We show NDVI in time-series because this index is a standard product, yet we provide change maps using NDVI* as scene-to-scene comparisons of change require the scaled version (hence NDVI* and not NDVI for change maps). We assessed NDVI and NDVI*, but our research results are primarily confined to data derived from EVI2: EVI2 for the peak growing season, EVI2 for the full year, ET with EVI2 as the input VI, and PAM ET using EVI2. We first report the absolute values, then the difference and percent change data from these seven periods: 2000–2020, 2000–2010, 2011–2020, 2000–2005, 2005–2010, 2011–2015, and 2016–2020.
3.1. Vegetation Index Time-Series from 2000 to 2020, EVI2
We produced time-series data for three VIs including NDVI, EVI, and EVI2, but because we found that EVI and EVI2 had an r
2 = 0.99, we proceeded with using only NDVI and EVI2 (
Figure 3). NDVI (
Figure 3a) and EVI2 (
Figure 3b) were produced following our methods which account for amplitude shifts across three sensors, Landsat5 (TM5), Landsat7 (ETM+), and Landsat8 (OLI).
Time series trend data, both NDVI and EVI2, since 2000are shown for each reach (R3 to R7) and the weighted average for the sum of the reaches (All) (
Figure 3). This beginning date was selected because, in our previous study, we used both Landsat and MODIS to compare how closely they provided results, and we determined that they are essentially measuring the same information [
14]. For that reason, we chose not to show MODIS data in this research, yet we begin the acquisition of Landsat time-series with the date that corresponds with the start of the MODIS collection. NDVI (
Figure 3a) and EVI2 (
Figure 3b) have some similarities and differences. The differences are that NDVI has a higher amplitude and nearly all of the results range between 0.2 and 0.4, whereas EVI2 ranges between 0.10 and 0.25, much lower. Furthermore, NDVI shows a deterioration of the signal beginning in 2015 through 2020, whereas EVI2 depicts these seasonal trends as having kept their shapes during this same period. NDVI does not capture the full spectrum the way EVI2 does; NDVI saturates at the high end and increases the signal amplitude, but EVI2 performs throughout the full red to near-infrared dynamic range [
122,
125,
126]. The similarities are that generally Reach 6 (R6) has the highest VI, followed by Reach 7 (R7), Reach 3 (R3), and Reach 4 (R4), while Reach 5 (R5) has the lowest VI. In both trends, we see similar decreases in VI, such as a drop of 0.34 to 0.275 for NDVI and 0.22 to 0.175 for EVI2 between 2000 and 2020.
Because we use EVI2 and not NDVI as input to the ET algorithm, we focus our discussion on EVI2. The EVI2 is used to compute the peak growing season average for each reach (R3 to R7) and the weighted average (All) for the 21 years in the study. These EVI2 data are in the Supplemental Information,
Appendix A,
Table A1 (
Appendix A,
Table A1). In
Appendix A,
Table A2, the EVI2 difference between years for the peak growing season, averaged between May 1 and October 30, is provided for each reach (R3 to R7) and the weighted average as All Reaches (
Appendix A,
Table A2). We provide the percent change of EVI2 between individual years for the five reaches and the full All reach (
Appendix A,
Table A3). Last, we also provide the percent change of EVI2 between groups of years for the peak growing season, averaged between May 1 and Oct. 30. We provide the percent change between 2000–2020, 2000–2010, 2011–2020, 2000–2005, 2006–2010, 2011–2015, and 2016–2020 in
Appendix A,
Table A4. This shows EVI2 for the peak growing season averaged per reach and All for each of the seven periods using to compare changes (
Appendix A,
Table A4).
EVI2, when averaged only over the growing season between May 1 and October 30 each year, decreases from 0.1993 to 0.1650 over the 21 years (
Appendix A,
Table A1). The decrease in EVI2 for R5 is smallest, 0.1689 to 0.1494, and R6 has the largest increase, from 0.2484 to 0.1604, over the same time. The EVI2 difference between each 1-year period is 12 decreasing and eight increasing years. We see that the largest increase in EVI2, a jump of 0.0181, occurred between 2004 and 2005, and the largest decline, a drop of 0.0196, occurred in the period 2010 to 2011 (
Appendix A,
Table A2). Knowing in which year the most or least greening occurred is a major contribution to our research findings. Another new contribution is the calculated percent change between years (
Appendix A,
Table A3). We broke the average peak growing season EVI2 into two ten-year periods, 2000–2010 and 2011–2020, and four five-year periods, 2000–2005 (six years), 2006–2010 (five years), 2011–2015 (five years), and 2016–2020 (five years), for further evaluation of the increases and decreases over these periods so we could state which period depicted the most greening and the most loss in vegetation health (
Appendix A,
Table A4). Similarly, we can determine the differences and percent changes for these periods within the five different river reaches. The data show the five-year data had a peak greenness, 0.1995, occurring during 2000–2005, and the maximum loss of green vegetation, 0.1719, happened between 2016 and 2020. A decline of −0.01 in EVI2 was observed between the first and last ten-year periods. R6 showed the maximum greenness, 0.2466, in the 2000–2005 period and the least green vegetation, 0.1599, in R5 during 2016–2020.
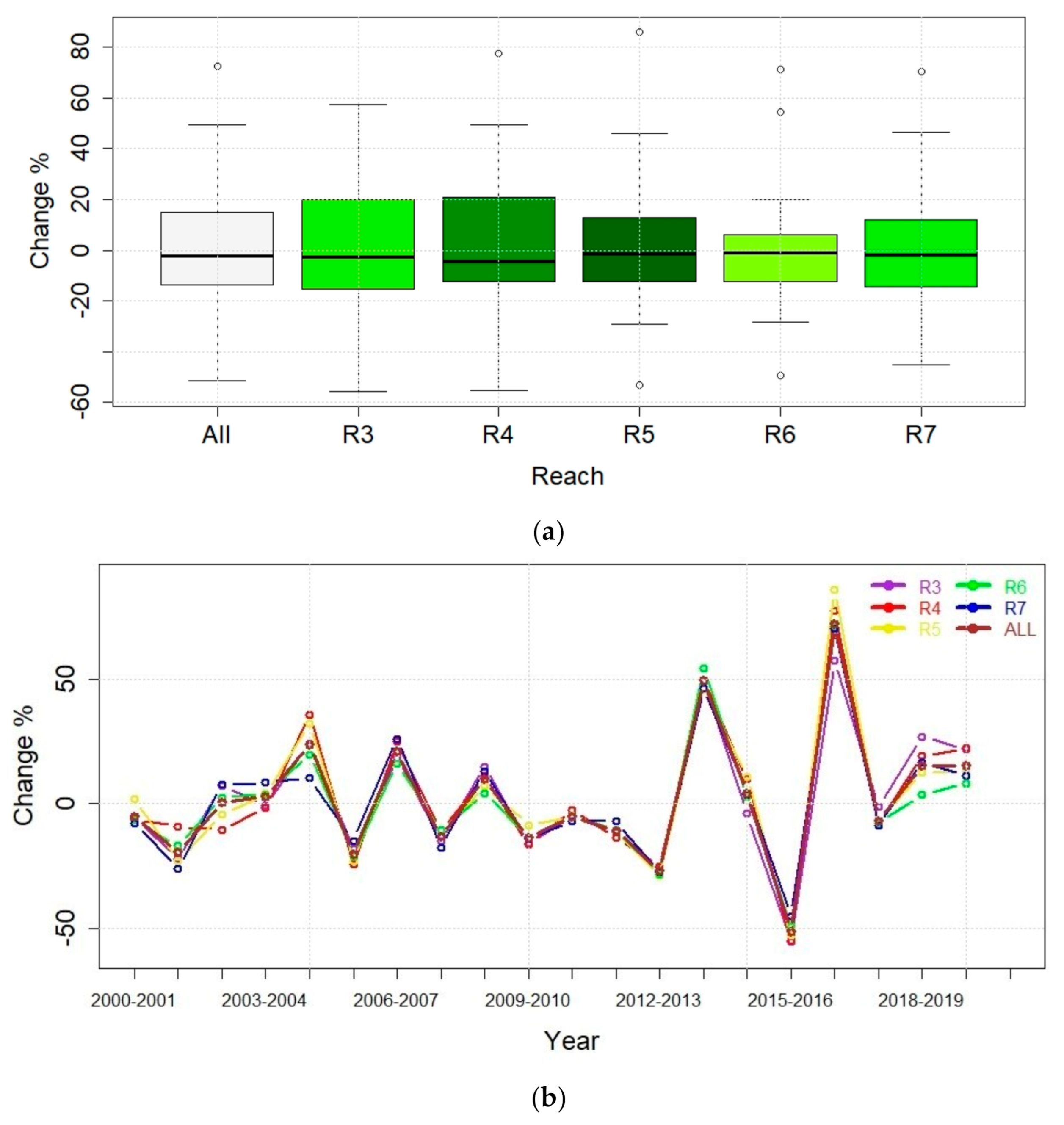
To visualize these changes,
Figure 4 shows five important statistics which include the minimum, first quantile (Q1, 25th percentile), median (Q2, 50th Percentile), third quantile (Q3, 75th percentile), and maximum, values that allow for a comparison of the reaches according to the EVI2 percent change (4a, top) and demonstrate the rate of change from year-to-year in all reaches (4b, bottom). In the top diagram, the darker the box, the larger the range of change. As an example, R4 shows the largest range of change in EVI2 (−8.33% to 17.49%), followed by R5 (−9.09% to 16.35%).
Because it can be difficult to ascertain the importance of these absolute differences in terms of their ups and downs over the growing seasons of these two decades, we provide the percent change of the EVI2 growing season VI data for these same time steps, year-to-year over 20 years, the two ten-year periods, and the four five-year periods.
Figure 4
shows the direction (positive or negative (-)) of the percent change in box plots EVI2 for the weighted average of five reaches (All) and the individual reaches for each year-to-year period (
Figure 4a) and the rate of change (
Figure 4b). In
Appendix A,
Table A3 the values corresponding to
Figure 4
are provided. Using these, the period between 2004 and 2005 shows large greening took place across all reaches, and the highest percent increases of all 20 years took place in this one one-year period where greening increased by 10.48% (R3), 17.49% (R4), 16.35% (R5), 6.00% (R6), and 2.29% (R7), an average of 9.57% increase (All).
This year 2004–2005 was also the period of most greening in the delta [
14] due to nearly three years of rainfall after nearly zero precipitation in 2002 as determined by data from the AZMET Yuma station. In 2009, precipitation was only 20 mmyr
−1, but was followed by the highest rainfall, 150 mmyr
−1, of the past 21 years we studied [
14]. The period between 2010 and 2011 shows large losses in greenness for the entire year and across reaches, where greening decreased by 9.65% (R3), 6.85% (R4), 9.09% (R5), 8.88% (R6), and 10.18% (R7), an average 9.38% decrease (All). This year was also the period of greatest decline in vegetation health as measured by VI for the delta [
14]. The preceding year, 2009, had very low rainfall followed by peak rainfall, then low rainfall again in 2011 (~45 mmyr
−1), so it is not surprising that vegetation declined not only over 2010–2011, but also in the following year, 2011–2012. The period 2011–2012 showed additional losses in vegetation greenness, i.e., R4 decreased −6.85% followed by −8.33%, compounding the declining trend. For R6, the period from 2018–2019 showed decreases of −14.43%, followed by a decrease of −9.18% the following year, 2019–2020. This was the most drastic percent change in the negative direction and is likely due to biocontrol of the vast salt cedar cover in the reach.
After the year-to-year comparisons were documented, we grouped the years into periods to do a comparison of percent change in EVI2 for the average peak growing season relative to other periods, for all reaches, and the full area (All) (
Table 1). In this last analysis step, we provide the percent change of the EVI2 between groups of years and compare them with one another as shown in the first columns, Periods 1 and 2 (
Table 1). Once again, the data are provided for each reach (R3 to R7) and the weighted average as All Reaches. The groups include the ten-year periods from 2000 to 2010 and 2011 to 2020 and four shorter periods of nearly five years: 2000–2005 (six years), 2006–2010, 2011–2015, and 2016–2020. We compare the changes in these time periods and determine percent change in EVI2 for each comparison made. A percent change in small amounts exist for R3 (2.06%), R4 (4.87%), and R7 (0.34%) when comparing 2000–2005 to 2006–2010; however, the greatest percent change was a −13.83% decrease in greenness which occurred for the five-year periods between 2000–2005 and 2016–2020. Over the 10-year comparisons, a decrease in green vegetation is as much as −10.46% between the two decades of 2000–2010 and 2011–2020.
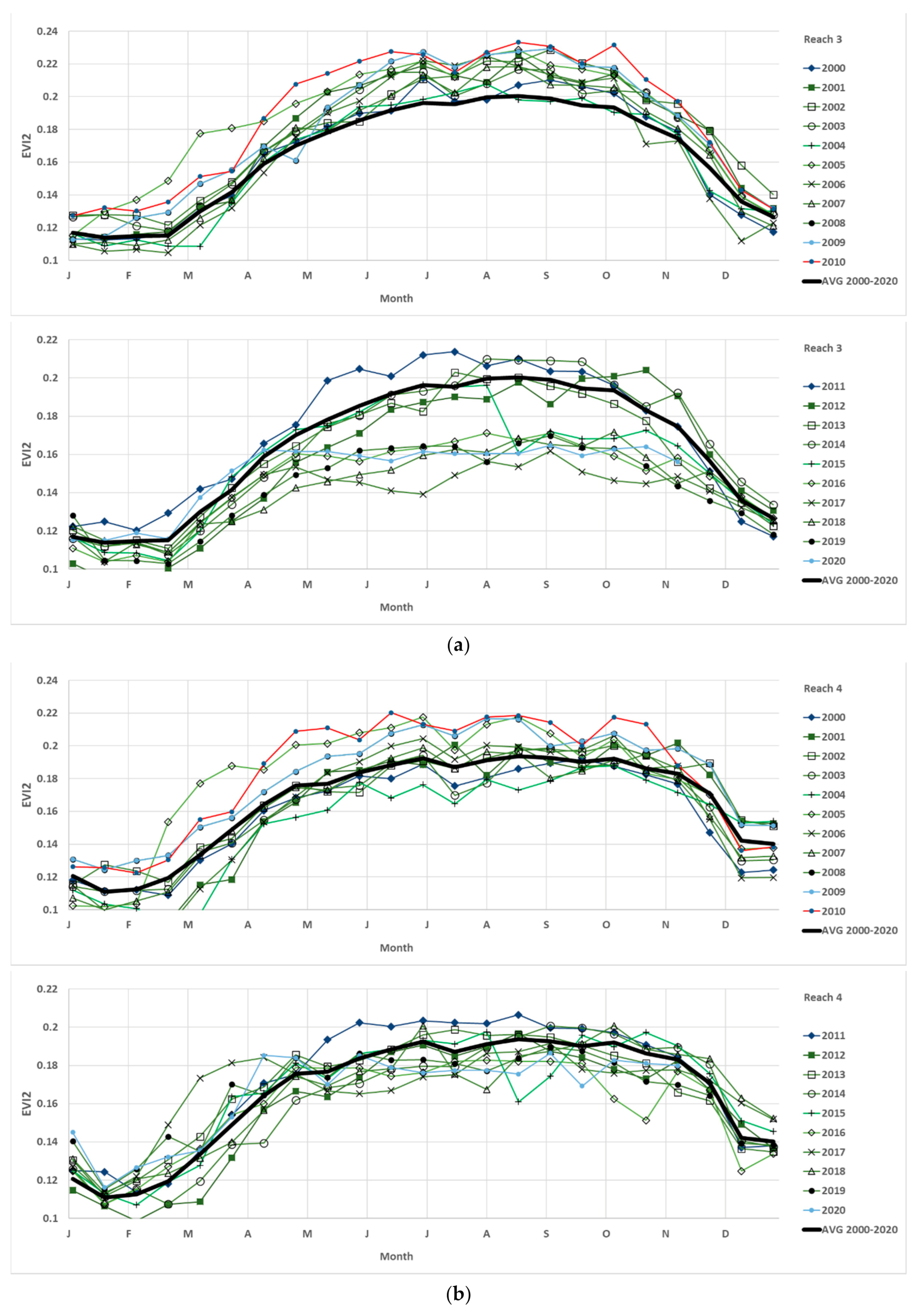
The EVI2 is plotted for the full year, January 1 to December 31, for each of the 21 years, with 2000–2010 (top) and 2011–2020 (bottom) to demonstrate spatiotemporal changes across the full LCR riparian zone, All reaches (
Figure 5). The same data are averaged across the full period, 2000–2020 (black solid line), shown for comparison. The same plots for 2000–2010 (top) and 2010–2020 (bottom) follow for the five individual reaches, R3 to R7 and are shown in
Appendix A (
Figure A1a–e). The data can be interpreted as the closer together the lines (each year), the less change over time and the more intact the phenology curve. Any change in the phenology curve can be due to either a shift in green-up and/or senescence or a disturbance to the growing cycle. Lines representing years below the average (bold black line) show stressed, defoliated, or dead/dying plant communities.
Seeing the anomalies overlaid with one another and in comparison with the 21 year mean (black line,
Figure 5), the EVI2 curves with each annual dataset plotted as lines for all 21 years, is a good way to determine in which months disturbances to the riparian plant system may have occurred. The important information conveyed in the plots show that the first decade has relatively normal riparian greenness and health and water use in All reaches (and for each reach) relative to the average for the study. The second decade, aside from the year 2011, shows the opposite, where all the curves fall under the mean for the study.
EVI2 is an indicator of riparian corridor vegetation health. We demonstrate the annual trends in green vegetation by showing the weighted average EVI2 for All reaches over the last ten years by plotting the individual years 2011–2020, as well as the average 2000–2010 period (red dashes), the average 2011–2020 period (orange short dashes), and the full period 2000–2020 (black solid line) for comparison (
Figure 5). Compared with the first decade (red dashes), all the individual years are less than this, indicating losses in green vegetation year after year. Furthermore, all but 2011 are less EVI2 than the average 21-year full period (black solid line). The collective loss of healthy riparian vegetation over the many months between April and December is indicated by the area under the curves. For example, decreases in magnitude of the curve in summer may be indicative of defoliation caused by the salt cedar biocontrol beetle, and the impact of not having any green canopy affects the overall annual riparian water use. It is the full-year measurement of the area under the ET curve, importantly derived in part from the EVI2 curve, that we use to determine the full-year amount of water use, which is determined by the phenology assessment metric.
The last VI figure shows average EVI2 for seven time-periods using time-series information for the full year for the average weighted total riparian area, All reaches (
Figure 6). Individual reaches are shown in
Appendix A,
Figure A1a–e. The periods shown are the average 2000–2020 period (black solid line), the average EVI2 over the first decade 2000–2010 (red solid line), the average EVI2 over the last decade 2011–2020 (red dotted line), plus four additional approximately five-year periods: 2000–2005 (blue line), 2006–2010 (green line), 2010–2015 (red dashed line), and 2016–2020 (green dashed line). The first decade has the highest EVI2, indicating it was also the greenest in terms of riparian vegetation health. The last decade, 2011–2020 has lower EVI2 with the most recent last five years having the very lowest EVI2, indicating tremendous loss in terms of the ecosystem greenness over the last five-year and ten-year periods. Despite recent on-the-ground research in the upper basin rivers, which found salt cedar-invaded systems to be stable after an 8-year study ending in 2018 [
118], these losses in the LCR are likely not only in part due to biocontrol, which undergoes periods of defoliation that may reduce EVI2, but also is more likely the result of a decrease in greenness in other species, due to recent very hot maximum temperatures and lack of rainfall. Our data support the stability of these ecosystems up to the recent few years, where we observe large declines across the riparian community. The average data over 21 years are nearly the same as the five-year period between 2010 and 2015.
The EVI2 is plotted for the full year, January 1 to December 31, for seven groups of years between 2000 and 2020 to demonstrate spatiotemporal changes in All reaches (
Figure 6). Shown for comparison, the data are averaged across the first decade, 2000–2010 (red solid line), and the second decade, 2011–2020 (red dashed line), as well as the average for the full period, 2000–2020 (black solid line), plus four additional approximately five-year periods: 2000–2005 (blue line), 2006–2010 (green line), 2010–2015 (red dashed line), and 2016–2020 (green dashed line).
These same plots for the five reaches, R3 to R7, are provided in
Appendix A (
Figure A2a–e). In
Appendix A,
Figure A2a for R3, the last five-year and ten-year periods are much lower than the full 21-year period and other period averages. The average data over 21 years is practically the same as the five-year period between 2010 and 2015. In
Appendix A,
Figure A2b, R4 and
Figure A2c, R5, the wide range of EVI2 values by groups of years is not present; the data are very tight indicating similar EVI2 data and time-series trends across all months and across all groups of years. In
Appendix A,
Figure A2d, R6, the first decade has very high EVI2, depicting much green foliage in this part of the riparian corridor. As with other reaches, the last decade indicates a significant loss of green foliage in R6. The last two five-year periods also show these losses, with the last five years, 2016–2020, showing drastic losses in riparian health. In
Appendix A,
Figure A2e, R7, the data are very tight, indicating similar EVI2 data and time-series trends across all months and across all groups of years, but the last decade and the last two five-year periods are lower EVI2 than the averages for the first decade and the full 21-year period.
3.2. ET Daily from VI-Based Time-Series from 2000 to 2020
ET (mmd
−1) was calculated following Equation 4, which used EVI2 and potential ET (ETo) from three nearby weather stations. We show the riparian ET for the full year in five reaches, R3 to R7, as time-series data using Landsat imagery collected ~16 days from 2000 to 2020 for the riparian zone (
Figure 7).
We provide values that correspond with
Figure 7 for ET (mmd
−1) in the peak growing season from 2000–2020 for each reach and the total weighted average in All reaches (
Appendix A,
Table A5). In
Appendix A,
Table A5, the absolute value of ET over all reaches (All) dropped over 1 mmd
−1 from 4.34 mmd
−1 in 2000 to 3.25 mmd
−1 in 2020.
Appendix A,
Table A6 shows the difference in daily ET values, demonstrated as positive or negative (−) change, as measured from one year to the next. The greatest differences across all reaches were observed between 2015 and 2016, possibly due to the initial wave of biocontrol defoliation, where negative change persisted with average daily ET ranging between a decrease of 1.44 mmd
−1 and 1.86 mmd
−1, and All reaches was defined by a decrease in average daily ET of 1.62 mmd
−1.
The percent change from year-to-year using the average daily ET (mmd
−1) for the peak growing season (May 1 to October 30) for each of the 21 years in the study is provided (
Figure 8). We also provide corresponding values of the percent change between years in
Appendix A Table A7. ET change is either positive or negative (−) and the data are shown for each reach and the weighted average in All reaches (
Appendix A,
Table A7).
A boxplot shows the range of daily ET changes across reaches individually and together as “All” with the darker the box, the larger the range of change (
Figure 8a). For example, R5 shows the largest range of change, followed by R4. Following this the rate of change over years is provided (
Figure 8b). Using the values in
Appendix A,
Table A7, as an example, R5 shows the largest range of change in daily ET (−53% to 86%, followed by R4 (−55% to 77%).
Figure 8 shows that the largest increase in ET was 2004–2005 with 160.8 mmyr
−1, followed by 2007–2008 with 152.2 mmyr
−1. The largest loss in ET was 189.5 mmyr
−1 for the year 2002–2003, followed by the second largest loss in ET, 134 mmyr
−1 for the year 2015–2016. The year-to-year percent change shows a decrease of as much as −51.12% between 2015 and 2016; however, the following year, 2016–2017, shows as much as a 69.53% increase in average daily ET.
Too much variance in the year-to-year change data makes it difficult to quantify the overall ecosystem change trends. Therefore, we employed seven groups of years for evaluation of the ET metric. We then averaged the daily water use for seven groups of years in the study (2000–2020, 2000–2010, 2011–2020, 2000–2005, 2006–2010, 2011–2015, and 2016–2020) and reported these averages for each reach and the weighted average for All reaches (
Table 2).
These average daily ET values for each reach and the entire riparian corridor (All) (
Table 2) are as follows: 3.24 mmd
−1 for the full study period, 2000–2020; 3.72 mmd
−1 for the first decade, a full 1 mmd
−1 less; 2.71 mmd
−1 for the most recent decade; and a steady decrease for the past four approximately five-year periods (see
Figure 9). These periods dropped from 3.78 mmd
−1 (2000–2005) to 3.66 mmd
−1 (2006–2010) to 2.87 mmd
−1 (2011–2015) to 2.54 mmd
−1 (2016–2020). Importantly, for all the riparian vegetation over these four periods of time since 2000, the average daily ET values were all under 4.5 mmd
−1. We plotted daily ET for the four groups of approximately five-year periods in a bar graph for individual reaches and All reaches (
Figure 9).
Finally, we quantified the difference in the average daily water use using ET for the peak growing season for grouped years to be compared in two periods. We used the following temporal groups: 2000–2010, 2011–2020, 2000–2005, 2006–2010, 2011–2015, and 2016–2020. We then subtracted one group of years from another group as demonstrated in the first and second columns, Periods 1 and 2, to determine percent change between these groups (
Table 3). The ET percent change between periods is shown for each reach and the weighted average in All reaches.
Table 3 shows the percent change in daily ET between these six periods, the two ten-year periods, and the four approximately five-year periods, with all change being negative (−). When comparing 2000–2005 to 2006–2010, the least amount of decrease in water use, 0.12 mmd
−1, occurred in the daily ET metric, a decrease of −2.95% for All reaches. The comparison between the first and last ca. five-year periods, 2000–2005 and 2016–2020, showed the largest decrease in average daily water use, a decrease of 1.24 mmd
−1, or −32.61% as measure by the daily ET metric. Another loss in the average peak growing season daily ET was 0.79 mmd
−1 when the years 2006–2010 were compared with 2011–2015, a decrease of −21.63%. After this large percent change decrease, the years 2011–2015 compared with 2016–2020, another but smaller decrease in water use was estimated to be 0.33 mmd
−1, or a change of −11.39%. The comparison between the two decades, first and last, also yielded large declines in daily water use, an average decrease of over 1 mmd
−1, or a change of −27.30%, as measured by the daily ET metric.
3.3. ET Annually, PAM ET, from VI-Based Time-Series from 2000 to 2020
We provide annual water use data as PAM ET (mmyr−1) for the LCR riparian area. The PAM ET provides ET on an annualized basis because it uses the full-year EVI plotted as time-series data and sums the area under the phenology curve. The annualized ET captures growth of the natural vegetation that happens outside of the peak growing season. Average peak growing season ET is mainly used when estimating water use of cultivated crops and has also been applied to riparian woodlands.
PAM ET (mmyr
−1) values are plotted as a bar graph for the last two decades, 2000–2020 for the five reaches, R3 to R7, and the entire area, All (
Figure 10). Each reach is also plotted in
Appendix A,
Figure A2a–e, and the corresponding values are in
Appendix A,
Table A7. Because PAM ET is based on the area under the EVI2 curves (see
Figure 5), the decrease in annual curve magnitude has a significant impact on the annual water use totals (
Figure 10). These data also show that the very low years of 2019 and 2020 in terms of EVI2 greenness do not impact annualized water use when the ETo and temperatures are higher than normal. We can use 2005 as an example of early green-up compared with the other years, and this may be due to high precipitation in 2004 (
Figure 2); but we note that the early green-up trends appear in R3, R4, and R5, but not in R6 and R7, which could be due to the presence of MSCP and/or other restoration activities.
The annual ET cycles show oscillations that occur in declines of three years and one of five years between 2008 and 2012 (
Figure 10). In recent years, we see that the data supports the data in the delta [
14] and 2020 is increasing rather than continuing the declining trend observed between 2000 and 2019. The ET algorithm relies not only on vegetation greenness but also on weather station data (see
Figure 2). Vegetation greenness using EVI2 was the lowest ever recorded in 2020 (
Figure 5); however, ETo was high in 2020, having increased to 275 mm from 250 mm in 2019, as temperature was higher by 1–3 degrees C, and there was no rainfall. This is likely the reason for the increase in PAM ET in 2020 (
Figure 10).
Several things are important to note: (i) the 2013 data are a mix of Landsat 7 (ETM+) and Landsat 8 (OLI); (ii) the salt cedar biocontrol beetle was first observed on the LCR in 2014 but reach-wide impacts from defoliation events take several years to make a measurable difference in ET; (iii) PAM ET measured over the full year, not just the peak growing season, shows 2016 is consistently the lowest ET across reaches; and (iv) the full year PAM ET is on average a little less than 800 mmyr−1 with declining trends in four 3-year periods, then followed by an increase in ET, i.e., 2002–2004, 2005–2007, 2010–2013, 2014–2016, and 2017–2019. There is one 5-year period from 2008–2012 before the increase in ET the following year, 2013.
The absolute values of the full-year water use, PAM ET (mmyr
−1), are shown for each of the 21 years from 2000 to 2020 for five reaches and the weighted average of All reaches along the LCR (
Table 4). The initial 10-year period was in fact greener with higher values of PAM ET than the last decade as shown in
Table 4. The All column in
Table 4 shows values that began at 1034 mmyr
−1 and end below 700 mmyr
−1 until 2020, when an increase occurred and is due to higher ETo and temperatures. Because of the different composition of species and cover reach by reach, our reach level results are also shown. A plot of PAM ET (mmyr
−1) is shown for each river reach in Supplemental Information (
Appendix A). The PAM ET for R3 is in
Appendix A,
Figure A2a, R4 is A2b, R5 is A2c, R6 is A2d, and R7 is A2e.
For year-to-year changes over 21 years, the percent change in PAM ET is demonstrated with positive and negative (−) box and line plots for each of the five reaches and the weighted average of All reaches along the LCR (
Figure 11). The percent change in PAM ET is shown by box plots with the range of change highlighted by the darker the box (11a, top), and the lines indicate the rate of change between years 2000 and 2019 (11b, bottom). For example, R4 shows the largest range of change, followed by R5. Using the difference values in
Appendix A,
Table A8, as an example, R4 shows the largest range of change in PAM ET, from −221.1 mmyr
−1 to 249.4 mmyr
−1 a range between −220% and 250%, followed by R5, from −207.8 mmyr
−1 to 206.8 mmyr
−1, a range between −200% and 200%. That the percent change in PAM ET year-to-year is as much as 200% in the positive direction and 200% in the negative direction is a key finding. This large amount of change was somewhat steady between 2007 and 2013 and corresponds to the period of lower ETo, although the three stations of weather data do not seem to have significant variation. Difference in PAM ET (mmyr
−1) between years for 21 years is provided in
Appendix A Table A8. PAM ET (mmyr
−1) change is either positive or negative and the data are shown for each reach and the weighted average in All reaches (
Appendix A,
Table A8).
The PAM ET (mmyr
−1) is provided for seven groups of years is shown for the periods 2000–2020, 2000–2010, 2011–2020, 2000–2005, 2006–2010, 2011–2015, and 2016–2020 for five reaches and the weighted average of All reaches along the LCR (
Table 5). Overall, the PAM ET was 870.76 mmyr
−1 over the past two decades. The first decade, 2000–2010, the PAM ET averaged 952.14 mmyr
−1, and the first years, 2000–2005, PAM ET averaged 993.50 mmyr
−1. The water use estimates dropped in the second, more recent decade to 781.23 mmyr
−1, and in the last years, 2016–2020, PAM ET averaged 737.15 mmyr
−1. A comparison of the average changes between the first three five-year periods shows decreases in PAM ET of 90.99 mmyr
−1, 77.20 mmyr
−1, and 88.16 mmyr
−1.
Percent change in PAM ET (mmyr
−1) is shown between two periods, Periods 1 and 2, with comparisons made for groups of years 2000–2010, 2011–2020, 2000–2005, 2006–2010, 2011–2015, and 2016–2020, and for five reaches and the weighted average of All reaches along the LCR (
Table 6). The percent change between these groups of years for PAM ET is all negative change (-) and shows large, significant decreases in the estimated annualized water use (
Table 6).
Table 6 shows the percent change in annual PAM ET between six periods, the two ten-year periods, and the four approximately five-year periods. Percent changes between periods compared and across reaches are denoted with a negative sign (−) to indicate that all were decreases in PAM ET (
Table 6). Importantly, all reaches for all compared time periods have negative change. The percent change in PAM ET between the first and last 5-year periods, 2000–2005 and 2006–2010, is equal to a −9.16% decline; between 2006–2010 and 2011–2015 PAM ET is equal to a −8.55% decline; and between 2011–2015 and 2016–2020 PAM ET is equal to a −10.68% decline. The comparison between the first and last periods, 2000–2005 and 2016–2020, shows a loss in PAM ET of 256.35 mmyr
−1 or a decrease of −25.80%. A comparison between the first and last decade also reflects these large losses. Results show a loss in PAM ET of −170.91 mmyr
−1 or a decrease of −17.95% and indicate a shift toward an unhealthy riparian corridor with less green vegetation and corresponding losses in reach level riparian ET.
3.4. Change Maps from Two Periods Showing Two Metrics, Scaled NDVI, and ET
Two types of map that represent spatiotemporal change (positive and negative) over two periods of time are provided. We produce change maps using the metrics of a scaled NDVI (referred to as NDVI*) and ET that was parameterized with EVI2. The primary reason for using the NDVI* scaling method is that it takes into account changing illumination conditions scene-to-scene. NDVI* is an additional correction to the NDVI, used here because it is widely known, to account for illumination conditions. In other words, it is a processing correction step to improve the usefulness of NDVI. NDVI* is not used to parameterize the ET model. The first set of maps compares the first to the last decade (
Figure 12) and is of the NDVI* (
Figure 12a, left) and ET using EVI2 (
Figure 12b, right). The second set of maps represent change between the first 5-year period and the last 5-year period (
Figure 13) with both NDVI* (
Figure 13a, left) and ET using EVI2 (
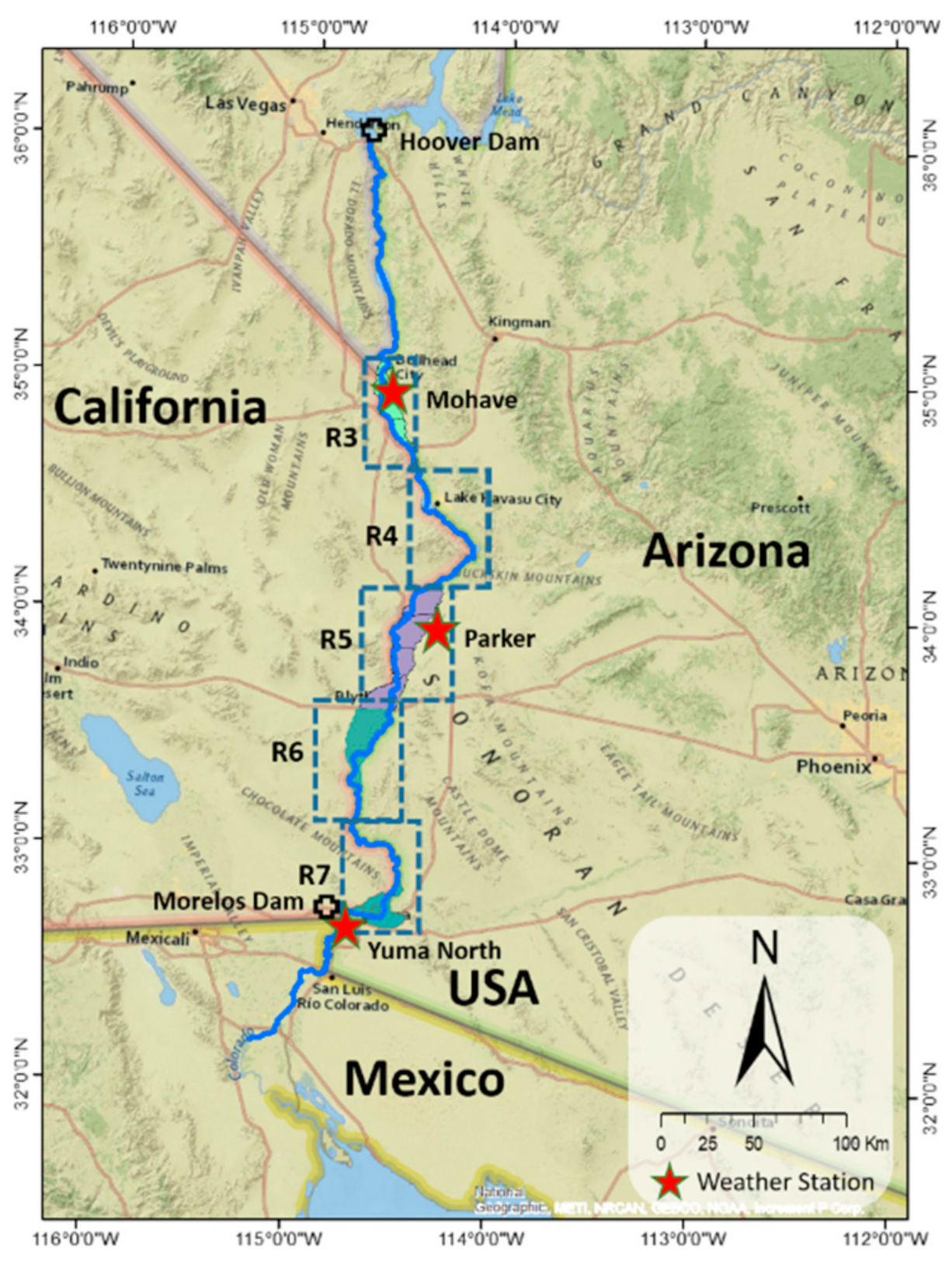
Figure 13b, right) change indicating areas of increasing and decreasing greenness. Each map has a histogram of the number of pixels in the map, and the distribution of positive and negative changes is displayed. Due to the long, narrow nature of the riparian corridor region, we show enlarged maps for the five reaches we studied, R3 to R7. Areas such as R4 had very few pixels representing riparian plants due to the physical geography of the reach.
In these sets of maps showing change in NDVI* and ET, we compare the first and last decades of our study (
Figure 12), as well as the first and last 5-year periods (
Figure 13). In individual reaches, boxes that are enlarged show R3 to R7, allowing the reader to observe changes to the riparian cover in these areas. Historically, R3 and R6 have been heavily dominated by salt cedar plants [
19,
20,
30,
45] and have been validated with on-the-ground observations [
15,
28,
73,
120]. R5 and R7 have salt cedar stands but have not been dominated by vast, monotypic swaths of salt cedar like those in R3 and R6, which are a few kilometers across in some areas. All reaches show negative trends, declines in greenness, ET, and PAM ET. R5 and R7 are not showing these dramatic declines in greenness and water use seen in R3 and R6; however, these reaches do have significant losses in vegetation greenness cover as seen in the bar graph of daily ET (
Figure 9). The percent change in PAM ET declines between the first and last 5-year periods and is smallest for R4, R7, and R5, but is still in the order of −20%. R3 shows declines of −34% and R6 declines of −29%, thus both reaches have considerably larger negative change than R5 and R7 (
Figure 10). In the box plots, PAM ET percent change is negative for each reach, but less so in R5 and R7.
Figure 12 depicts the first decade, 2000–2010, in comparison to the last, most recent decade, 2011–2020. The two reaches, R3 and R6, have displayed the greatest decline or negative change in greenness and water use. Furthermore, Reach 5 (
Appendix A,
Figure A1c) and Reach 7 (
Figure A1e) show the smallest ranges of annual cycle data; Reach 3 (
Figure A1a) and Reach 6 (
Figure A1d) show the widest ranges, with decreases prominent in the last few years (see
Appendix Figure A1a-e). These results are in line with findings for other parts of the Colorado River basin where salt cedar cover across sites declined on average of −40–50% in response to the beetle, but then recovered [
8,
23,
25,
47,
82]. However, we are now seeing that when we compare the initial five years and the last five years (
Figure 13) with the last decade (
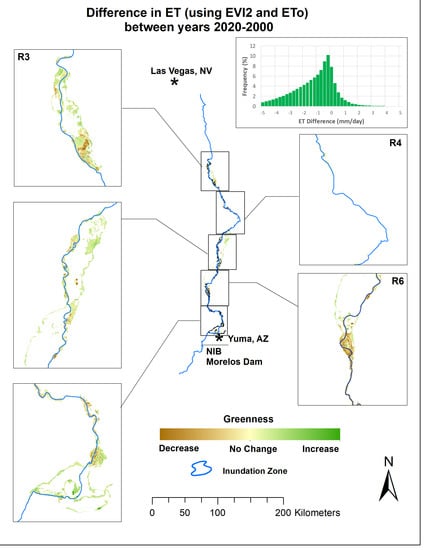
Figure 12), the reaches dominated by salt cedar have undergone decreases in vegetation and show large declines in its water use due to the eradication of plant cover that has not been replaced by regrown salt cedar or native species. From the change maps that use NDVI* and ET from EVI2, for the two periods of change (
Figure 12 and
Figure 13), we see much darker browns where salt cedar exists at Topock Marsh and Cibola NWR. In R3 and R6, we know these darker brown patches, indicating maximum browning or maximum negative change in greenness, are where salt cedar has been validated on the ground. In R5 and R7, the color indicates that there has been less browning change in comparison to the color scheme in the other reaches. Other areas near the international border and the city of Yuma, Arizona, such as R5 and R7, have experienced more subtle declines in riparian health. There has been active restoration in R7 and these small-scale and healthy areas may have positively impacted the this reach.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}