
Establishing the Position and Drivers of the Eastern Andean Treeline with Automated Transect Sampling


Abstract
:1. Introduction
2. Materials and Methods
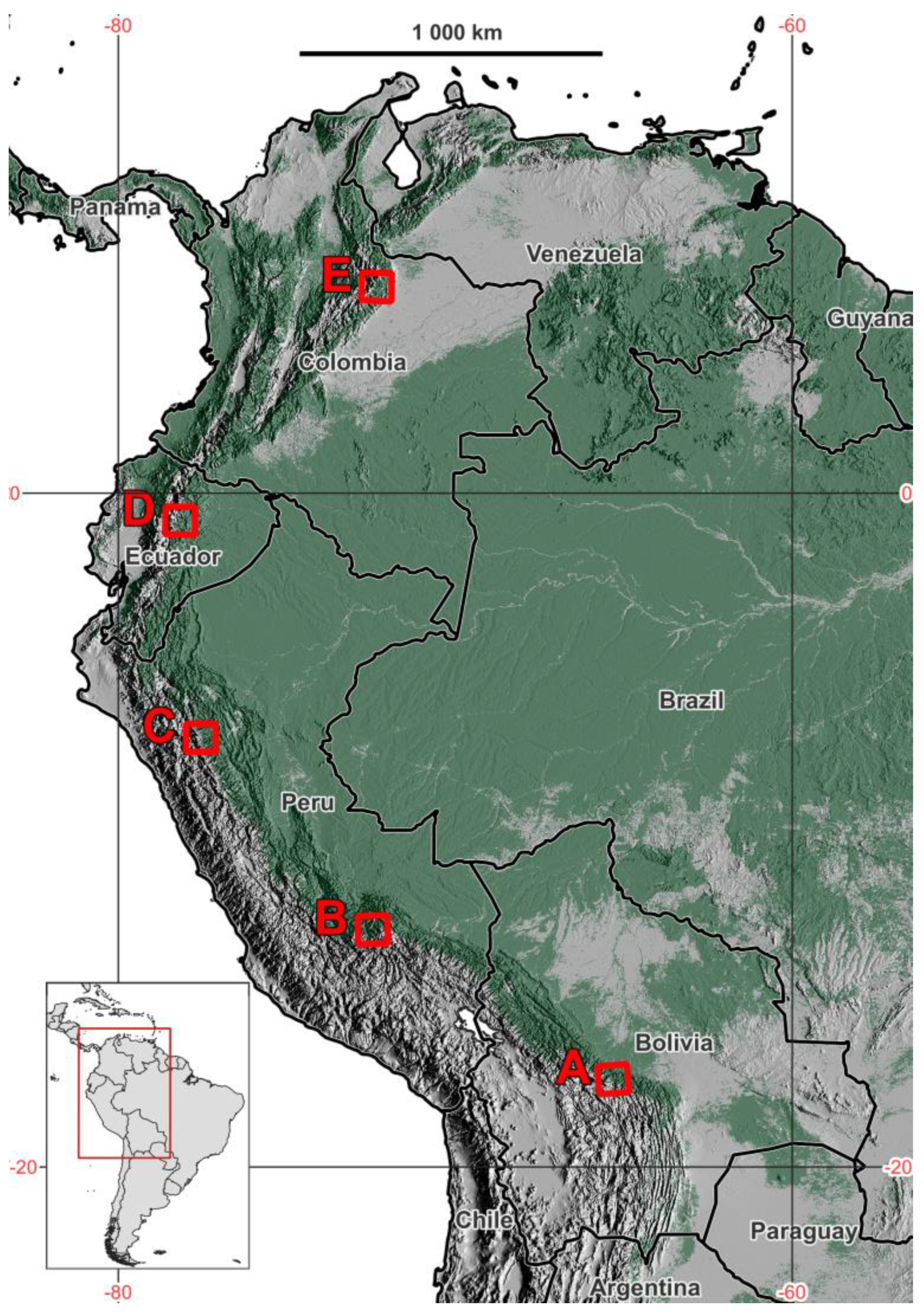
2.1. Study Area
2.2. Materials
2.3. Methods
3. Results
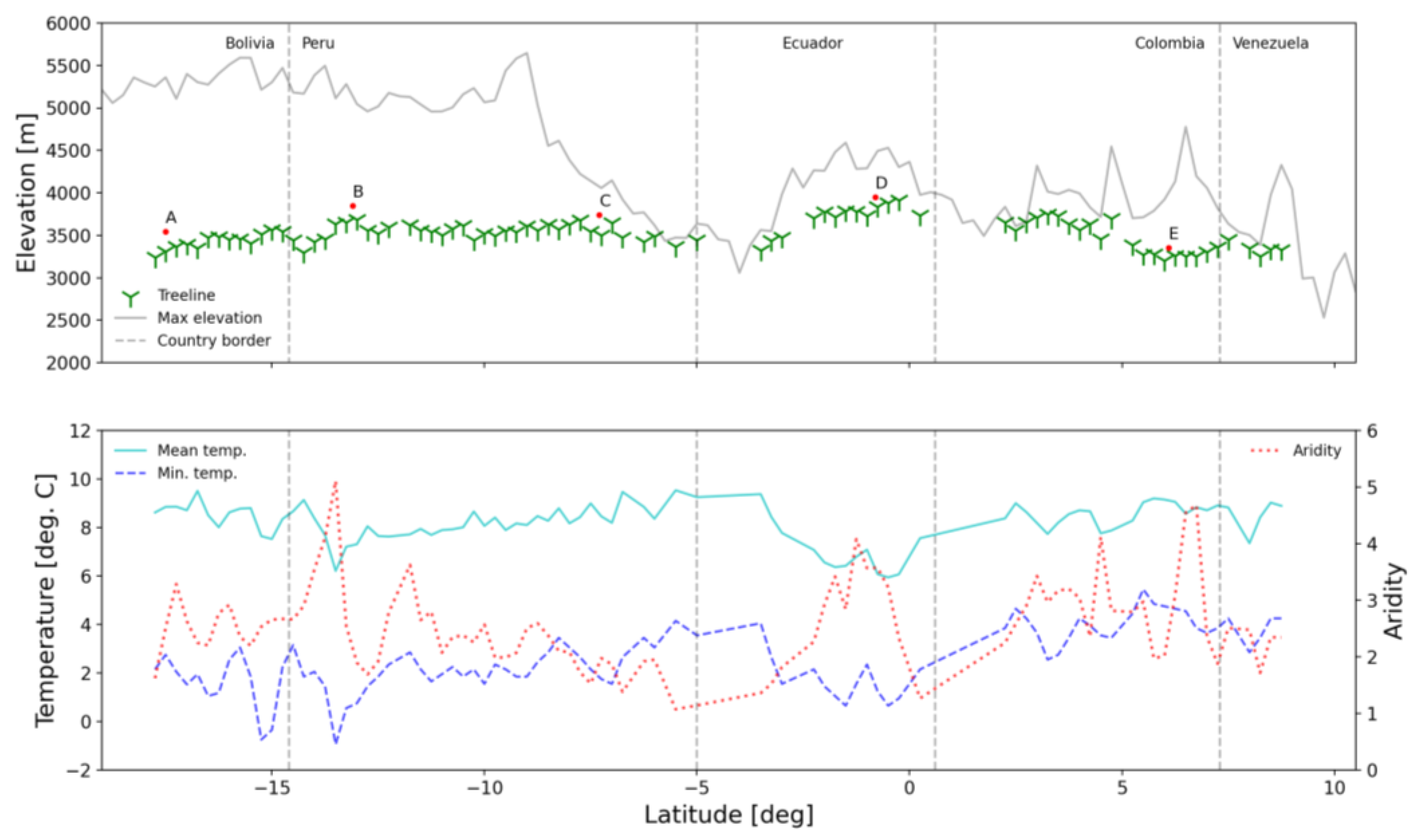
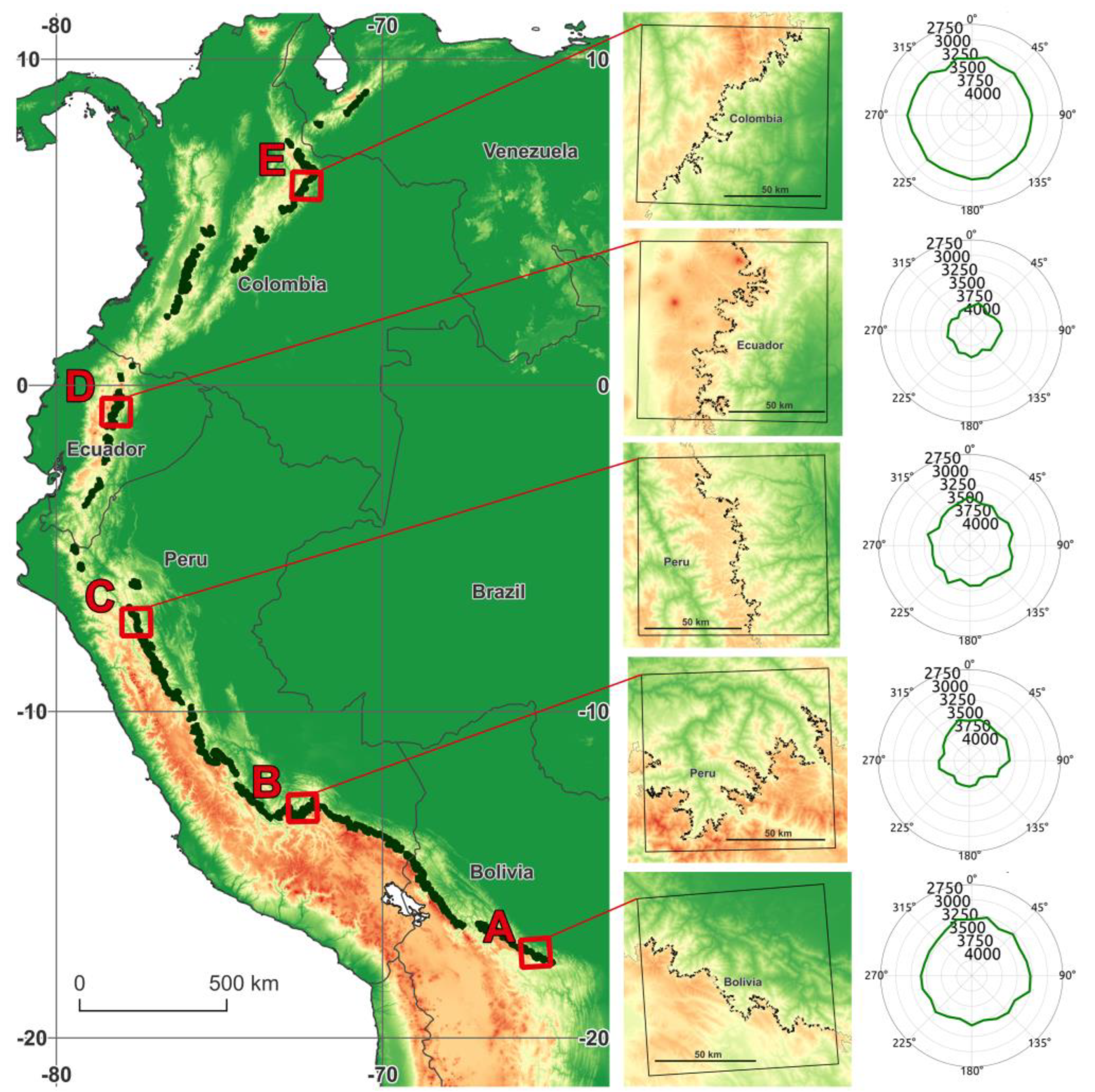
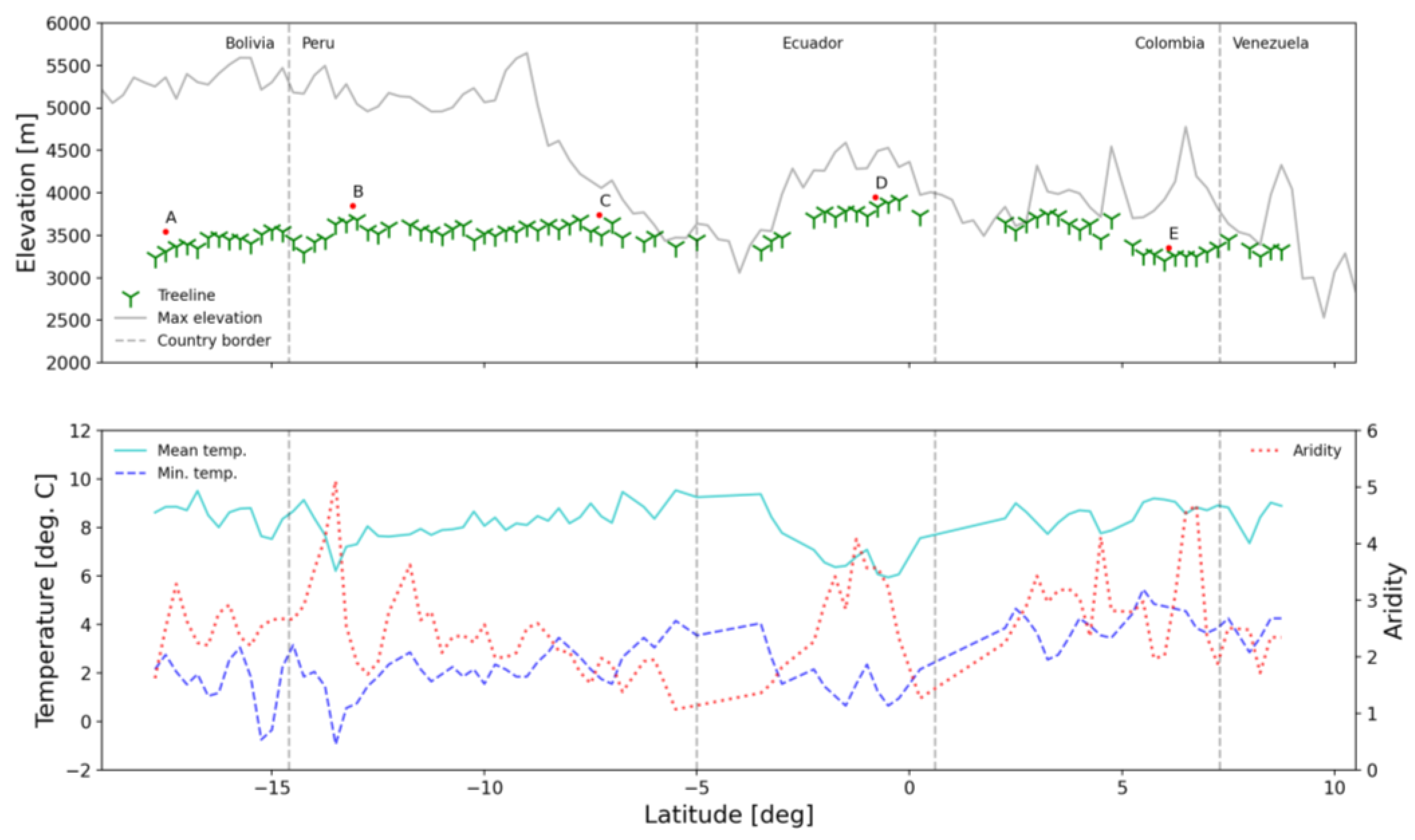
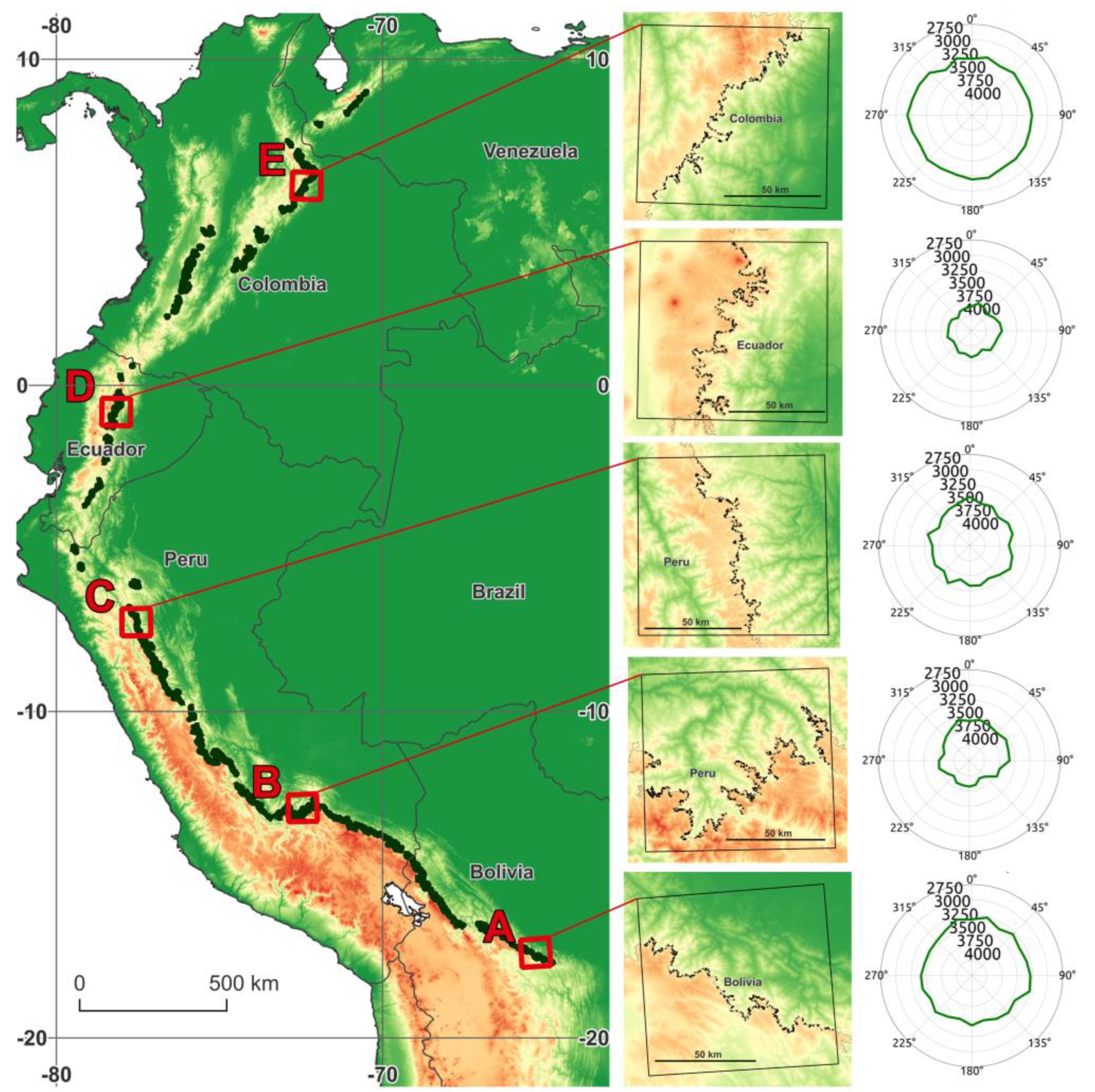
3.1. Contemporary Treeline Extent and Environmental Controls
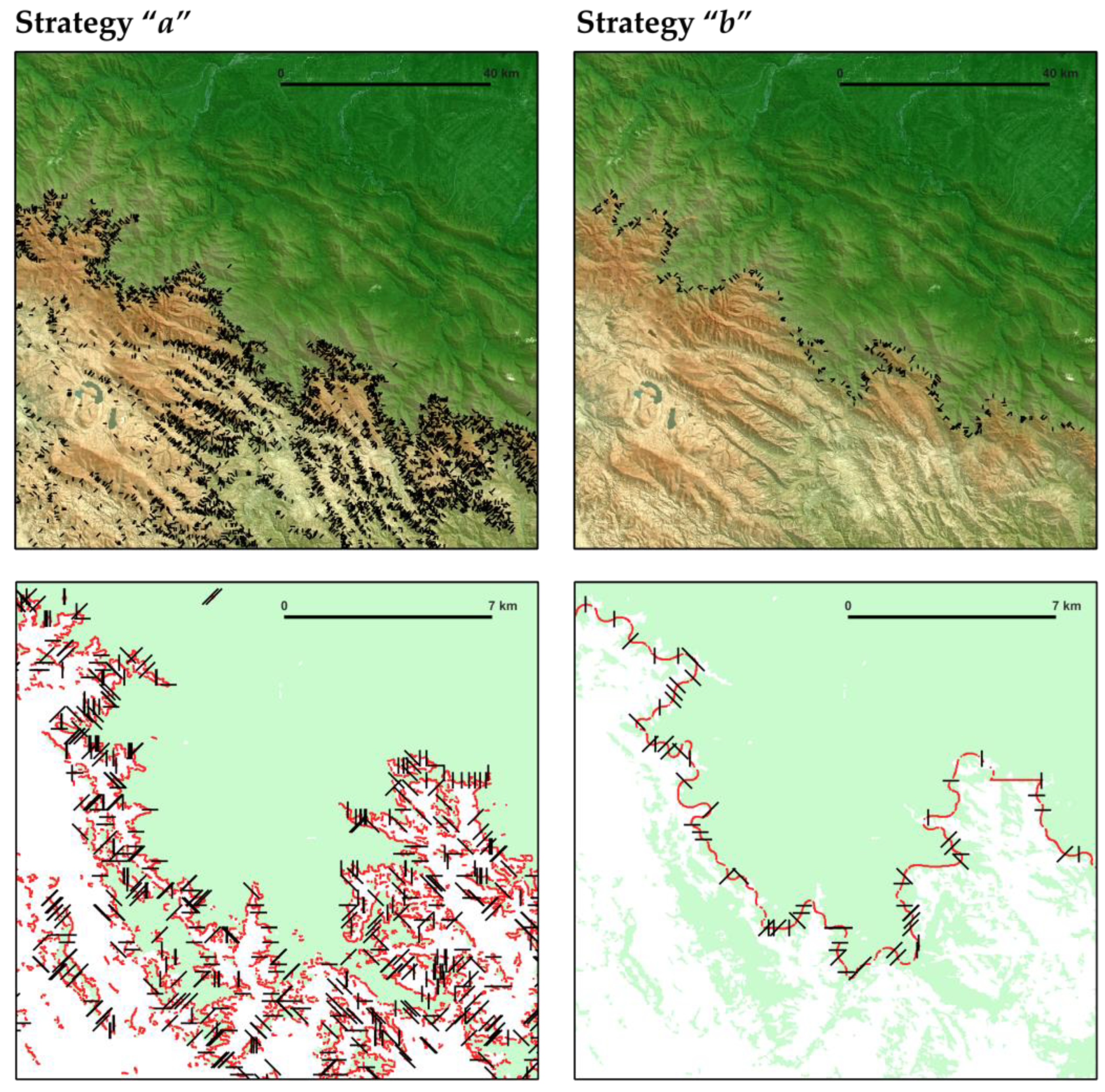
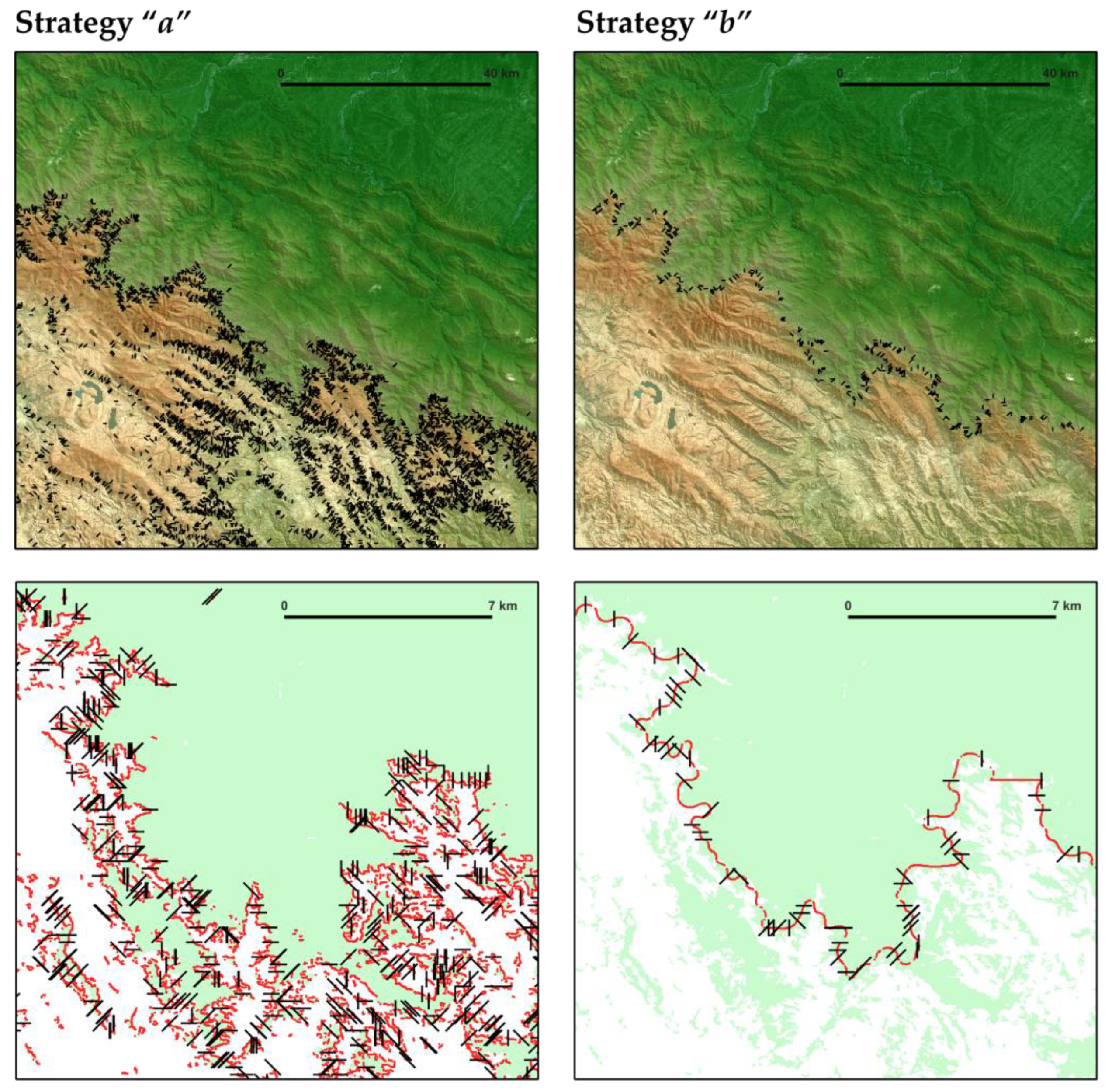
3.2. Assessment of Strategies for Automated Sampling with Transects
3.3. Insight from Elevational Transects
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Pabón-Caicedo, J.D.; Arias, P.A.; Carril, A.F.; Espinoza, J.C.; Borrel, L.F.; Goubanova, K.; Lavado-Casimiro, W.; Masiokas, M.; Solman, S.; Villalba, R. Observed and Projected Hydroclimate Changes in the Andes. Front. Earth Sci. 2020, 8, 61. [Google Scholar] [CrossRef]
- Vuille, M.; Francou, B.; Wagnon, P.; Juen, I.; Kaser, G.; Mark, B.G.; Bradley, R.S. Climate Change and Tropical Andean Glaciers: Past, Present and Future. Earth-Sci. Rev. 2008, 89, 79–96. [Google Scholar] [CrossRef]
- Morueta-Holme, N.; Engemann, K.; Sandoval-Acuña, P.; Jonas, J.D.; Segnitz, R.M.; Svenning, J.-C. Strong Upslope Shifts in Chimborazo’s Vegetation over Two Centuries since Humboldt. Proc. Natl. Acad. Sci. USA 2015, 112, 12741–12745. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global Warming, Elevational Range Shifts, and Lowland Biotic Attrition in the Wet Tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Meir, P.; Revilla, N.S.; Quisiyupanqui, M.N.R.; Saatchi, S. Upslope Migration of Andean Trees. J. Biogeogr. 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are Treelines Advancing? A Global Meta-Analysis of Treeline Response to Climate Warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Helmer, E.H.; Gerson, E.A.; Baggett, L.S.; Bird, B.J.; Ruzycki, T.S.; Voggesser, S.M. Neotropical Cloud Forests and Páramo to Contract and Dry from Declines in Cloud Immersion and Frost. PLoS ONE 2019, 14, e0213155. [Google Scholar] [CrossRef]
- Paulsen, J.; Körner, C. A Climate-Based Model to Predict Potential Treeline Position around the Globe. Alp. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef]
- Körner, C. The Cold Range Limit of Trees. Trends Ecol. Evol. 2021, 36, 979–989. [Google Scholar] [CrossRef]
- Kessler, M.; Böhner, J.; Kluge, J. Modelling Tree Height to Assess Climatic Conditions at Tree Lines in the Bolivian Andes. Ecol. Model. 2007, 207, 223–233. [Google Scholar] [CrossRef]
- Sarmiento, G.; Pinillos, M. The Tropical Alpine Treeline: A Case Study of a Changing Ecosystem Boundary. In Applying Ecological Knowledge to Land use Decisions; Inter-American Institute for Global Change Research: Montevideo, Uruguay, 2008; pp. 111–122. [Google Scholar]
- Sarmiento, F.O.; Frolich, L.M. Andean Cloud Forest Tree Lines. Mt. Res. Dev. 2002, 22, 278–287. [Google Scholar] [CrossRef]
- Lutz, D.A.; Powell, R.L.; Silman, M.R. Four Decades of Andean Timberline Migration and Implications for Biodiversity Loss with Climate Change. PLoS ONE 2013, 8, e74496. [Google Scholar] [CrossRef] [PubMed]
- Young, K.R.; Ponette-González, A.G.; Polk, M.H.; Lipton, J.K. Snowlines and Treelines in the Tropical Andes. Ann. Am. Assoc. Geogr. 2017, 107, 429–440. [Google Scholar] [CrossRef]
- Chacón-Moreno, E.; Rodríguez-Morales, M.; Paredes, D.; Suárez del Moral, P.; Albarrán, A. Impacts of Global Change on the Spatial Dynamics of Treeline in Venezuelan Andes. Front. Ecol. Evol. 2021, 9, 615223. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G.E. Treeline Advance—Driving Processes and Adverse Factors. Landsc. Online 2007, 1, 1–33. [Google Scholar] [CrossRef]
- Rita, A.; Bonanomi, G.; Allevato, E.; Borghetti, M.; Cesarano, G.; Mogavero, V.; Rossi, S.; Saulino, L.; Zotti, M.; Saracino, A. Topography Modulates Near-Ground Microclimate in the Mediterranean Fagus Sylvatica Treeline. Sci. Rep. 2021, 11, 8122. [Google Scholar] [CrossRef]
- Quadri, P.; Silva, L.C.R.; Zavaleta, E.S. Climate-Induced Reversal of Tree Growth Patterns at a Tropical Treeline. Sci. Adv. 2021, 7, eabb7572. [Google Scholar] [CrossRef]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraer, M.; Caceres, E.B.; Forsythe, N.; Fowler, H.; Greenwood, G.; Hashmi, M.Z.; Liu, X.D.; et al. Elevation-Dependent Warming in Mountain Regions of the World. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar] [CrossRef]
- Tovar, C.; Carril, A.F.; Gutiérrez, A.G.; Ahrends, A.; Fita, L.; Zaninelli, P.; Flombaum, P.; Abarzúa, A.M.; Alarcón, D.; Aschero, V.; et al. Understanding Climate Change Impacts on Biome and Plant Distributions in the Andes: Challenges and Opportunities. J. Biogeogr. 2022, 49, 1420–1442. [Google Scholar] [CrossRef]
- Storey, J.C.; Rengarajan, R.; Choate, M.J. Bundle Adjustment Using Space-Based Triangulation Method for Improving the Landsat Global Ground Reference. Remote Sens. 2019, 11, 1640. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.C.; Kommareddy, I.; Kommareddy, A.; Turubanova, S.; Pickens, A.; Adusei, B.; Tyukavina, A.; Ying, Q. Landsat Analysis Ready Data for Global Land Cover and Land Cover Change Mapping. Remote Sens. 2020, 12, 426. [Google Scholar] [CrossRef]
- Young, K.R.; Leon, B. Distribution and Conservation of Peru’s Montane Forests: Interactions between the Biota and Human Society. In Tropical Montane Cloud Forests; Springer: Cham, Switzerland, 1993; pp. 237–246. [Google Scholar]
- Sulla-Menashe, D.; Gray, J.M.; Abercrombie, S.P.; Friedl, M.A. Hierarchical Mapping of Annual Global Land Cover 2001 to Present: The MODIS Collection 6 Land Cover Product. Remote Sens. Environ. 2019, 222, 183–194. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, 183. [Google Scholar] [CrossRef]
- Potapov, P.; Hansen, M.C.; Pickens, A.; Hernandez-Serna, A.; Tyukavina, A.; Turubanova, S.; Zalles, V.; Li, X.; Khan, A.; Stolle, F.; et al. The Global 2000–2020 Land Cover and Land Use Change Dataset Derived from the Landsat Archive: First Results. Front. Remote Sens. 2022, 3, 18. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at High Resolution for the Earth’s Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Zomer, R.J.; Xu, J.; Trabucco, A. Version 3 of the Global Aridity Index and Potential Evapotranspiration Database. Sci. Data 2022, 9, 409. [Google Scholar] [CrossRef]
- Bader, M.Y.; Llambí, L.D.; Case, B.S.; Buckley, H.L.; Toivonen, J.M.; Camarero, J.J.; Cairns, D.M.; Brown, C.D.; Wiegand, T.; Resler, L.M. A Global Framework for Linking Alpine-Treeline Ecotone Patterns to Underlying Processes. Ecography 2021, 44, 265–292. [Google Scholar] [CrossRef]
- Korner, C.; Paulsen, J. A World-Wide Study of High Altitude Treeline Temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Kok, K.V.; Kok, P.A.; Beukema, H. Effects of Cutting and Grazing on Andean Treeline Vegetation. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; New York Botanical Garden: New York, NY, USA, 1995. [Google Scholar]
- Sarmiento, F. Anthropogenic Change in the Landscapes of Highland Ecuador. Geogr. Rev. 2002, 92, 213–234. [Google Scholar] [CrossRef]
- Sexton, J.O.; Song, X.-P.; Feng, M.; Noojipady, P.; Anand, A.; Huang, C.; Kim, D.-H.; Collins, K.M.; Channan, S.; DiMiceli, C.; et al. Global, 30-m Resolution Continuous Fields of Tree Cover: Landsat-Based Rescaling of MODIS Vegetation Continuous Fields with Lidar-Based Estimates of Error. Int. J. Digit. Earth 2013, 6, 427–448. [Google Scholar] [CrossRef]
- Mukul, M.; Srivastava, V.; Mukul, M. Accuracy Analysis of the 2014–2015 Global Shuttle Radar Topography Mission (SRTM) 1 Arc-Sec C-Band Height Model Using International Global Navigation Satellite System Service (IGS) Network. J. Earth Syst. Sci. 2016, 125, 909–917. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Smith, T.; Bookhagen, B. Climatic and Biotic Controls on Topographic Asymmetry at the Global Scale. J. Geophys. Res. Earth Surf. 2021, 126, e2020JF005692. [Google Scholar] [CrossRef]
- Grace, J.; Berninger, F.; Nagy, L. Impacts of Climate Change on the Tree Line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef]
- Myneni, R.B.; Dong, J.; Tucker, C.J.; Kaufmann, R.K.; Kauppi, P.E.; Liski, J.; Zhou, L.; Alexeyev, V.; Hughes, M.K. A Large Carbon Sink in the Woody Biomass of Northern Forests. Proc. Natl. Acad. Sci. USA 2001, 98, 14784–14789. [Google Scholar] [CrossRef]
- Olthof, I.; Pouliot, D. Treeline Vegetation Composition and Change in Canada’s Western Subarctic from AVHRR and Canopy Reflectance Modeling. Remote Sens. Environ. 2010, 114, 805–815. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Focus Region | ||||
|---|---|---|---|---|---|
| A | B | C | D | E | |
| Coordinates (midpoint) | 65.3°W, 17.5°S | 72.4°W, 13.1°S | 77.5°W, 7.3°S | 78.2°W, 0.8°S | 72.4°W, 6.1°N |
| Country | Bolivia | Peru | Peru | Ecuador | Columbia |
| Overall orientation | N-E (28°) | N (327°) | E (78°) | E (108°) | S-E (129°) |
| Length [km] | 296 | 754 | 304 | 489 | 378 |
| Elevation; avg. ± SD [m] | 3466 ± 152 | 3766 ± 200 | 3656 ± 172 | 3878 ± 130 | 3291 ± 158 |
| Diff. from regional estimate [m] | +211 | +243 | +154 | +140 | +112 |
| SW-NE elev. diff. [m] | 127 | 201 | 81 | 50 | −63 |
| Annual mean temp. [°C] | 9.13 ± 0.78 | 7.86 ± 1.00 | 8.91 ± 0.74 | 6.75 ± 0.65 | 9.48 ± 0.67 |
| Annual min. temp. [°C] | 2.9 ± 0.92 | 2.07 ± 1.18 | 2.15 ± 0.69 | 1.8 ± 0.78 | 5.24 ± 0.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zelazowski, P.; Jozefowicz, S.; Feeley, K.J.; Malhi, Y. Establishing the Position and Drivers of the Eastern Andean Treeline with Automated Transect Sampling. Remote Sens. 2023, 15, 2679. https://doi.org/10.3390/rs15102679
Zelazowski P, Jozefowicz S, Feeley KJ, Malhi Y. Establishing the Position and Drivers of the Eastern Andean Treeline with Automated Transect Sampling. Remote Sensing. 2023; 15(10):2679. https://doi.org/10.3390/rs15102679
Chicago/Turabian StyleZelazowski, Przemyslaw, Stefan Jozefowicz, Kenneth J. Feeley, and Yadvinder Malhi. 2023. "Establishing the Position and Drivers of the Eastern Andean Treeline with Automated Transect Sampling" Remote Sensing 15, no. 10: 2679. https://doi.org/10.3390/rs15102679
APA StyleZelazowski, P., Jozefowicz, S., Feeley, K. J., & Malhi, Y. (2023). Establishing the Position and Drivers of the Eastern Andean Treeline with Automated Transect Sampling. Remote Sensing, 15(10), 2679. https://doi.org/10.3390/rs15102679