
Differentiation of Six Grassland/Forage Types in Three Canadian Ecoregions Based on Spectral Characteristics


Abstract
:1. Introduction
2. Materials and Methods
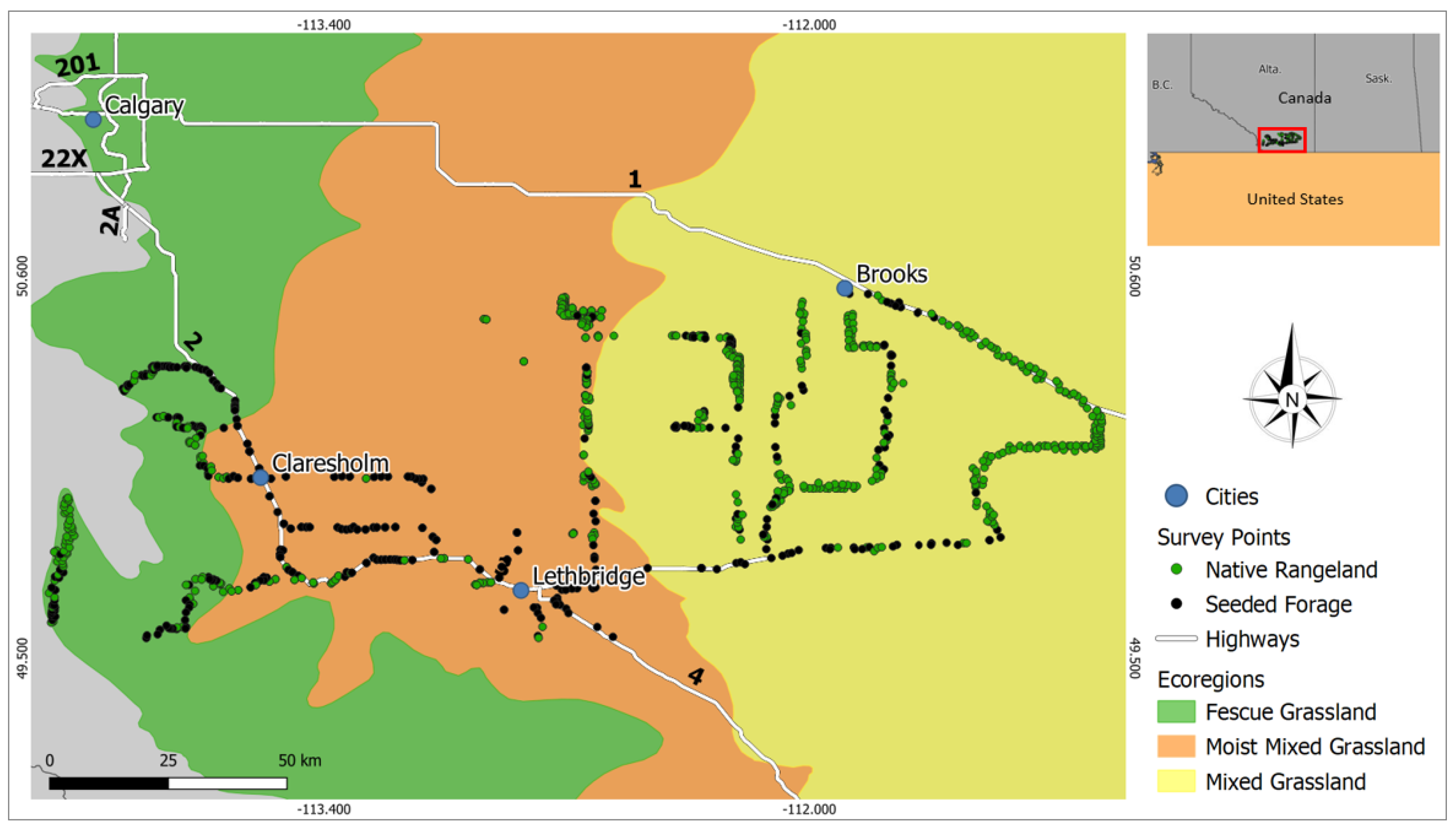
2.1. Study Area
2.2. Reference Data Collection from Surveys
2.3. Image Acquisition and Preprocessing
2.4. Statistical Analysis
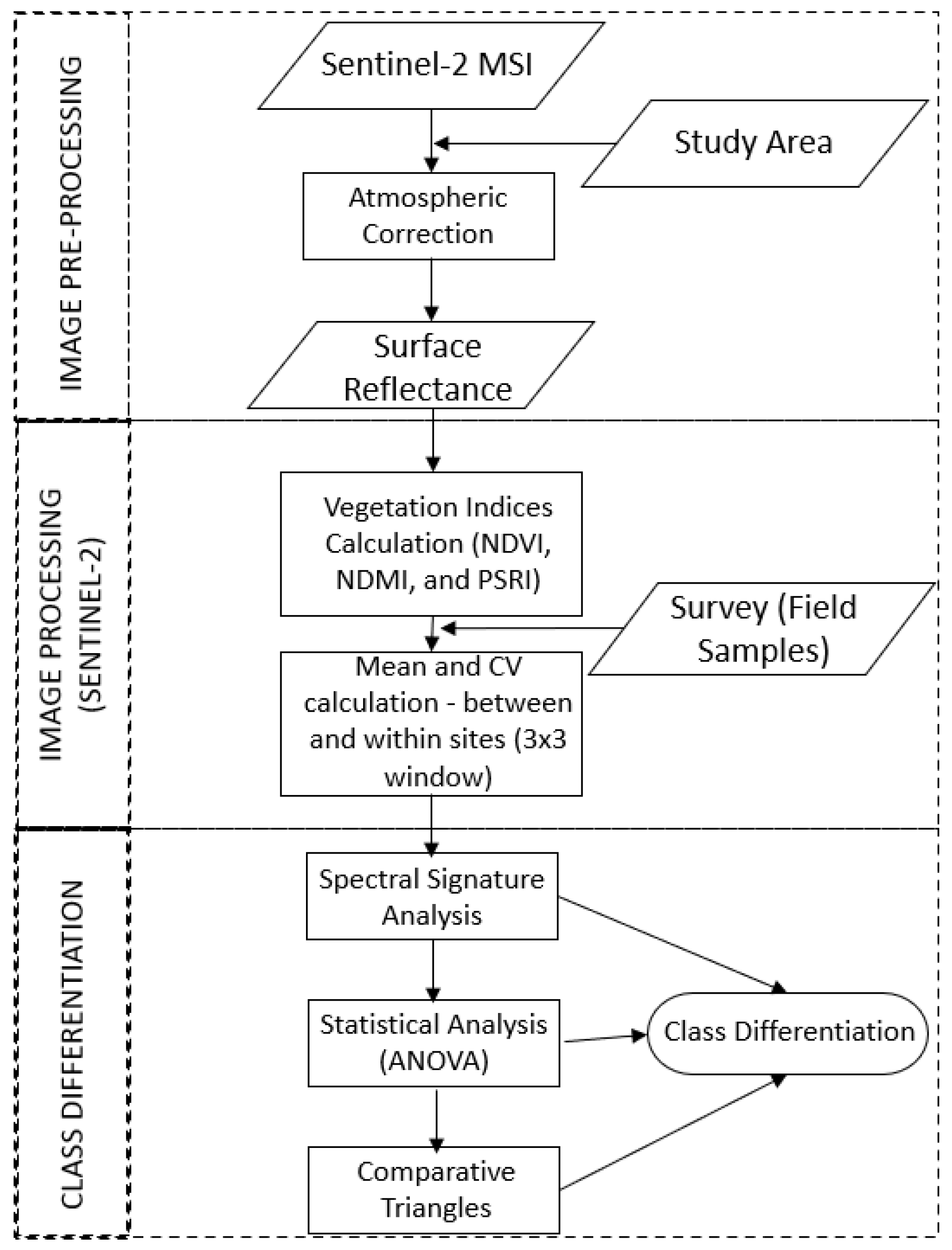
2.5. General Workflow
3. Results
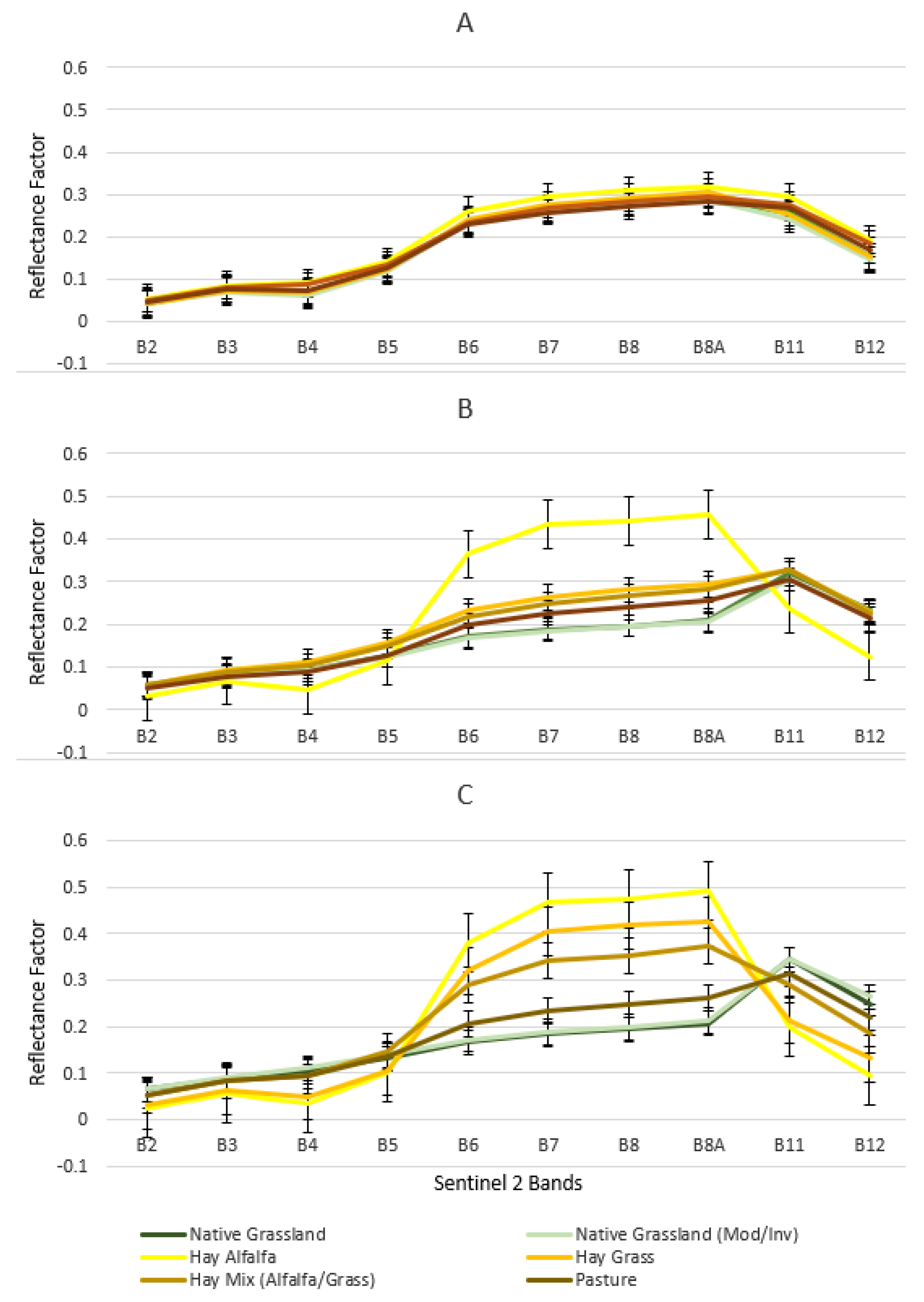
3.1. Spectral Signature of Six Grassland/Forage Land Covers
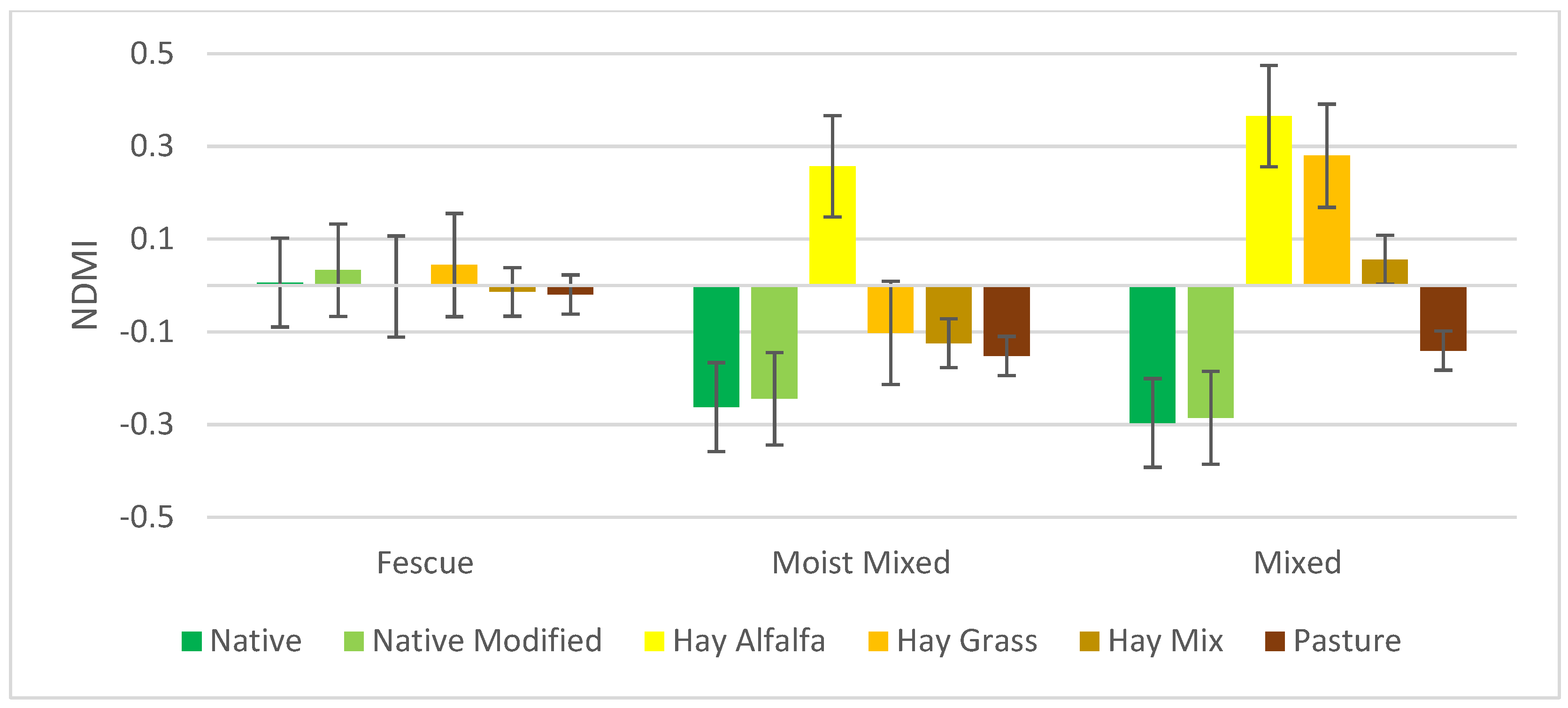
3.2. NDVI, NDMI, and PSRI
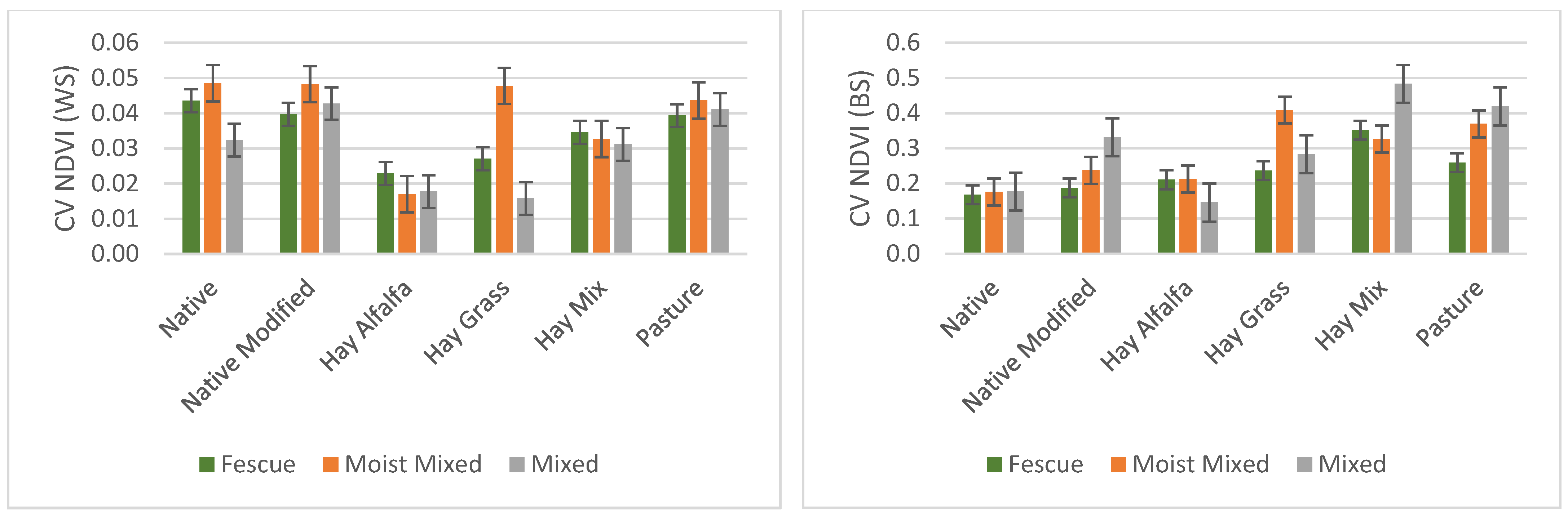
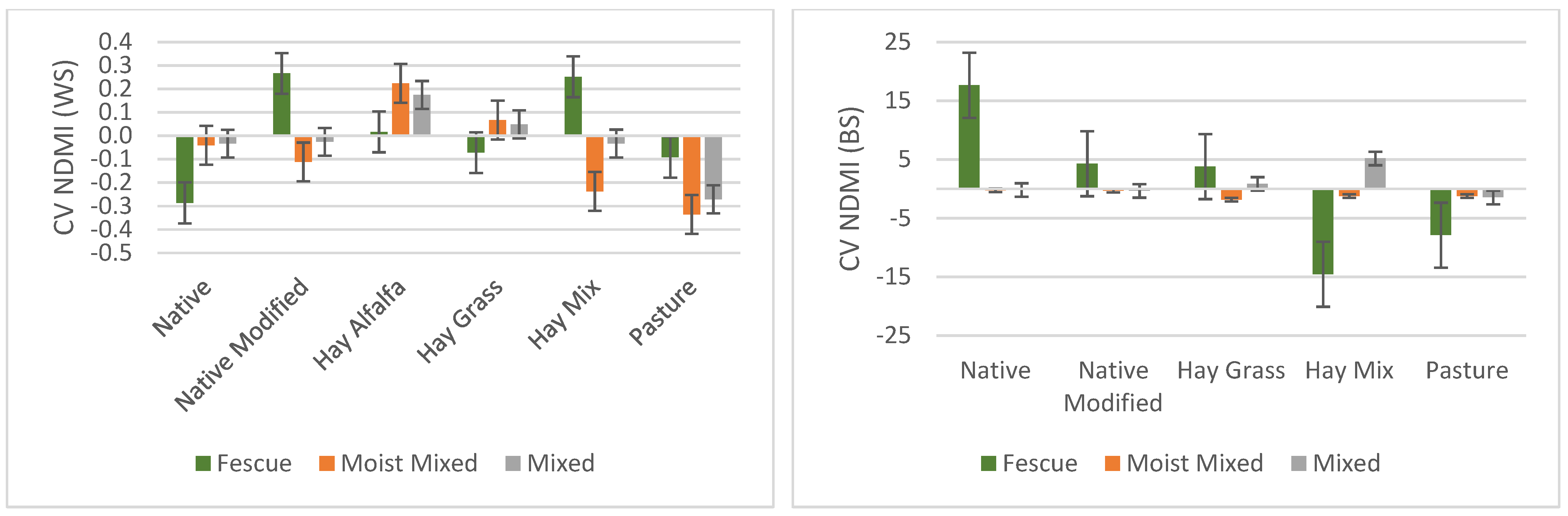
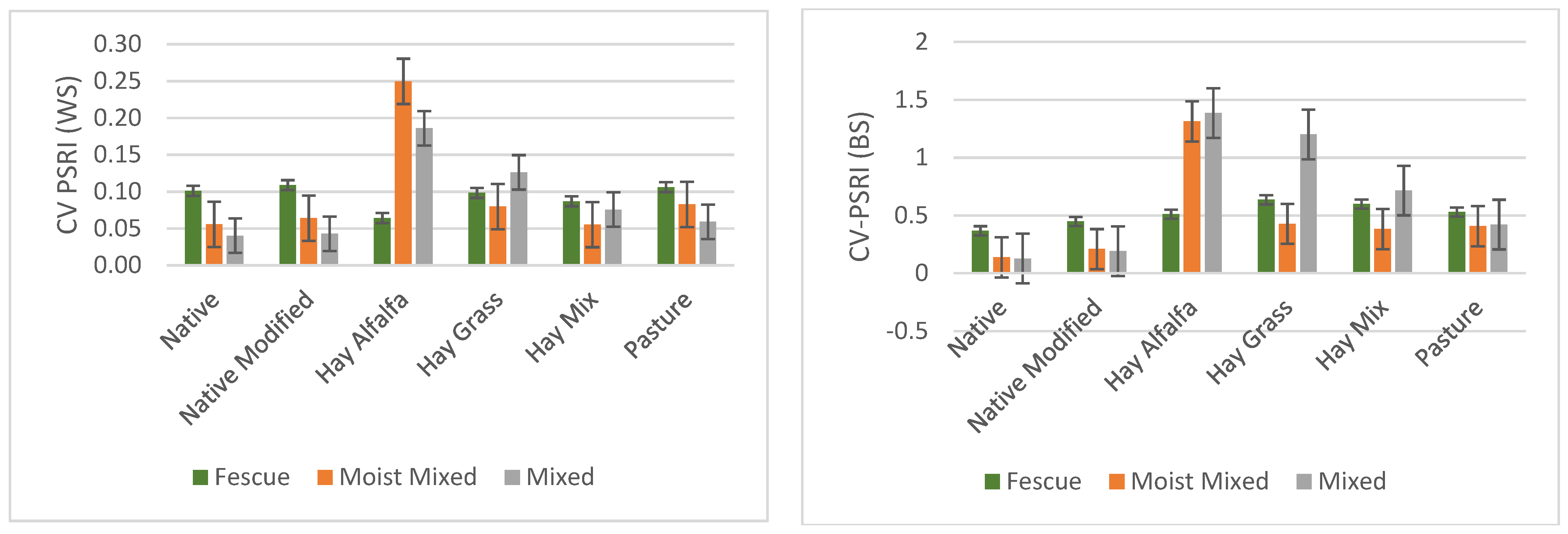
3.3. Variation within and between Sites for Different Grassland/Forage Land Covers
3.4. Separating Classes Using Comparative Triangles
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Conant, R.T.; Cerri, C.E.P.; Osborne, B.B.; Paustian, K. Grassland Management Impacts on Soil Carbon Stocks: A New Synthesis. Ecol. Appl. 2017, 27, 662–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinehart, L. Pasture, Rangeland and Grazing Management. National Sustainable Agricultural Information Service; ATTRA Publication #IP306: Butte, MT, USA, 2006. [Google Scholar]
- O’Mara, F.P. The Role of Grasslands in Food Security and Climate Change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filho, M.G.; Kuplich, T.M.; De Quadros, F.L.F. Estimating Natural Grassland Biomass by Vegetation Indices Using Sentinel 2 Remote Sensing Data. Int. J. Remote Sens. 2020, 41, 2861–2876. [Google Scholar] [CrossRef]
- Jin, Y.; Yang, X.; Qiu, J.; Li, J.; Gao, T.; Wu, Q.; Zhao, F.; Ma, H.; Yu, H.; Xu, B. Remote Sensing-Based Biomass Estimation and Its Spatio-Temporal Variations in Temperate Grassland, Northern China. Remote Sens. 2014, 6, 1496–1513. [Google Scholar] [CrossRef] [Green Version]
- Marsett, R.C.; Qi, J.; Heilman, P.; Biedenbender, S.H.; Carolyn Watson, M.; Amer, S.; Weltz, M.; Goodrich, D.; Marsett, R. Remote Sensing for Grassland Management in the Arid Southwest. Rangel. Ecol. Manag. 2006, 59, 530–540. [Google Scholar] [CrossRef]
- Zha, Y.; Gao, J.; Ni, S.; Liu, Y.; Jiang, J.; Wei, Y. A Spectral Reflectance-Based Approach to Quantification of Grassland Cover from Landsat TM Imagery. Remote Sens. Environ. 2003, 87, 371–375. [Google Scholar] [CrossRef]
- Aragón, R.; Oesterheld, M. Linking Vegetation Heterogeneity and Functional Attributes of Temperate Grasslands through Remote Sensing. Appl. Veg. Sci. 2008, 11, 117–130. [Google Scholar] [CrossRef]
- Cui, T.; Martz, L.; Guo, X. Grassland Phenology Response to Drought in the Canadian Prairies. Remote Sens. 2017, 9, 1258. [Google Scholar] [CrossRef] [Green Version]
- Fontana, F.; Rixen, C.; Jonas, T.; Aberegg, G.; Wunderle, S. Alpine Grassland Phenology as Seen in AVHRR, VEGETATION, and MODIS NDVI Time Series—A Comparison with In Situ Measurements. Sensors 2008, 8, 2833–2853. [Google Scholar] [CrossRef] [Green Version]
- Ali, I.; Cawkwell, F.; Dwyer, E.; Barrett, B.; Green, S. Satellite Remote Sensing of Grasslands: From Observation to Management. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Guo, X. Some Insights on Grassland Health Assessment Based on Remote Sensing. Sensors 2015, 15, 3070–3089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Yang, H.; Huang, L.; Chen, C.; Lin, X.; Hu, Z.; Li, J. Grassland Degradation Remote Sensing Monitoring and Driving Factors Quantitative Assessment in China from 1982 to 2010. Ecol. Indic. 2017, 83, 303–313. [Google Scholar] [CrossRef]
- Cleland, E.E.; Collins, S.L.; Dickson, T.L.; Farrer, E.C.; Gross, K.L.; Gherardi, L.A.; Hallett, L.M.; Hobbs, R.J.; Hsu, J.S.; Turnbull, L.; et al. Sensitivity of Grassland Plant Community Composition to Spatial vs. Temporal Variation in Precipitation. Ecology 2013, 94, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, A.P.; Faber-Langendoen, D.; Josse, C.; Morrison, J.; Loucks, C.J.; Dixon, A. Distribution Mapping of World Grassland Types. Wiley Online Libr. 2014, 41, 2003–2019. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ma, W.; Wang, W. Relationship between Variability in Aboveground Net Primary Production and Precipitation in Global Grasslands. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Fisette, T.; Davidson, A.; Daneshfar, B.; Rollin, P.; Aly, Z.; Campbell, L. Annual Space-Based Crop Inventory for Canada: 2009–2014. In Proceedings of the International Geoscience and Remote Sensing Symposium (IGARSS), Quebec City, QC, Canada, 13–18 July 2014; pp. 5095–5098. [Google Scholar]
- Pogue, S.J.; Kröbel, R.; Janzen, H.H.; Beauchemin, K.A.; Legesse, G.; de Souza, D.M.; Iravani, M.; Selin, C.; Byrne, J.; McAllister, T.A. Beef Production and Ecosystem Services in Canada’s Prairie Provinces: A Review. Agric. Syst. 2018, 166, 152–172. [Google Scholar] [CrossRef]
- Smith, A.M.; Buckley, J.R. Investigating RADARSAT-2 as a Tool for Monitoring Grassland in Western Canada. Can. J. Remote Sens. 2011, 37, 93–102. [Google Scholar] [CrossRef]
- Fisher, R.J.; Sawa, B.; Prieto, B. A Novel Technique Using LiDAR to Identify Native-Dominated and Tame-Dominated Grasslands in Canada. Remote Sens. Environ. 2018, 218, 201–206. [Google Scholar] [CrossRef]
- Yang, X.; Smith, A.M.; Hill, M.J. Updating the Grassland Vegetation Inventory Using Change Vector Analysis and Functionally-Based Vegetation Indices. Can. J. Remote Sens. 2017, 43, 62–78. [Google Scholar] [CrossRef]
- McInnes, W.S.; Smith, B.; McDermid, G.J. Discriminating Native and Nonnative Grasses in the Dry Mixedgrass Prairie with MODIS NDVI Time Series. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 1395–1403. [Google Scholar] [CrossRef]
- Lindsay, E.J.; King, D.J.; Davidson, A.M.; Daneshfar, B. Canadian Prairie Rangeland and Seeded Forage Classification Using Multiseason Landsat 8 and Summer RADARSAT-2. Rangel. Ecol. Manag. 2019, 72, 92–102. [Google Scholar] [CrossRef]
- Olimb, S.K.; Dixon, A.P.; Dolfi, E.; Engstrom, R.; Anderson, K. Prairie or Planted? Using Time-Series NDVI to Determine Grassland Characteristics in Montana. GeoJournal 2018, 83, 819–834. [Google Scholar] [CrossRef]
- Shorthouse, J.D. Ecoregions of Canada’s Prairie Grasslands. Arthropods Can. Grassl. 2010, 1, 53–81. [Google Scholar] [CrossRef]
- Bailey, A.W.; Schellenberg, M.P.; McCartney, D. Management of Canadian Prairie Rangeland; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2010.
- Gitelson, A.A.; Merzlyak, M.N.; Zur, Y. Non-Destructive and Remote Sensing Techniques for Estimation of Vegetation Status. In Proceedings of the 3rd European Conference on Precision Agriculture, Montpelier, France, 18–20 June 2001; University of Nebraska: Lincoln, NE, USA, 2001. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the Radiometric and Biophysical Performance of the MODIS Vegetation Indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Tucker, C.J.; Elgin, J.H.; McMurtrey, J.E.; Fan, C.J. Monitoring Corn and Soybean Crop Development with Hand-Held Radiometer Spectral Data. Remote Sens. Environ. 1979, 8, 237–248. [Google Scholar] [CrossRef]
- Galvao, L.; Formaggio, A.; Environment, D.T.-R. Discrimination of Sugarcane Varieties in Southeastern Brazil with EO-1 Hyperion Data. Remote Sens. Environ. 2005, 94, 523–534. [Google Scholar] [CrossRef]
- Wilson, E.H.; Sader, S.A. Detection of Forest Harvest Type Using Multiple Dates of Landsat TM Imagery. Remote Sens. Environ. 2002, 80, 385–396. [Google Scholar] [CrossRef]
- Anderegg, J.; Yu, K.; Aasen, H.; Walter, A.; Liebisch, F.; Hund, A. Spectral Vegetation Indices to Track Senescence Dynamics in Diverse Wheat Germplasm. Front. Plant Sci. 2020, 10, 1749. [Google Scholar] [CrossRef] [Green Version]
- Apan, A.; Held, A.; Phinn, S.; Markley, J. Formulation and Assessment of Narrow-Band Vegetation Indices from EO-1 Hyperion Imagery for Discriminating Sugarcane Disease. In Proceedings of the Spatial Sciences Institute Biennial Conference (SSC 2003): Spatial Knowledge without Boundaries, Canberra, Australia, 22–26 September 2003; pp. 1–13. [Google Scholar]
- Sims, D.A.; Gamon, J.A. Relationships between Leaf Pigment Content and Spectral Reflectance across a Wide Range of Species, Leaf Structures and Developmental Stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Hall-Beyer, M. Patterns in the Yearly Trajectory of Standard Deviation of NDVI over 25 Years for Forest, Grasslands and Croplands across Ecological Gradients in Alberta, Canada. Int. J. Remote Sens. 2012, 33, 2725–2746. [Google Scholar] [CrossRef]
- Environment Alberta. 2016 Range Reference Area Report for the Southern Grasslands; Environment Alberta: Edmonton, AB, Canada, 2017.
- Li, Q.; Wang, C.; Zhang, B.; Lu, L. Object-Based Crop Classification with Landsat-MODIS Enhanced Time-Series Data. Remote Sens. 2015, 7, 16091–16107. [Google Scholar] [CrossRef] [Green Version]
- Griffith, J.A.; Price, K.P.; Martinko, E.A. A Multivariate Analysis of Biophysical Parameters of Tallgrass Prairie Among Land Management Practices and Years. Environ. Monit. Assess. 2001, 68, 249–271. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, F.; Schneider, M.K.; Keller, A.; Mayr, A.; Rentschler, T.; Meuli, R.G.; Schaepman, M.; Liebisch, F. Spatial Monitoring of Grassland Management Using Multi-Temporal Satellite Imagery. Ecol. Indic. 2020, 113, 106201. [Google Scholar] [CrossRef]
- Gómez Giménez, M.; de Jong, R.; Della Peruta, R.; Keller, A.; Schaepman, M.E. Determination of Grassland Use Intensity Based on Multi-Temporal Remote Sensing Data and Ecological Indicators. Remote Sens. Environ. 2017, 198, 126–139. [Google Scholar] [CrossRef]
- Peeters, A.; Beaufoy, G.; Canals, R.M.; De Vliegher, A.; Huyghe, C.; Isselstein, J.; Jones, J.; Kessler, W.; Kirilovsky, D.; Van Den Pol-Van Dasselaar, A. Grassland Term Definitions and Classifications Adapted to the Diversity of European Grassland-Based Systems. In Proceedings of the 25th EGF General Meeting on “EGF at 50: The Future of European Grasslands”, Aberystwyth, UK, 7–11 September 2014; pp. 743–750. [Google Scholar]
- Kizeková, M.; Hopkins, A.; Kanianska, R.; Makovníková, J.; Pollák, Š.; Pálka, B. Changes in the Area of Permanent Grassland and Its Implications for the Provision of Bioenergy: Slovakia as a Case Study. Grass Forage Sci. 2018, 73, 218–232. [Google Scholar] [CrossRef] [Green Version]
- Porqueddu, C.; Ates, S.; Louhaichi, M.; Kyriazopoulos, A.P.; Moreno, G.; del Pozo, A.; Ovalle, C.; Ewing, M.A.; Nichols, P.G.H. Grasslands in ‘Old World’ and ‘New World’ Mediterranean-Climate Zones: Past Trends, Current Status and Future Research Priorities. Grass Forage Sci. 2016, 71, 1–35. [Google Scholar] [CrossRef]
- Hudewenz, A.; Klein, A.-M.; Scherber, C.; Stanke, L.; Tscharntke, T.; Vogel, A.; Weigelt, A.; Weisser, W.W.; Ebeling, A. Herbivore and Pollinator Responses to Grassland Management Intensity along Experimental Changes in Plant Species Richness. Biol. Conserv. 2012, 150, 42–52. [Google Scholar] [CrossRef]
- Rose, L.; Leuschner, C. The Diversity–Productivity Relationship in a Permanent Temperate Grassland: Negative Diversity Effect, Dominant Influence of Management Regime. Plant Ecol. Divers. 2012, 5, 265–274. [Google Scholar] [CrossRef]
- Wang, C.; Jamison, B.E.; Spicci, A.A. Trajectory-Based Warm Season Grassland Mapping in Missouri Prairies with Multi-Temporal ASTER Imagery. Remote Sens. Environ. 2010, 114, 531–539. [Google Scholar] [CrossRef]
- Gómez, C.; White, J.C.; Wulder, M.A. Optical Remotely Sensed Time Series Data for Land Cover Classification: A Review. ISPRS J. Photogramm. Remote Sens. 2016, 116, 55–72. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class Level-1 | Class Level-2 | Code * | Samples Fescue | Samples Moist-Mixed | Samples Mixed | Description |
|---|---|---|---|---|---|---|
| Native grassland | Native | 101 | 77 | 93 | 269 | Land dominated by native grasses and managed as a natural ecosystem |
| Modified/Invaded | 105 | 25 | 21 | 69 | Land dominated by introduced or invasive species and managed as a natural ecosystem | |
| Seeded forage | Hay Alfalfa | 201 | 7 | 27 | 37 | Land devoted to hay production (mainly alfalfa) |
| Hay Grass | 203 | 20 | 27 | 9 | Land devoted to hay production (mainly grass) | |
| Hay Alfalfa/Grass | 204 | 25 | 30 | 15 | Land devoted to hay production (mixture of alfalfa and grass) | |
| Pasture | 210 | 51 | 53 | 50 | Lands isolated by fences, which receive periodic cultural treatments, composed of forage plants used to feed grazing animals |
| NDVI Mean [Standard Deviation] | |||
|---|---|---|---|
| Grassland Types | Fescue | Moist-Mixed | Mixed |
| Native | 0.61[0.025]a * | 0.36[0.019]b # | 0.31[0.010]d ^ |
| Native Modified | 0.63[0.023]a * | 0.36[0.019]bc # | 0.28[0.012]d ^ |
| Hay Alfalfa | 0.56[0.012]a * | 0.78[0.012]a # | 0.82[0.013]a # |
| Hay Grass | 0.62[0.016]a * | 0.43[0.019]bc # | 0.74[0.011]a * |
| Hay Mixture | 0.54[0.017]a * | 0.45[0.014]c * | 0.54[0.012]b * |
| Pasture | 0.58[0.022]a * | 0.45[0.019]c # | 0.43[0.016]c # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frank, T.; Smith, A.; Houston, B.; Lindsay, E.; Guo, X. Differentiation of Six Grassland/Forage Types in Three Canadian Ecoregions Based on Spectral Characteristics. Remote Sens. 2022, 14, 2121. https://doi.org/10.3390/rs14092121
Frank T, Smith A, Houston B, Lindsay E, Guo X. Differentiation of Six Grassland/Forage Types in Three Canadian Ecoregions Based on Spectral Characteristics. Remote Sensing. 2022; 14(9):2121. https://doi.org/10.3390/rs14092121
Chicago/Turabian StyleFrank, Thiago, Anne Smith, Bill Houston, Emily Lindsay, and Xulin Guo. 2022. "Differentiation of Six Grassland/Forage Types in Three Canadian Ecoregions Based on Spectral Characteristics" Remote Sensing 14, no. 9: 2121. https://doi.org/10.3390/rs14092121
APA StyleFrank, T., Smith, A., Houston, B., Lindsay, E., & Guo, X. (2022). Differentiation of Six Grassland/Forage Types in Three Canadian Ecoregions Based on Spectral Characteristics. Remote Sensing, 14(9), 2121. https://doi.org/10.3390/rs14092121