
Response of Vegetation Photosynthetic Phenology to Urbanization in Dongting Lake Basin, China

 ,
,
Abstract
:1. Introduction
2. Data and Methods
2.1. Study Area
2.2. Data Sources
2.3. Vegetation Photosynthesis Extraction
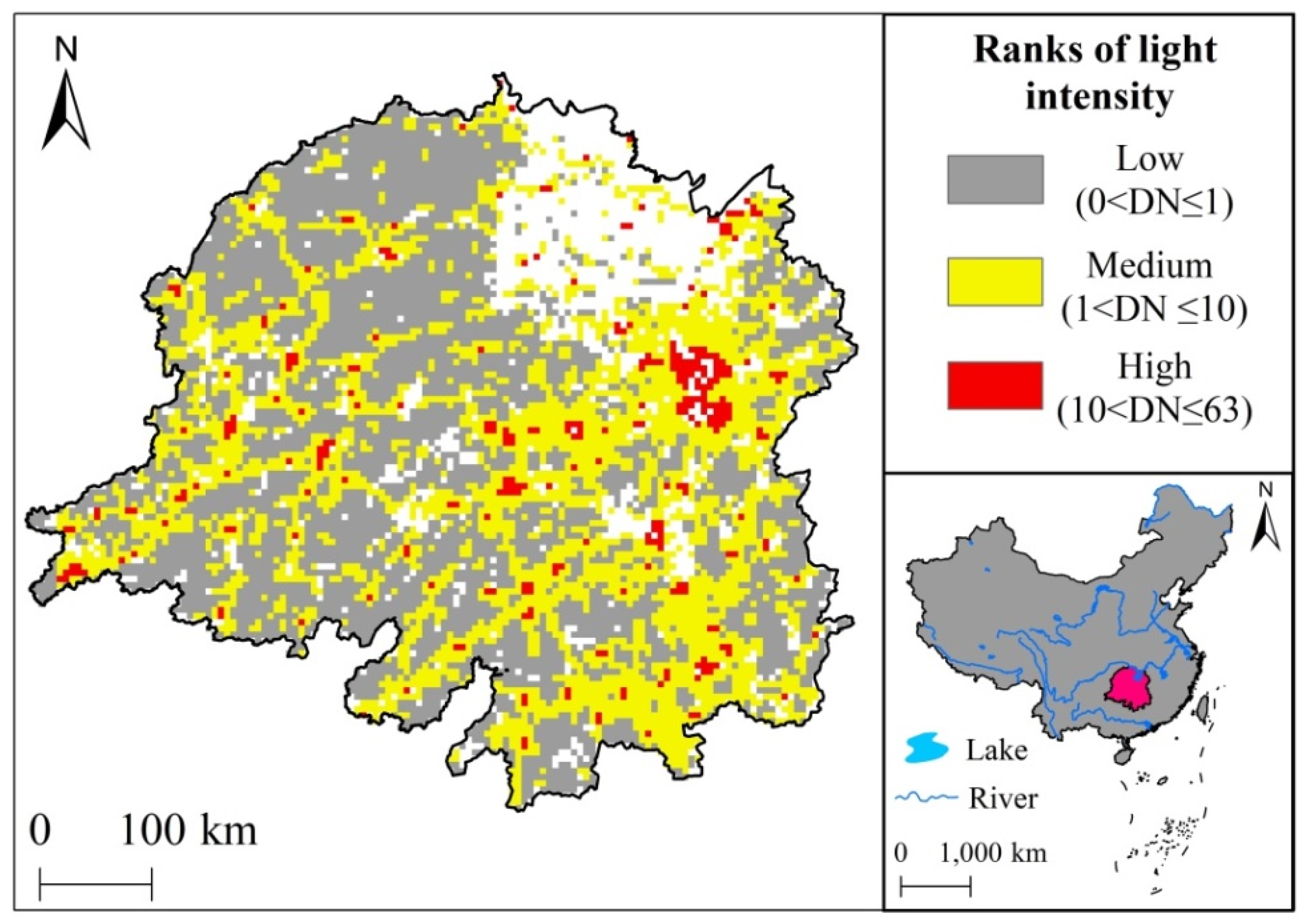
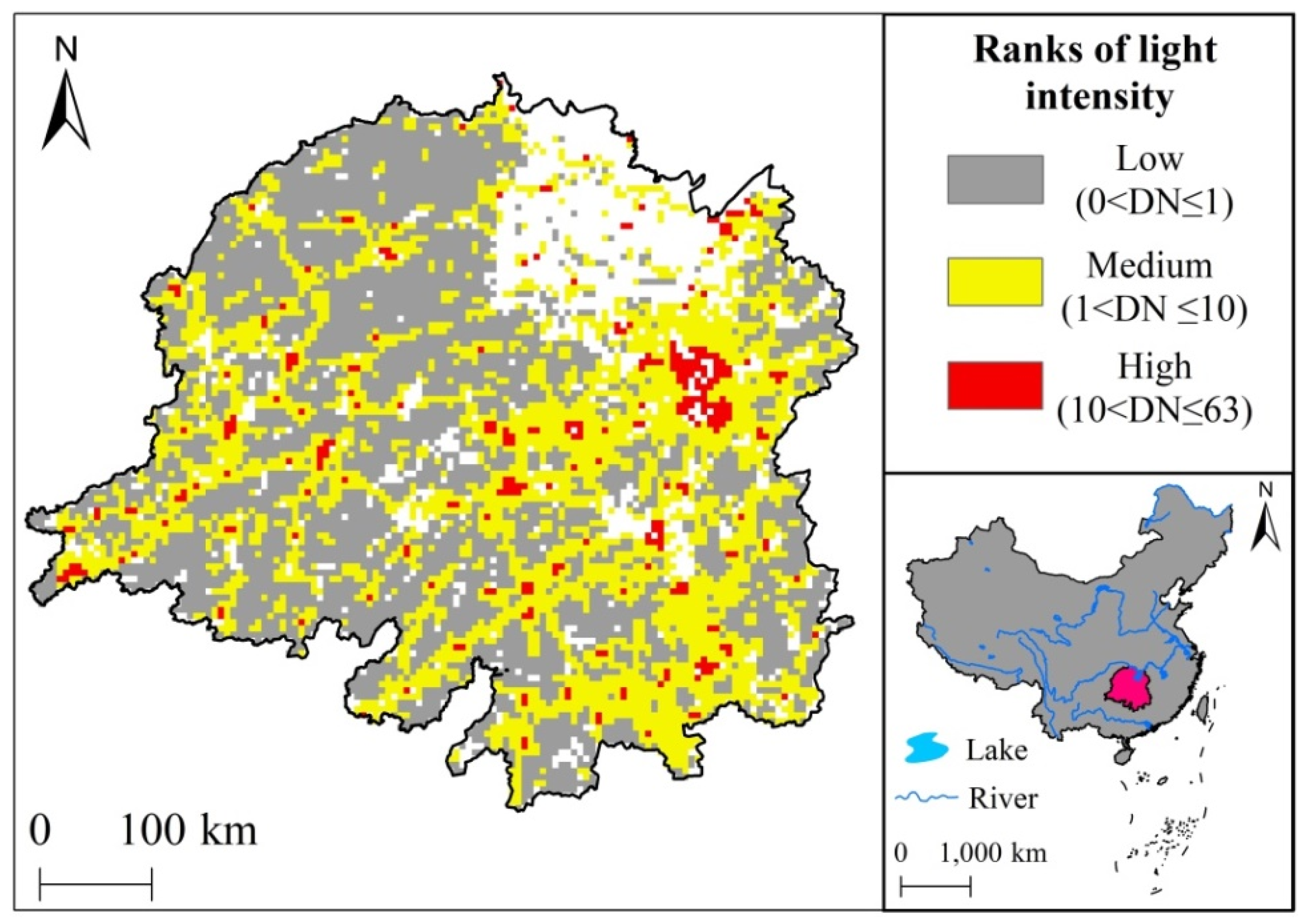
2.4. DMSP Nighttime Light Classification Statistics
2.5. Analysis
3. Results
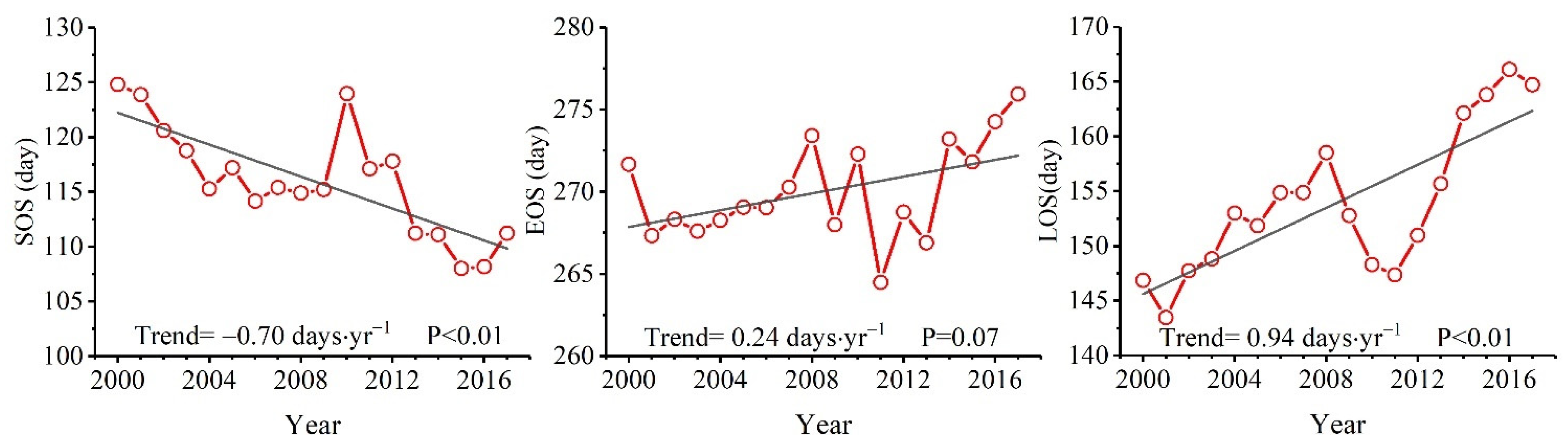
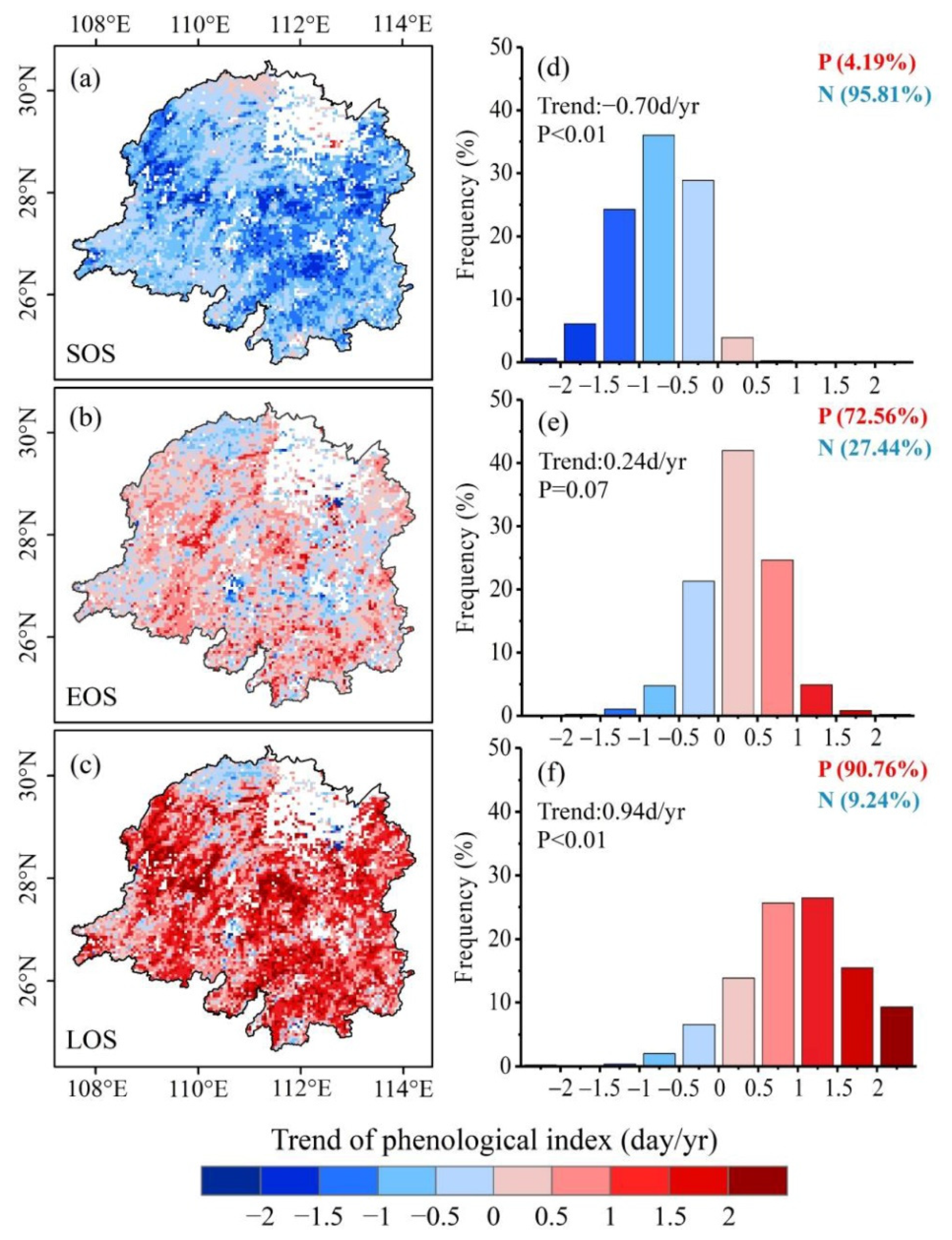
3.1. Vegetation Photosynthetic Phenology Trends in Dongting Lake Basin
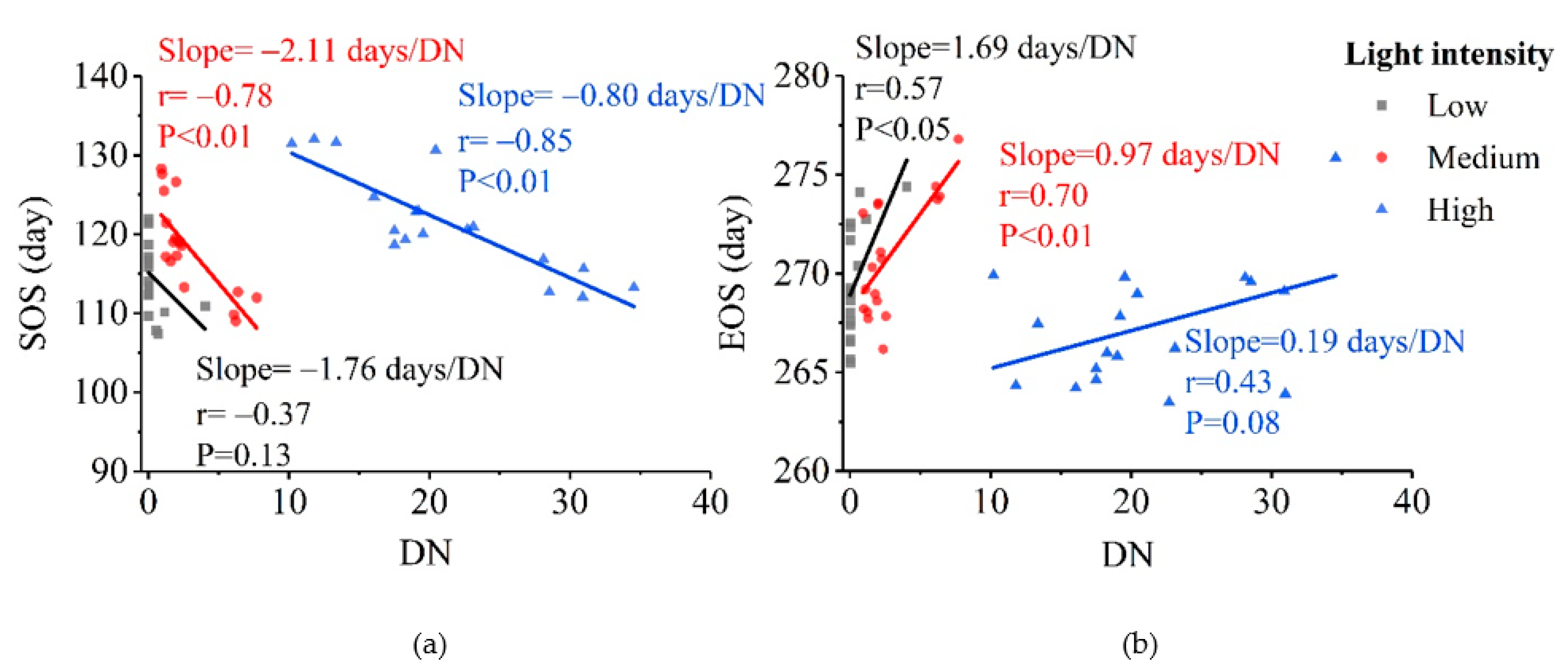
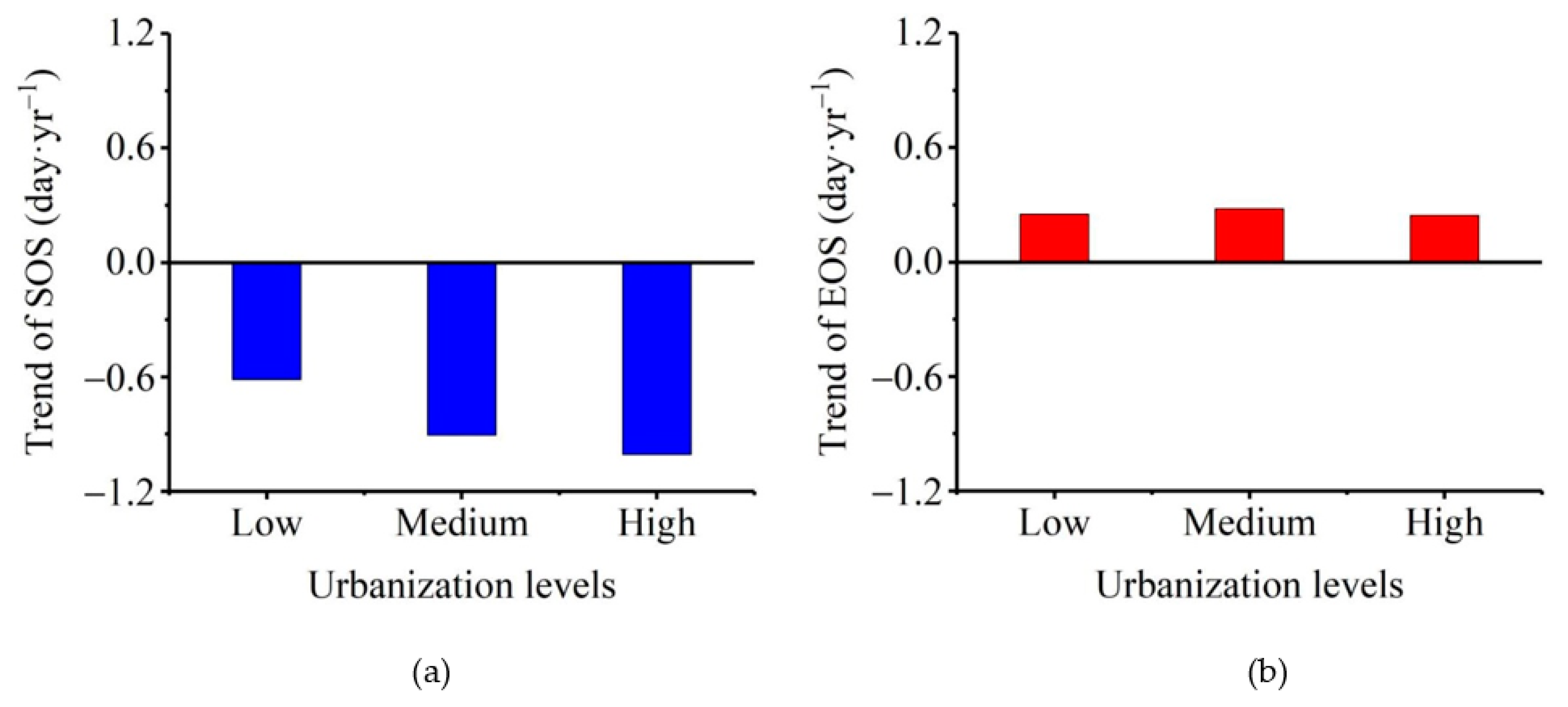
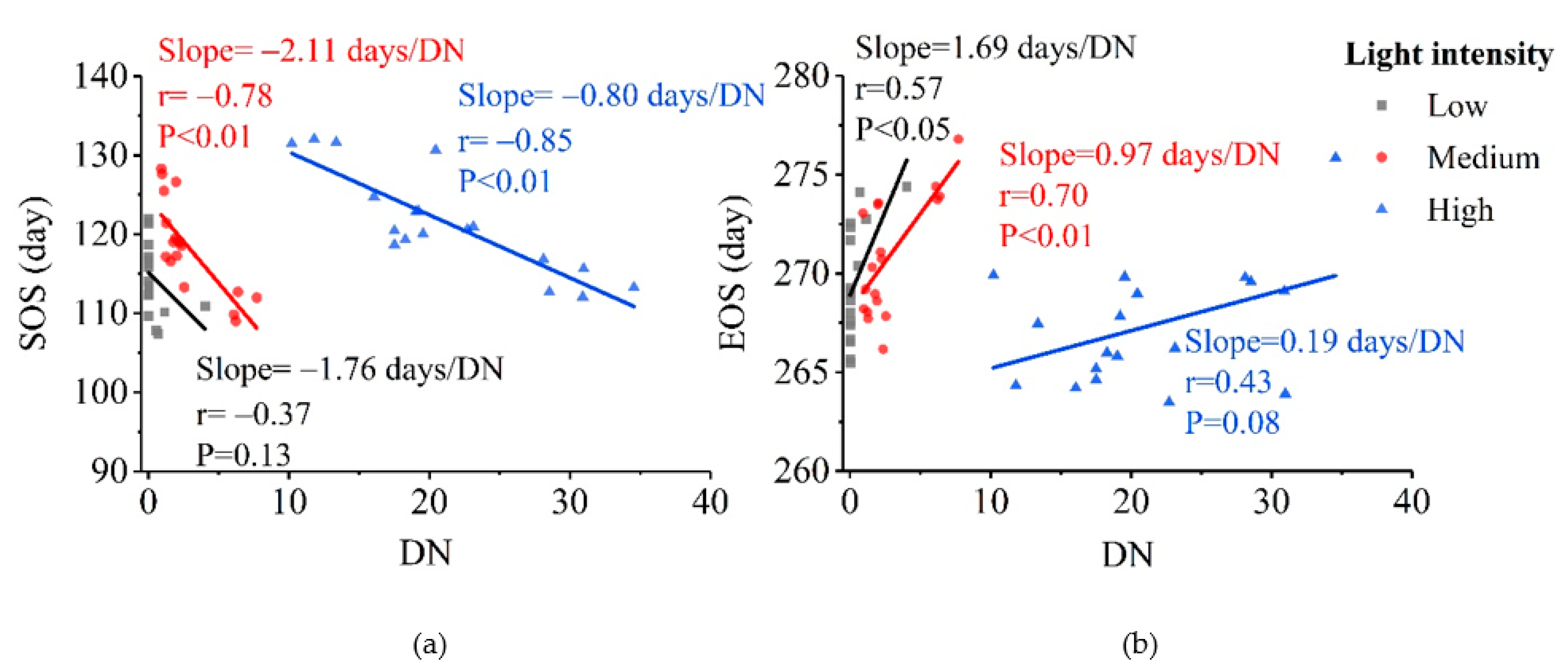
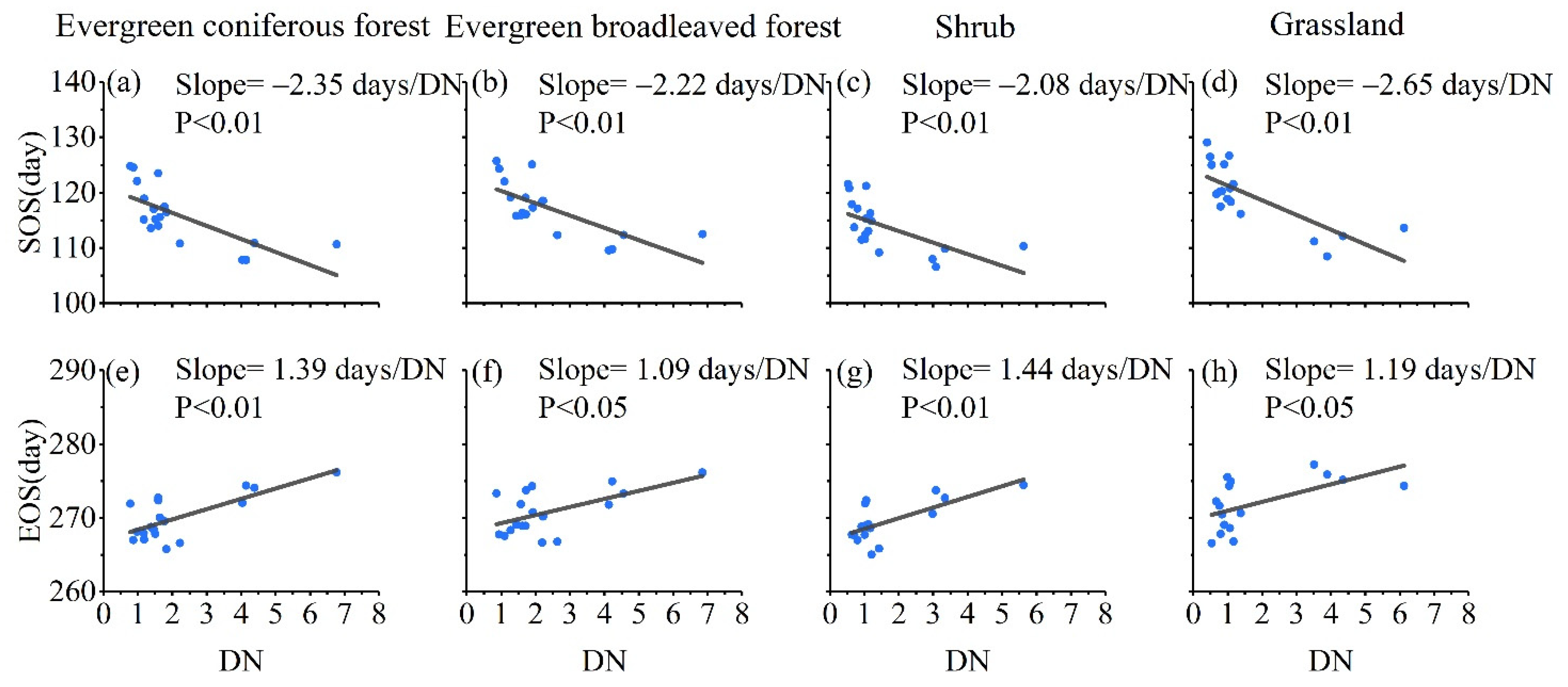
3.2. Patterns of Vegetation Photosynthetic Phenological Changes with Increased Urbanization
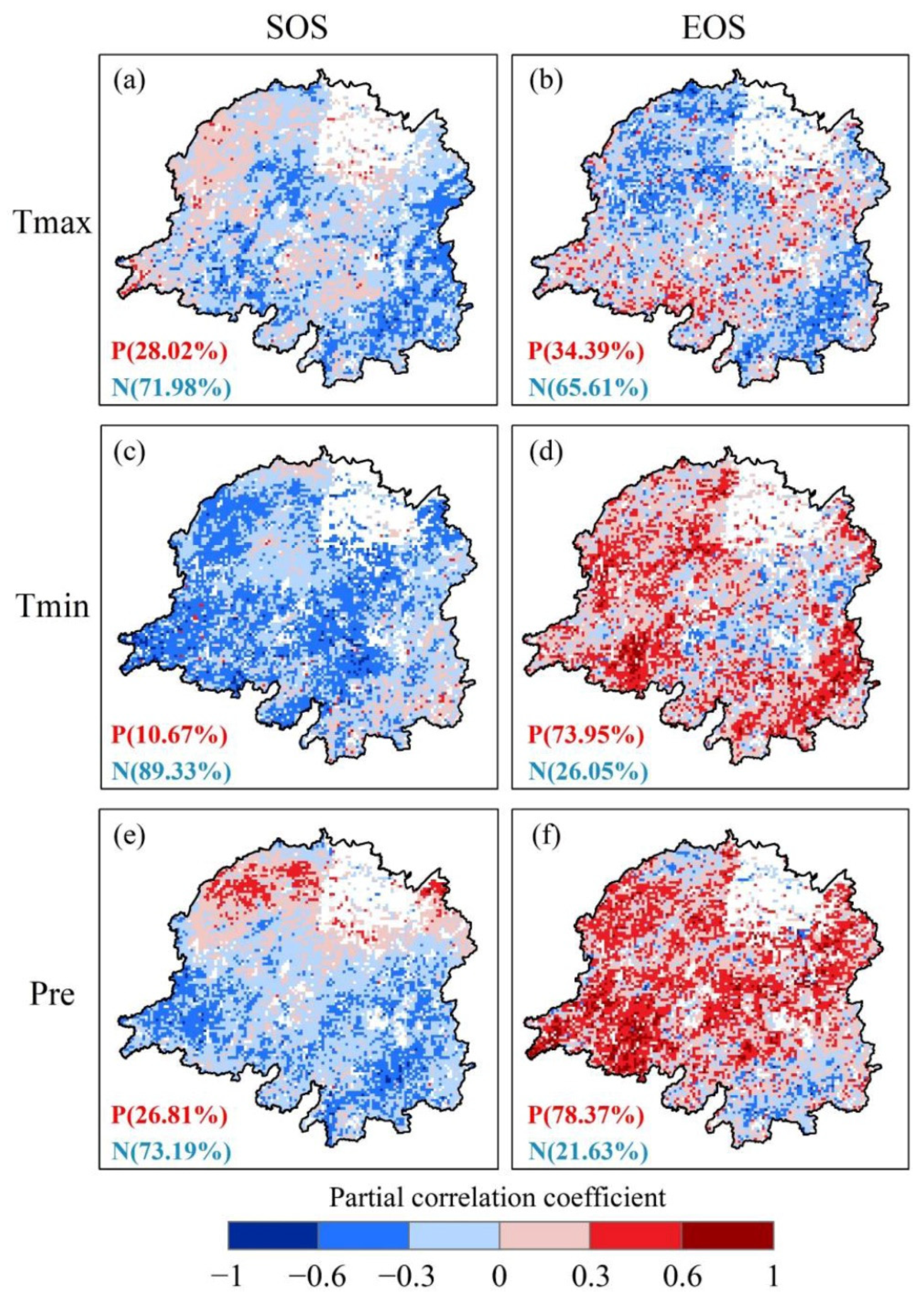
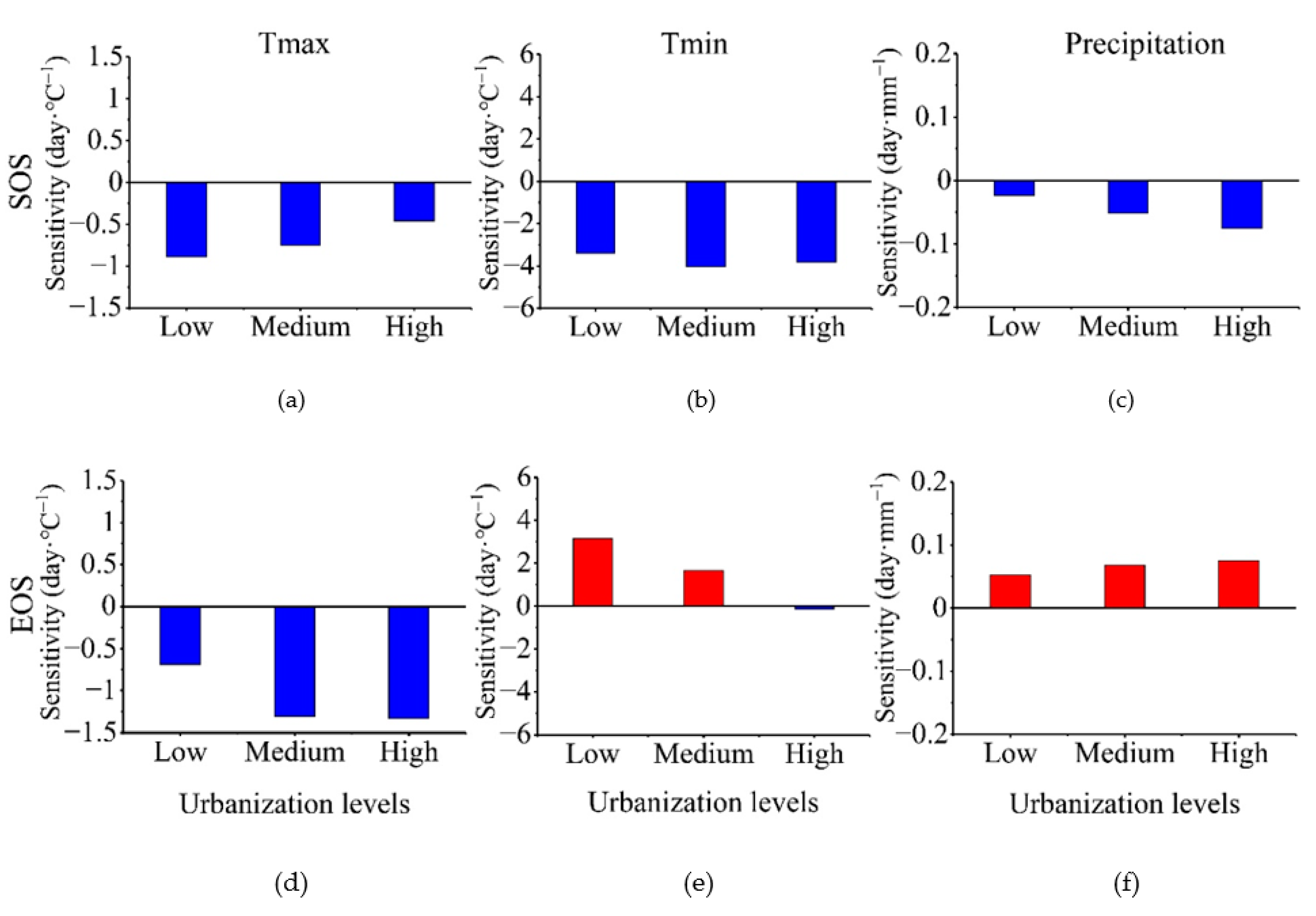
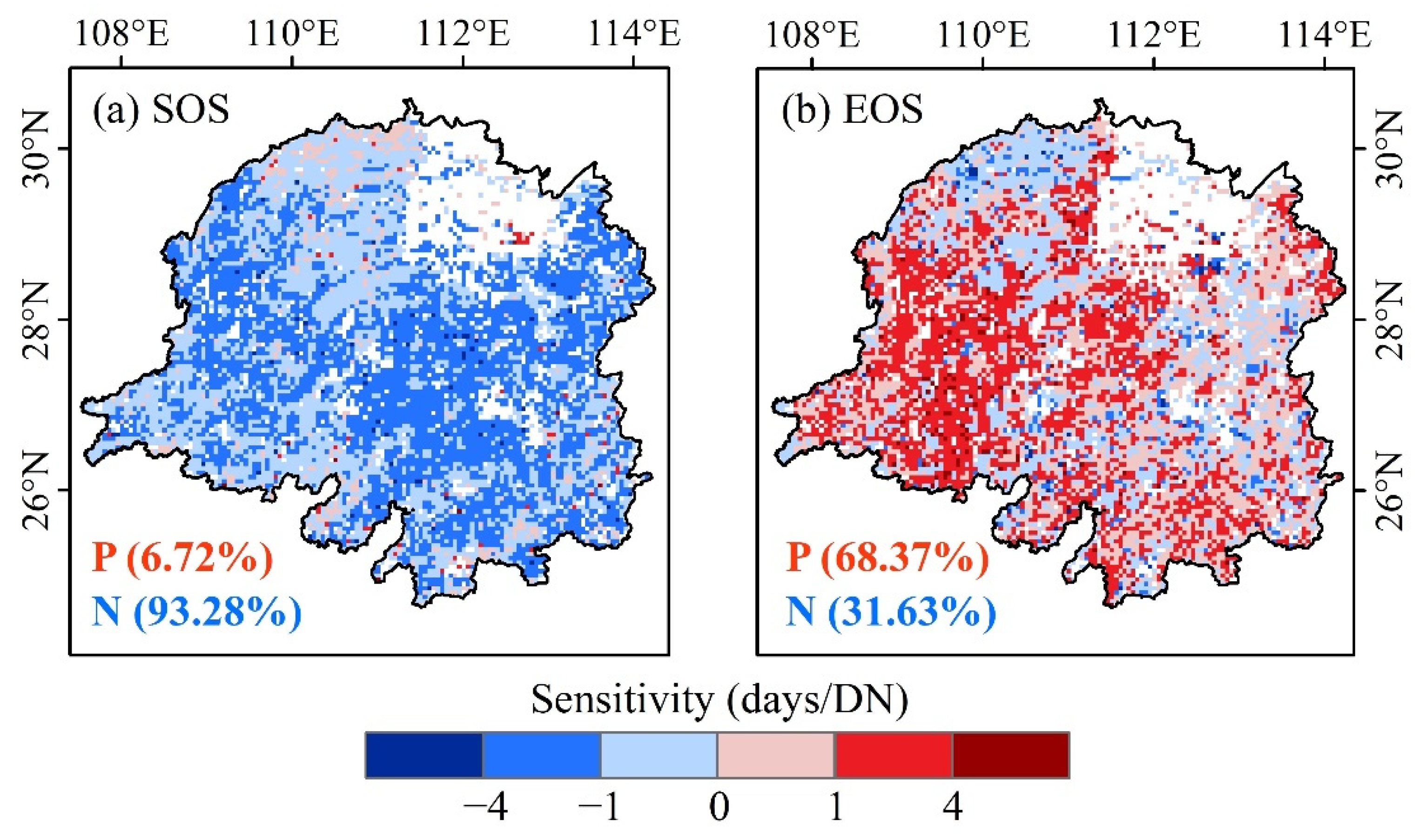
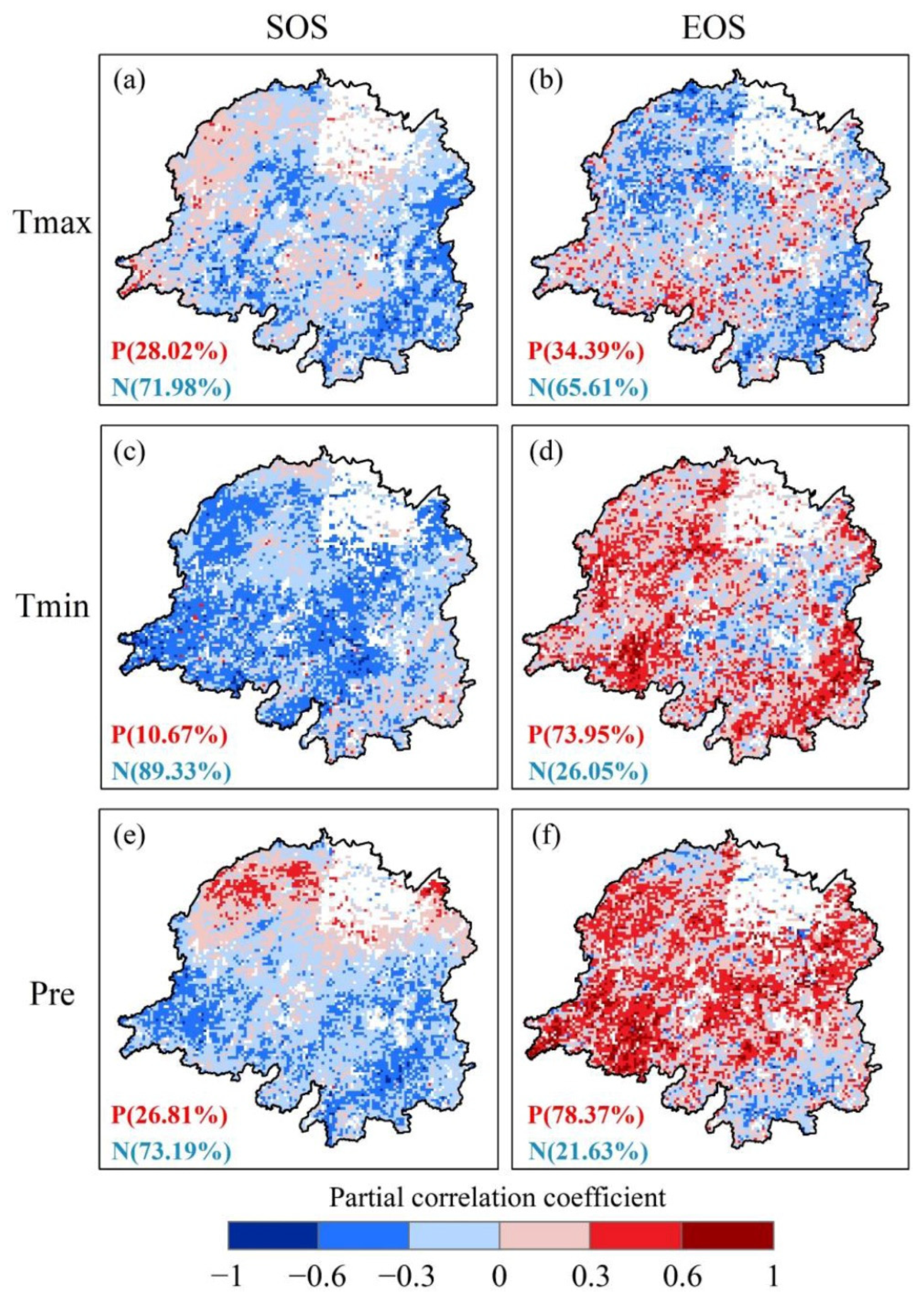
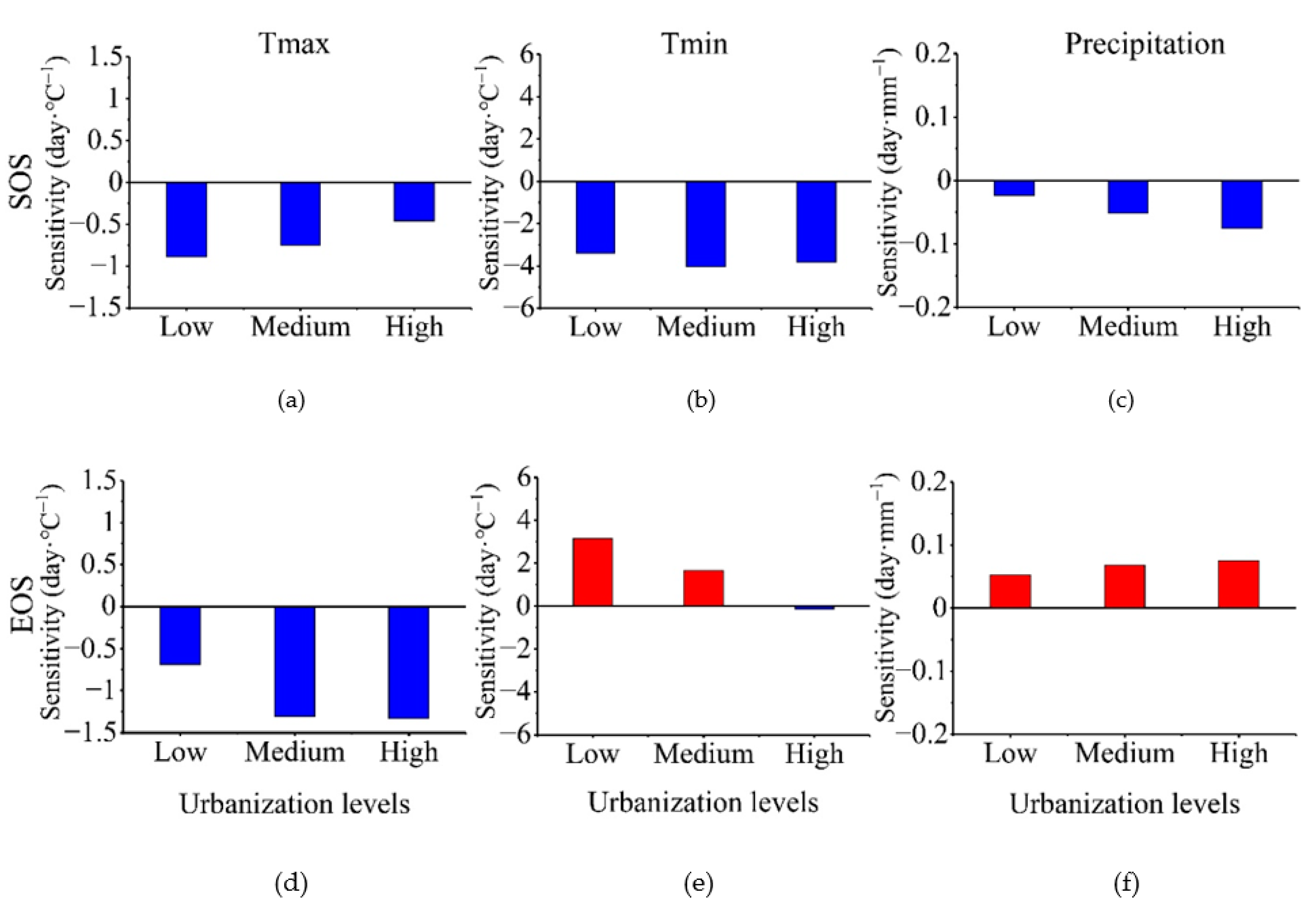
3.3. Climate Sensitivity of Vegetation Phenology According to Urbanization Gradient
4. Discussion
4.1. Changes of Vegetation Photosynthetic Phenology in the Dongting Lake Basin
4.2. Response of Vegetation Phenology Changes to Urbanization
4.3. Effects of Urbanization on the Climatic Sensitivity of Vegetation Photosynthetic Phenology
4.4. Uncertainty and Insights
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, K.Z.; Wan, M.W. Phenology; Hunan Education Press: Changsha, China, 1999. [Google Scholar]
- Ge, Q.S.; Wang, H.J.; Rutishauser, T.; Dai, J.H. Phenological response to climate change in China: A meta-analysis. Glob. Chang. Biol. 2015, 21, 265–274. [Google Scholar] [CrossRef]
- Fu, Y.S.; Zhao, H.F.; Piao, S.L.; Peaucelle, M.; Peng, S.S.; Zhou, G.Y.; Ciais, P.; Huang, M.T.; Menzel, A.; Penuelas, J.; et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.S.; Piao, S.L.; Zhou, X.C.; Geng, X.J.; Hao, F.H.; Vitasse, Y.; Janssens, I.A. Short photoperiod reduces the temperature sensitivity of leaf-out in saplings of Fagus sylvatica but not in horse chestnut. Glob. Chang. Biol. 2019, 25, 1696–1703. [Google Scholar] [CrossRef] [PubMed]
- Linderholm, H.W. Growing season changes in the last century. Agric. For. Meteorol. 2006, 137, 1–14. [Google Scholar] [CrossRef]
- Menzel, A.; Yuan, Y.; Matiu, M.; Sparks, T.; Scheifinger, H.; Gehrig, R.; Estrella, N. Climate change fingerprints in recent European plant phenology. Glob. Chang. Biol. 2020, 26, 2599–2612. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.L.; Liu, Q.; Chen, A.P.; Janssens, I.A.; Fu, Y.S.; Dai, J.H.; Liu, L.L.; Lian, X.; Shen, M.G.; Zhu, X.L. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Liu, X.T.; Zhou, L.; Shi, H.; Wang, S.Q.; Chi, Y.G. Phenological characteristics of temperate coniferous and broad-leaved mixed forests based on multiple remote sensing vegetation indices, chlorophyll fluorescence and CO2 flux data. Acta Ecol. Sin. 2018, 38, 3482–3494. [Google Scholar] [CrossRef]
- Wu, C.Y.; Wang, X.Y.; Wang, H.J.; Ciais, P.; Peñuelas, J.; Myneni, R.B.; Desai, A.R.; Gough, C.M.; Gonsamo, A.; Black, A.T.; et al. Contrasting responses of autumn-leaf senescence to daytime and night-time warming. Nat. Clim. Chang. 2018, 8, 1092–1096. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, Z.L.; Zhou, X.L.; Xie, B.G.; Li, Z.W.; Luo, Y.P.; Zhu, Q.A.; Peng, C.H. Combined control of multiple extreme climate stressors on autumn vegetation phenology on the Tibetan Plateau under past and future climate change. Agric. For. Meteorol. 2021, 308–309. [Google Scholar] [CrossRef]
- Huang, W.L.; Zhang, Q.; Kong, D.D.; Gu, X.H.; Sun, P.; Hu, P. Response of vegetation phenology to drought in Inner Mongolia from 1982 to 2013. Acta Ecol. Sin. 2019, 39, 4953–4965. [Google Scholar] [CrossRef]
- Chen, M.X. Research progress and scientific issues in the field of urbanization. Geogr. Res. 2015, 34, 614–630. [Google Scholar] [CrossRef]
- Buyantuyev, A.; Wu, J. Urbanization diversifies land surface phenology in arid environments: Interactions among vegetation, climatic variation, and land use pattern in the Phoenix metropolitan region, USA. Landsc. Urban Plan. 2012, 105, 149–159. [Google Scholar] [CrossRef]
- Qiu, T.; Song, C.H.; Zhang, Y.L.; Liu, H.S.; Vose, J.M. Urbanization and climate change jointly shift land surface phenology in the northern mid-latitude large cities. Remote Sens. Environ. 2020, 236, 111477. [Google Scholar] [CrossRef]
- Li, D.; Stucky, B.J.; Deck, J.; Baiser, B.; Guralnick, R.P. The effect of urbanization on plant phenology depends on regional temperature. Nat. Ecol. Evol. 2019, 3, 1661–1667. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Tomelleri, E.; Hammerle, A. The urban imprint on plant phenology. Nat. Ecol. Evol. 2019, 3, 1668–1674. [Google Scholar] [CrossRef]
- Meng, L.; Mao, J.F.; Zhou, Y.Y.; Richardson, A.D.; Lee, X.; Thornton, P.E.; Ricciuto, D.M.; Li, X.C.; Dai, Y.J.; Shi, X.Y.; et al. Urban warming advances spring phenology but reduces the response of phenology to temperature in the conterminous United States. Proc. Natl. Acad. Sci. USA 2020, 117, 4228–4233. [Google Scholar] [CrossRef]
- Zhou, D.C.; Zhao, S.Q.; Zhang, L.X.; Liu, S.G. Remotely sensed assessment of urbanization effects on vegetation phenology in China’s 32 major cities. Remote Sens. Environ. 2016, 176, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Bennie, J.; Davies, T.W.; Cruse, D.; Gaston, K.J.; Swenson, N. Ecological effects of artificial light at night on wild plants. J. Ecol. 2016, 104, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Viera-Perez, M.; Hernandez-Calvento, L.; Hesp, P.A.; Santana-Del Pino, A. Effects of artificial light on flowering of foredune vegetation. Ecology 2019, 100, e02678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennie, J.; Davies, T.W.; Cruse, D.; Bell, F.; Gaston, K.J.; James, J. Artificial light at night alters grassland vegetation species composition and phenology. J. Appl. Ecol. 2018, 55, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Neil, K.; Wu, J. Effects of urbanization on plant flowering phenology: A review. Urban Ecosyst. 2006, 9, 243–257. [Google Scholar] [CrossRef]
- Ren, P.X.; Liu, Z.L.; Zhou, X.L.; Peng, C.H.; Xiao, J.F.; Wang, S.H.; Li, X.; Li, P. Strong controls of daily minimum temperature on the autumn photosynthetic phenology of subtropical vegetation in China. For. Ecosyst. 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Jochner, S.; Menzel, A. Urban phenological studies—Past, present, future. Environ. Pollut. 2015, 203, 250–261. [Google Scholar] [CrossRef]
- Karkauskaite, P.; Tagesson, T.; Fensholt, R. Evaluation of the Plant Phenology Index (PPI), NDVI and EVI for start-of-season trend analysis of the northern hemisphere boreal zone. Remote Sens. 2017, 9, 485. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Commane, R.; Zhou, S.; Williams, A.P.; Gentine, P. Light limitation regulates the response of autumn terrestrial carbon uptake to warming. Nat. Clim. Chang. 2020, 10, 739–743. [Google Scholar] [CrossRef]
- Jeong, S.J.; Schimel, D.; Frankenberg, C.; Drewry, D.T.; Fisher, J.B.; Verma, M.; Berry, J.A.; Lee, J.-E.; Joiner, J. Application of satellite solar-induced chlorophyll fluorescence to understanding large-scale variations in vegetation phenology and function over northern high latitude forests. Remote Sens. Environ. 2017, 190, 178–187. [Google Scholar] [CrossRef]
- Wang, S.H.; Ju, W.M.; Penuelas, J.; Cescatti, A.; Zhou, Y.S.; Fu, Y.S.; Huete, A.; Liu, M.; Zhang, Y.G. Urban-rural gradients reveal joint control of elevated CO2 and temperature on extended photosynthetic seasons. Nat. Ecol. Evol. 2019, 3, 1076–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Dannenberg, M.P.; Yan, D.; Jones, M.O.; Kimball, J.S.; Moore, D.J.P.; Leeuwen, W.J.D.; Didan, K.; Smith, W.K. Globally consistent patterns of asynchrony in vegetation phenology derived from optical, microwave, and fluorescence satellite data. J. Geophys. Res. Biogeosciences 2020, 125, e2020JG005732. [Google Scholar] [CrossRef]
- Walther, S.; Voigt, M.; Thum, T.; Gonsamo, A.; Zhang, Y.; Kohler, P.; Jung, M.; Varlagin, A.; Guanter, L. Satellite chlorophyll fluorescence measurements reveal large-scale decoupling of photosynthesis and greenness dynamics in boreal evergreen forests. Glob. Chang. Biol. 2016, 22, 2979–2996. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.M.; Wang, Z.C.; Li, C.H.; Zhao, J.; Li, Y. Assessment and prediction of land ecological environment quality change based on remote sensing-a case study of the Dongting lake area in China. IOP Conf. Ser. Earth Environ. Sci. 2018, 121, 032053. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J.F. Global climatic controls on interannual variability of ecosystem productivity: Similarities and differences inferred from solar-induced chlorophyll fluorescence and enhanced vegetation index. Agric. For. Meteorol. 2020, 288, 108018. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J.F. A global, 0.05-degree product of solar-induced chlorophyll fluorescence derived from OCO-2, MODIS, and reanalysis data. Remote Sens. 2019, 11, 517. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, J.F. Mapping photosynthesis solely from solar-induced chlorophyll fluorescence: A global, fine-resolution dataset of gross primary production derived from OCO-2. Remote Sens. 2019, 11, 2563. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Y.; Smith, S.J.; Zhao, K.; Imhoff, M.; Thomson, A.; Bond-Lamberty, B.; Asrar, G.R.; Zhang, X.S.; He, C.Y.; Elvidge, C.D. A global map of urban extent from nightlights. Environ. Res. Lett. 2015, 10. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, Y.Y.; Li, X.C.; Zhou, C.H.; Cheng, W.M.; Li, M.C.; Huang, K. Building a series of consistent night-time light data (1992–2018) in Southeast Asia by integrating DMSP-OLS and NPP-VIIRS. IEEE Trans. Geosci. Remote. Sens. 2020, 58, 1843–1856. [Google Scholar] [CrossRef]
- Xu, W.T.; Wu, B.F.; Yan, C.Z.; Huang, H.P. China land cover 2000 using SPOT VGT S10 data. J. Remote. Sens. 2005, 9, 204–214. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Zhou, L.; Chi, Y.G.; Liu, X.T.; Dai, X.Q.; Yang, F.T. Land surface phenology tracked by remotely sensed sun-induced chlorophyll fluorescence in subtropical evergreen coniferous forests. Acta Ecol. Sin. 2020, 40, 4114–4125. [Google Scholar] [CrossRef]
- Li, P.; Peng, C.H.; Zhang, J.; Wang, M.; Zhang, J.J.; Ding, J.H.; Zhou, X.L.; Zhu, Q. Change in autumn vegetation phenology and the climate controls from 1982 to 2012 on the Qinghai-Tibet Plateau. Front. Plant Sci. 2019, 10, 1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.X.; Fu, Y.S.; Chen, S.Z.; Xiao, J.F.; Yin, G.D.; Li, X.; Zhang, X.S.; Geng, X.J.; Wu, Z.F.; Zhou, X.C.; et al. Increasing importance of precipitation in spring phenology with decreasing latitudes in subtropical forest area in China. Agric. For. Meteorol. 2021, 304, 108427. [Google Scholar] [CrossRef]
- Wu, J.S.; Liu, H.; Peng, J.; Ma, L. Hierarchical structure and spatial pattern of China’s urban system: Evidence from DMSP/OLS nightlight data. Acta Geogr. Sin. 2014, 69, 759–770. [Google Scholar] [CrossRef]
- Wang, X.H.; Piao, S.L.; Xu, X.T.; Ciais, P.; MacBean, N.; Myneni, R.B.; Li, L. Has the advancing onset of spring vegetation green-up slowed down or changed abruptly over the last three decades? Glob. Ecol. Biogeogr. 2015, 24, 621–631. [Google Scholar] [CrossRef]
- Shen, X.J.; Liu, B.H.; Henderson, M.; Wang, L.; Wu, Z.F.; Wu, H.T.; Jiang, M.; Lu, X.G. Asymmetric effects of daytime and nighttime warming on spring phenology in the temperate grasslands of China. Agric. For. Meteorol. 2018, 259, 240–249. [Google Scholar] [CrossRef]
- Zhu, W.Q.; Tian, H.Q.; Xu, X.F.; Pan, Y.Z.; Chen, G.S.; Lin, W.P. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- Liu, Q.; Fu, Y.S.; Zeng, Z.Z.; Huang, M.T.; Li, X.R.; Piao, S.L. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob. Chang. Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [Green Version]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Piao, S.L.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.Y.; Barr, A.; Chen, A.P.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef]
- Li, X.C.; Zhou, Y.Y.; Asrar, G.R.; Mao, J.F.; Li, X.M.; Li, W.Y. Response of vegetation phenology to urbanization in the conterminous United States. Glob. Chang. Biol. 2017, 23, 2818–2830. [Google Scholar] [CrossRef]
- Tai, X.L.; Epstein, H.E.; Li, B. Elevation and climate effects on vegetation greenness in an arid mountain-basin system of Central Asia. Remote Sens. 2020, 12, 1665. [Google Scholar] [CrossRef]
- Jonsson, S.; Segall, P.; Pedersen, R.; Bjornsson, G. Post-earthquake ground movements correlated to pore-pressure transients. Nature 2003, 424, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Zohner, C.M. Phenology and the city. Nat. Ecol. Evol. 2019, 3, 1618–1619. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, S.; Jiang, B.; Ren, Y.; Wang, H.; Vause, J.; Yu, H. Temporal trend of green space coverage in China and its relationship with urbanization over the last two decades. Sci. Total Environ. 2013, 442, 455–465. [Google Scholar] [CrossRef]
- Oliveira, S.; Andrade, H.; Vaz, T. The cooling effect of green spaces as a contribution to the mitigation of urban heat: A case study in Lisbon. Build. Environ. 2011, 46, 2186–2194. [Google Scholar] [CrossRef]
- Huang, M.; Cui, P.; He, X. Study of the cooling effects of urban green space in Harbin in terms of reducing the heat island effect. Sustainability 2018, 10, 1101. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.L.; Dai, H.; Hou, F.; LI, E.Z. Spatio-temporal change of urban-rural vegetation phenology and its response to land surface temperature in Northeast China. Acta Ecol. Sin. 2020, 40, 4137–4145. [Google Scholar] [CrossRef]
- Jia, W.X.; Zhao, S.Q.; Zhang, X.Y.; Liu, S.G.; Henebry, G.; Liu, L.L. Urbanization imprint on land surface phenology: The urban–rural gradient analysis for Chinese cities. Glob. Chang. Biol. 2021, 27, 2895–2904. [Google Scholar] [CrossRef] [PubMed]
- Han, G.F.; Xu, J.H. Land surface phenology and land surface temperature changes along an urban-rural gradient in Yangtze River Delta, China. Environ. Manag. 2013, 52, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Decina, S.M.; Templer, P.H.; Hutyra, L.R. Atmospheric inputs of nitrogen, carbon, and phosphorus across an Urban Area: Unaccounted fluxes and canopy influences. Earths Future 2018, 6, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Do, F.C.; Goudiaby, V.A.; Gimenez, O.; Diagne, A.L.; Diouf, M.; Rocheteau, A.; Akpo, L.E. Environmental influence on canopy phenology in the dry tropics. For. Ecol. Manag. 2005, 215, 319–328. [Google Scholar] [CrossRef]
- Lian, X.H.; Jiao, L.M.; Zhong, J.; Jia, Q.Q.; Liu, J.F.; Liu, Z.J. Artificial light pollution inhibits plant phenology advance induced by climate warming. Environ. Pollut. 2021, 291, 118110. [Google Scholar] [CrossRef]
- Peng, J.D.; Zhao, H.; Chen, X.C. Effect of urbanization process on local climate in Changsha City. Meteorol. Environ. Sci. 2017, 40, 42–48. [Google Scholar] [CrossRef]
- Peng, L.L.; Luo, B.L.; Sun, J.Q. Variation of extreme precipitation during the urbanization of Changsha-Zhuzhou-Xiangtan urban agglomeration. Torrential Rain Disasters 2015, 34, 191–196. [Google Scholar] [CrossRef]
- Xiao, F.; Zhu, B.; Zhu, T. Inconsistent urbanization effects on summer precipitation over the typical climate regions in central and eastern China. Theor. Appl. Climatol. 2020, 143, 73–85. [Google Scholar] [CrossRef]
- Xin, Q.C.; Li, J.; Li, Z.M.; Li, Y.M.; Zhou, X.W. Evaluations and comparisons of rule-based and machine-learning-based methods to retrieve satellite-based vegetation phenology using MODIS and USA National Phenology Network data. Int. J. Appl. Earth Obs. Geoinf. 2020, 93. [Google Scholar] [CrossRef]
- Izar, A.; Noelia, O.; Marco, Q.; Igor, G.O.; Diego, G.; Susanna, M. Comparison of climate reanalysis and remote-sensing data for predicting olive phenology through machine-learning methods. Remote Sens. 2021, 13, 1224. [Google Scholar] [CrossRef]
- Guo, Y.H.; Fu, Y.S.; Hao, F.H.; Zhang, X.; Wu, W.X.; Jin, X.L.; Robin Bryant, C.; Senthilnath, J. Integrated phenology and climate in rice yields prediction using machine learning methods. Ecol. Indic. 2021, 120, 106935. [Google Scholar] [CrossRef]
- Guo, Y.H.; Yin, G.D.; Sun, H.Y.; Wang, H.X.; Chen, S.Z.; Senthilnath, J.; Wang, J.Z.; Fu, Y.S. Scaling effects on chlorophyll content estimations with RGB camera mounted on a UAV platform using machine-learning methods. Sensors 2020, 20, 5130. [Google Scholar] [CrossRef]
- Pope, K.S.; Dose, V.; Da Silva, D.; Brown, P.H.; Leslie, C.A.; Dejong, T.M. Detecting nonlinear response of spring phenology to climate change by Bayesian analysis. Glob. Chang. Biol. 2013, 19, 1518–1525. [Google Scholar] [CrossRef]
- Shen, M.G.; Piao, S.L.; Cong, N.; Zhang, G.X.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Level | Maximum Temperature (°C/DN) | Minimum Temperature (°C/DN) | Precipitation | |||
|---|---|---|---|---|---|---|
| SOS | EOS | SOS | EOS | SOS | EOS | |
| Low | 0.15 | 0.15 | 0.56 * | 0.77 | 0.02 | 0.02 * |
| Medium | 0.33 | 0.29 | 1.51 ** | 2.18 | 0.03 | 0.04 ** |
| High | 0.98 | 1.81 | 4.74 ** | 8.94 * | 0.07 | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Sun, M.; Liu, Y.; Ren, P.; Peng, C.; Zhou, X.; Tang, J. Response of Vegetation Photosynthetic Phenology to Urbanization in Dongting Lake Basin, China. Remote Sens. 2021, 13, 3722. https://doi.org/10.3390/rs13183722
Li P, Sun M, Liu Y, Ren P, Peng C, Zhou X, Tang J. Response of Vegetation Photosynthetic Phenology to Urbanization in Dongting Lake Basin, China. Remote Sensing. 2021; 13(18):3722. https://doi.org/10.3390/rs13183722
Chicago/Turabian StyleLi, Peng, Mai Sun, Yuxin Liu, Peixin Ren, Changhui Peng, Xiaolu Zhou, and Jiayi Tang. 2021. "Response of Vegetation Photosynthetic Phenology to Urbanization in Dongting Lake Basin, China" Remote Sensing 13, no. 18: 3722. https://doi.org/10.3390/rs13183722
APA StyleLi, P., Sun, M., Liu, Y., Ren, P., Peng, C., Zhou, X., & Tang, J. (2021). Response of Vegetation Photosynthetic Phenology to Urbanization in Dongting Lake Basin, China. Remote Sensing, 13(18), 3722. https://doi.org/10.3390/rs13183722