
Tracking Individual Scots Pine (Pinus sylvestris L.) Height Growth Using Multi-Temporal ALS Data from North-Eastern Poland


Abstract
:1. Introduction
2. Materials and Methods
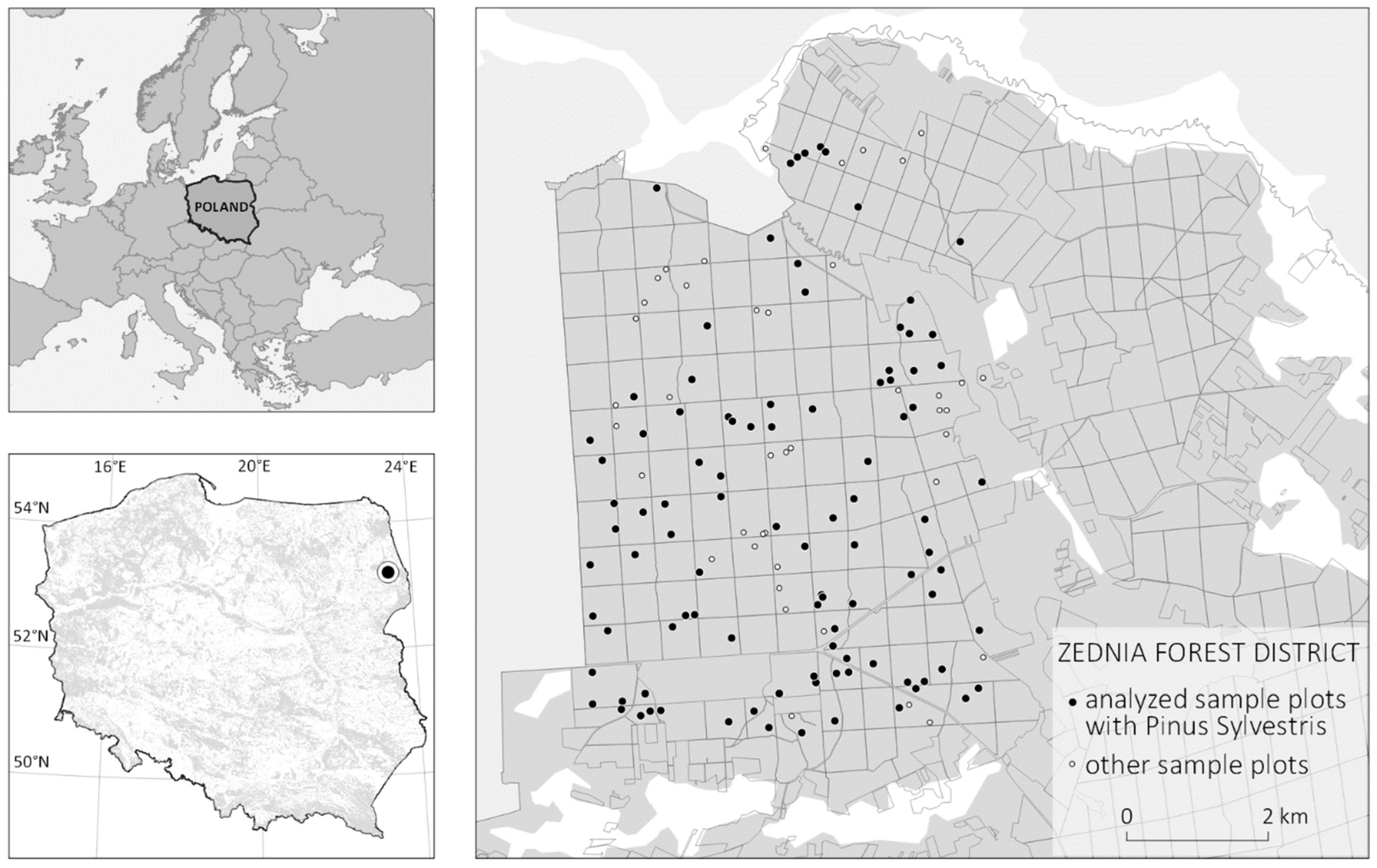
2.1. Study Area
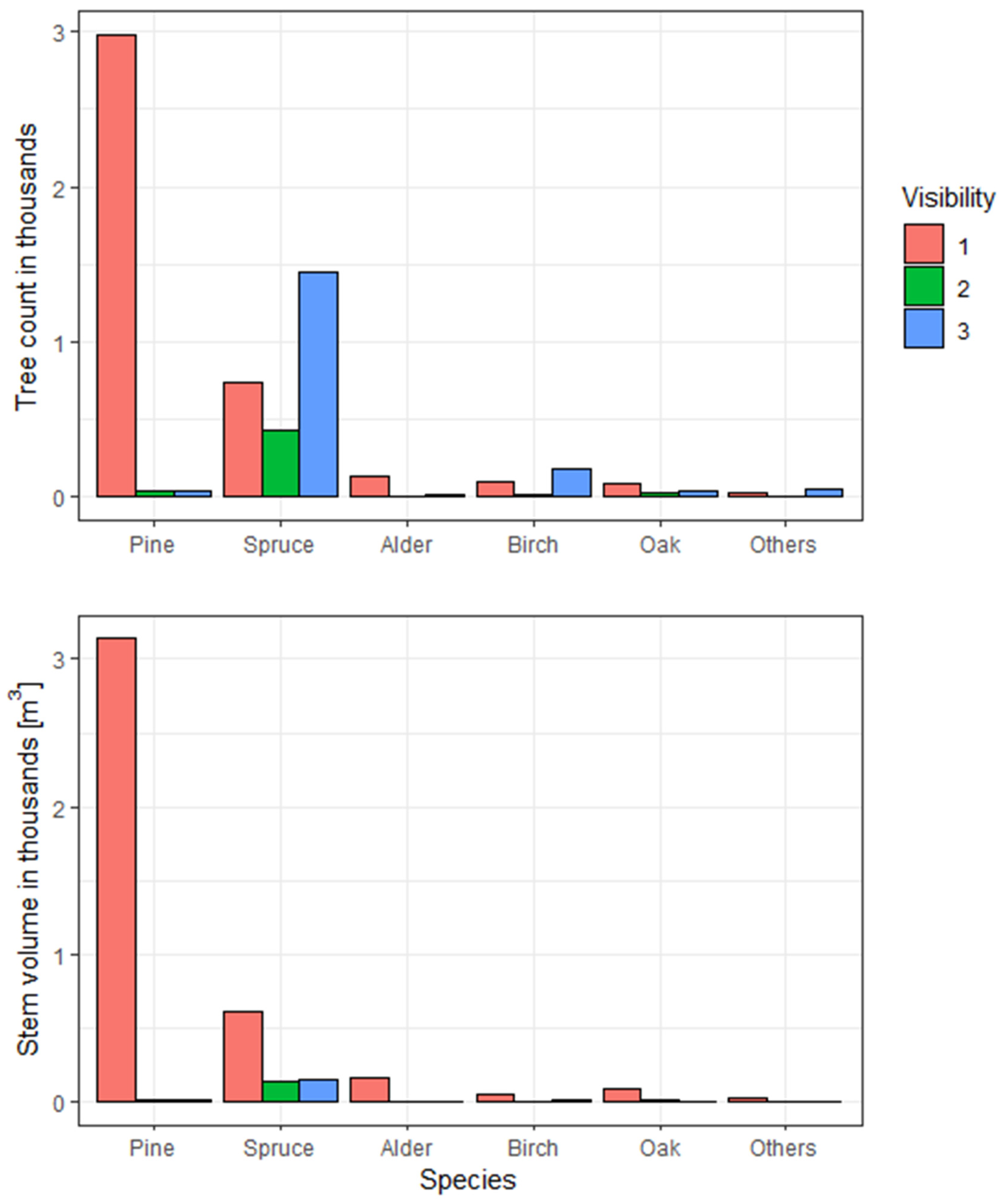
2.2. Field Inventory Data
- (a)
- HCfs—hydrogenic coniferous forest sites;
- (b)
- MCfs—mineral coniferous forest sites;
- (c)
- MMfs—mineral mixed forest sites.
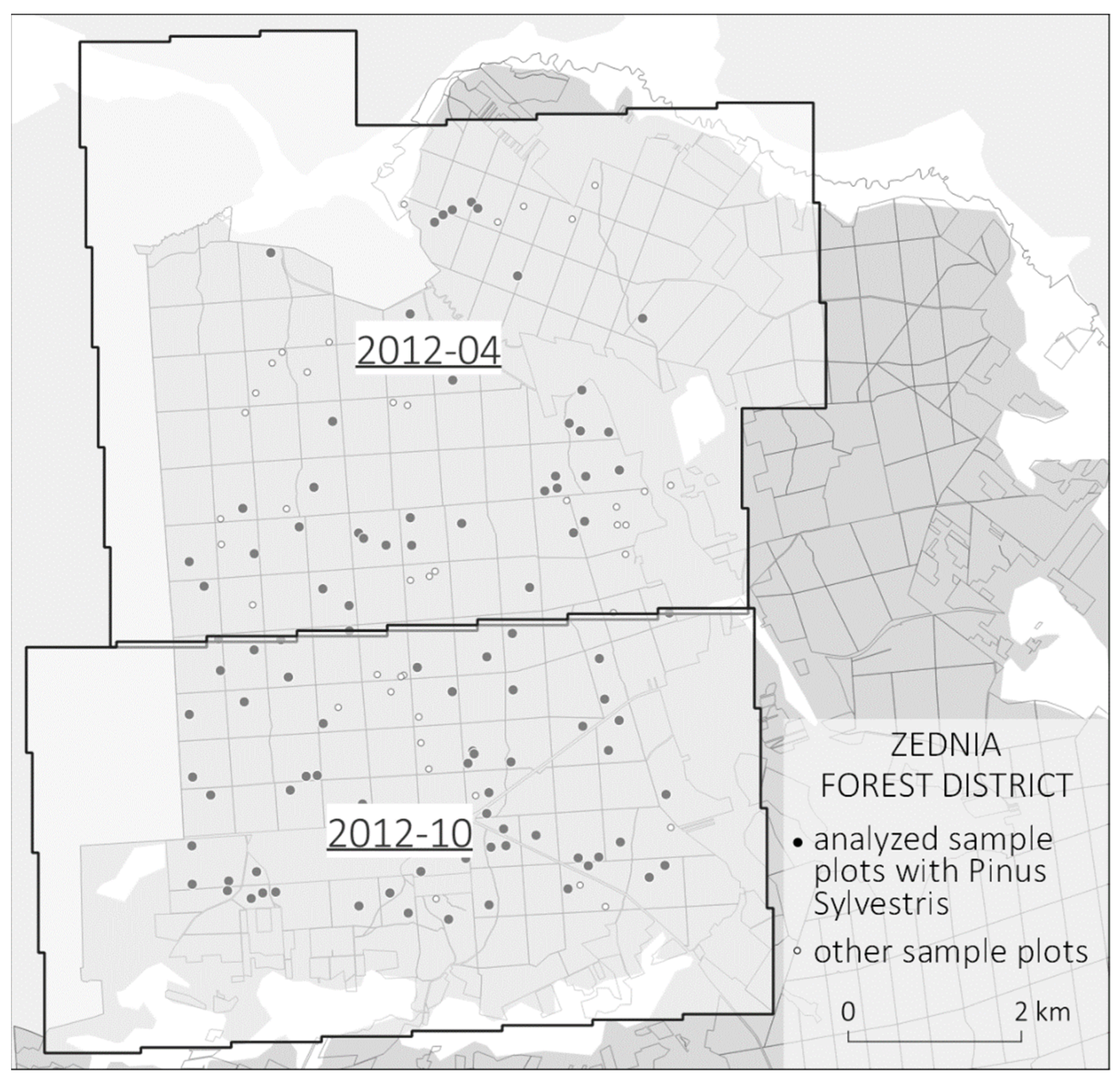
2.3. Airborne Laser Scanning Point Clouds
- (a)
- Point cloud density [pt/m2]—14.08 (2007), 8.65 (2012), 19.64 (2017);
- (b)
- Density of single and last returns [pt/m2] —6.84 (2007), 4.54 (2012), 4.06 (2017).
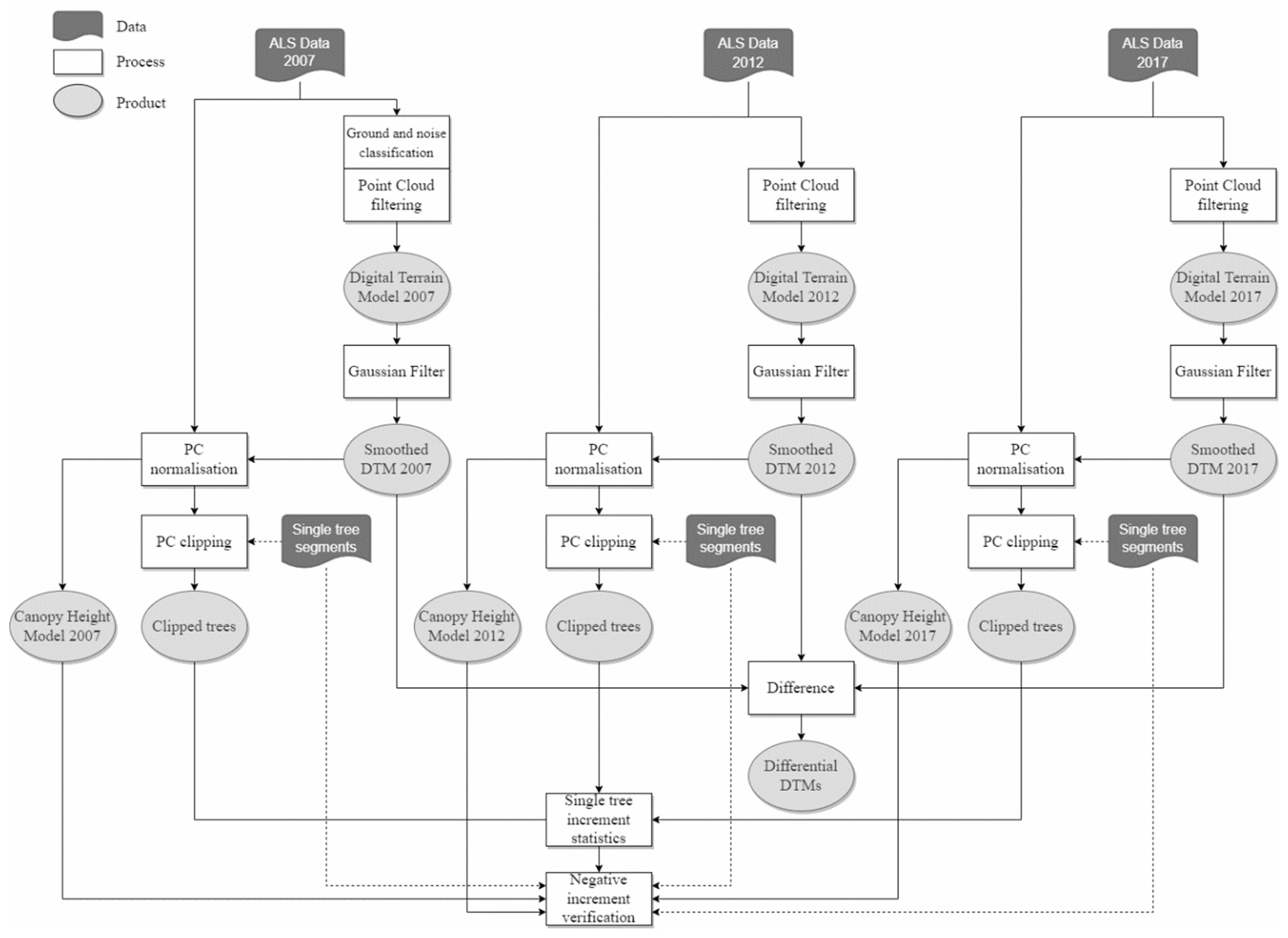
2.4. Point Clouds Processing and Derivative Products
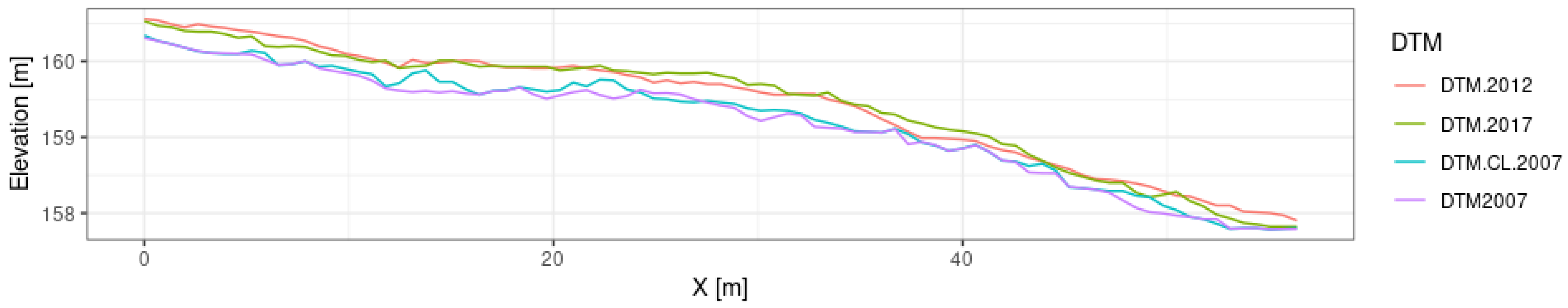
2.5. Comparative Analysis of DTM Generated from Multi-Temporal Point Clouds
2.6. Tracking individual Scots Pine Height Growth
2.7. Modeling of Individual Scots Pine Height Growth
3. Results
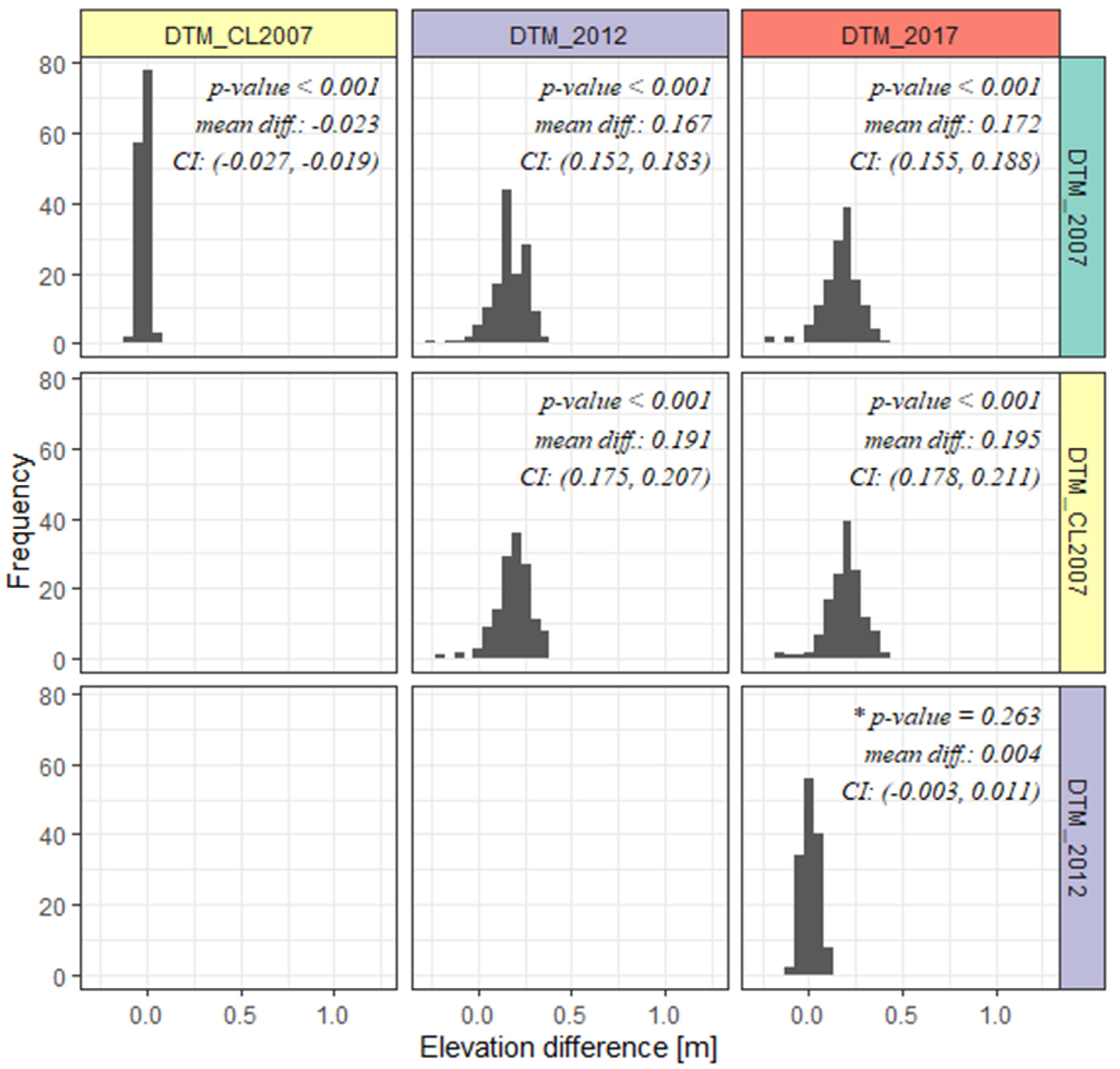
3.1. Comparative Analysis of DTMs Generated from Multi-Temporal Point Clouds
3.2. Verification of Tree Heights 2007–2017
- (a)
- Correctness/unambiguity of top detection—the use of point clouds with different density parameters, non-uniform point distribution, or even the same data acquisition technology (impulse scanning, full-wave registration) may cause the real vertex (the highest point in the segment) to not always be correctly identified on multitemporal materials. As a consequence, this may lead to the generation of negative height increments, especially if the data acquisition interval is small (1–2 years) and the analyzed trees are characterized by low annual growth dynamics.
- (b)
- Removal of individual trees (due to management activities or natural processes)—segments developed on 2017 CHM data are used as reference data in this analysis. When a neighboring tree (that was taller) was removed of the analyzed individual, a portion of crown of the taller neighboring tree that was removed may be obscuring the analyzed segment when calculating height in previous time points (2007 and/or 2012). The situation is visualized in Figure 8.
- (c)
- Terminal phase of trees—Scots pine trees entering the terminal phase are characterized by the priority of growth in thickness and not in height; moreover, their tops show a tendency to bend, which, in the examined time interval, may give minimal or even negative growth in height.
- (d)
- (e)
- Incorrectly determined (delineated) crown contour boundary, which results in incorrect height statistics and uncertainty in tree growth analysis.
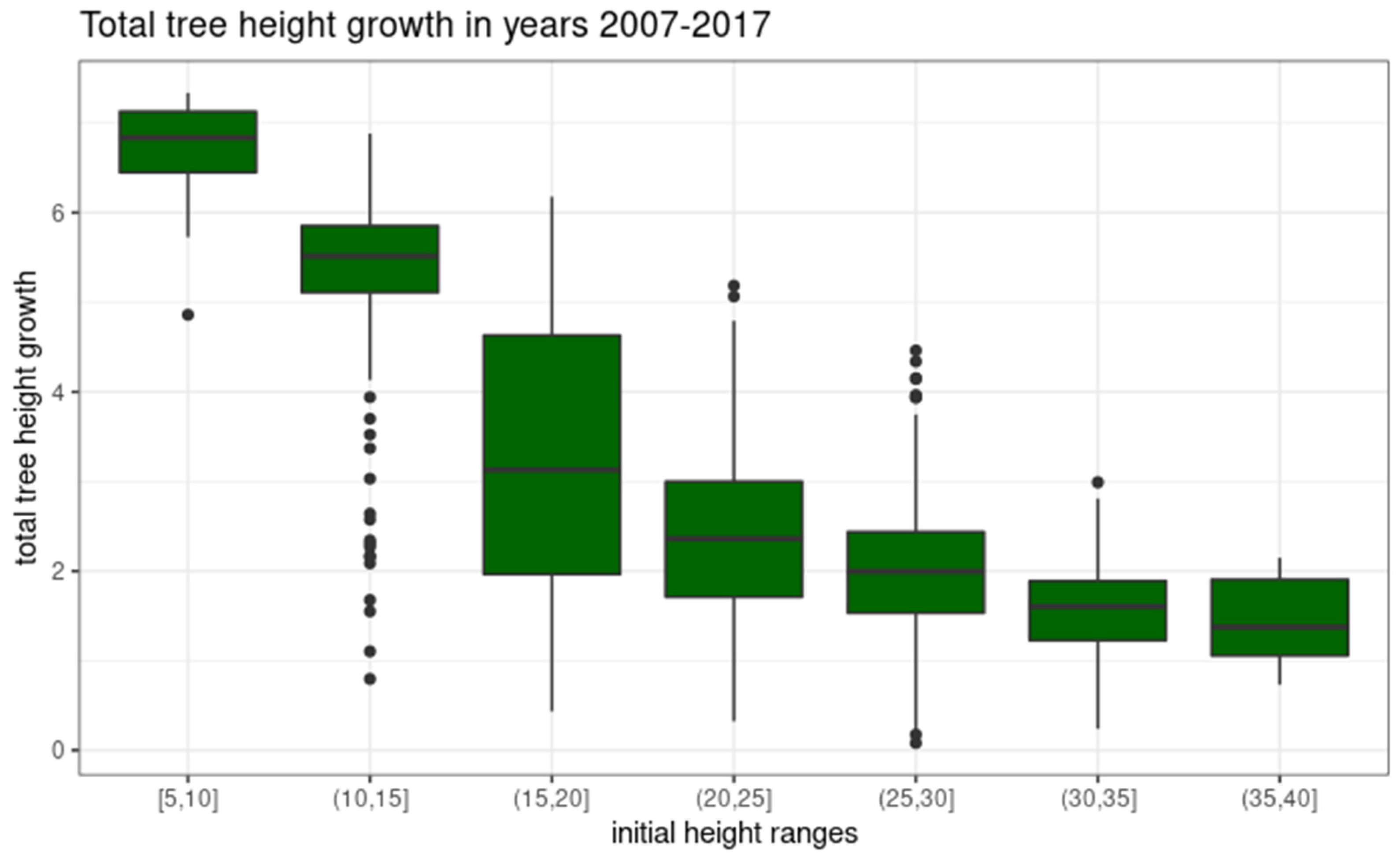
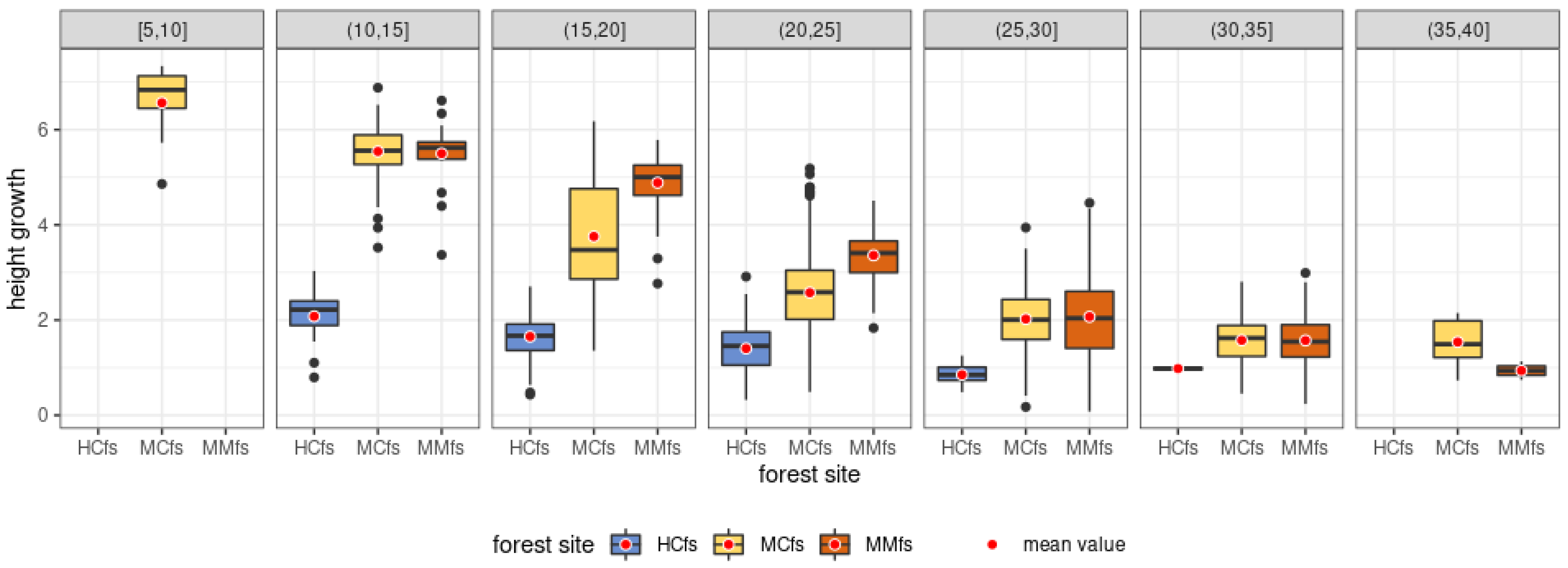
3.3. Scots Pine Height Growth Rate in 2007–2017 for the Whole Sample
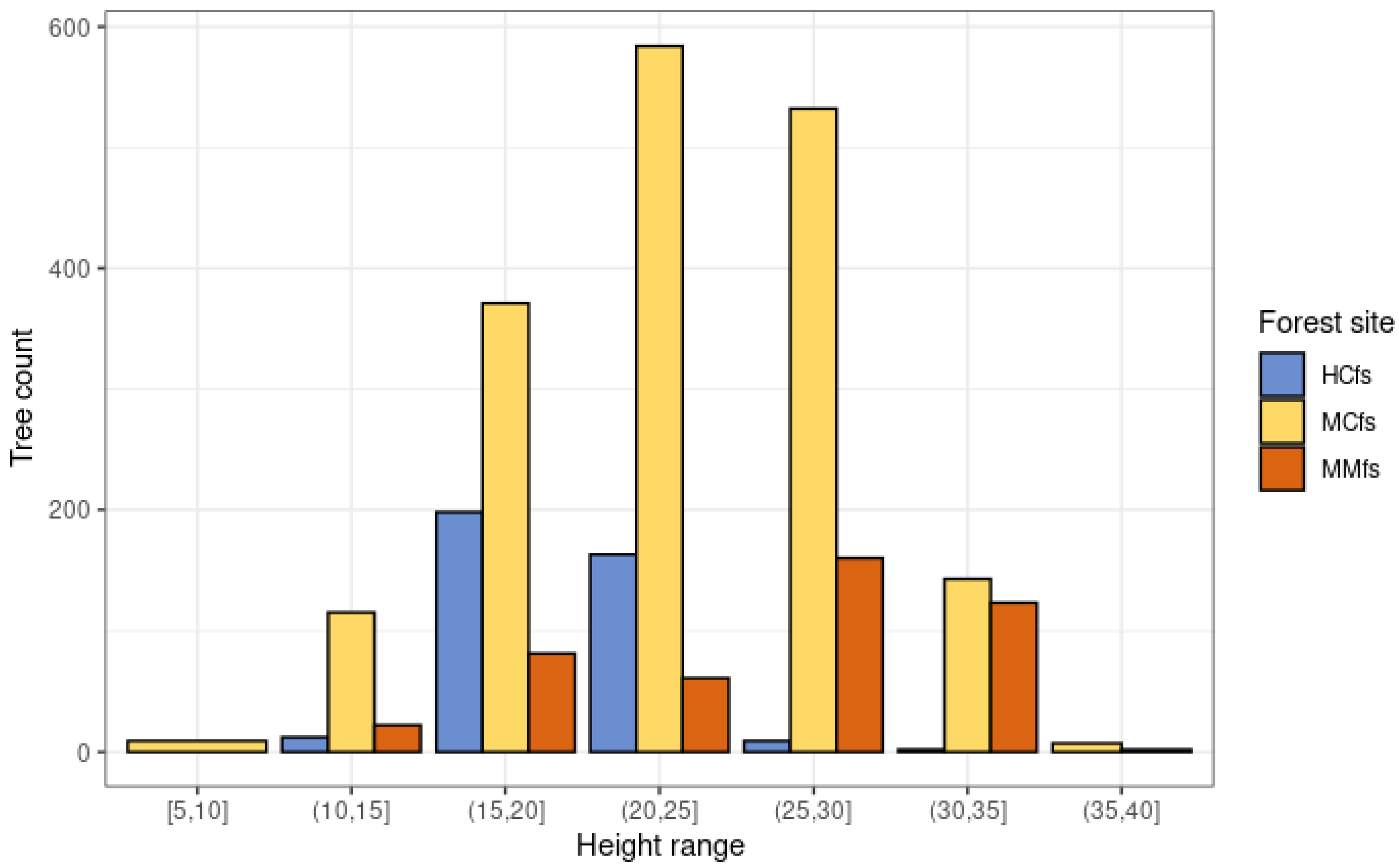
3.4. Average Scots Pine Height Growth Rate in 2007–2017 across Different Forest Site Groups
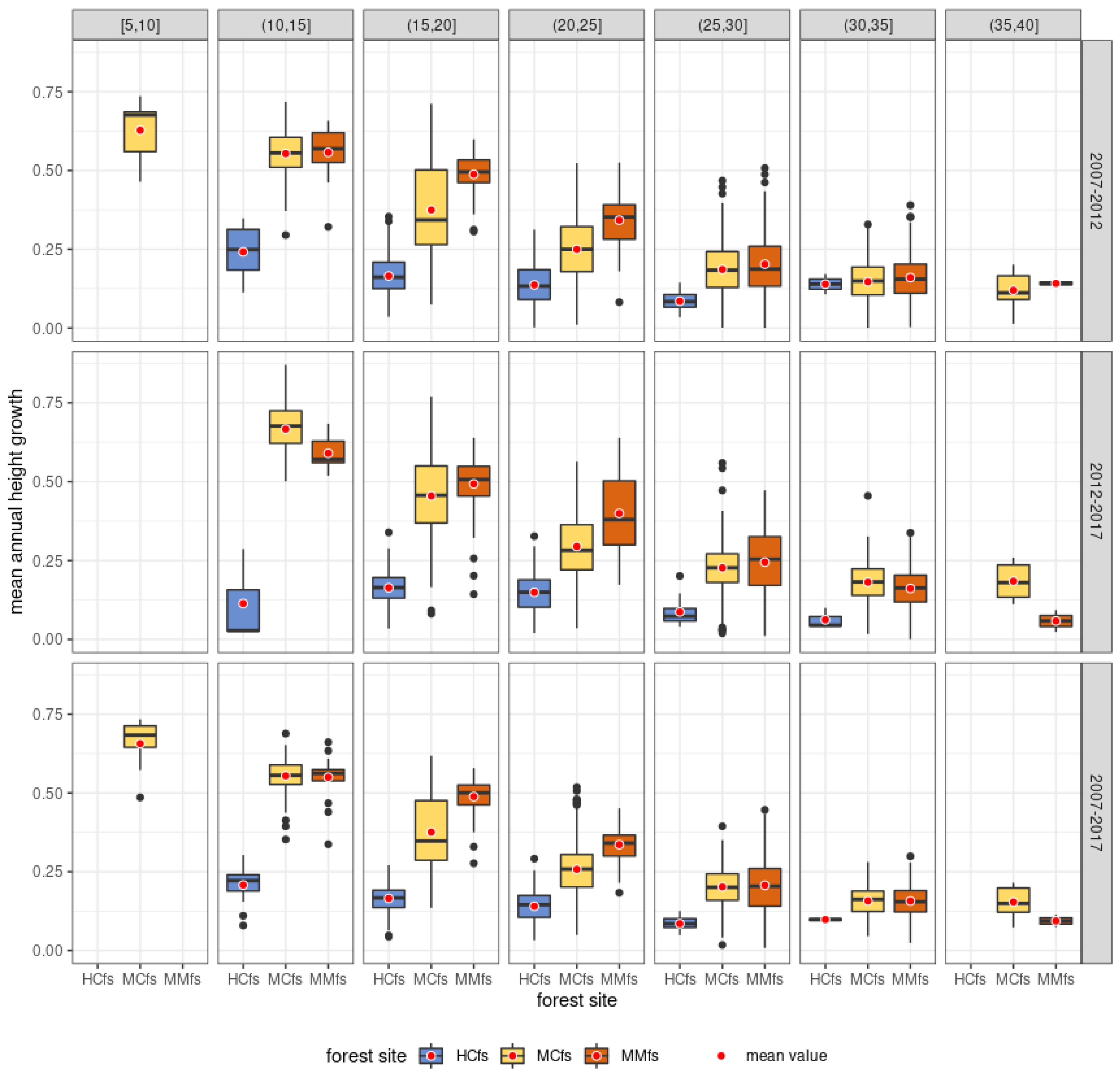
3.5. Dynamics of Annual Height Growth Rate in Time Ranges 2007–2012 and 2012–2017
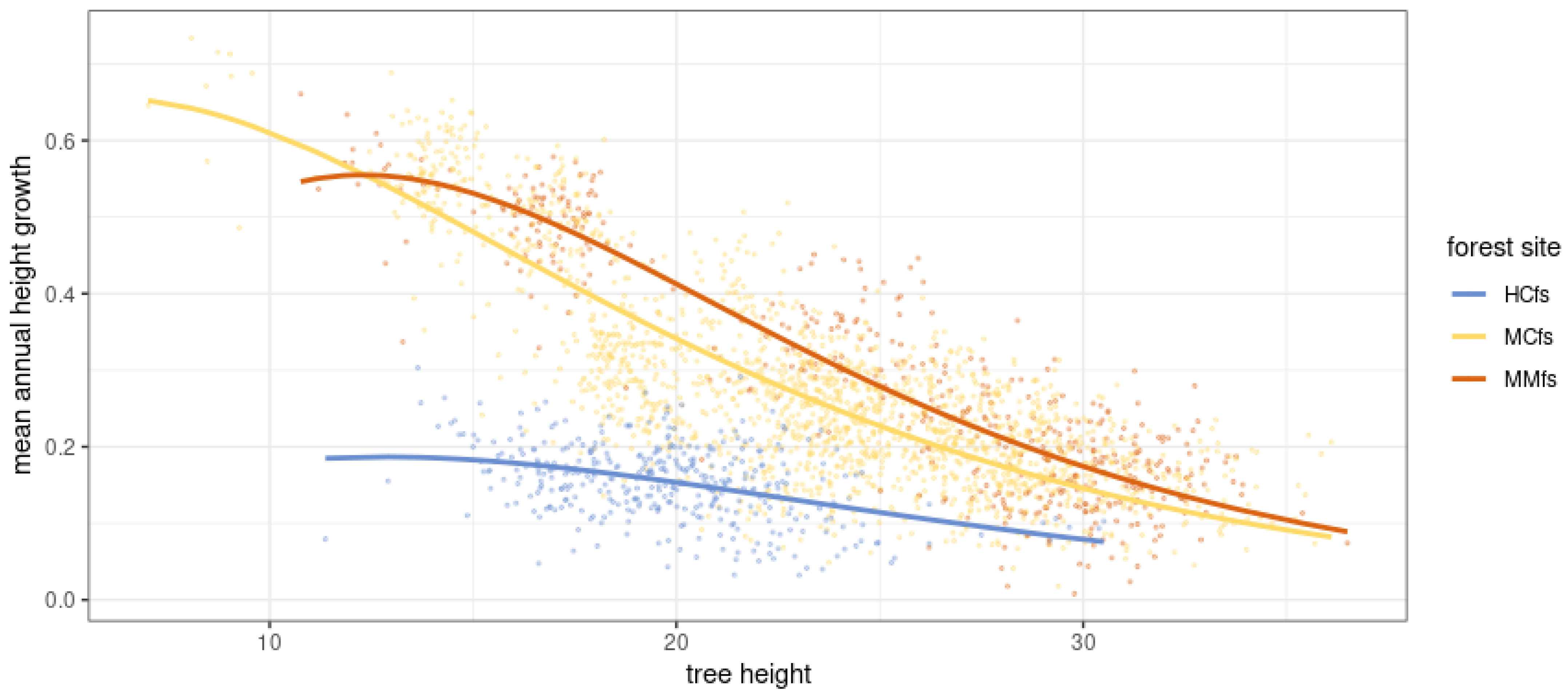
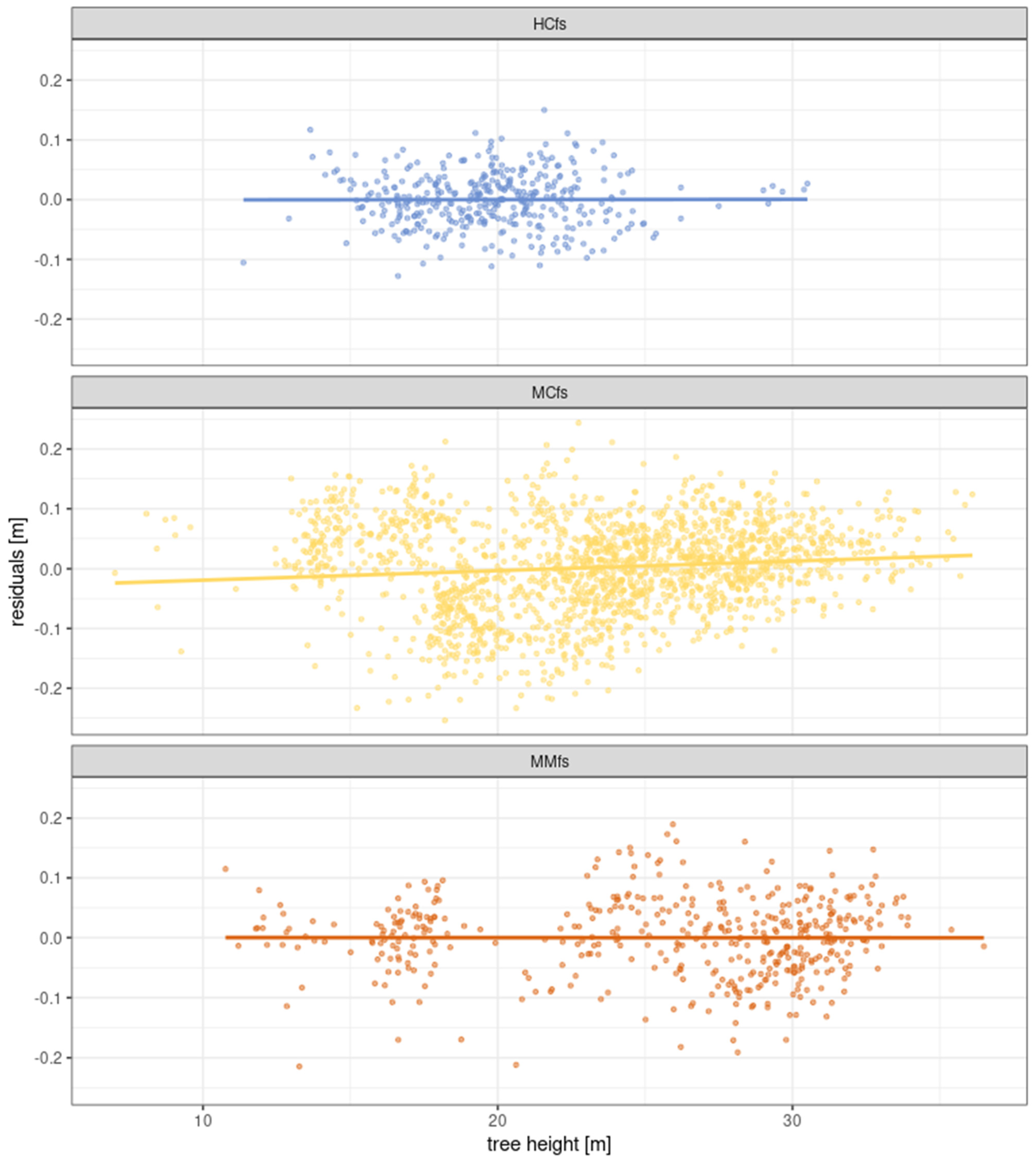
3.6. Modeling Scots Pine Height Growth including Forest Site Variability—Fitting a Gompetrz Curve
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jiang, Z.D.; Owens, P.R.; Ashworth, A.J.; Fuentes, B.A.; Thomas, A.L.; Sauer, T.J.; Wang, Q.B. Evaluating tree growth factors into species-specific functional soil maps for improved agroforestry system efficiency. Agrofor. Syst. 2022, 96, 479–490. [Google Scholar] [CrossRef]
- Hynynen, J.; Repola, J.; Mielikäinen, K. The effects of species mixture on the growth and yield of mid-rotation mixed stands of Scots pine and silver birch. For. Ecol. Manag. 2011, 262, 1174–1183. [Google Scholar] [CrossRef]
- Borowski, M. Przyrost Drzew I Drzewostanow; Panstwowe Wydawnictwo Rolnicze i Lesne: Warszawa, Poland, 1974. [Google Scholar]
- Gizachew, B.; Brunner, A. Density-growth relationships in thinned and unthinned Norway spruce and Scots pine stands in Norway. Scand. J. For. Res. 2011, 26, 543–554. [Google Scholar] [CrossRef]
- Rudnicki, M.; Silins, U.; Lieffers, V.J. Crown cover is correlated with relative density, tree slenderness, and tree height in lodgepole pine. For. Sci. 2004, 50, 356–363. [Google Scholar]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Sibona, E.; Vitali, A.; Meloni, F.; Caffo, L.; Dotta, A.; Lingua, E.; Motta, R.; Garbarino, M. Direct measurement of tree height provides different results on the assessment of LiDAR accuracy. Forests 2016, 8, 7. [Google Scholar] [CrossRef]
- Fardusi, M.J.; Chianucci, F.; Barbati, A. Concept to practice of geospatial-information tools to assist forest management and planning under precision forestry framework: A review. Ann. Silvic. Res. 2017, 41, 3–14. [Google Scholar]
- Wunder, J.; Brzeziecki, B.; Żybura, H.; Reineking, B.; Bigler, C.; Bugmann, H. Growth-mortality relationships as indicators of life-history strategies: A comparison of nine tree species in unmanaged European forests. Oikos 2008, 117, 815–828. [Google Scholar] [CrossRef]
- Zhen, Z.; Quackenbush, L.J.; Zhang, L. Trends in automatic individual tree crown detection and delineation—Evolution of LiDAR data. Remote Sens. 2016, 8, 333. [Google Scholar] [CrossRef]
- Moskal, L.M.; Erdody, T.; Kato, A.; Richardson, J.; Zheng, G.; Briggs, D. Lidar applications in precision forestry. In Proceedings of the Silvilaser 2009, College Station, TX, USA, 14–16 October 2009; pp. 154–163. [Google Scholar]
- Goodbody, T.R.; Coops, N.C.; Marshall, P.L.; Tompalski, P.; Crawford, P. Unmanned aerial systems for precision forest inventory purposes: A review and case study. For. Chron. 2017, 93, 71–81. [Google Scholar] [CrossRef]
- Instytut Badawczy Leśnictwa (Forest Research Institute). Remote Sensing Based Assessment of Woody Biomass and Carbon Storage in Forests (RemBioFor). 2015. Available online: http://rembiofor.pl/en/305-2/ (accessed on 1 May 2022).
- Instytut Badawczy Leśnictwa (Forest Research Institute). Comprehensive Monitoring of Stand Dynamics in Białowieża Forest Supported with Remote Sensing Techniques (ForBioSensing). 2014. Available online: http://www.forbiosensing.pl/about-project (accessed on 1 May 2022).
- Tymińska-Czabańska, L.; Hawryło, P.; Socha, J. Assessment of the effect of stand density on the height growth of Scots pine using repeated ALS data. Int. J. Appl. Earth Obs. Geoinf. 2022, 108, 102763. [Google Scholar] [CrossRef]
- Zawieja, B.; Kazmierczak, K. Longitudinal analysis of annual height increment differentiation in Scots pine (Pinus sylvestris L.) stands of different age classes. Folia For. Polonica. Ser. A For. 2014, 56, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Hyyppä, J.; Kaartinen, H.; Hyyppä, H.; Maltamo, M.; Rönnholm, P. Measuring the growth of individual trees using multi-temporal airborne laser scanning point clouds. In Proceedings of the ISPRS Workshop Laser Scanning, Enschede, The Netherlands, 12–14 September 2005; Volume 2005, pp. 204–208. [Google Scholar]
- Ma, Q.; Su, Y.; Tao, S.; Guo, Q. Quantifying individual tree growth and tree competition using bi-temporal airborne laser scanning data: A case study in the Sierra Nevada Mountains, California. Int. J. Digit. Earth 2018, 11, 485–503. [Google Scholar] [CrossRef]
- Wang, Y.; Lehtomäki, M.; Liang, X.; Pyörälä, J.; Kukko, A.; Jaakkola, A.; Liu, J.; Feng, Z.; Chen, R.; Hyyppä, J. Is field-measured tree height as reliable as believed—A comparison study of tree height estimates from field measurement, airborne laser scanning and terrestrial laser scanning in a boreal forest. ISPRS J. Photogramm. Remote Sens. 2019, 147, 132–145. [Google Scholar] [CrossRef]
- Loehle, C.; Solarik, K.A. Forest growth trends in Canada. For. Chron. 2019, 95, 183–195. [Google Scholar] [CrossRef]
- Kazmierczak, K. The current growth increment of pine tree stands comprising three different age classes. Leśne Pr. Badaw. 2013, 74, 93. [Google Scholar] [CrossRef]
- Arumäe, T.; Lang, M.; Laarmann, D. Thinning-and tree-growth-caused changes in canopy cover and stand height and their estimation using low-density bitemporal airborne lidar measurements–a case study in hemi-boreal forests. Eur. J. Remote Sens. 2020, 53, 113–123. [Google Scholar] [CrossRef]
- Kamińska, A.; Lisiewicz, M.; Stereńczak, K. Single Tree Classification Using Multi-Temporal ALS Data and CIR Imagery in Mixed Old-Growth Forest in Poland. Remote Sens. 2021, 13, 5101. [Google Scholar] [CrossRef]
- Zajączkowski, G.; Jabłoński, M.; Jabłoński, T.; Szmidla, H.; Kowalska, A.; Małachowska, J.; Piwnicki, J. Raport o Stanie Lasów W Polsce 2019 [Report on the State of Forests in Poland 2019]; Centrum Informacyjne Lasów Państwowych: Warsaw, Poland, 2020. [Google Scholar]
- Evans, D. Building the European Union’s Natura 2000 network. Nat. Conserv. 2012, 1, 11–26. [Google Scholar] [CrossRef]
- Gajko, K.; Myszkowski, M.; Ksepko, M. Eksperyment w obrębie Zajma. Geod. Mag. Geoinformacyjny 2009, 1, 60–62. [Google Scholar]
- Kolendo, Ł.; Ksepko, M. Selection of optimal tree top detection parameters in a context of effective forest management. Ekonomia i Środowisko 2019, 68, 67–85. [Google Scholar]
- Dyrekcja Generalna Lasów Państwowych (General Directorate of the State Forests). Instrukcja Urządzania Lasu. Część II [Instruction for Forest Management. Part II]; Ośrodek Rozwojowo-Wdrożeniowy Lasów Państwowych: Bedoń, Poland, 2012. [Google Scholar]
- Stereńczak, K.; Kraszewski, B.; Mielcarek, M.; Piasecka, Ż.; Lisiewicz, M.; Heurich, M. Mapping individual trees with airborne laser scanning data in an European lowland forest using a self-calibration algorithm. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102191. [Google Scholar] [CrossRef]
- BULiGL. Operat Glebowo-Siedliskowy dla Obszaru Nadleśnictwa Żednia. In Soil and Forest Site Survey for the Żednia Forest District; 2018; Unpublished Report. [Google Scholar]
- Vanguelova, E.I.; Nortcliff, S.; Moffat, A.J.; Kennedy, F. Morphology, biomass and nutrient status of fine roots of Scots pine (Pinus sylvestris) as influenced by seasonal fluctuations in soil moisture and soil solution chemistry. Plant Soil 2005, 270, 233–247. [Google Scholar] [CrossRef]
- Wężyk, P. Podręcznik dla uczestników szkoleń z wykorzystania produktów LiDAR. In The Handbook for LiDAR Data Utilization Training; Główny Urząd Geodezji i Kartografii: Warsaw, Poland, 2014. [Google Scholar]
- Państwowe Gospodarstwo Wodne Wody Polskie Informatyczny System Ochrony Kraju. 2009. Available online: https://isok.gov.pl/o-projekcie.html (accessed on 2 June 2022).
- American Society for Photogrammetry and Remote Sensing. LAS Specification. Version 1.2. 2008. Available online: https://www.asprs.org/wp-content/uploads/2010/12/asprs_las_format_v12.pdf (accessed on 1 February 2022).
- Roussel, J.R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.; Meador, A.S.; Bourdon, J.F.; de Boissieu, F.; Achim, A. lidR: An R package for analysis of Airborne Laser Scanning (ALS) data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Zhang, K.; Chen, S.C.; Whitman, D.; Shyu, M.L.; Yan, J.; Zhang, C. A progressive morphological filter for removing nonground measurements from airborne LIDAR data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 872–882. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for automated geoscientific analyses (SAGA) v. 2.1. 4. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef]
- Kolendo, Ł.; Kozniewski, M.; Ksepko, M.; Chmur, S.; Neroj, B. Parameterization of the Individual Tree Detection Method Using Large Dataset from Ground Sample Plots and Airborne Laser Scanning for Stands Inventory in Coniferous Forest. Remote Sens. 2021, 13, 2753. [Google Scholar] [CrossRef]
- Miklewska, J. Including the growth function into the renewable resources optimal control models. Folia Universitatis Agriculturae Stetinensis. Oeconomica 2007, 48, 233–242. [Google Scholar]
- Jarosz, K.; Klapec, B. Modelowanie wzrostu drzewostanow z wykorzystaniem funkcji Gompertza. Sylwan 2002, 146, 35–42. [Google Scholar]
- El-Gohary, A.; Alshamrani, A.; Al-Otaibi, A.N. The generalized Gompertz distribution. Appl. Math. Model. 2013, 37, 13–24. [Google Scholar] [CrossRef]
- Hanusz, Z.; Siarkowski, Z.; Ostrowski, K. Zastosowanie modelu Gompertz’a w inżynierii rolniczej. Inżynieria Rol. 2008, 12, 71–77. [Google Scholar]
- Rymuza, K.; Bombik, A.; Radzka, E. Zastosowanie wybranych nieliniowych modeli do opisu wzrostu rutwicy wschodniej (Galega orientalis Lam.). Acta Agrophysica 2018, 25, 373–383. [Google Scholar] [CrossRef]
- Mielcarek, M.; Kamińska, A.; Stereńczak, K. Digital Aerial Photogrammetry (DAP) and Airborne Laser Scanning (ALS) as Sources of Information about Tree Height: Comparisons of the Accuracy of Remote Sensing Methods for Tree Height Estimation. Remote Sens. 2020, 12, 1808. [Google Scholar] [CrossRef]
- Sačkov, I.; Hlasny, T.; Bucha, T.; Juriš, M. Integration of tree allometry rules to treetops detection and tree crowns delineation using airborne lidar data. Iforest-Biogeosciences For. 2017, 10, 459–467. [Google Scholar] [CrossRef]
- Lisiewicz, M.; Kamińska, A.; Stereńczak, K. Recognition of specified errors of Individual Tree Detection methods based on Canopy Height Model. Remote Sens. Appl.: Soc. Environ. 2022, 25, 100690. [Google Scholar] [CrossRef]
- Lisiewicz, M.; Kamińska, A.; Kraszewski, B.; Stereńczak, K. Correcting the Results of CHM-Based Individual Tree Detection Algorithms to Improve Their Accuracy and Reliability. Remote Sens. 2022, 14, 1822. [Google Scholar] [CrossRef]
- Van Lier, O.R.; Luther, J.E.; White, J.C.; Fournier, R.A.; Côté, J. Effect of scan angle on ALS metrics and area-based predictions of forest attributes for balsam fir dominated stands. Forestry 2022, 95, 49–72. [Google Scholar] [CrossRef]
- Keränen, J.; Maltamo, M.; Packalen, P. Effect of flying altitude, scanning angle and scanning mode on the accuracy of ALS based forest inventory. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 349–360. [Google Scholar] [CrossRef]
- Socha, J.; Solberg, S.; Tymińska-Czabańska, L.; Tompalski, P.; Vallet, P. Height growth rate of Scots pine in Central Europe increased by 29% between 1900 and 2000 due to changes in site productivity. For. Ecol. Manag. 2021, 490, 119102. [Google Scholar] [CrossRef]
- Socha, J.; Tymińska-Czabańska, L.; Bronisz, K.; Zięba, S.; Hawryło, P. Regional height growth models for Scots pine in Poland. Sci. Rep. 2021, 11, 10330. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height Range [m] | Min | P5 | P25 | Median | Mean | P75 | P95 | Max | Count |
|---|---|---|---|---|---|---|---|---|---|
| 5–10 | 4.86 | 5.21 | 6.45 | 6.84 | 6.56 | 7.13 | 7.26 | 7.34 | 9 |
| 10–15 | 0.80 | 2.44 | 5.13 | 5.47 | 5.25 | 5.81 | 6.30 | 6.88 | 129 |
| 15–20 | 0.48 | 1.25 | 1.98 | 3.30 | 3.39 | 4.82 | 5.58 | 6.37 | 606 |
| 20–25 | 0.32 | 0.94 | 1.70 | 2.36 | 2.42 | 3.05 | 3.99 | 6.01 | 770 |
| 25–30 | 0.18 | 0.99 | 1.61 | 2.08 | 2.11 | 2.58 | 3.24 | 4.46 | 711 |
| 30–35 | 0.08 | 0.73 | 1.27 | 1.62 | 1.62 | 1.95 | 2.53 | 3.14 | 353 |
| 35–40 | 0.73 | 0.74 | 1.06 | 1.14 | 1.37 | 1.91 | 2.08 | 2.15 | 16 |
| sum | 2594 |
| Forest Site Group | Min | P5 | P25 | Median | Mean | P75 | P95 | Max | Count |
|---|---|---|---|---|---|---|---|---|---|
| HCfs | 0.32 | 0.66 | 1.21 | 1.56 | 1.54 | 1.85 | 2.31 | 3.03 | 384 |
| MCfs | 0.18 | 1.15 | 1.87 | 2.52 | 2.79 | 3.31 | 5.53 | 7.34 | 1761 |
| MMfs | 0.08 | 0.85 | 1.54 | 2.36 | 2.78 | 3.91 | 5.42 | 6.61 | 449 |
| sum | 2594 |
| Forest Site Group | a | b | c | R2 | adj. R2 | MAE | RMSE |
|---|---|---|---|---|---|---|---|
| HCfs | −0.0972 | 5.2243 | 12.7975 | 0.1621 | 0.1554 | 0.0356 | 0.0450 |
| MCfs | −0.0733 | 39815 | −32.2856 | 0.6757 | 0.6752 | 0.0593 | 0.0745 |
| MCfs (bounded) | −0.1006 | 17.7132 | 6.0000 | 0.6644 | 0.6638 | 0.0609 | 0.0760 |
| MMfs | −0.1143 | 13.2014 | 12.2731 | 0.8136 | 0.8124 | 0.0503 | 0.0647 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozniewski, M.; Kolendo, Ł.; Ksepko, M.; Chmur, S. Tracking Individual Scots Pine (Pinus sylvestris L.) Height Growth Using Multi-Temporal ALS Data from North-Eastern Poland. Remote Sens. 2022, 14, 4170. https://doi.org/10.3390/rs14174170
Kozniewski M, Kolendo Ł, Ksepko M, Chmur S. Tracking Individual Scots Pine (Pinus sylvestris L.) Height Growth Using Multi-Temporal ALS Data from North-Eastern Poland. Remote Sensing. 2022; 14(17):4170. https://doi.org/10.3390/rs14174170
Chicago/Turabian StyleKozniewski, Marcin, Łukasz Kolendo, Marek Ksepko, and Szymon Chmur. 2022. "Tracking Individual Scots Pine (Pinus sylvestris L.) Height Growth Using Multi-Temporal ALS Data from North-Eastern Poland" Remote Sensing 14, no. 17: 4170. https://doi.org/10.3390/rs14174170