

Vegetation Types Variations to the South of Ngoring Lake from 2013 to 2020, Analyzed by Hyperspectral Imaging



Abstract
:1. Introduction
2. Materials and Methods
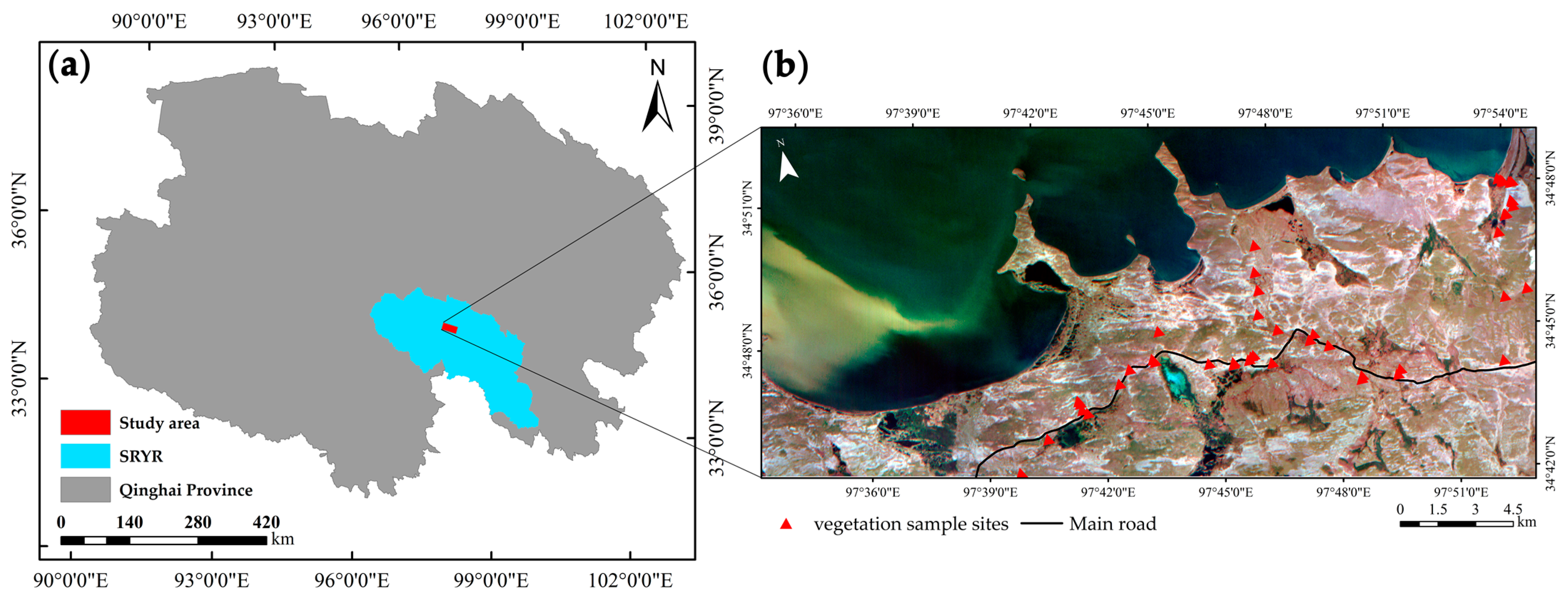
2.1. Study Area
2.2. Data and Preprocessing
2.3. Methods
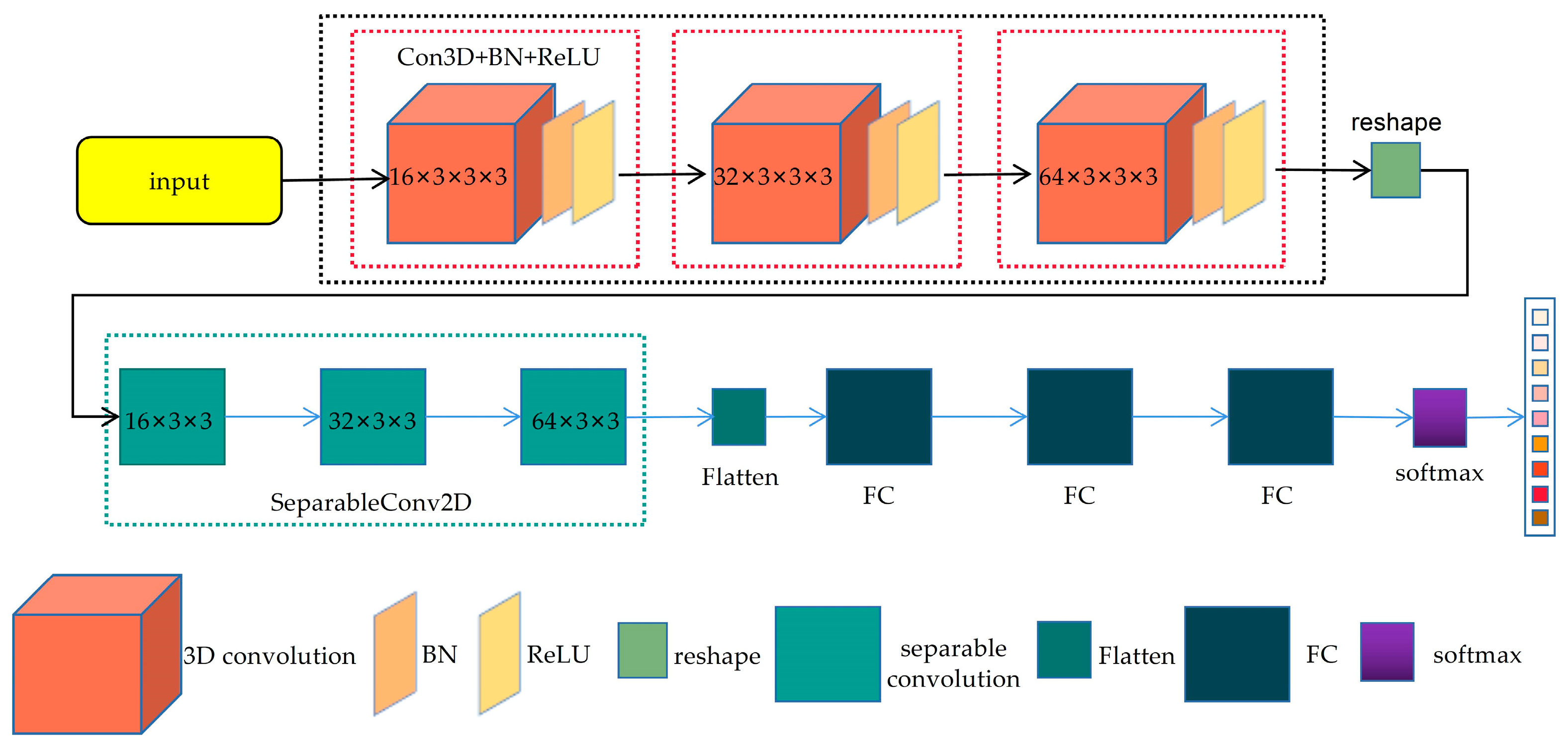
2.3.1. Hybrid Spectral Convolutional Neural Network (HybridSN)
2.3.2. Transition Matrix
2.3.3. Linear Regression Trend Model
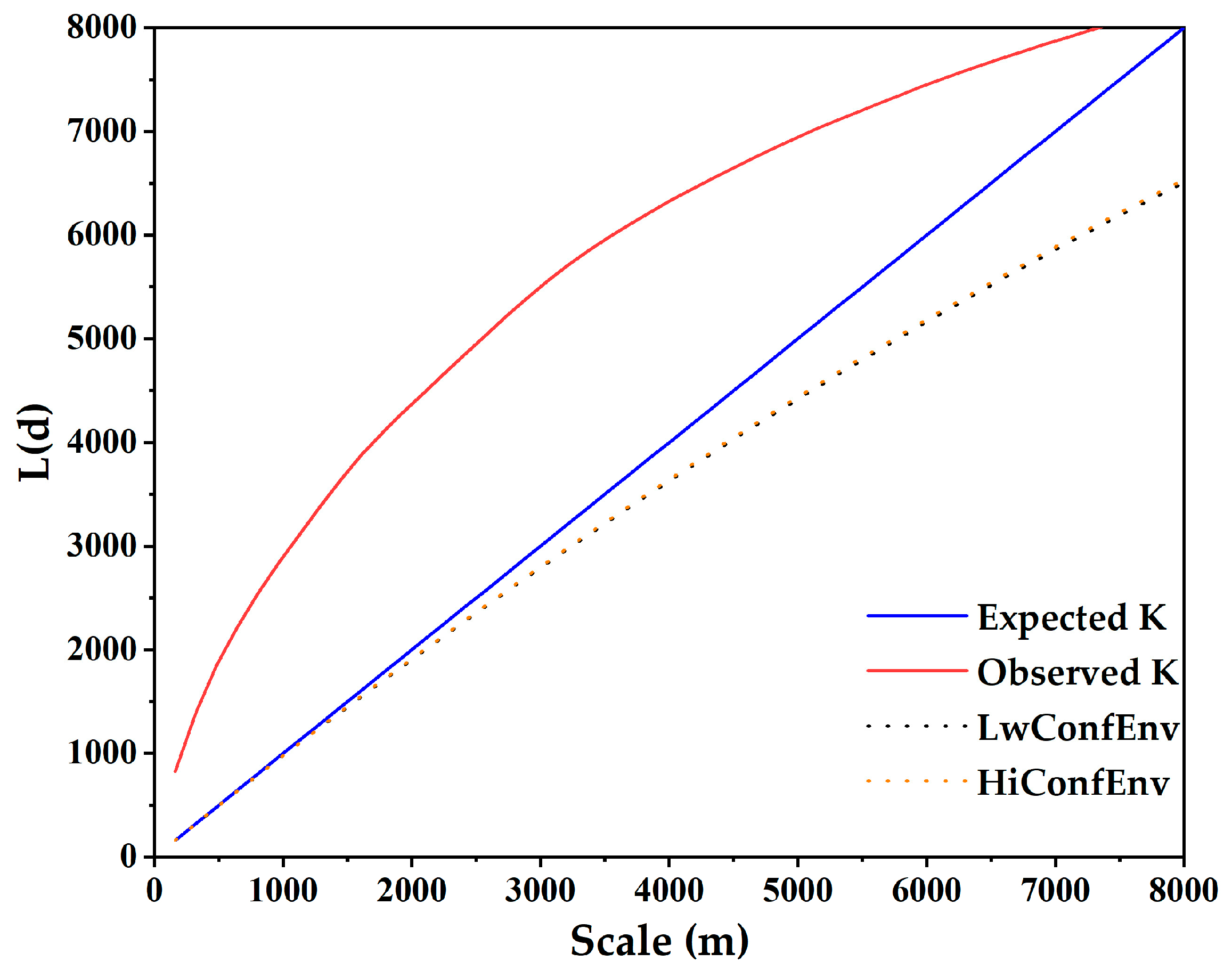
2.3.4. Ripley’s K Function
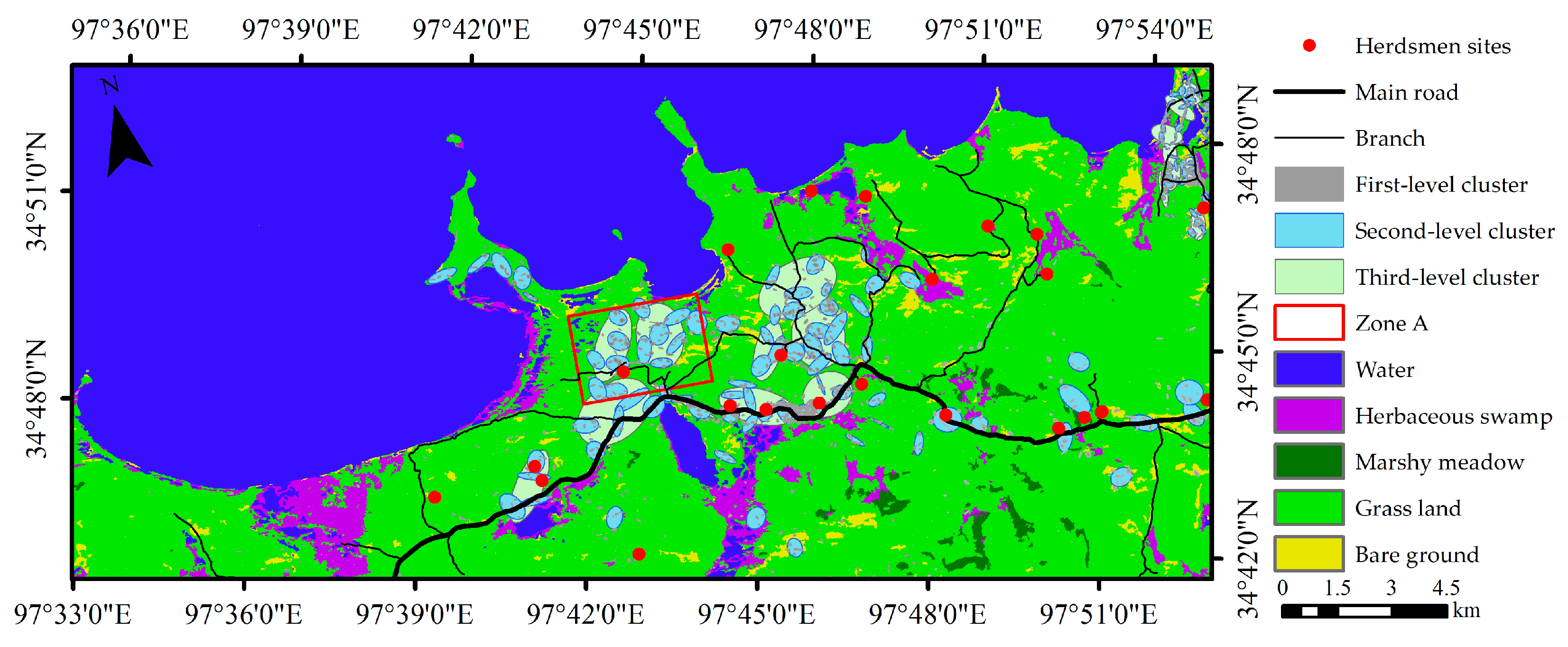
2.3.5. Nearest Neighbor Hierarchical Spatial Cluster (NNH)
3. Results
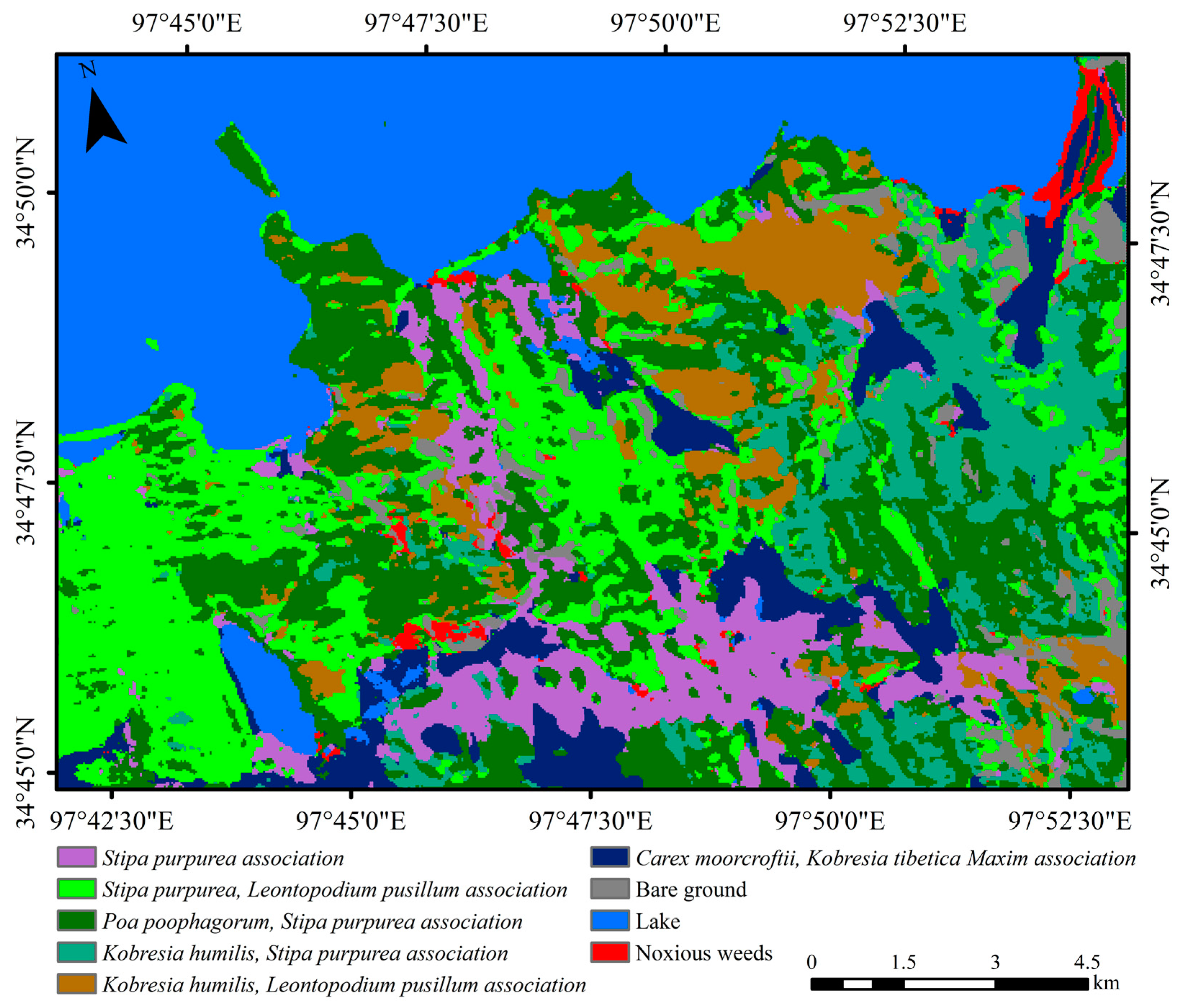
3.1. Classification of Vegetation Associations
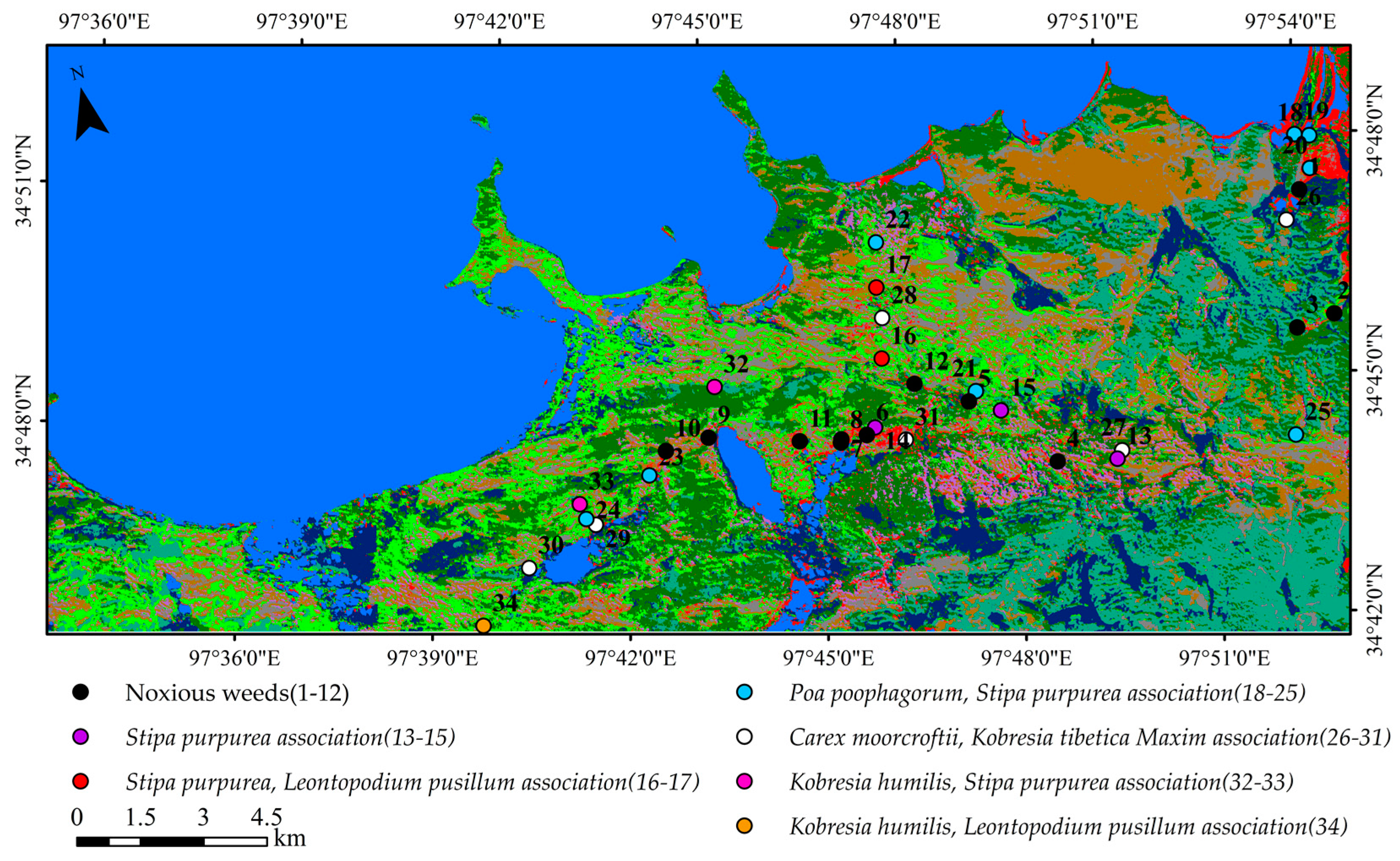
3.1.1. Definition of Vegetation Association and Sample Selection
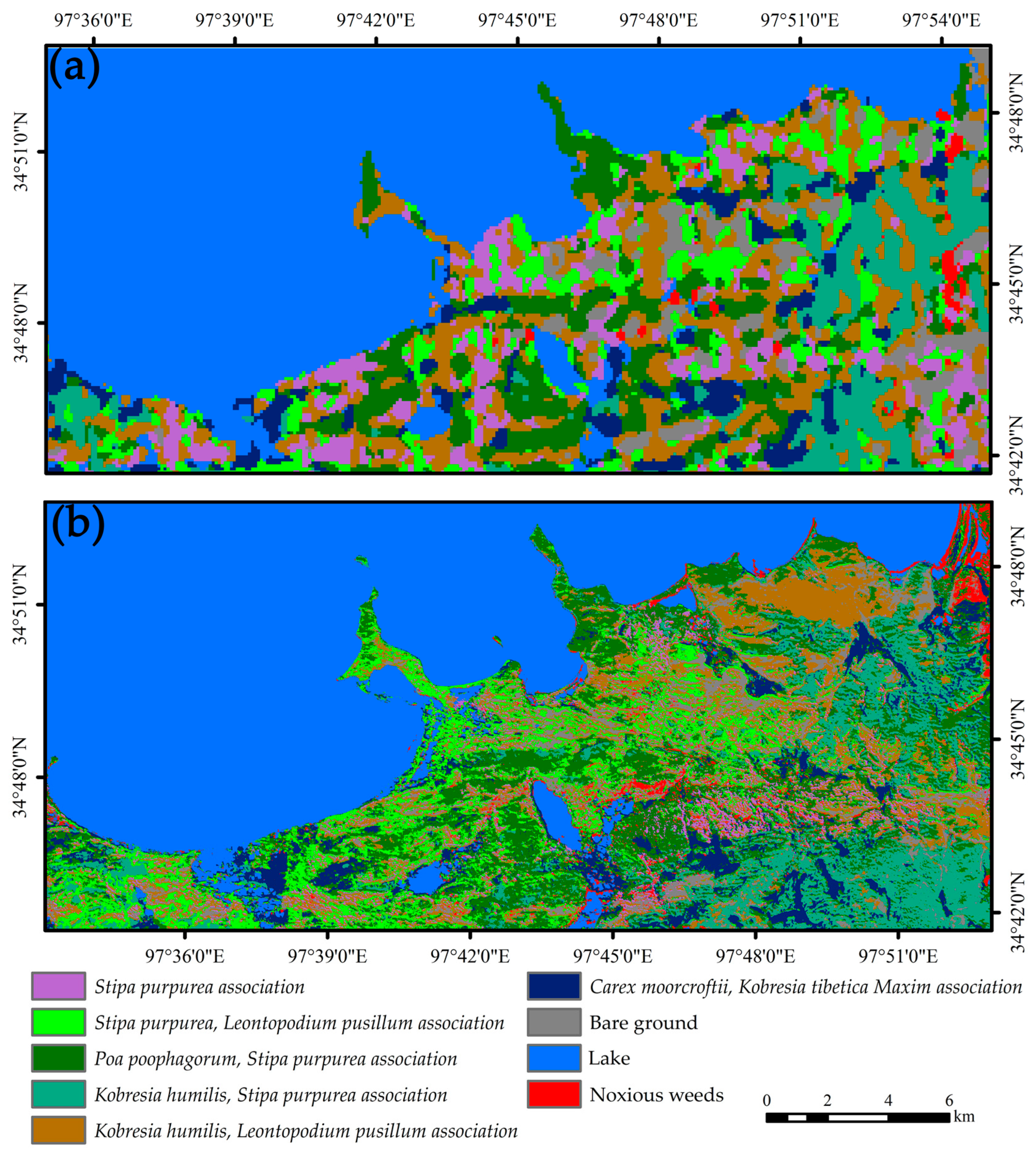
3.1.2. Vegetation Association Classification Results
3.2. Classification Accuracy Verification
3.3. Vegetation Association Variation Analysis
3.4. Temperature and Precipitation Trend Analysis
3.5. Noxious Weed Variation across Altitude and Slope Gradients
3.6. Thermopsis Lanceolata Association Spatial Distribution Pattern
3.7. Thermopsis Lanceolata Association Diffusion Mechanism Analysis
4. Discussion
4.1. Influencing Factors of Vegetation Association Variation
4.2. Spatial Distribution and Diffusion of Noxious Weeds
4.3. Future Research Direction
5. Conclusions
- The vegetation cover area increased, where the noxious weeds area increased more rapidly. The surface temperature and annual precipitation increases were conducive to vegetation restoration, while human activities had the opposite effect on vegetation.
- The noxious weeds area decreased gradually with increasing altitude. Most of noxious weeds were located in the bottomland or on the gentle slope of 4250–4350 m. The noxious weeds area increased in the range of 4250–4400 m from 2013 to 2020, and the increase was highest at 4250–4300 m. The noxious weeds area decreased mainly at 4400–4500 m, and the decrease was largest in the range of 4450–4500 m.
- The Thermopsis lanceolata association distribution was characterized by aggregation, and its diffusion direction was mainly along the road and the herdsmen sites located near the road and around the swamp. The diffusion was attributed to human activities such as human transportation, overgrazing, or the degradation of swamp wetlands.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, Q.G.; Huang, Y.F. Spatial and temporal changes and driving factors of desertification in the Source Region of the Yellow River, China. Nat. Environ. Pollut. Technol. 2020, 19, 1435–1442. [Google Scholar] [CrossRef]
- Lan, Y.C.; Zhao, G.H.; Zhang, Y.N.; Wen, J.; Liu, J.Q.; Hu, X.L. Response of runoff in the source region of the Yellow River to climate warming. Quat. Int. 2010, 226, 60–65. [Google Scholar] [CrossRef]
- Hu, Y.R.; Maskey, S.; Uhlenbrook, S.; Zhao, H.L. Streamflow trends and climate linkages in the source region of the Yellow River, China. Hydrol. Process. 2011, 25, 3399–3411. [Google Scholar] [CrossRef]
- Liu, L.L.; Sayer, E.J.; Deng, M.F.; Li, P.; Liu, W.X.; Wang, X.; Yang, S.; Huang, J.S.; Luo, J.; Su, Y.J.; et al. The grassland carbon cycle: Mechanisms, responses to global changes, and potential contribution to carbon neutrality. Fundam. Res. 2022, 3, 209–218. [Google Scholar] [CrossRef]
- Ibarra-Manriquez, G.; González-Espinosa, M.; Martínez-Ramos, M.; Meave, J.A. From vegetation ecology to vegetation science: Current trends and perspectives. Bot. Sci. 2022, 100, S137–S174. [Google Scholar] [CrossRef]
- Wang, D.N.; Zhao, Y.T.; Yang, W.X.; Ma, K.X.; Hao, T.X.; Zhao, J.W.; Tang, R.; Pu, Y.F.; Zhang, X.J.; Mujtaba, K.G.; et al. Ecological–economic assessment and managerial significance of water conservation in the Headwaters of the Yellow River. Water 2022, 14, 2553. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Wang, W.W.; Tang, X.J.; Li, H.; Hu, H.W.; Wang, X.F. Classification of alpine grasslands in cold and high altitudes based on multispectral Landsat-8 images: A case study in Sanjiangyuan National Park, China. Remote Sens. 2022, 14, 3714. [Google Scholar] [CrossRef]
- Liu, X.F.; Zhang, J.S.; Zhu, X.F.; Pan, Y.Z.; Liu, Y.X.; Zhang, D.H.; Lin, Z.H. Spatiotemporal changes in vegetation coverage and its driving factors in the Three-River Headwaters Region during 2000–2011. J. Geogr. Sci. 2014, 24, 288–302. [Google Scholar] [CrossRef]
- Verrall, B.; Pickering, C.M. Alpine vegetation in the context of climate change: A global review of past research and future directions. Sci. Total Environ. 2020, 748, 141344. [Google Scholar] [CrossRef]
- Salhab, J.; Wang, J.L.; Anjum, S.A.; Chen, Y.X. Assessment of the grassland degradation in the southeastern part of the source region of the Yellow River from 1994 to 2001. J. Food Agric. Environ. 2010, 8, 1367–1372. [Google Scholar]
- Yi, X.S.; Li, G.S.; Yin, Y.Y. The impacts of grassland vegetation degradation on soil hydrological and ecological effects in the source region of the Yellow River--A case study in Junmuchang region of Maqin country. Procedia Environ. Sci. 2012, 13, 967–981. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.T.; Chen, J.J. Research on the variation characteristics of vegetation change and its terrain influence factors in alpine grassland in the Source Region of the Yellow River. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2020, XLII-3/W10, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wang, H.; Jia, H.; Pen, M.; Liu, N.; Wei, J.; Zhou, B. Ecological niche and interspecific association of plant communities in alpine desertification grasslands: A case study of Qinghai Lake Basin. Plants 2022, 11, 2724. [Google Scholar] [CrossRef]
- Callaway, R.M.; DeLuca, T.H.; Belliveau, W.M. Biological-control herbivores may increase competitive ability of the noxious weed Centaurea Maculosa. Ecology 1999, 80, 1196–1201. [Google Scholar] [CrossRef]
- Rocchini, D.; Andreo, V.; Förster, M.; Garzon-Lopez, C.X.; Gutierrez, A.P.; Gillespie, T.W.; Hauffe, H.C.; He, K.S.; Kleinschmit, B.; Mairota, P. Potential of remote sensing to predict species invasions: A modelling perspective. Prog. Phys. Geogr. 2015, 39, 283–309. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Dong, S.K.; Liu, S.L.; Wang, X.X.; Wen, L.; Wu, Y. The interaction between poisonous plants and soil quality in response to grassland degradation in the alpine region of the Qinghai-Tibetan Plateau. Plant Ecol. 2014, 215, 809–819. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.J.; Qin, Q.T.; You, H.T.; Han, X.W.; Zhou, G.Q. Patch pattern and ecological risk assessment of alpine grassland in the Source Region of the Yellow River. Remote Sens. 2020, 12, 3460. [Google Scholar] [CrossRef]
- Piao, S.L.; Mohammat, A.; Fang, J.Y.; Cai, Q.; Feng, J.M. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang.-Hum. Policy Dimens. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Wang, G.X.; Li, Y.S.; Wu, Q.B.; Wang, Y.B. Impacts of permafrost changes on alpine ecosystem in Qinghai-Tibet Plateau. Sci. China Ser. D Earth Sci. 2006, 49, 1156–1169. [Google Scholar] [CrossRef]
- Zhao, Z.P.; Wu, X.P.; Li, G.; Li, J.S. The cause of grassland degradation in Golog Tibetan Autonomous Prefecture in the Three Rivers Headwaters Region of Qinghai Province. Shengtai Xuebao/ Acta Ecol. Sin. 2013, 33, 6577–6586. [Google Scholar] [CrossRef]
- Yang, Z.P.; Gao, J.X.; Zhou, C.P.; Shi, P.L.; Zhao, L.; Shen, W.S.; Ouyang, H. Spatio-temporal changes of NDVI and its relation with climatic variables in the source regions of the Yangtze and Yellow rivers. J. Geogr. Sci. 2011, 21, 979–993. [Google Scholar] [CrossRef]
- Liu, X.F.; Zhu, X.F.; Zhu, W.Q.; Pan, Y.Z.; Zhang, C.; Zhang, D.H. Changes in spring phenology in the Three-Rivers Headwater region from 1999 to 2013. Remote Sens. 2014, 6, 9130–9144. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Wang, M.; Pang, Z.; Jiang, W.; Lu, J.; Yang, K.; Li, L.; Li, X.; Qu, W. Spatiotemporal variation of NDVI in the Yellow River Source Region from 1998 to 2016. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2020, V-3-2020, 739–744. [Google Scholar] [CrossRef]
- Shen, X.J.; An, R.; Feng, L.; Ye, N.; Zhu, L.J.; Li, M.H. Vegetation changes in the Three-River Headwaters Region of the Tibetan Plateau of China. Ecol. Indic. 2018, 93, 804–812. [Google Scholar] [CrossRef]
- Liu, Y.N. Development of hyperspectral imaging remote sensing technology. J. Remote Sens. 2021, 25, 439–459. [Google Scholar] [CrossRef]
- An, R.; Lu, C.H.; Wang, H.L.; Jiang, D.P.; Sun, M.Q.; Ballard, J.A.Q. Remote sensing identification of rangeland degradation using Hyperion hyperspectral image in a typical area for Three-River Headwater Region, Qinghai, China. Geomat. Inf. Sci. Wuhan Univ. 2018, 43, 399–405. [Google Scholar] [CrossRef]
- Xing, F.; An, R.; Wang, B.; Miao, J.; Jiang, T.; Huang, X.; Hu, Y. Mapping the occurrence and spatial distribution of noxious weed species with multisource data in degraded grasslands in the Three-River Headwaters Region, China. Sci. Total Environ. 2021, 801, 149714. [Google Scholar] [CrossRef]
- Sonobe, R.; Yamaya, Y.; Tani, H.; Wang, X.; Kobayashi, N.; Mochizuki, K.-I. Crop classification from sentinel-2-derived vegetation indices using ensemble learning. J. Appl. Remote Sens. 2018, 12, 026019. [Google Scholar] [CrossRef] [Green Version]
- Dobrinić, D.; Gašparović, M.; Medak, D. Sentinel-1 and 2 time-series for vegetation mapping using random forest classification: A case study of northern Croatia. Remote Sens. 2021, 13, 2321. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, L.; Zhang, X.L. Three-dimensional convolutional neural network model for tree species classification using airborne hyperspectral images. Remote Sens. Environ. 2020, 247, 111938. [Google Scholar] [CrossRef]
- Dahiya, N.; Singh, S.; Gupta, S. A review on deep learning classifier for hyperspectral imaging. Int. J. Image Graph. 2022, 2350036. [Google Scholar] [CrossRef]
- Wang, P.; Wang, L.; Leung, H.; Zhang, G. Super-resolution mapping based on spatial–spectral correlation for spectral imagery. IEEE Trans. Geosci. Remote Sens. 2021, 59, 2256–2268. [Google Scholar] [CrossRef]
- Shang, X.; Song, M.; Wang, Y.; Yu, C.; Yu, H.; Li, F.; Chang, C.-I. Target-constrained interference-minimized band selection for hyperspectral target detection. IEEE Trans. Geosci. Remote Sens. 2021, 59, 6044–6064. [Google Scholar] [CrossRef]
- Hussain, S.A.; Tahir, A.; Khan, J.A.; Salman, A. Pixel-based classification of hyperspectral images using Convolutional Neural Networks. PFG-J. Photogramm. Remote Sens. Geoinf. Sci. 2019, 87, 33–45. [Google Scholar] [CrossRef]
- Chen, Y.S.; Jiang, H.L.; Li, C.Y.; Jia, X.P.; Ghamisi, P. Deep feature extraction and classification of hyperspectral images based on convolutional neural networks. IEEE Trans. Geosci. Remote Sens. 2016, 54, 6232–6251. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Luo, S.Q.; Li, Z.G.; Wang, S.Y.; Li, Z.H. The freeze/thaw process and the surface energy budget of the seasonally frozen ground in the source region of the Yellow River. Theor. Appl. Climatol. 2019, 138, 1631–1646. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Gu, X.; Yu, T.; He, H.; Zhu, L.; Wang, F. Study of data preprocess for HJ-1A satellite HSI image. Remote Sens. Environ. 2015, 9669, 79–84. [Google Scholar] [CrossRef]
- Chen, W.H.; Pan, J.; Sun, Y.L. Tree species classification based on fusion images by GF-5 and Sentinel-2A. Remote Sens. 2022, 14, 5088. [Google Scholar] [CrossRef]
- Dai, J.; Roberts, D.A.; Stow, D.A.; An, L.; Hall, S.J.; Yabiku, S.T.; Kyriakidis, P.C. Mapping understory invasive plant species with field and remotely sensed data in Chitwan, Nepal. Remote Sens. Environ. 2020, 250, 112037. [Google Scholar] [CrossRef]
- Hu, P.P. Analyzing the Spatiotemporal Variations of the Land Surface Temperature on the Tibetan Plateau from 2003–2020 Based on MODIS Data; China University of Geosciences: Beijing, China, 2021. (In Chinese) [Google Scholar]
- Li, P.H. Extraction of Wetland Information in Typical Areas of the Source of the Yellow River Based on Multi-Source Data Feature Optimization; China University of Geosciences: Beijing, China, 2021. (In Chinese) [Google Scholar]
- Alzubaidi, L.; Zhang, J.; Humaidi, A.J.; Al-Dujaili, A.; Duan, Y.; Al-Shamma, O.; Santamaria, J.; Fadhel, M.A.; Al-Amidie, M.; Farhan, L. Review of deep learning: Concepts, CNN architectures, challenges, applications, future directions. J. Big Data 2021, 8, 53. [Google Scholar] [CrossRef]
- Teja, K.; Jens, L.; Felix, S.; Stefan, H. Review on Convolutional Neural Networks (CNN) in vegetation remote sensing. ISPRS J. Photogramm. Remote Sens. 2021, 173, 24–49. [Google Scholar] [CrossRef]
- Chollet, F. Xception: Deep Learning with Depthwise Separable Convolutions. In Proceedings of the 30th Ieee Conference On Computer Vision And Pattern Recognition (Cvpr 2017), Honolulu, HI, USA, 21–26 July 2017; pp. 1800–1807. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.K.; Krishna, G.; Dubey, S.R.; Chaudhuri, B.B. HybridSN: Exploring 3-D–2-D CNN feature hierarchy for hyperspectral image classification. IEEE Geosci. Remote Sens. Lett. 2020, 17, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Qian, Y.R.; Du, J.; Fan, Y.G. The application of the convolution neural network to grassland classification in remote sensing images. J. Northeast Norm. Univ. Nat. Sci. Ed. 2019, 51, 53–58. (In Chinese) [Google Scholar]
- Hu, Y.; Batunacun. An analysis of Land-Use and Land-Cover change in the Zhujiang–Xijiang economic belt, China, from 1990 to 2017. Appl. Sci. 2018, 8, 1524. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zhang, L.J.; Zhang, Z.Q.; Li, X.Y.; Wang, H.Y.; Xi, X. Spatiotemporal variation and influence factors of vegetation cover in the Yellow River Basin (1982–2021) based on GIMMS NDVI and MOD13A1. Water 2022, 14, 3274. [Google Scholar] [CrossRef]
- Yan, X.; Wang, R.; Niu, Z.G. Response of China’s wetland NDVI to climate changes. Wetlands 2022, 42, 55. [Google Scholar] [CrossRef]
- Yao, J.; Zhang, C.Y.; Zhao, X.H. Species spatial distribution patterns and species associations in a broad-leaved korean pine forest in Jiaohe, Jilin Province. Linye Kexue/Sci. Silvae Sin. 2018, 54, 23–31. [Google Scholar] [CrossRef]
- Dong, Q.M.; Ma, Y.S.; Xu, C.J.; Shi, J.J.; Wang, Y.; Wang, Y.L.; Shang, L.; Li, S.X. Study of classification and gradation, restoration of black-soil beach degraded grassland in the Headwaters of Three Rivers. Acta Agrestia Sin. 2015, 3, 441–447. (In Chinese) [Google Scholar]
- Chen, J. The Distribution Patternd and the Major Influencing Factors of Invasive Alien Species in China. Ph.D. Thesis, Nanjing Forestry University, Nanjing, China, 2020. (In Chinese). [Google Scholar]
- Hou, Q.C.; Feng, Y.L.; Zhou, Y.J.; Ao, Y.M.; Chen, C.X.; Xing, Y.J.; Wang, Q.G.; Yan, G.Y. Main hypotheses on mechanisms underlying plant invasion: A review. J. Appl. Ecol. 2022, 33, 3105–3115. (In Chinese) [Google Scholar]
- Xu, L.; Zhang, Y.; Mohamad, O.; Jiang, C.Y.; Friman, V. Mesorhizobium zhangyense sp. nov., isolated from wild Thermopsis lanceolate in northwestern China. Arch. Microbiol. 2018, 200, 603–610. [Google Scholar] [CrossRef]
- Jin, X.Y.; Jin, H.J.; Luo, D.L.; Sheng, Y.; Wu, Q.B.; Wu, J.C.; Wang, W.H.; Huang, S.; Li, X.Y.; Liang, S.H.; et al. Impacts of permafrost degradation on hydrology and vegetation in the Source Area of the Yellow River on Northeastern Qinghai-Tibet Plateau, Southwest China. Front. Earth Sci. 2022, 10, 845824. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Zhang, X.F. Mechanisms of osmotic adjustment of Themopsis lanceolata test–tuble plantlets in adapting to salt stress. Acta Bot. Boreali-Occident. Sin. 2014, 34, 963–969. (In Chinese) [Google Scholar]
- Kalisz, S.; Kivlin, S.N.; Bialic-Murphy, L. Allelopathy is pervasive in invasive plants. Biol. Invasions 2021, 23, 367–371. [Google Scholar] [CrossRef]
- Wang, G.Q.; Li, J.Y.; Sivakumar, B.; Li, T.J.; Chen, C. Attribution of growing season vegetation activity to climate change and human activities in the Three-River Headwaters Region, China. J. Hydroinform. 2020, 22, 186–204. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.C.; Jin, X.M. Vegetation dynamics and responses to climate change and anthropogenic activities in the Three-River Headwaters Region, China. Ecol. Indic. 2021, 131, 108223. [Google Scholar] [CrossRef]
- Xu, K.J.; Zhang, Z.Y.; Yu, W.W.; Zhao, P.; Yue, J.B.; Deng, Y.P.; Geng, J. How spatial resolution affects forest phenology and tree-species classification based on satellite and up-scaled time-series images. Remote Sens. 2021, 13, 2716. [Google Scholar] [CrossRef]
- Roberts, D.A.; Gardner, M.E.; Church, R.; Ustin, S.L.; Green, R.O. Optimum Strategies for Mapping Vegetation Using Multiple-Endmember Spectral Mixture Models. In Proceedings of the Imaging spectrometry III, San Diego, CA, USA, 28–30 July 1997; pp. 108–119. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Acquisition Date | Sensor | Satellite | Number of Bands | Spatial Resolution (m) |
|---|---|---|---|---|---|
| 1 | 16 August 2013 | HSI | HJ-1A | 115 | 100 |
| 2 | 13 August 2019 | AHSI | GF-5 | 330 | 30 |
| 3 | 24 August 2020 | CMOS | OHS-3C | 32 | 10 |
| Vegetation Types | Longitude and Latitude | Photos |
|---|---|---|
| Carex moorcroftii, Kobresia tibetica Maxim cluster | 34°48’N, 97°53’E |  |
| Kobresia humilis, Leontopodium pusillum association | 34°45’N, 97°53’E |  |
| Kobresia humilis, Stipa purpurea association | 34°45’N, 97°46’E |  |
| Poa poophagorum, Stipa purpurea association | 34°45’N, 97°41’E |  |
| Stipa purpurea association | 34°47’N, 97°46’E |  |
| Stipa purpurea, Leontopodium pusillum association | 34°46’E, 97°46’N |  |
| Noxious weeds (Oxytropis ochrocephala Bunge in the picture) | 34°45’E, 97°45’N |  |
| Vegetation Association | Dominant Species | Subdominant Species | Companion Species |
|---|---|---|---|
| Stipa purpurea association | Stipa purpurea | Elymus dahuricus Turcz, Roegneria thoroldiana, Kobresia humilis, etc. | |
| Carex moorcroftii, Kobresia tibetica Maxim association | Carex moorcroftii | Kobresia tibetica Maxim | Scirpus distigmaticus, Pteridophyta, Cremanthodium Benth, etc. |
| Kobresia humilis, Stipa purpurea association | Kobresia humilis | Stipa purpurea | Poa poophagorum, Kobresia humilis, Leontopodium pusillum, etc. |
| Poa poophagorum, Stipa purpurea association | Poa poophagorum | Stipa purpurea | Elymus dahuricus Turcz, Kobresia humilis, etc. |
| Kobresia humilis, Leontopodium pusillum association | Kobresia humilis | Leontopodium pusillum | Oxytropis, Stipa purpurea, Poa poophagorum, etc. |
| Stipa purpurea, Leontopodium pusillum association | Stipa purpurea | Leontopodium pusillum | Oxytropis, Ajania khartensis, Poa poophagorum, etc. |
| Noxious weeds | Thermopsis lanceolate, Ligularia virgaurea, Aconitum pendulum, Oxytropis, Leontopodium pusillum, Saussurea japonica, Polygonum sibiricum Laxm, Ajania khartensis, Pedicularis kansuensis Maxim, etc. | ||
| Categories | HJ-1A | GF-5 | OHS-3C | |||
|---|---|---|---|---|---|---|
| Number of Training Samples | Number of Testing Samples | Numbersof Training Samples | Number of Testing Samples | Number of Training Samples | Number of Testing Samples | |
| Lake | 410 | 176 | 758 | 325 | 816 | 350 |
| Bare ground | 102 | 44 | 162 | 70 | 729 | 312 |
| Noxious weeds | 13 | 6 | 48 | 20 | 319 | 137 |
| Stipa purpurea association | 67 | 29 | 276 | 118 | 509 | 218 |
| Carex moorcroftii, Kobresia tibetica Maxim association | 87 | 37 | 206 | 88 | 888 | 380 |
| Kobresia humilis, Stipa purpurea association | 86 | 37 | 223 | 95 | 1165 | 499 |
| Poa poophagorum, Stipa purpurea association | 86 | 37 | 269 | 115 | 1194 | 512 |
| Kobresia humilis, Leontopodium pusillum association | 111 | 47 | 134 | 58 | 792 | 340 |
| Stipa purpurea, Leontopodium pusillum association | 90 | 38 | 263 | 113 | 869 | 373 |
| Total | 1052 | 451 | 2339 | 1002 | 7281 | 3121 |
| Categories | UA (%) | PA (%) | ||||
|---|---|---|---|---|---|---|
| HJ-1A | GF-5 | OHS-3C | HJ-1A | GF-5 | OHS-3C | |
| Noxious weeds | 83.33 | 100.00 | 97.46 | 100.00 | 100.00 | 99.14 |
| Lake | 99.78 | 99.75 | 100.00 | 100.00 | 100.00 | 100.00 |
| Bare ground | 100.00 | 100.00 | 100.00 | 97.73 | 100.00 | 100.00 |
| Carex moorcroftii, Kobresia tibetica Maxim association | 100.00 | 100.00 | 98.58 | 100.00 | 97.98 | 99.71 |
| Kobresia humilis, Leontopodium pusillum association | 100.00 | 100.00 | 98.04 | 100.00 | 98.82 | 99.34 |
| Kobresia humilis, Stipa purpurea association | 97.71 | 97.35 | 100.00 | 98.46 | 100.00 | 99.33 |
| Poa poophagorum, Stipa purpurea association | 98.96 | 98.79 | 99.41 | 99.31 | 99.19 | 98.82 |
| Stipa purpurea, Leontopodium pusillum association | 99.42 | 99.32 | 98.55 | 97.16 | 96.08 | 96.88 |
| Stipa purpurea association | 100.00 | 96.47 | 96.44 | 99.00 | 98.80 | 98.19 |
| Categories | 2013 (km2) | Proportion (%) | 2020 (km2) | Proportion (%) | Growth Rate (%) |
|---|---|---|---|---|---|
| Lake | 208 | 47.59% | 195.44 | 44.71% | −6.04% |
| Bare ground | 19.23 | 4.40% | 17.24 | 3.94% | −10.35% |
| Noxious weeds | 2.88 | 0.66% | 9.02 | 2.06% | 213.19% |
| Stipa purpurea association | 28.62 | 6.55% | 14.7 | 3.36% | −48.64% |
| Carex moorcroftii, Kobresia tibetica Maxim association | 20.72 | 4.74% | 24.13 | 5.52% | 16.46% |
| Poa poophagorum, Stipa purpurea association | 42.56 | 9.74% | 57.42 | 13.14% | 34.92% |
| Kobresia humilis, Stipa purpurea association | 29.8 | 6.82% | 40.19 | 9.19% | 34.87% |
| Kobresia humilis, Leontopodium pusillum association | 57.47 | 13.15% | 38.05 | 8.70% | −33.79% |
| Stipa purpurea, Leontopodium pusillum association | 27.83 | 6.37% | 40.92 | 9.36% | 47.04% |
| Categories | Proportion in 2019 (%) | Proportion in 2020 (%) | Difference (%) |
|---|---|---|---|
| Lake | 27.82% | 25.81% | 2.01% |
| Bare ground | 4.55% | 3.10% | 1.45% |
| Noxious weeds | 1.65% | 1.41% | 0.24% |
| Stipa purpurea association | 7.14% | 8.62% | −1.48% |
| Carex moorcroftii, Kobresia tibetica Maxim association | 6.82% | 3.80% | 3.02% |
| Poa poophagorum, Stipa purpurea association | 20.68% | 22.34% | −1.66% |
| Kobresia humilis, Stipa purpurea association | 7.37% | 9.31% | −1.94% |
| Kobresia humilis, Leontopodium pusillum association | 8.46% | 11.17% | −2.71% |
| Stipa purpurea, Leontopodium pusillum association | 15.51% | 14.44% | 1.07% |
| 2013 | NW | Lake | BG | CK | KL | KS | PS | SL | SC | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | |||||||||||
| NW | 0.37 | 2.10 | 1.47 | 0.45 | 1.76 | 0.31 | 0.85 | 0.70 | 1.01 | 9.02 | |
| Lake | 0.09 | 191.74 | 0.18 | 0.90 | 0.91 | 0.08 | 0.59 | 0.55 | 0.4 | 195.44 | |
| BG | 0.14 | 0.43 | 4.35 | 0.28 | 4.77 | 0.68 | 0.84 | 2.92 | 2.83 | 17.24 | |
| CK | 0.21 | 2.74 | 0.53 | 7.54 | 2.89 | 4.07 | 3.54 | 1.29 | 1.32 | 24.13 | |
| KL | 0.36 | 0.96 | 5.08 | 0.66 | 11.99 | 1.82 | 3.52 | 7.01 | 6.65 | 38.05 | |
| KS | 0.62 | 1.58 | 1.79 | 3.83 | 8.44 | 13.04 | 5.40 | 2.77 | 2.72 | 40.19 | |
| PS | 0.78 | 4.10 | 1.92 | 4.63 | 11.81 | 7.00 | 17.95 | 4.53 | 4.70 | 57.42 | |
| SL | 0.16 | 3.66 | 2.49 | 2.03 | 10.55 | 1.96 | 7.89 | 5.88 | 6.30 | 40.92 | |
| SC | 0.15 | 0.69 | 1.42 | 0.40 | 4.35 | 0.84 | 1.98 | 2.18 | 2.69 | 14.70 | |
| Total | 2.88 | 208.00 | 19.23 | 20.72 | 57.47 | 29.80 | 42.56 | 27.83 | 28.62 | 437.11 | |
| Scheme 0. | 0–7 | 7–25 |
|---|---|---|
| Grade | Bottomland | Gentle slope |
| Altitude/m | 4200–4250 | 4250–4300 | 4300–4350 | 4350–4400 | 4400–4450 | 4450–4500 | 4500–4550 |
|---|---|---|---|---|---|---|---|
| Grade | I | II | III | IV | V | VI | VII |
| Categories | Area in 2013 (km2) | Area in 2020 (km2) | Difference (km2) | Growth Rate (%) |
|---|---|---|---|---|
| Water | 1.09 | 0.94 | −0.15 | −13.76 |
| Herbaceous swamp | 1.00 | 0.69 | −0.31 | −31.00 |
| Grass land | 29.65 | 31.42 | 1.77 | 6.00 |
| Bare ground | 3.25 | 1.94 | −1.31 | −40.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wang, G.; Shi, Y.; Liang, S.; Jia, J. Vegetation Types Variations to the South of Ngoring Lake from 2013 to 2020, Analyzed by Hyperspectral Imaging. Remote Sens. 2023, 15, 3174. https://doi.org/10.3390/rs15123174
Liu X, Wang G, Shi Y, Liang S, Jia J. Vegetation Types Variations to the South of Ngoring Lake from 2013 to 2020, Analyzed by Hyperspectral Imaging. Remote Sensing. 2023; 15(12):3174. https://doi.org/10.3390/rs15123174
Chicago/Turabian StyleLiu, Xiaole, Guangjun Wang, Yu Shi, Sihai Liang, and Jinzhang Jia. 2023. "Vegetation Types Variations to the South of Ngoring Lake from 2013 to 2020, Analyzed by Hyperspectral Imaging" Remote Sensing 15, no. 12: 3174. https://doi.org/10.3390/rs15123174
APA StyleLiu, X., Wang, G., Shi, Y., Liang, S., & Jia, J. (2023). Vegetation Types Variations to the South of Ngoring Lake from 2013 to 2020, Analyzed by Hyperspectral Imaging. Remote Sensing, 15(12), 3174. https://doi.org/10.3390/rs15123174