sens. 2000, 21, 2705–2717. [Google Scholar] [CrossRef]
- Mason, D.C.; Anderson, G.Q.A.; Bradbury, R.B.; Cobby, D.M.; Davenport, I.; Vandepoll, M.; Wilson, J.D. Measurement of habitat predictor variables for organism-habitat models using remote sensing and image segmentation. Int. J. Remote Sens. 2003, 24, 2515–2532. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, R.B.; Hill, R.A.; Mason, D.C.; Hinsley, S.A.; Wilson, J.D.; Balzter, H.; Anderson, G.Q.A.; Whittingham, M.J.; Davenport, I.; Bellamy, P.E. Modelling relationships between birds and vegetation structure using airborne LiDAR data: A review with case studies from agricultural and woodland environments. Ibis 2005, 147, 443–452. [Google Scholar] [CrossRef]
- Redhead, J.; Pywell, R.; Bellamy, P.; Broughton, R.; Hill, R.; Hinsley, S. Great tits Parus major and blue tits Cyanistes caeruleus as indicators of agri-environmental habitat quality. Agric. Ecosyst. Environ. 2013, 178, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Trainor, A.M.; Walters, J.R.; Morris, W.F.; Sexton, J.; Moody, A. Empirical estimation of dispersal resistance surfaces: A case study with red-cockaded woodpeckers. Landsc. Ecol. 2013, 28, 755–767. [Google Scholar] [CrossRef]
- Melin, M.; Hinsley, S.A.; Broughton, R.K.; Bellamy, P.; Hill, R.A. Living on the edge: Utilising lidar data to assess the importance of vegetation structure for avian diversity in fragmented woodlands and their edges. Landsc. Ecol. 2018, 33, 895–910. [Google Scholar] [CrossRef] [Green Version]
- Sillero, N.; Gonçalves-Seco, L. Spatial structure analysis of a reptile community with airborne LiDAR data. Int. J. Geogr. Inf. Sci. 2014, 28, 1709–1722. [Google Scholar] [CrossRef]
- Zabihi, K.; Driese, K.L.; Paige, G.B.; Hild, A.K. Application of Ground-Based Lidar and Gap Intercept Measurements to Quantify a Shrub Configuration Metric within Greater Sage-Grouse Nesting Habitat. West. N. Am. Nat. 2019, 79, 500–514. [Google Scholar] [CrossRef]
- Müller, J.; Stadler, J.; Brandl, R. Composition versus physiognomy of vegetation as predictors of bird assemblages: The role of lidar. Remote Sens. Environ. 2010, 114, 490–495. [Google Scholar] [CrossRef]
- Vierling, K.T.; Bassler, C.; Brandl, R.; Vierling, L.A.; Weis, I.; Müller, J. Spinning a laser web: Predicting spider distributions using LiDAR. Ecol. Appl. 2011, 21, 577–588. [Google Scholar] [CrossRef]
- Renner, S.C.; Baur, S.; Possler, A.; Winkler, J.; Kalko, E.K.V.; Bates, P.J.J.; Mello, M. Food Preferences of Winter Bird Communities in Different Forest Types. PLoS ONE 2012, 7, e53121. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Bonardi, A.; Mucher, S.; Gilissen, N.L.; Schioppa, E.P. How many predictors in species distribution models at the landscape scale? Land use versus LiDAR-derived canopy height. Int. J. Geogr. Inf. Sci. 2014, 28, 1723–1739. [Google Scholar] [CrossRef]
- Sasaki, T.; Imanishi, J.; Fukui, W.; Morimoto, Y. Fine-scale characterization of bird habitat using airborne LiDAR in an urban park in Japan. Urban For. Urban Green. 2016, 17, 16–22. [Google Scholar] [CrossRef]
- Müller, J.; Brandl, R.; Buchner, J.; Pretzsch, H.; Seifert, S.; Strätz, C.; Veith, M.; Fenton, B. From ground to above canopy—Bat activity in mature forests is driven by vegetation density and height. For. Ecol. Manag. 2013, 306, 179–184. [Google Scholar] [CrossRef]
- Hill, R.A.; Hinsley, S.A. Airborne Lidar for Woodland Habitat Quality Monitoring: Exploring the Significance of Lidar Data Characteristics when Modelling Organism-Habitat Relationships. Remote Sens. 2015, 7, 3446–3466. [Google Scholar] [CrossRef] [Green Version]
- Vierling, K.T.; Swift, C.E.; Hudak, A.T.; Vogeler, J.C.; Vierling, L.A. How much does the time lag between wildlife field-data collection and LiDAR-data acquisition matter for studies of animal distributions? A case study using bird communities. Remote Sens. Lett. 2014, 5, 185–193. [Google Scholar] [CrossRef]
- Peura, M.; Gonzalez, R.S.; Müller, J.; Heurich, M.; Vierling, L.A.; Mönkkönen, M.; Bässler, C. Mapping a ‘cryptic kingdom’: Performance of lidar derived environmental variables in modelling the occurrence of forest fungi. Remote Sens. Environ. 2016, 186, 428–438. [Google Scholar] [CrossRef]
- Hull, I.T.; Shipley, L.A. Testing the Ability of Airborne LiDAR to Measure Forage Resources for Wild Ungulates in Conifer Forests. J. For. 2019, 117, 492–503. [Google Scholar] [CrossRef]
- Tweedy, P.J.; Moriarty, K.M.; Bailey, J.D.; Epps, C.W. Using fine scale resolution vegetation data from LiDAR and ground-based sampling to predict Pacific marten resting habitat at multiple spatial scales. For. Ecol. Manag. 2019, 452, 117556. [Google Scholar] [CrossRef]
- Dunkin, L.; Reif, M.; Altman, S.; Swannack, T. A Spatially Explicit, Multi-Criteria Decision Support Model for Loggerhead Sea Turtle Nesting Habitat Suitability: A Remote Sensing-Based Approach. Remote Sens. 2016, 8, 573. [Google Scholar] [CrossRef] [Green Version]
- Halls, J.N.; Randall, A.L. Nesting Patterns of Loggerhead Sea Turtles (Caretta caretta): Development of a Multiple Regression Model Tested in North Carolina, USA. ISPRS Int. J. Geo-Inf. 2018, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Lane, C.; Liu, H. An Effective Method for Detecting Potential Woodland Vernal Pools Using High-Resolution LiDAR Data and Aerial Imagery. Remote Sens. 2014, 6, 11444–11467. [Google Scholar] [CrossRef] [Green Version]
- Ehmke, G.; Maguire, G.S.; Bird, T.; Ierodiaconou, D.; Weston, M.A. An obligate beach bird selects sub-, inter- and supra-tidal habitat elements. Estuar. Coast. Shelf Sci. 2016, 181, 266–276. [Google Scholar] [CrossRef]
- Pittman, S.; Costa, B.M.; Battista, T.A. Using Lidar Bathymetry and Boosted Regression Trees to Predict the Diversity and Abundance of Fish and Corals. J. Coast. Res. 2009, 10053, 27–38. [Google Scholar] [CrossRef]
- Casella, E.; Collin, A.; Harris, D.; Ferse, S.; Bejarano, S.; Parravicini, V.; Hench, J.L.; Rovere, A. Mapping coral reefs using consumer-grade drones and structure from motion photogrammetry techniques. Coral Reefs 2017, 36, 269–275. [Google Scholar] [CrossRef]
- Collin, A.; Ramambason, C.; Pastol, Y.; Casella, E.; Rovere, A.; Thiault, L.; Espiau, B.; Siu, G.; Lerouvreur, F.; Nakamura, N.; et al. Very high resolution mapping of coral reef state using airborne bathymetric LiDAR surface-intensity and drone imagery. Int. J. Remote Sens. 2018, 39, 5676–5688. [Google Scholar] [CrossRef] [Green Version]
- Wedding, L.M.; Friedlander, A.M.; McGranaghan, M.; Yost, R.S.; Monaco, M.E. Using bathymetric lidar to define nearshore benthic habitat complexity: Implications for management of reef fish assemblages in Hawaii. Remote Sens. Environ. 2008, 112, 4159–4165. [Google Scholar] [CrossRef]
- Wedding, L.M.; Jorgensen, S.; Lepczyk, C.A.; Friedlander, A.M. Remote sensing of three-dimensional coral reef structure enhances predictive modeling of fish assemblages. Remote Sens. Ecol. Conserv. 2019, 5, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Pimm, S.L.; Alibhai, S.; Bergl, R.; Dehgan, A.; Giri, C.; Jewell, Z.; Joppa, L.; Kays, R.; Loarie, S. Emerging Technologies to Conserve Biodiversity. Trends Ecol. Evol. 2015, 30, 685–696. [Google Scholar] [CrossRef]
- Zielewska-Büttner, K.; Heurich, M.; Müller, J.; Braunisch, V. Remotely Sensed Single Tree Data Enable the Determination of Habitat Thresholds for the Three-Toed Woodpecker (Picoides tridactylus). Remote Sens. 2018, 10, 1972. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, L.; Giam, X.; Papeş, M.; Sheldon, K.S. Metrics of Lidar-Derived 3D Vegetation Structure Reveal Contrasting Effects of Horizontal and Vertical Forest Heterogeneity on Bird Species Richness. Remote Sens. 2019, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- Bergen, K.M.; Goetz, S.; Dubayah, R.O.; Henebry, G.; Hunsaker, C.T.; Imhoff, M.L.; Nelson, R.F.; Parker, G.; Radeloff, V.C. Remote sensing of vegetation 3-D structure for biodiversity and habitat: Review and implications for lidar and radar spaceborne missions. J. Geophys. Res. Space Phys. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Valbuena, R.; O’Connor, B.; Zellweger, F.; Simonson, W.; Vihervaara, P.; Maltamo, M.; Silva, C.; de Almeida, D.R.A.; Danks, F.; Morsdorf, F.; et al. Standardizing Ecosystem Morphological Traits from 3D Information Sources. Trends Ecol. Evol. 2020, 35, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.J.; Dennison, P.E.; Hudak, A.T.; Parham, L.M.; Butler, B.W. Quantifying understory vegetation density using small-footprint airborne lidar. Remote Sens. Environ. 2018, 215, 330–342. [Google Scholar] [CrossRef]
- Hagar, J.C.; Yost, A.; Haggerty, P.K. Incorporating LiDAR metrics into a structure-based habitat model for a canopy-dwelling species. Remote Sens. Environ. 2020, 236, 111499. [Google Scholar] [CrossRef]
- Keim, J.L.; DeWitt, P.D.; Fitzpatrick, J.J.; Jenni, N.S. Estimating plant abundance using inflated beta distributions: Applied learnings from a lichen-caribou ecosystem. Ecol. Evol. 2016, 7, 486–493. [Google Scholar] [CrossRef]
- Sovern, S.G.; Lesmeister, D.; Dugger, K.M.; Pruett, M.S.; Davis, R.J.; Jenkins, J.M. Activity center selection by northern spotted owls. J. Wildl. Manag. 2019, 83, 714–727. [Google Scholar] [CrossRef] [Green Version]
- Linnell, M.A.; Davis, R.J.; Lesmeister, D.; Swingle, J.K. Conservation and relative habitat suitability for an arboreal mammal associated with old forest. For. Ecol. Manag. 2017, 402, 1–11. [Google Scholar] [CrossRef]
- Gossner, M.M.; Wende, B.; Levick, S.; Schall, P.; Floren, A.; Linsenmair, K.E.; Steffan-Dewenter, I.; Schulze, E.-D.; Weisser, W. Deadwood enrichment in European forests–Which tree species should be used to promote saproxylic beetle diversity? Biol. Conserv. 2016, 201, 92–102. [Google Scholar] [CrossRef]
- Korpela, I.; Haapanen, R.; Korrensalo, A.; Tuittila, E.; Vesala, T. Fine-resolution mapping of microforms of a boreal bog using aerial images and waveform-recording LiDAR. Mires Peat 2020, 26, 3. [Google Scholar] [CrossRef]
- Hess, A.N.; Falkowski, M.J.; Webster, C.R.; Storer, A.J.; Pocewicz, A.; Martinuzzi, S. Employing lidar data to identify butterfly habitat characteristics of four contrasting butterfly species across a diverse landscape. Remote Sens. Lett. 2013, 4, 354–363. [Google Scholar] [CrossRef]
- Kroeger, A.J.; Moorman, C.E.; Lashley, M.A.; Chitwood, M.C.; Harper, C.A.; DePerno, C.S. White-tailed deer use of overstory hardwoods in longleaf pine woodlands. For. Ecol. Manag. 2020, 464, 118046. [Google Scholar] [CrossRef]
- Klein, J.; Haverkamp, P.J.; Lindberg, E.; Griesser, M.; Eggers, S. Remotely sensed forest understory density and nest predator occurrence interact to predict suitable breeding habitat and the occurrence of a resident boreal bird species. Ecol. Evol. 2020, 10, 2238–2252. [Google Scholar] [CrossRef] [PubMed]
- Vogeler, J.C.; Yang, Z.; Cohen, W.B. Mapping Suitable Lewis’s Woodpecker Nesting Habitat in a Post-Fire Landscape. Northwest Sci. 2016, 90, 421–432. [Google Scholar] [CrossRef]
- Vepakomma, U.; Cormier, D. Potential of multi-temporal uav-borne lidar in assessing effectiveness of silvicultural treatments. ISPRS-Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2017, XLII-2/W6, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, R.A.; Muller-Landau, H.; Rahman, K.A.; Bebber, D.P.; Bin, Y.; Bohlman, S.A.; Bourg, N.; Brinks, J.; Bunyavejchewin, S.; Butt, N.; et al. Scale-dependent relationships between tree species richness and ecosystem function in forests. J. Ecol. 2013, 101, 1214–1224. [Google Scholar] [CrossRef]
- Kakaes, K.; Greenwood, F.; Lippincott, M.; Dosemagen, S.; Meier, P.; Wich, S. Drones and Aerial Observation: New Technologies for Property Rights, Human Rights, and Global Development: A Primer; New America: Washington, DC, USA, 2015; p. 103. [Google Scholar]
- Brack, I.V.; Kindel, A.; Oliveira, L.F.B. Detection errors in wildlife abundance estimates from Unmanned Aerial Systems (UAS) surveys: Synthesis, solutions, and challenges. Methods Ecol. Evol. 2018, 9, 1864–1873. [Google Scholar] [CrossRef]
- Gomes Pereira, L.; Fernandez, P.; Mourato, S.; Matos, J.; Mayer, C.; Marques, F. Quality Control of Outsourced LiDAR Data Acquired with a UAV: A Case Study. Remote Sens. 2021, 13, 419. [Google Scholar] [CrossRef]
- Calders, K.; Adams, J.; Armston, J.; Bartholomeus, H.; Bauwens, S.; Bentley, L.P.; Chave, J.; Danson, F.M.; Demol, M.; Disney, M.; et al. Terrestrial laser scanning in forest ecology: Expanding the horizon. Remote Sens. Environ. 2020, 251, 112102. [Google Scholar] [CrossRef]
- Kokalj, Ž.; Mast, J. Space lidar for archaeology? Reanalyzing GEDI data for detection of ancient Maya buildings. J. Archaeol. Sci. Rep. 2021, 36, 102811. [Google Scholar] [CrossRef]
- Li, W.; Niu, Z.; Shang, R.; Qin, Y.; Wang, L.; Chen, H. High-resolution mapping of forest canopy height using machine learning by coupling ICESat-2 LiDAR with Sentinel-1, Sentinel-2 and Landsat-8 data. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102163. [Google Scholar] [CrossRef]
- Ma, X.; Mahecha, M.D.; Migliavacca, M.; Van Der Plas, F.; Benavides, R.; Ratcliffe, S.; Kattge, J.; Richter, R.; Musavi, T.; Baeten, L.; et al. Inferring plant functional diversity from space: The potential of Sentinel-2. Remote Sens. Environ. 2019, 233, 111368. [Google Scholar] [CrossRef]
- Zhang, Y.; Migliavacca, M.; Penuelas, J.; Ju, W. Advances in hyperspectral remote sensing of vegetation traits and functions. Remote Sens. Environ. 2021, 252, 112121. [Google Scholar] [CrossRef]
- Marvin, D.C.; Koh, L.P.; Lynam, A.; Wich, S.; Davies, A.; Krishnamurthy, R.; Stokes, E.; Starkey, R.; Asner, G.P. Integrating technologies for scalable ecology and conservation. Glob. Ecol. Conserv. 2016, 7, 262–275. [Google Scholar] [CrossRef] [Green Version]




| Canopy Height | Canopy Cover | Canopy Vertical Distribution | Understory Structure | Terrain Topography | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | |
| Birds | 68 | 68.7 | 86.8 | 66 | 66.7 | 92.4 | 49 | 49.5 | 77.6 | 39 | 39.4 | 76.9 | 17 | 17.2 | 76.5 |
| Mammals | 23 | 69.7 | 73.9 | 21 | 63.6 | 90.5 | 16 | 48.5 | 68.8 | 11 | 33.3 | 81.8 | 9 | 27.3 | 88.9 |
| R&A | 1 | 14.3 | 100 | 2 | 28.6 | 100 | 0 | 0 | 5 | 71.4 | 100 | ||||
| Invertebrates | 7 | 38.9 | 85.7 | 5 | 27.8 | 100 | 5 | 27.8 | 100 | 3 | 16.7 | 100 | 12 | 66.7 | 91.7 |
| BLF | 4 | 40 | 100 | 4 | 40 | 100 | 4 | 40 | 50 | 2 | 20 | 100 | 8 | 80 | 100 |
| Several | 5 | 83.3 | 80 | 3 | 50 | 100 | 4 | 66.7 | 50 | 3 | 50 | 100 | 1 | 16.7 | 0 |
| Canopy Height | Canopy Cover | Canopy Vertical Distribution | Understory Structure | Terrain Topography | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | n | Use | Sig. | |
| Temperate forest | 72 | 62.1 | 84.7 | 76 | 65.5 | 93.4 | 57 | 49.1 | 77.2 | 44 | 37.9 | 75 | 30 | 25.9 | 90 |
| Rainforest | 8 | 100 | 100 | 4 | 50 | 75 | 5 | 62.5 | 60 | 0 | 1 | 12.5 | 100 | ||
| Taiga | 11 | 73.3 | 90.9 | 9 | 60 | 100 | 9 | 60 | 55.6 | 7 | 46.7 | 100 | 1 | 6.7 | 100 |
| Riparian forest | 2 | 66.7 | 100 | 1 | 33.3 | 100 | 2 | 66.7 | 100 | 1 | 33.3 | 100 | 0 | ||
| Tundra | 1 | 50 | 0 | 1 | 50 | 100 | 0 | 0 | 1 | 50 | 100 | ||||
| Savannah | 4 | 57.1 | 25 | 3 | 42.9 | 100 | 1 | 14.3 | 100 | 1 | 14.3 | 100 | 5 | 71.4 | 60 |
| Agroecosystem | 9 | 81.8 | 88.9 | 5 | 45.5 | 80 | 4 | 36.4 | 75 | 4 | 36.4 | 100 | 5 | 45.5 | 60 |
| Arid & semi-arid ecosystem | 0 | 2 | 40 | 100 | 0 | 1 | 20 | 100 | 4 | 80 | 100 | ||||
| Inland and coastal waters | 1 | 16.7 | 100 | 0 | 0 | 0 | 5 | 83.3 | 100 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acebes, P.; Lillo, P.; Jaime-González, C. Disentangling LiDAR Contribution in Modelling Species–Habitat Structure Relationships in Terrestrial Ecosystems Worldwide. A Systematic Review and Future Directions. Remote Sens. 2021, 13, 3447. https://doi.org/10.3390/rs13173447
Acebes P, Lillo P, Jaime-González C. Disentangling LiDAR Contribution in Modelling Species–Habitat Structure Relationships in Terrestrial Ecosystems Worldwide. A Systematic Review and Future Directions. Remote Sensing. 2021; 13(17):3447. https://doi.org/10.3390/rs13173447
Chicago/Turabian StyleAcebes, Pablo, Paula Lillo, and Carlos Jaime-González. 2021. "Disentangling LiDAR Contribution in Modelling Species–Habitat Structure Relationships in Terrestrial Ecosystems Worldwide. A Systematic Review and Future Directions" Remote Sensing 13, no. 17: 3447. https://doi.org/10.3390/rs13173447
APA StyleAcebes, P., Lillo, P., & Jaime-González, C. (2021). Disentangling LiDAR Contribution in Modelling Species–Habitat Structure Relationships in Terrestrial Ecosystems Worldwide. A Systematic Review and Future Directions. Remote Sensing, 13(17), 3447. https://doi.org/10.3390/rs13173447