



Spatiotemporal Variations of Global Terrestrial Typical Vegetation EVI and Their Responses to Climate Change from 2000 to 2021
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data and Processing
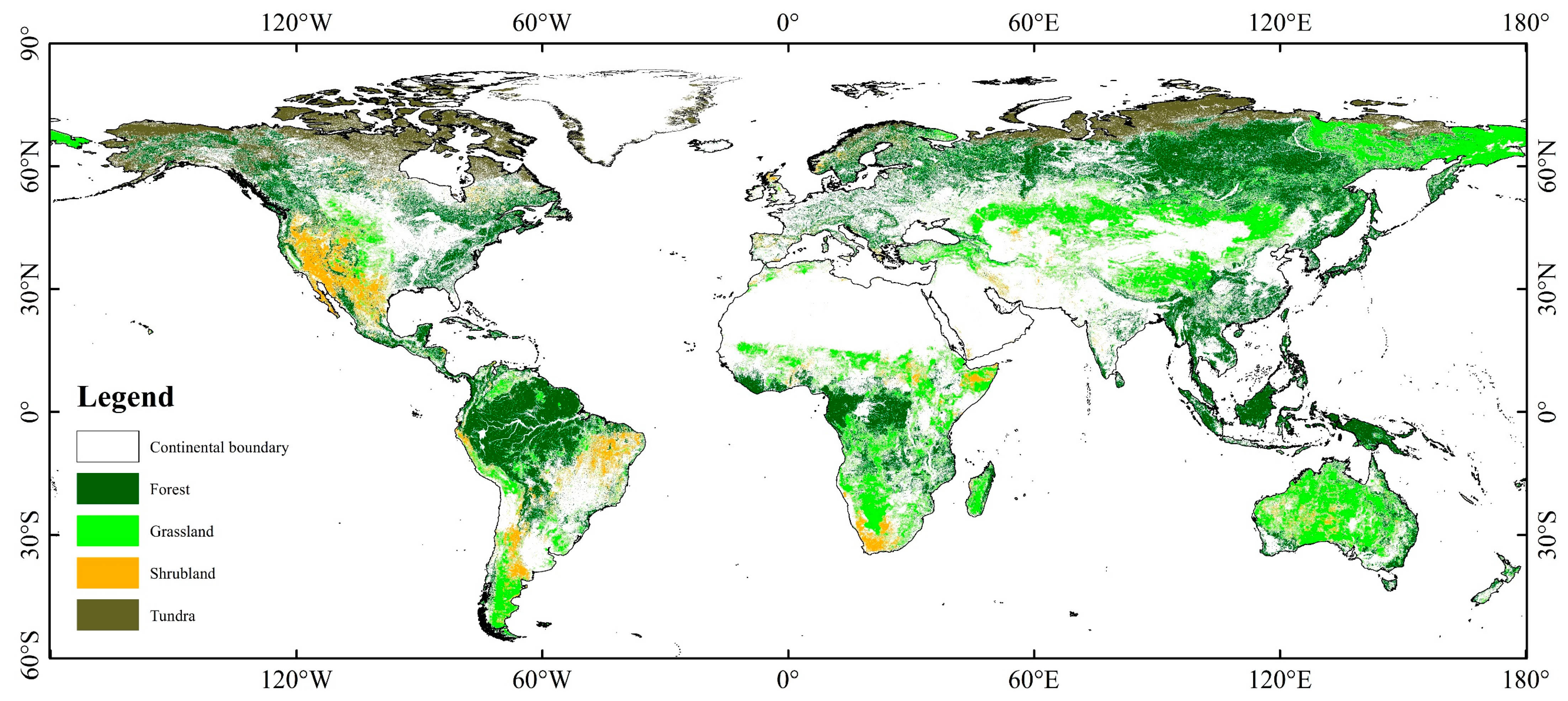
2.1.1. Land Cover Data
2.1.2. EVI Data
2.1.3. Meteorological Data
2.2. Methods
2.2.1. Liner Regression Analysis
2.2.2. Partial Correlation Analysis
2.2.3. Extreme Climate Indices
2.2.4. Grey Relation Analysis
- Establish a comparison sequence and a reference sequence.
- 2.
- Standardized sequence.
- 3.
- Calculate the relation coefficient.
- 4.
- Calculate the grey relation grade.
3. Results
3.1. Spatiotemporal Characteristics of EVI in FGST
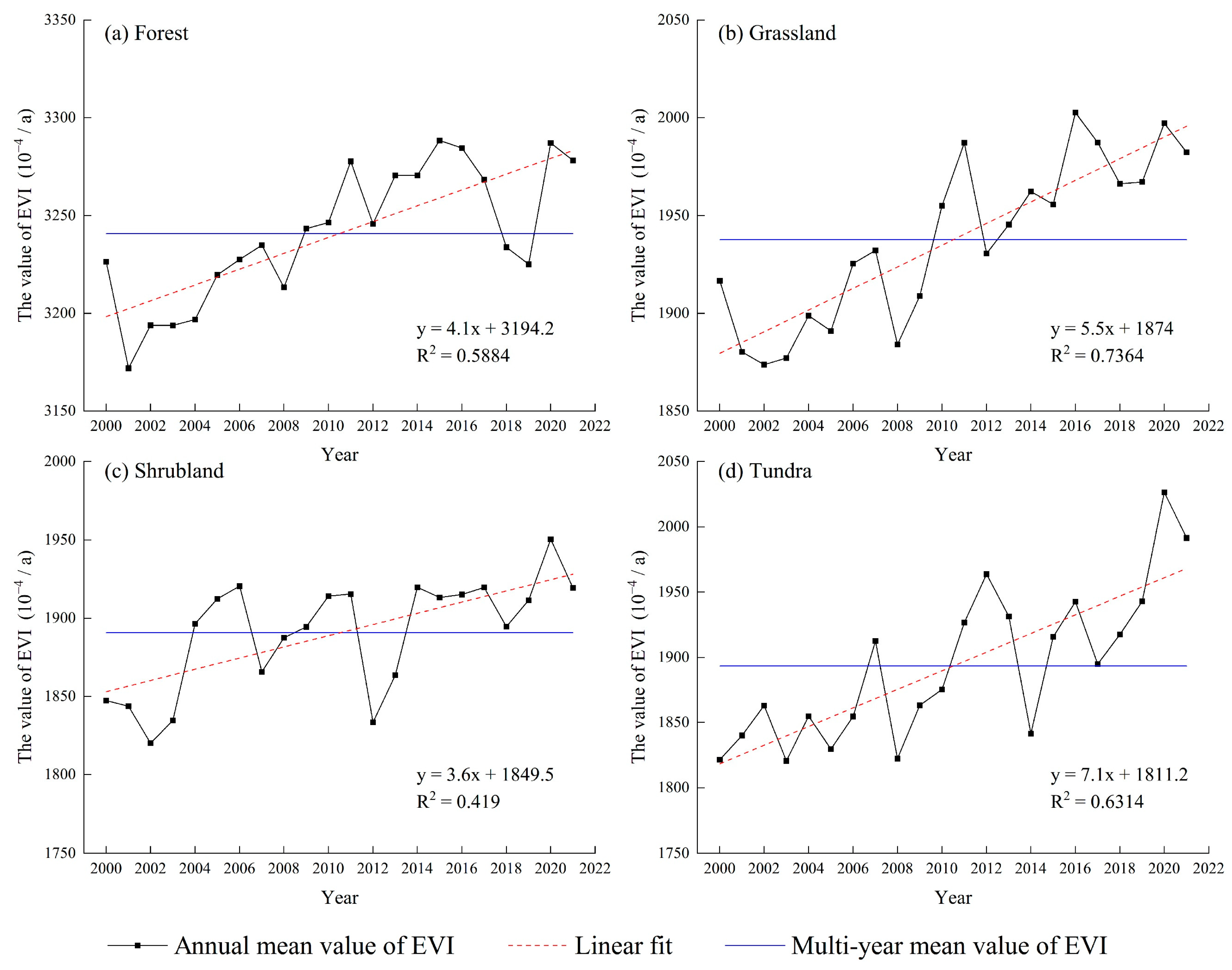
3.1.1. Interannual Variation Characteristics of EVI in FGST
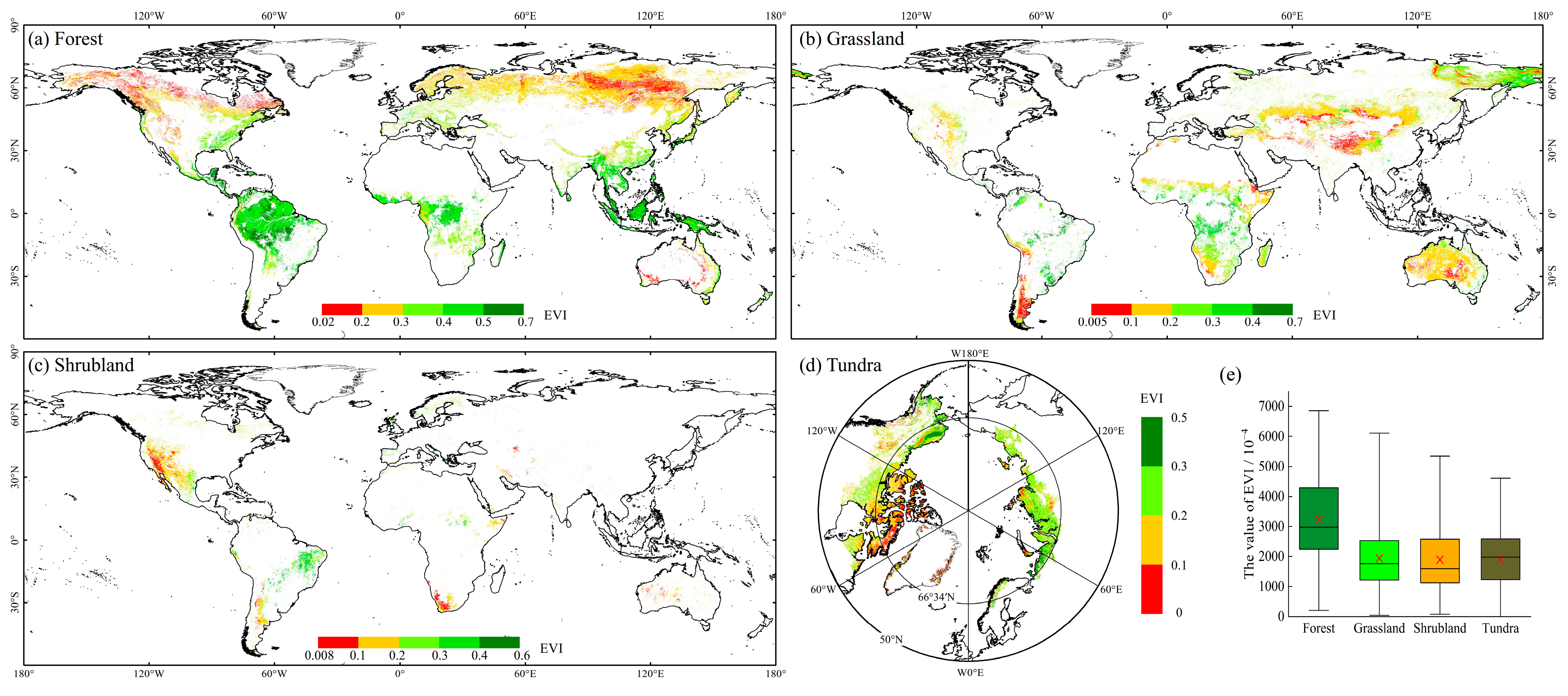
3.1.2. Spatial Characteristics of EVI in FGST
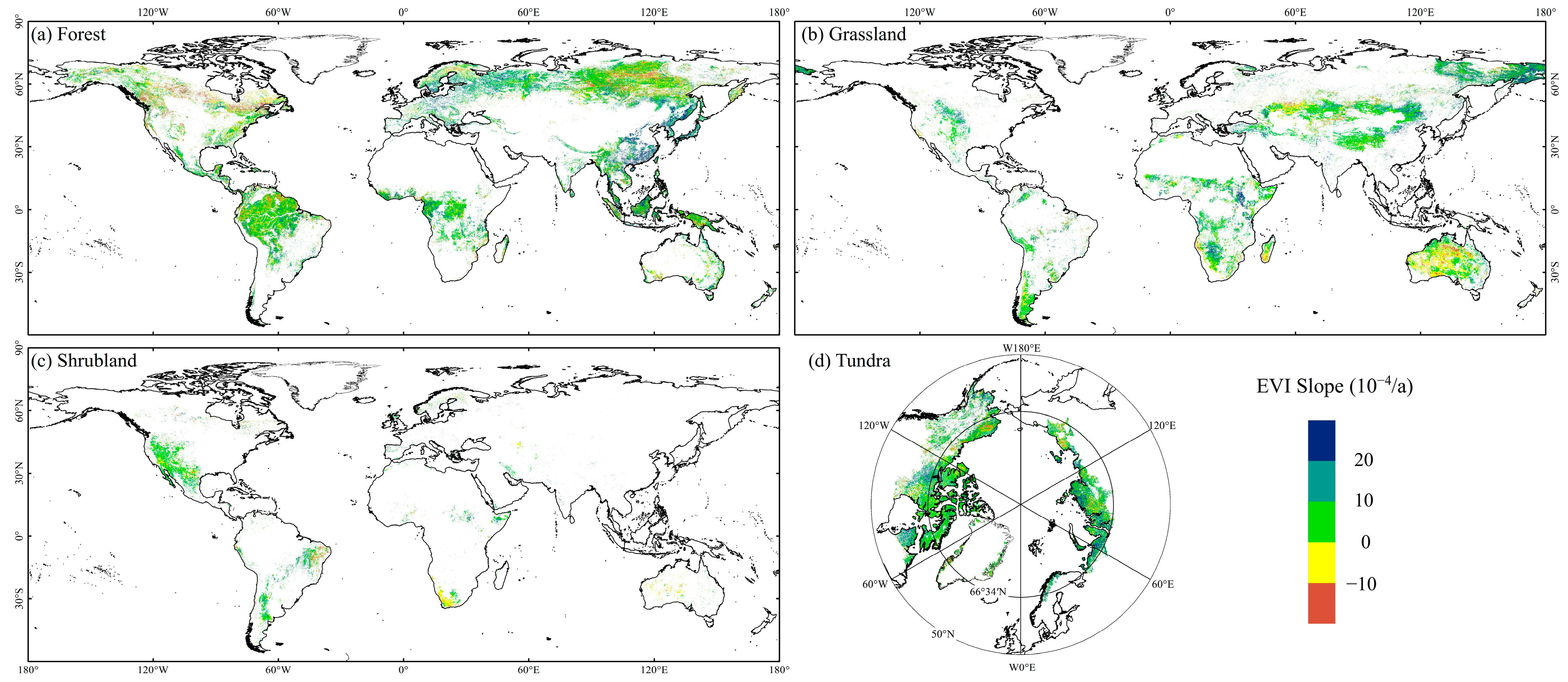
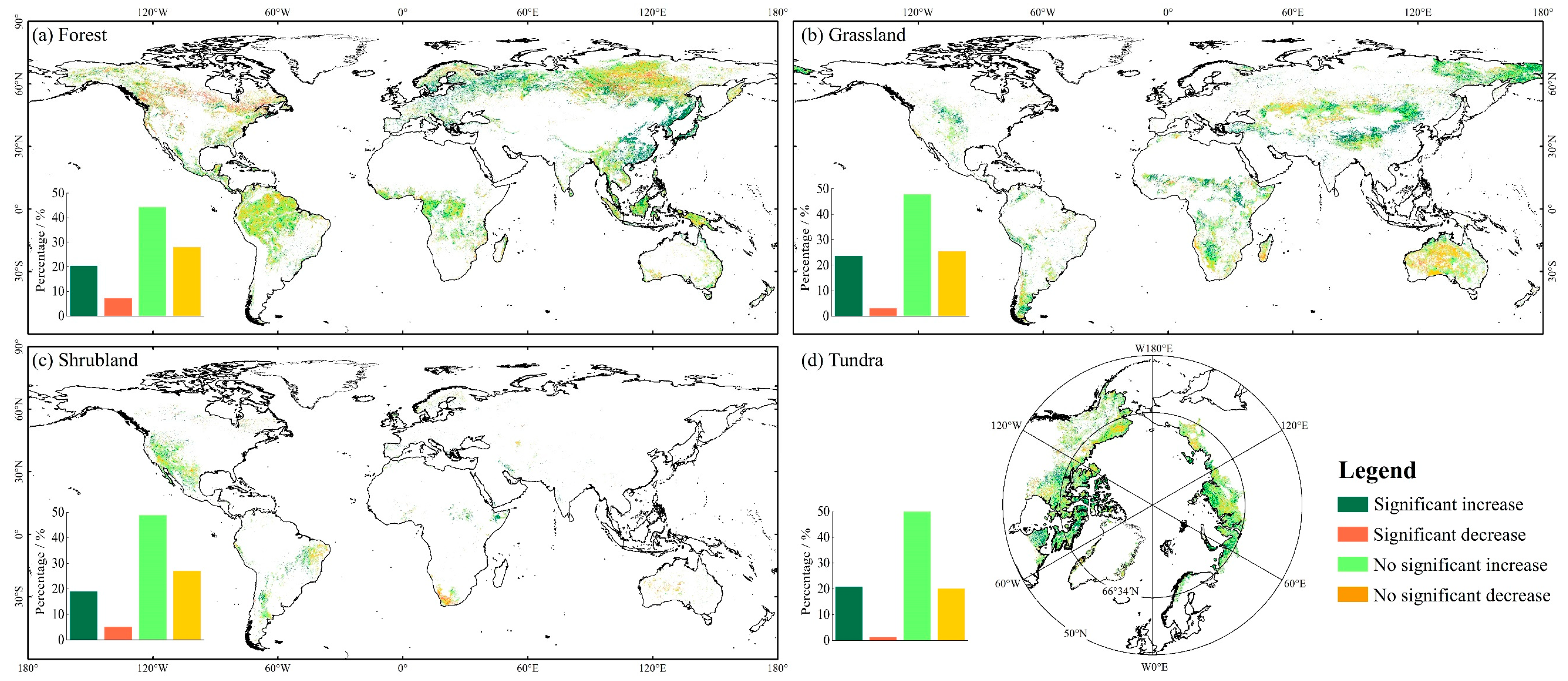
3.1.3. Spatial Variation of EVI in FGST
3.2. Relationship between EVI and Climate Factors in FGST
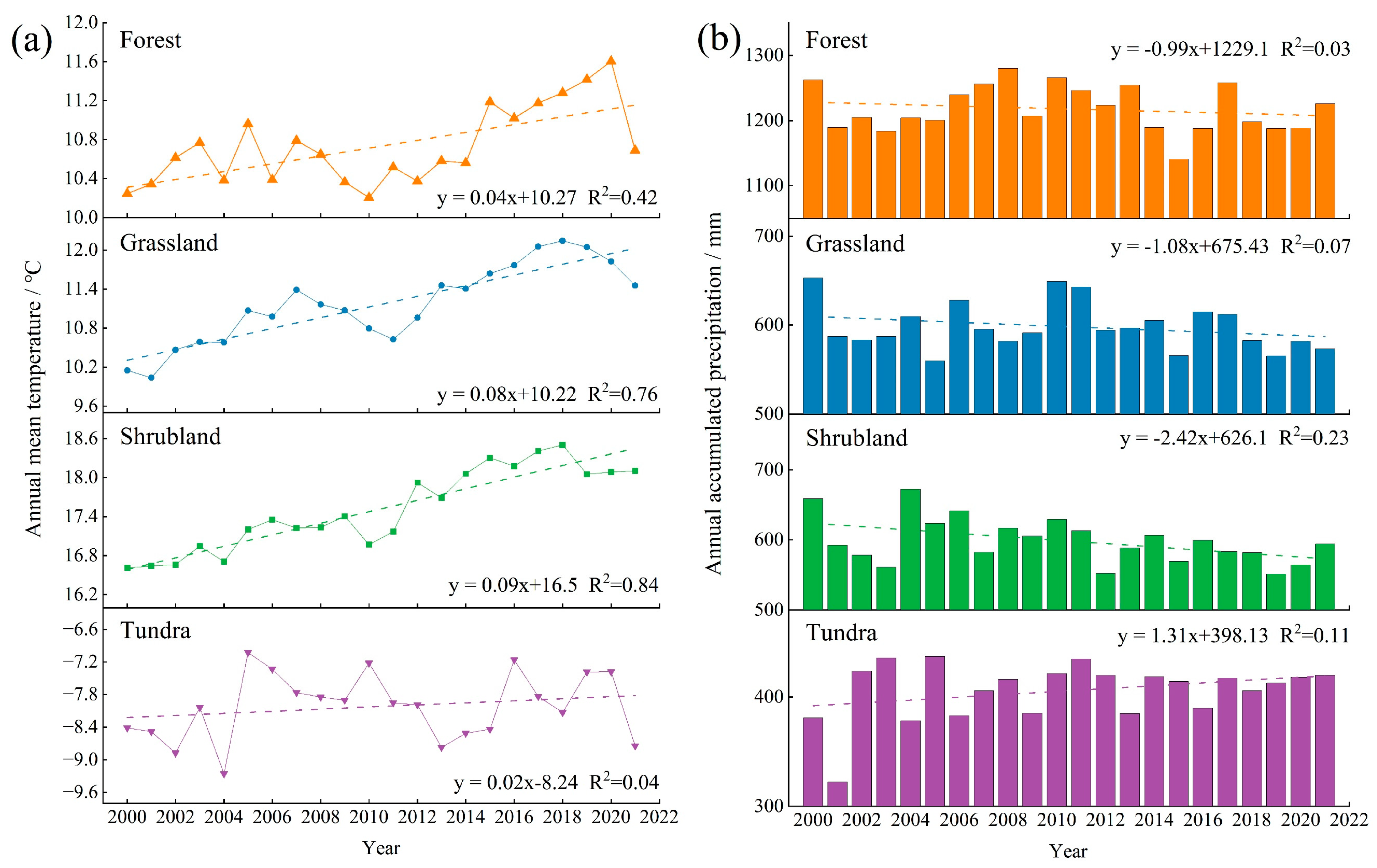
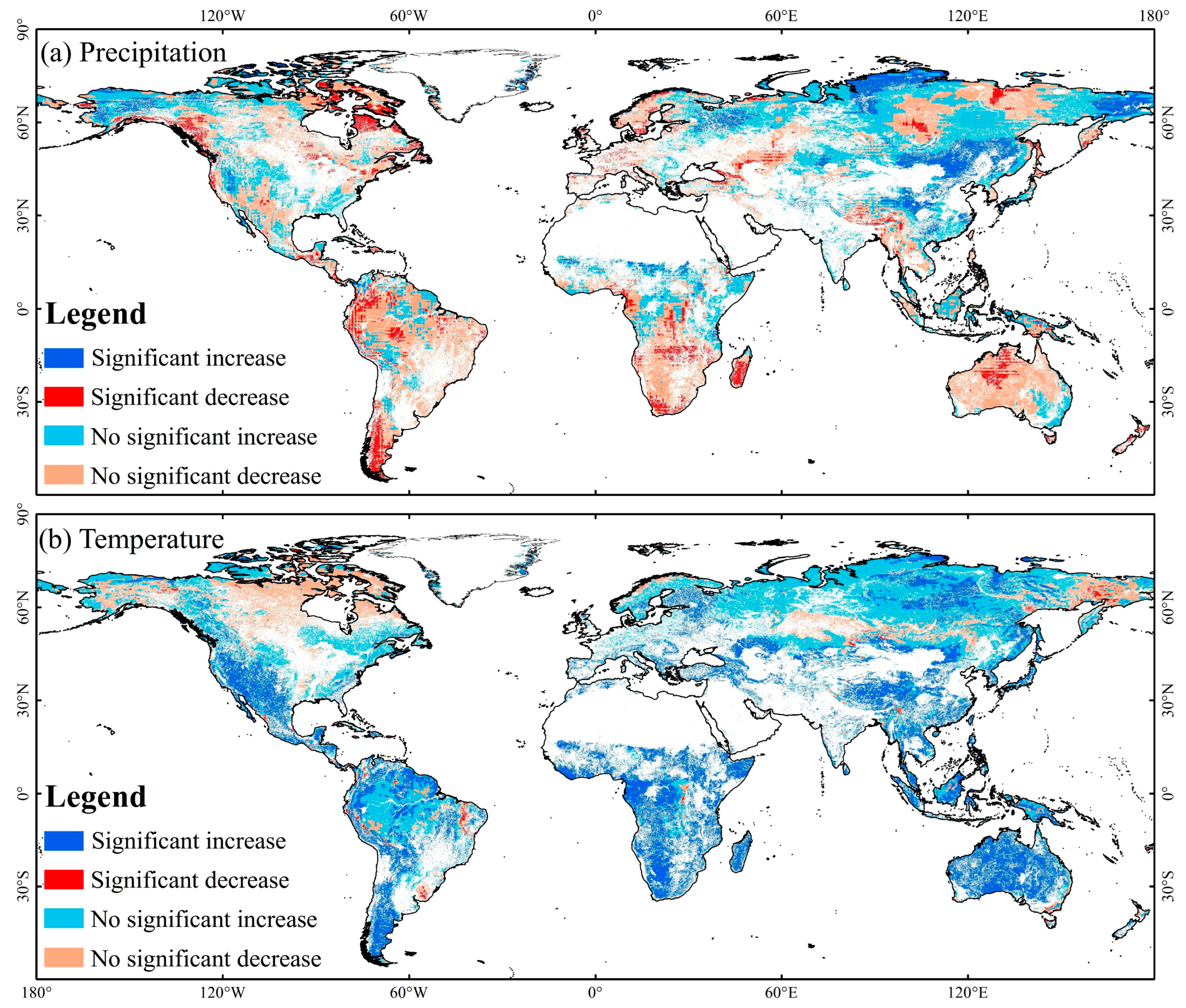
3.2.1. Spatiotemporal Variations of Temperature and Precipitation in FGST
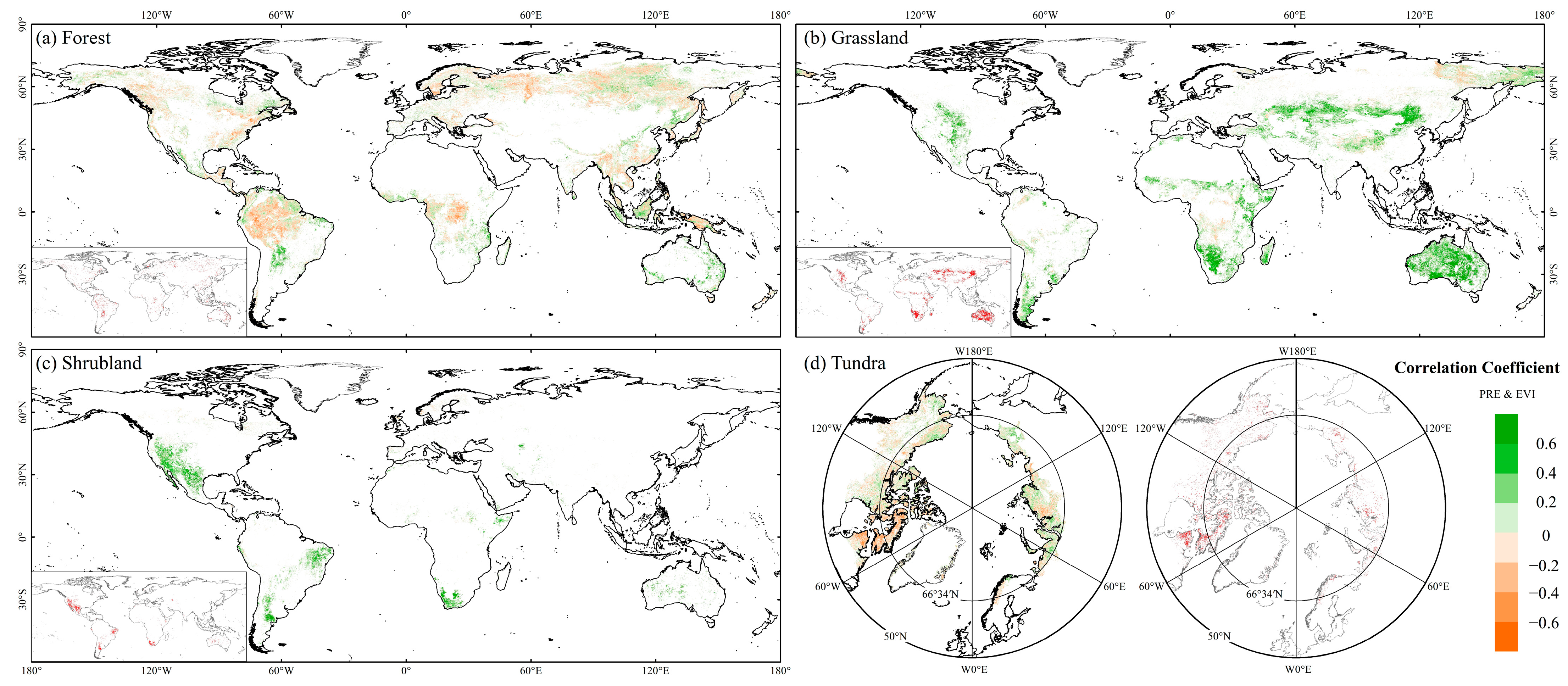
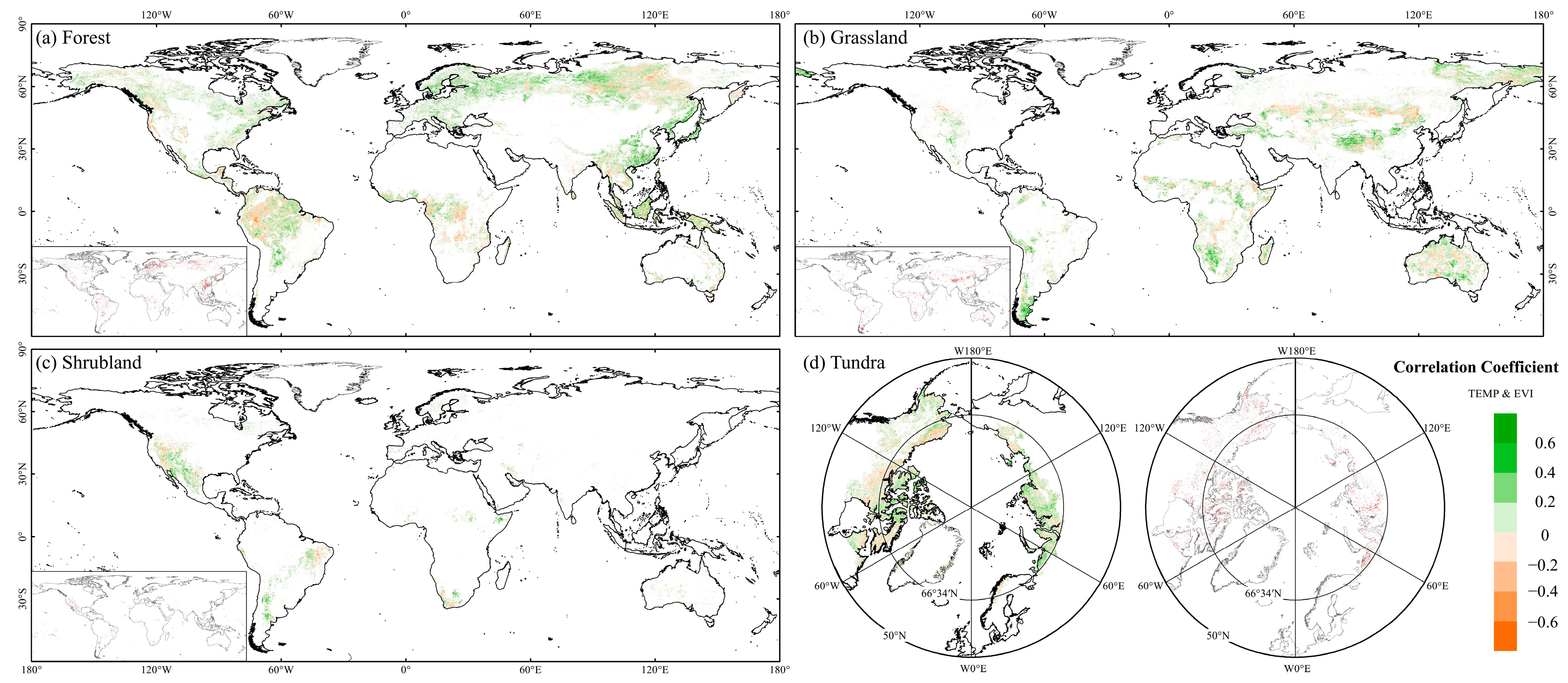
3.2.2. Effects of Climatic Factor Variations on EVI in FGST
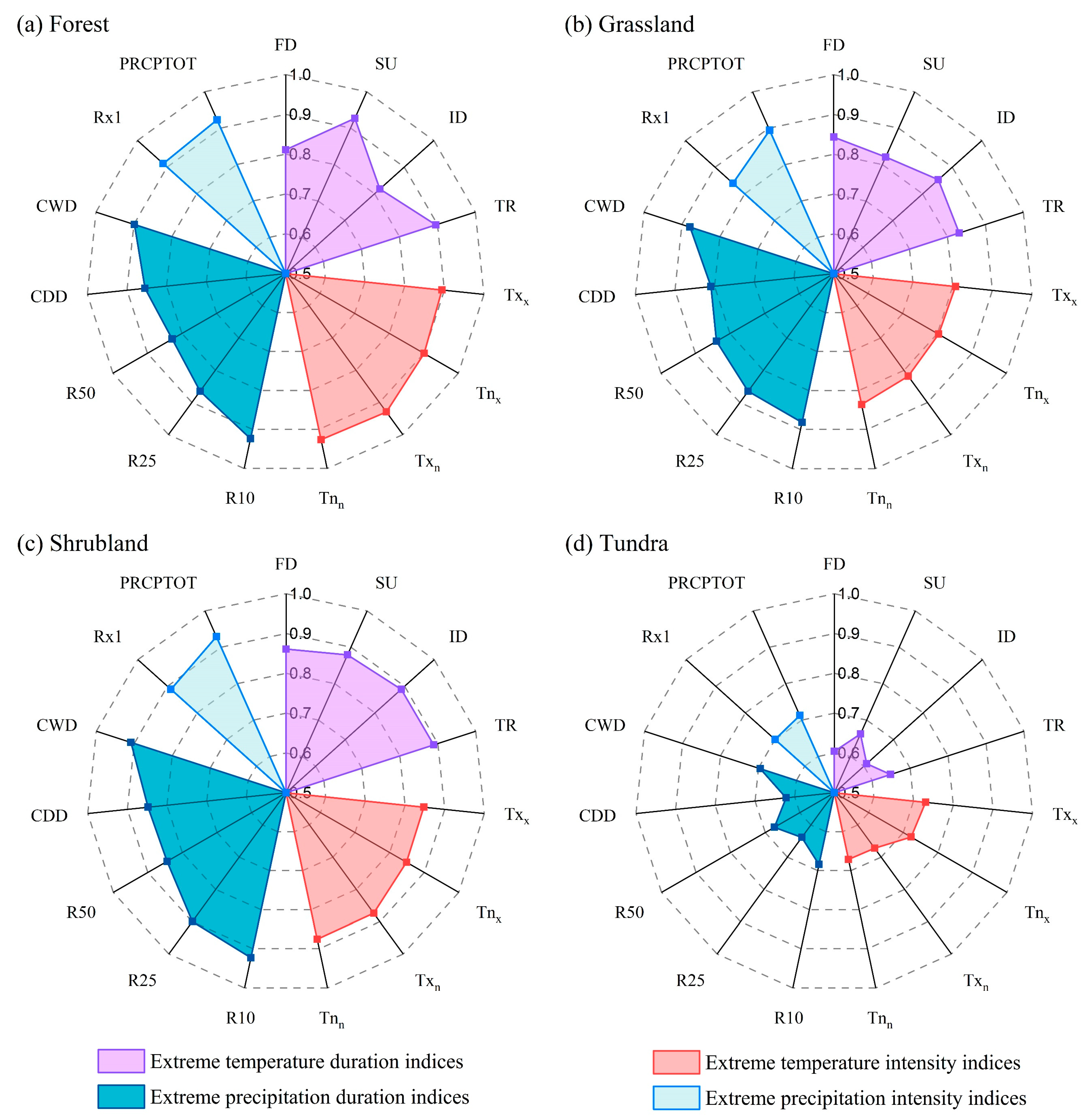
3.3. Association between EVI and Extreme Climate Indices in FGST
4. Discussion
4.1. Global FGST’s Spatiotemporal Variation of EVI and Its Response to Precipitation and Temperature
4.2. Responses of Different Types of Vegetation EVI to Extreme Events
4.3. Limitations of the Study and Further Directions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Code | Definition |
|---|---|---|
| Forest | 20 | It refers to the lands covered with trees, the top density of which occupies over 30%. Deciduous broadleaf forest, evergreen broadleaf forest, deciduous coniferous forest, evergreen coniferous forest, mixed forest, and sparse woodland, the top density of which covers 10–30%, are included in this category. It is mainly planted with crops and rarely with fruit trees or other trees that are not included in this category. |
| Grassland | 30 | It refers to the lands covered by natural grass with a cover density of over 10%. The prairies, meadow steppes, alpine grasslands, desert steppes, and lawns, etc., are included in this category. Cultivated pastures are not included in this category. |
| Shrubland | 40 | It refers to the lands covered with shrubs with a cover density of over 30%. Mountain shrubs, deciduous and evergreen shrubs, and desert jungle in desert areas with a cover density of over 10% are included in this category. Tea gardens, coffee gardens, and other economic croplands, etc., are not included in this category. |
| Tundra | 70 | It refers to the lands covered by lichen, moss, hardy perennial herbs, and shrubs in cold and high-altitude mountain areas. Shrub tundra, grass tundra, wet tundra, alpine tundra, and barren tundra, etc., are included in this category. |
| Classification | Slope | t |
|---|---|---|
| Significant increase | >0 | t > tα/2 |
| Significant decrease | <0 | t > tα/2 |
| No significant increase | >0 | t < tα/2 |
| No significant decrease | <0 | t < tα/2 |
| Category | Extreme Climate Index Name | Definition | Unit |
|---|---|---|---|
| Extreme temperature duration indices | Number of frost days, FD | Annual count of days when daily minimum temperature < 0 °C | d |
| Number of summer days, SU | Annual count of days when daily maximum temperature > 25 °C | d | |
| Number of icing days, ID | Annual count of days when daily maximum temperature < 0 °C | d | |
| Number of tropical nights, TR | Annual count of days when daily minimum temperature > 20 °C | d | |
| Extreme temperature intensity indices | Annual maximum value of daily maximum temperature, TXx | The maximum daily maximum temperature for each year | °C |
| Annual maximum value of daily minimum temperature, TNx | The maximum daily minimum temperature for each year | °C | |
| Annual minimum value of daily maximum temperature, TXn | The minimum daily maximum temperature for each year | °C | |
| Annual minimum value of daily minimum temperature, TNn | The minimum daily minimum temperature for each year | °C | |
| Extreme precipitation duration indices | Number of moderate rain days, R10mm | Annual count of days when the daily precipitation amount ≥ 10 mm | d |
| Number of heavy rain days, R25mm | Annual count of days when the daily precipitation amount ≥ 25 mm | d | |
| Number of torrential rain days, R50mm | Annual count of days when the daily precipitation amount ≥ 50 mm | d | |
| Maximum length of dry spell, CDD | The maximum number of consecutive days with the daily precipitation amount < 1 mm | d | |
| Maximum length of wet spell, CWD | The maximum number of consecutive days with the daily precipitation amount ≥ 1 mm | d | |
| Extreme precipitation intensity indices | Annual maximum value of daily maximum precipitation, Rx1 | The maximum daily maximum precipitation for each year | mm |
| Annual total precipitation in wet days, PRCPTOT | The sum of precipitation in a year when the daily precipitation amount ≥ 1 mm | mm |

References
- Feng, X.; Liu, C.; Xie, F.; Lu, J.; Chiu, L.S.; Tintera, G.; Chen, B. Precipitation Characteristic Changes Due to Global Warming in a High-Resolution (16 Km) ECMWF Simulation. Q. J. R. Meteorol. Soc. 2019, 145, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tian, Y.; Myneni, R.B.; Ciais, P.; Saatchi, S.; Liu, Y.Y.; Piao, S.; Chen, H.; Vermote, E.F.; Song, C.; et al. Widespread Decline of Congo Rainforest Greenness in the Past Decade. Nature 2014, 509, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, X.; Wilson, A.M.; Silander, J.A. Predicting Autumn Phenology: How Deciduous Tree Species Respond to Weather Stressors. Agric. For. Meteorol. 2018, 250–251, 127–137. [Google Scholar] [CrossRef]
- Qiu, T.; Song, C.; Clark, J.S.; Seyednasrollah, B.; Rathnayaka, N.; Li, J. Understanding the Continuous Phenological Development at Daily Time Step with a Bayesian Hierarchical Space-Time Model: Impacts of Climate Change and Extreme Weather Events. Remote Sens. Environ. 2020, 247, 111956. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and Its Drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.; Ciais, P.; Tømmervik, H.; et al. Characteristics, Drivers and Feedbacks of Global Greening. Nat. Rev. Earth Environ. 2020, 1, 14–27. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Primary Forests Are Irreplaceable for Sustaining Tropical Biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef]
- Leßmeister, A.; Bernhardt-Römermann, M.; Schumann, K.; Thiombiano, A.; Wittig, R.; Hahn, K. Vegetation Changes over the Past Two Decades in a West African Savanna Ecosystem. Appl. Veg. Sci. 2019, 22, 230–242. [Google Scholar] [CrossRef]
- Wu, L.; Chen, H.; Chen, D.; Wang, S.; Wu, Y.; Wang, B.; Liu, S.; Yue, L.; Yu, J.; Bai, Y. Soil Biota Diversity and Plant Diversity Both Contributed to Ecosystem Stability in Grasslands. Ecol. Lett. 2023, 26, 858–868. [Google Scholar] [CrossRef]
- Sheil, D. Forests, Atmospheric Water and an Uncertain Future: The New Biology of the Global Water Cycle. For. Ecosyst. 2018, 5, 19. [Google Scholar] [CrossRef]
- Gang, C.; Wang, Z.; Chen, Y.; Yang, Y.; Li, J.; Cheng, J.; Qi, J.; Odeh, I. Drought-Induced Dynamics of Carbon and Water Use Efficiency of Global Grasslands from 2000 to 2011. Ecol. Indic. 2016, 67, 788–797. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.-L.; Wu, H.; Zhou, C.; Liu, X.; Leng, P.; Yang, P.; Wu, W.; Tang, R.; Shang, G.-F.; et al. Biophysical Impacts of Earth Greening Can Substantially Mitigate Regional Land Surface Temperature Warming. Nat. Commun. 2023, 14, 121. [Google Scholar] [CrossRef]
- Zhang, Y.; Piao, S.; Sun, Y.; Rogers, B.M.; Li, X.; Lian, X.; Liu, Z.; Chen, A.; Peñuelas, J. Future Reversal of Warming-Enhanced Vegetation Productivity in the Northern Hemisphere. Nat. Clim. Chang. 2022, 12, 581–586. [Google Scholar] [CrossRef]
- Diepstraten, R.A.E.; Jessen, T.D.; Fauvelle, C.M.D.; Musiani, M.M. Does Climate Change and Plant Phenology Research Neglect the Arctic Tundra? Ecosphere 2018, 9, e02362. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, W.; Schwalm, C.R.; Gentine, P.; Smith, W.K.; Ciais, P.; Kimball, J.S.; Gazol, A.; Kannenberg, S.A.; Chen, A.; et al. Widespread Spring Phenology Effects on Drought Recovery of Northern Hemisphere Ecosystems. Nat. Clim. Chang. 2023, 13, 182–188. [Google Scholar] [CrossRef]
- Chen, J.; Shao, Z.; Deng, X.; Huang, X.; Dang, C. Vegetation as the Catalyst for Water Circulation on Global Terrestrial Ecosystem. Sci. Total Environ. 2023, 895, 165071. [Google Scholar] [CrossRef]
- Na, R.; Na, L.; Du, H.; He, H.S.; Shan, Y.; Zong, S.; Huang, L.; Yang, Y.; Wu, Z. Vegetation Greenness Variations and Response to Climate Change in the Arid and Semi-Arid Transition Zone of the Mongo-Lian Plateau during 1982–2015. Remote Sens. 2021, 13, 4066. [Google Scholar] [CrossRef]
- Cui, J.; Lian, X.; Huntingford, C.; Gimeno, L.; Wang, T.; Ding, J.; He, M.; Xu, H.; Chen, A.; Gentine, P.; et al. Global Water Availability Boosted by Vegetation-Driven Changes in Atmospheric Moisture Transport. Nat. Geosci. 2022, 15, 982–988. [Google Scholar] [CrossRef]
- Sun, Z.; Ouyang, Z.; Zhang, X.; Ren, W. A New Global Dataset of Phase Synchronization of Temperature and Precipitation: Its Climatology and Contribution to Global Vegetation Productivity. Geosci. Data J. 2019, 6, 126–136. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate Change Impacts and Adaptation in Forest Management: A Review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef]
- Wang, G.; Mang, S.L.; Riehl, B.; Huang, J.; Wang, G.; Xu, L.; Huang, K.; Innes, J. Climate Change Impacts and Forest Adaptation in the Asia–Pacific Region: From Regional Experts’ Perspectives. J. For. Res. 2019, 30, 277–293. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Wu, M.; Han, Y.; Xia, J.; Li, L.; Wan, S. Water-Mediated Responses of Ecosystem Carbon Fluxes to Climatic Change in a Temperate Steppe. New Phytol. 2008, 177, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Wei, J.-Z.; Pei, J.-Y.; Cheng, H.; Zhang, X.-L.; Huang, F.-Q.; Li, F.-M.; Ye, J.-S. Impacts of Climate Change and Human Activities on Grassland Vegetation Variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Cao, G.; Ma, Z.; Li, Y.; Zhang, F.; Zhao, X.; Zhao, X.; Jiang, L.; Sanders, N.J.; et al. Alpine Grassland Plants Grow Earlier and Faster but Biomass Remains Unchanged over 35 Years of Climate Change. Ecol. Lett. 2020, 23, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Gang, C.; Zhou, W.; Chen, Y.; Wang, Z.; Sun, Z.; Li, J.; Qi, J.; Odeh, I. Quantitative Assessment of the Contributions of Climate Change and Human Activities on Global Grassland Degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Henry, G.H.R.; Hollister, R.D.; Klanderud, K.; Björk, R.G.; Bjorkman, A.D.; Elphinstone, C.; Jónsdóttir, I.S.; Molau, U.; Petraglia, A.; Oberbauer, S.F.; et al. The International Tundra Experiment (ITEX): 30 Years of Research on Tundra Ecosystems. Arct. Sci. 2022, 8, 550–571. [Google Scholar] [CrossRef]
- May, J.L.; Hollister, R.D.; Betway, K.R.; Harris, J.A.; Tweedie, C.E.; Welker, J.M.; Gould, W.A.; Oberbauer, S.F. NDVI Changes Show Warming Increases the Length of the Green Season at Tundra Communities in Northern Alaska: A Fine-Scale Analysis. Front. Plant Sci. 2020, 11, 1174. [Google Scholar] [CrossRef]
- Field, C.B.; Barros, V.; Stocker, T.F.; Qin, D.; Dokken, D.J.; Ebi, K.L.; Mastrandrea, M.D.; Mach, K.J.; Plattner, G.-K.; Allen, S.K.; et al. (Eds.) Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation: Special Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2012; ISBN 978-1-107-02506-6. [Google Scholar]
- Diffenbaugh, N.S.; Singh, D.; Mankin, J.S.; Horton, D.E.; Swain, D.L.; Touma, D.; Charland, A.; Liu, Y.; Haugen, M.; Tsiang, M.; et al. Quantifying the Influence of Global Warming on Unprecedented Extreme Climate Events. Proc. Natl. Acad. Sci. USA 2017, 114, 4881–4886. [Google Scholar] [CrossRef]
- Sun, S.; Shi, P.; Zhang, Q.; Wang, J.; Wu, J.; Chen, D. Evolution of Future Precipitation Extremes: Viewpoint of Climate Change Classification. Int. J. Climatol. 2022, 42, 1220–1230. [Google Scholar] [CrossRef]
- Didan, K.; Munoz, A.B.; Solano, R.; Huete, A. 34.MODIS Vegetation Index User’s Guide (MOD13 Series); Version 3.1.; Vegetation Index and Phenology Lab, University of Arizona: Tucson, AZ, USA, 2015. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the Radiometric and Biophysical Performance of the MODIS Vegetation Indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Zhang, H.; Zhan, C.; Xia, J.; Yeh, P.J.-F. Responses of Vegetation to Changes in Terrestrial Water Storage and Temperature in Global Mountainous Regions. Sci. Total Environ. 2022, 851, 158416. [Google Scholar] [CrossRef]
- Yuan, Z.; Xu, J.J.; Chen, J.; Wang, Y.Q.; Yin, J. EVI Indicated Spatial-Temporal Variations in Vegetation and Their Responses to Climatic and Anthropogenic Factors in the Chinese Mainland Since 2000s. J. Environ. Inform. 2021, 40, 157–175. [Google Scholar] [CrossRef]
- Zhang, D.; Geng, X.; Chen, W.; Fang, L.; Yao, R.; Wang, X.; Zhou, X. Inconsistency of Global Vegetation Dynamics Driven by Climate Change: Evidences from Spatial Regression. Remote Sens. 2021, 13, 3442. [Google Scholar] [CrossRef]
- Sa, C.; Meng, F.; Luo, M.; Li, C.; Wang, M.; Adiya, S.; Bao, Y. Spatiotemporal Variation in Snow Cover and Its Effects on Grassland Phenology on the Mongolian Plateau. J. Arid Land 2021, 13, 332–349. [Google Scholar] [CrossRef]
- Na, L.; Na, R.; Zhang, J.; Tong, S.; Shan, Y.; Ying, H.; Li, X.; Bao, Y. Vegetation Dynamics and Diverse Responses to Extreme Climate Events in Different Vegetation Types of Inner Mongolia. Atmosphere 2018, 9, 394. [Google Scholar] [CrossRef]
- Kazemzadeh, M.; Hashemi, H.; Jamali, S.; Uvo, C.B.; Berndtsson, R.; Huffman, G.J. Linear and Nonlinear Trend Analyzes in Global Satellite-Based Precipitation, 1998–2017. Earths Future 2021, 9, e2020EF001835. [Google Scholar] [CrossRef]
- Fan, M.; Xu, J.; Yu, W.; Chen, Y.; Wang, M.; Dai, W.; Wang, Y. Recent Tianshan Warming in Relation to Large-Scale Climate Teleconnections. Sci. Total Environ. 2023, 856, 159201. [Google Scholar] [CrossRef]
- Lafleur, P.M.; Humphreys, E.R. Tundra Shrub Effects on Growing Season Energy and Carbon Dioxide Exchange. Environ. Res. Lett. 2018, 13, 055001. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Fang, G.; Xia, Z.; Yang, Y.; Duan, W.; Xia, Q.; Li, S. Future Changes in Extreme Precipitation from 1.0 °C More Warming in the Tienshan Mountains, Central Asia. J. Hydrol. 2022, 612, 128269. [Google Scholar] [CrossRef]
- Isenlin COVID-19 Has a Major Impact on World Forestry. Available online: http://www.isenlin.cn/sf_2F72B31295974CDE929EFBFAB0CBFF2F_209_43659777456.html (accessed on 1 June 2023).
- Su, F.; Fu, D.; Yan, F.; Xiao, H.; Pan, T.; Xiao, Y.; Kang, L.; Zhou, C.; Meadows, M.; Lyne, V.; et al. Rapid Greening Response of China’s 2020 Spring Vegetation to COVID-19 Restrictions: Implications for Climate Change. Sci. Adv. 2021, 7, eabe8044. [Google Scholar] [CrossRef]
- Deb, J.; Phinn, S.; Butt, N.; McAlpine, C. Climate Change Impacts on Tropical Forests: Identifying Risks for Tropical Asia. J. Trop. For. Sci. 2018, 30, 182–194. [Google Scholar]
- Chang, J.; Tian, J.; Zhang, Z.; Chen, X.; Chen, Y.; Chen, S.; Duan, Z. Changes of Grassland Rain Use Efficiency and NDVI in Northwestern China from 1982 to 2013 and Its Response to Climate Change. Water 2018, 10, 1689. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Ju, W.; Ruan, H.; Qin, Z.; Huang, Y.; Jeelani, N.; Padarian, J.; Propastin, P. Quantitative Assessments of Water-Use Efficiency in Temperate Eurasian Steppe along an Aridity Gradient. PLoS ONE 2017, 12, e0179875. [Google Scholar] [CrossRef]
- Zhang, Z.; Chang, J.; Xu, C.-Y.; Zhou, Y.; Wu, Y.; Chen, X.; Jiang, S.; Duan, Z. The Response of Lake Area and Vegetation Cover Variations to Climate Change over the Qinghai-Tibetan Plateau during the Past 30 years. Sci. Total Environ. 2018, 635, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Sala, O.E.; Lauenroth, W.K. Small Rainfall Events: An Ecological Role in Semiarid Regions. Oecologia 1982, 53, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Schwinning, S.; Sala, O.E.; Loik, M.E.; Ehleringer, J.R. Thresholds, Memory, and Seasonality: Understanding Pulse Dynamics in Arid/Semi-Arid Ecosystems. Oecologia 2004, 141, 191–193. [Google Scholar] [CrossRef]
- Song, Y.; Lu, Y.; Guo, Z.; Xu, X.; Liu, T.; Wang, J.; Wang, W.; Hao, W.; Wang, J. Variations in Soil Water Content and Evapotranspiration in Relation to Precipitation Pulses within Desert Steppe in Inner Mongolia, China. Water 2019, 11, 198. [Google Scholar] [CrossRef]
- Dou, W.; Xiao, B.; Yao, X.; Kidron, G.J. Asymmetric Responses of Biocrust Respiration to Precipitation Manipulation under a Changing Semiarid Climate. Geoderma 2023, 430, 116318. [Google Scholar] [CrossRef]
- Sponseller, R.A.; Hall, S.J.; Huber, D.P.; Grimm, N.B.; Kaye, J.P.; Clark, C.M.; Collins, S.L. Variation in Monsoon Precipitation Drives Spatial and Temporal Patterns of Larrea Tridentata Growth in the Sonoran Desert. Funct. Ecol. 2012, 26, 750–758. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, X.; Li, H.; Guo, L.; Lv, T.; Wu, P. Exotic Shrub Species (Caragana Korshinskii) Is More Resistant to Extreme Natural Drought than Native Species (Artemisia Gmelinii) in a Semiarid Revegetated Ecosystem. Agric. For. Meteorol. 2018, 263, 207–216. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Mommer, L.; Sabatini, F.M.; Bruelheide, H.; Kuyper, T.W.; McCormack, M.L.; Bergmann, J.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Iversen, C.M.; et al. Root Traits Explain Plant Species Distributions along Climatic Gradients yet Challenge the Nature of Ecological Trade-Offs. Nat. Ecol. Evol. 2021, 5, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Fensholt, R.; Verbesselt, J.; Grogan, K.; Horion, S.; Wang, Y. Evaluating Temporal Consistency of Long-Term Global NDVI Datasets for Trend Analysis. Remote Sens. Environ. 2015, 163, 326–340. [Google Scholar] [CrossRef]
- Lyapustin, A.; Wang, Y.; Xiong, X.; Meister, G.; Platnick, S.; Levy, R.; Franz, B.; Korkin, S.; Hilker, T.; Tucker, J.; et al. Scientific Impact of MODIS C5 Calibration Degradation and C6+ Improvements. Atmos. Meas. Tech. 2014, 7, 4353–4365. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Song, Y.; Qin, T.; Yan, D.; Zhang, X.; Zhu, L.; Dorjsuren, B.; Khalid, H. Spatiotemporal Variations of Global Terrestrial Typical Vegetation EVI and Their Responses to Climate Change from 2000 to 2021. Remote Sens. 2023, 15, 4245. https://doi.org/10.3390/rs15174245
Li C, Song Y, Qin T, Yan D, Zhang X, Zhu L, Dorjsuren B, Khalid H. Spatiotemporal Variations of Global Terrestrial Typical Vegetation EVI and Their Responses to Climate Change from 2000 to 2021. Remote Sensing. 2023; 15(17):4245. https://doi.org/10.3390/rs15174245
Chicago/Turabian StyleLi, Chenhao, Yifan Song, Tianling Qin, Denghua Yan, Xin Zhang, Lin Zhu, Batsuren Dorjsuren, and Hira Khalid. 2023. "Spatiotemporal Variations of Global Terrestrial Typical Vegetation EVI and Their Responses to Climate Change from 2000 to 2021" Remote Sensing 15, no. 17: 4245. https://doi.org/10.3390/rs15174245
APA StyleLi, C., Song, Y., Qin, T., Yan, D., Zhang, X., Zhu, L., Dorjsuren, B., & Khalid, H. (2023). Spatiotemporal Variations of Global Terrestrial Typical Vegetation EVI and Their Responses to Climate Change from 2000 to 2021. Remote Sensing, 15(17), 4245. https://doi.org/10.3390/rs15174245