





Unmanned Aerial Vehicle (UAV) for Detection and Prediction of Damage Caused by Potato Cyst Nematode G. pallida on Selected Potato Cultivars

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling and Estimating Initial Population Densities (Pi)
2.3. Potato Cultivars
2.4. Field Management (Planting to Harvest)
2.5. Data Analysis and Modelling
2.5.1. The First Dataset
- y = fresh tuber yield in tons (ha)−1;
- Ymax = yield at Pi = 0 in tons (ha)−1;
- m = relative minimum yield when Pi → ∞;
- T = tolerance limit, the densityin number of eggs (g dry soil)−1 above which yield starts to decline.
2.5.2. UAV Data Collection
- Ccpt = Canopy temperature (°C);
- acpt = Maximum rate of canopy temperature increase;
- Mcpt = Maximum canopy temperature reached (°C);
- = Initial population density in number of eggs (g dry soil)−1.
3. Results and Discussion
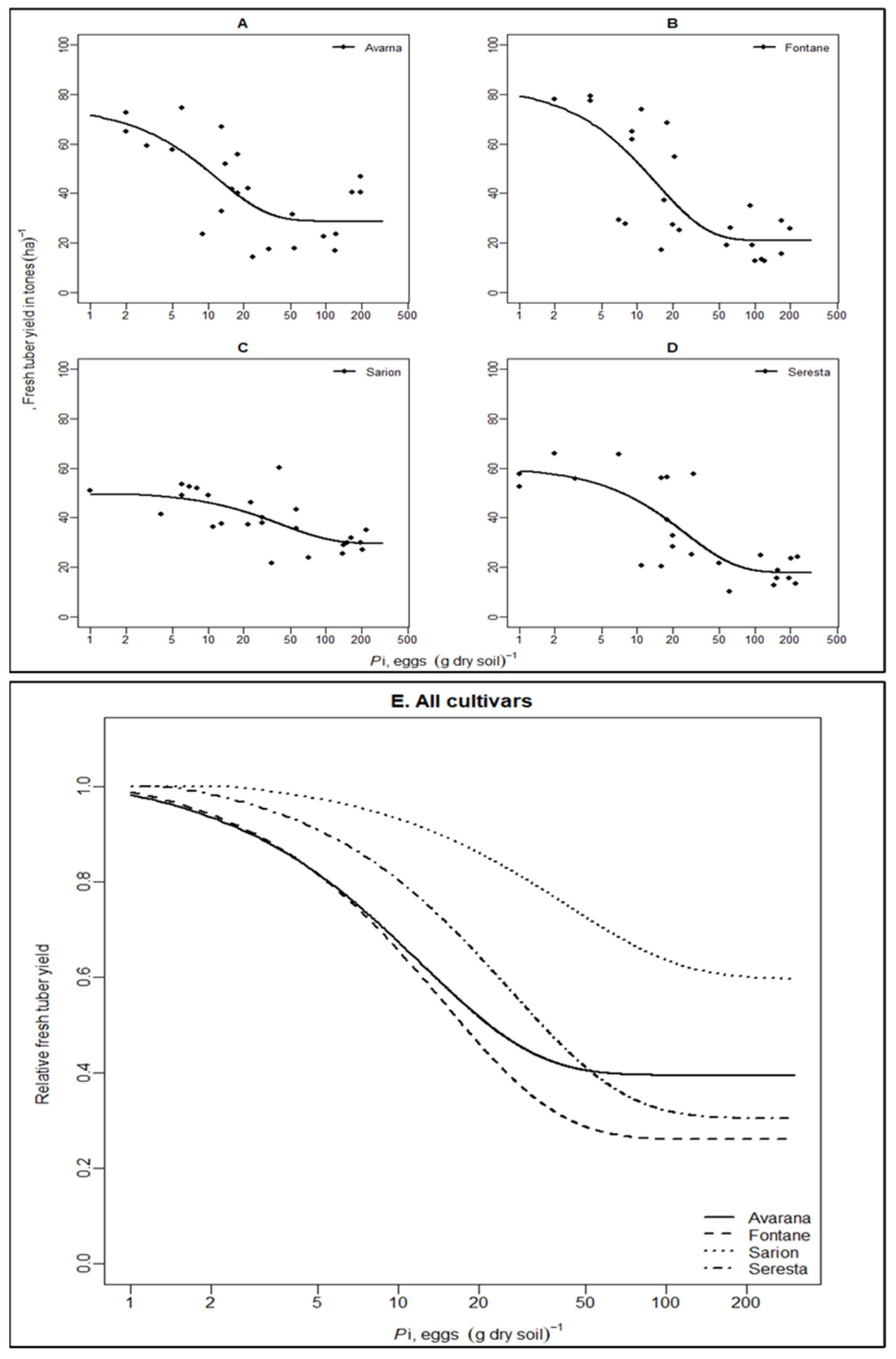
3.1. The Relationship between Pre-Plant Nematodes Densities (Pi) and Yield
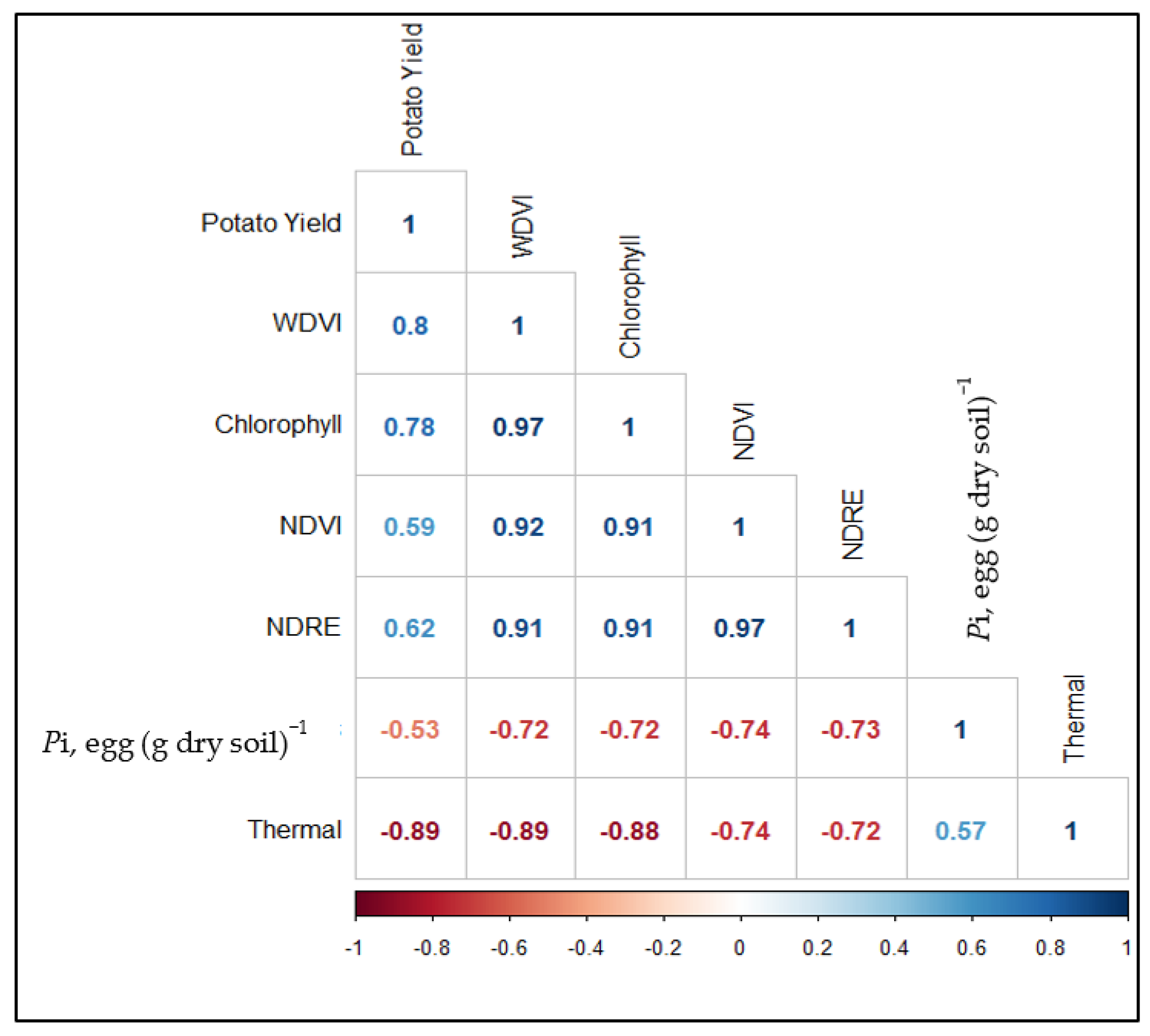
3.2. Correlation Matrix
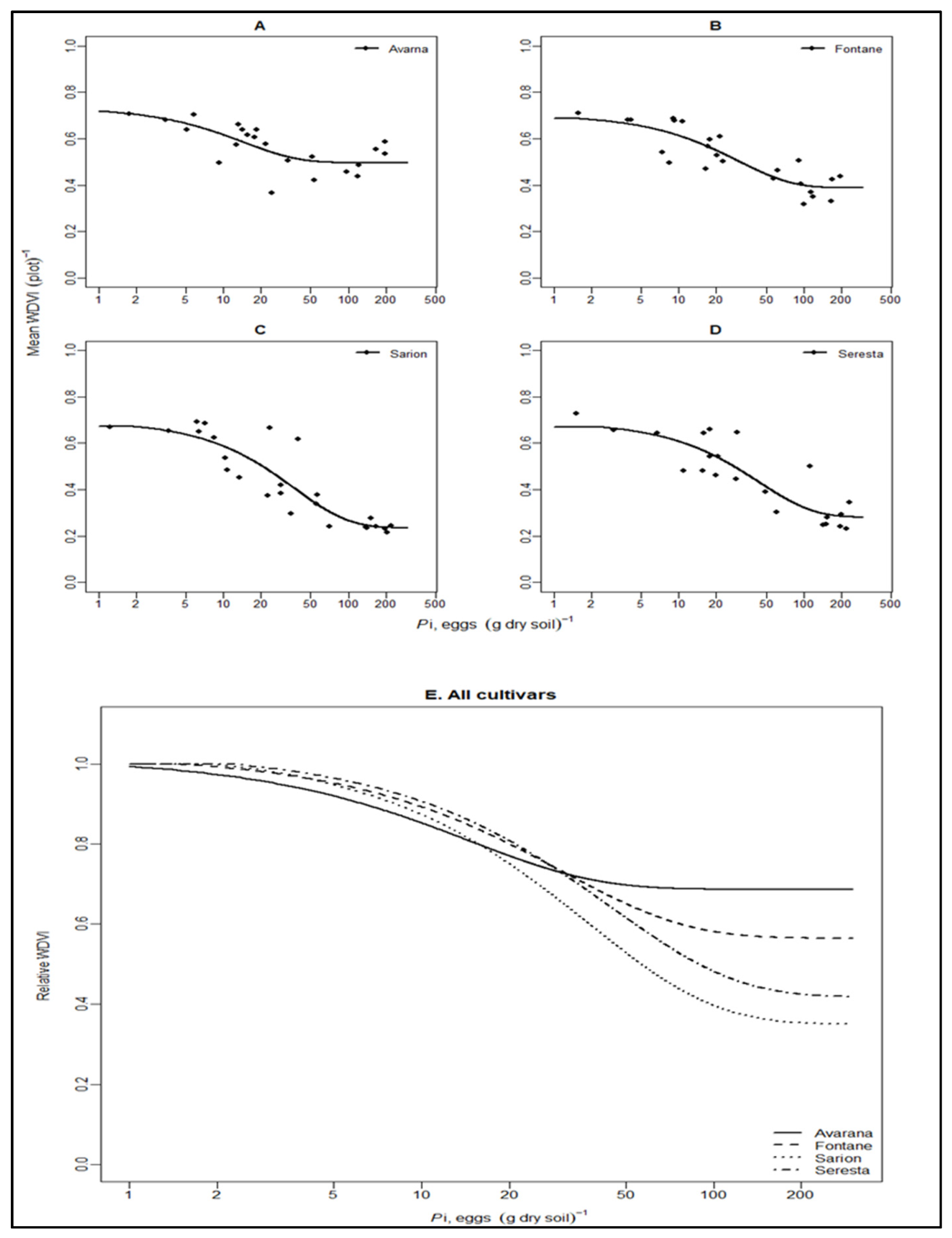
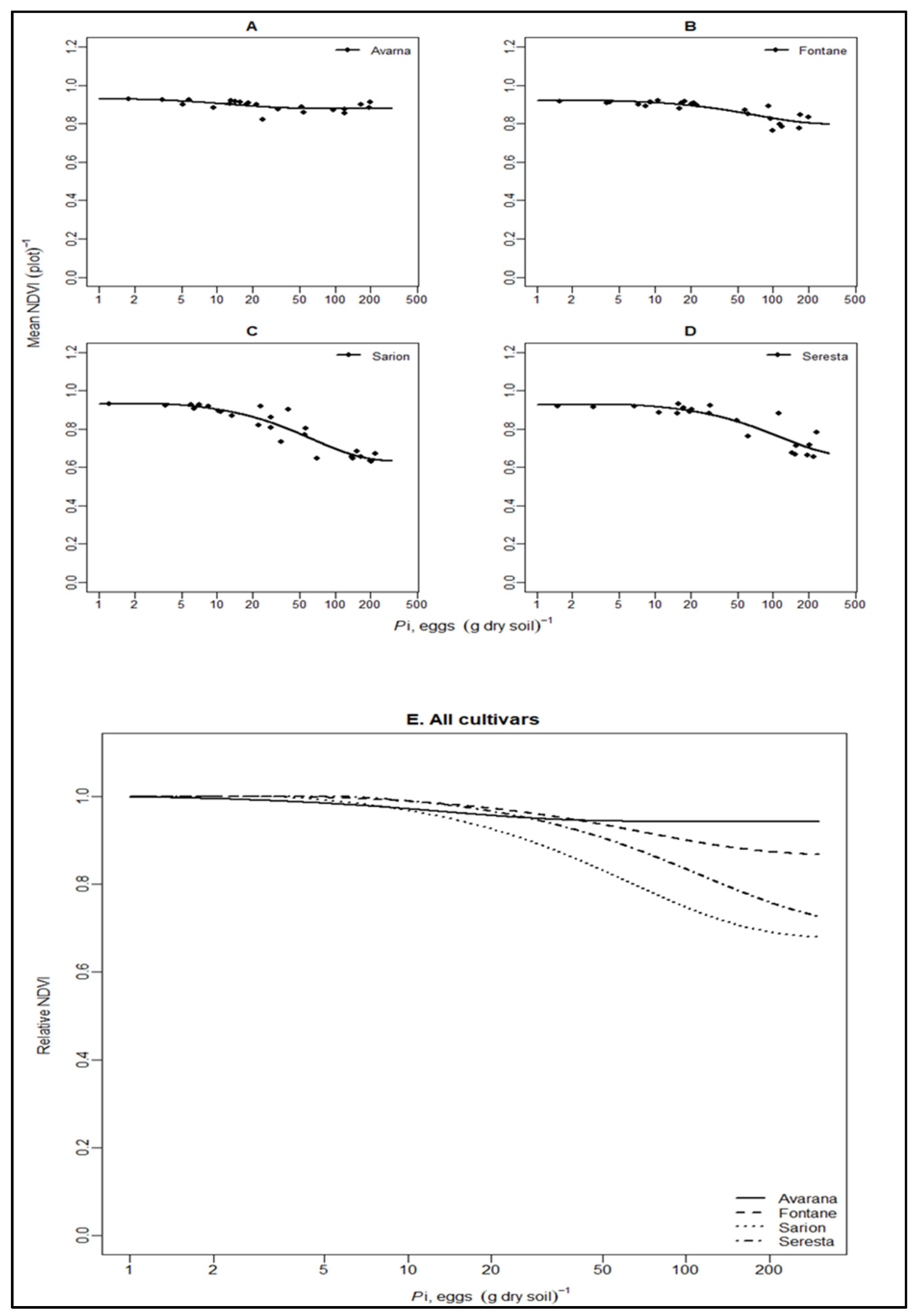
3.3. Relationship between Field Data and UAV Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Been, T.H.; Schomaker, C.H. Quantitative Studies on the Management of Potato Cyst Nematodes (Globodera spp.) in The Netherlands. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 1998. [Google Scholar]
- Teklu, M.G.; Schomaker, C.H.; Been, T.H.; Molendijk, L.P. Tuber Yield, Quality and Infestation Levels of Potato Genotypes, Resistant to the Root-Knot Nematode, Meloidogyne chitwoodi. Potato Res. 2022, 66, 105–135. [Google Scholar] [CrossRef]
- Seinhorst, J. Water Consumption of Plants Attacked By Nematodes and Mechanisms of Growth Reduction. Nematologica 1981, 27, 34–51. [Google Scholar] [CrossRef] [Green Version]
- Seinhorst, J.W. Effect of nematode attack the growth and yield of crop plants. In Cyst Nematodes; Lamberti, F., Taylor, C.E., Eds.; Plenum Press: New York, NY, USA, 1986; pp. 191–210. [Google Scholar]
- Gartner, U.; Hein, I.; Brown, L.H.; Chen, X.; Mantelin, S.; Sharma, S.K.; Dandurand, L.-M.; Kuhl, J.C.; Jones, J.T.; Bryan, G.J.; et al. Resisting Potato Cyst Nematodes With Resistance. Front. Plant Sci. 2021, 12, 661194. [Google Scholar] [CrossRef] [PubMed]
- Asano, K.; Kobayashi, A.; Tsuda, S.; Nishinaka, M.; Tamiya, S. DNA marker-assisted evaluation of potato genotypes for potential resistance to potato cyst nematode pathotypes not yet invading into Japan. Breed. Sci. 2012, 62, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotton, J.A.; Lilley, C.J.; Jones, L.M.; Kikuchi, T.; Reid, A.J.; Thorpe, P.; Tsai, I.J.; Beasley, H.; Blok, V.; Cock, P.J.A.; et al. The genome and life-stage specific transcriptomes of Globodera pallida elucidate key aspects of plant parasitism by a cyst nematode. Genome Biol. 2014, 15, R43. [Google Scholar] [CrossRef] [Green Version]
- Ochola, J.; Cortada, L.; Ng’Ang’A, M.; Hassanali, A.; Coyne, D.; Torto, B. Mediation of Potato–Potato Cyst Nematode, G. rostochiensis Interaction by Specific Root Exudate Compounds. Front. Plant Sci. 2020, 11, 649. [Google Scholar] [CrossRef]
- Hockland, S.; Niere, B.; Grenier, E.; Blok, V.; Phillips, M.; Den Nijs, L.; Anthoine, G.; Pickup, J.; Viaene, N. An evaluation of the implications of virulence in non-European populations of Globodera pallida and G. rostochiensis for potato cultivation in Europe. Nematology 2012, 14, 1–13. [Google Scholar] [CrossRef]
- Mburu, H.; Cortada, L.; Haukeland, S.; Ronno, W.; Nyongesa, M.; Kinyua, Z.; Bargul, J.L.; Coyne, D. Potato Cyst Nematodes: A New Threat to Potato Production in East Africa. Front. Plant Sci. 2020, 11, 670. [Google Scholar] [CrossRef]
- Zasada, I.A.; Ingham, R.E.; Baker, H.; Phillips, W.S. Impact of Globodera ellingtonae on yield of potato (Solanum tuberosum). J. Nematol. 2019, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- EPPO. PM 9/26(1) National regulatory control system for Globodera pallida and Globodera rostochiensis. EPPO Bull. 2018, 48, 516–532. [Google Scholar] [CrossRef] [Green Version]
- Hillnhütter, C.; Schweizer, A.; Kühnhold, V.; Sikora, R.A. Remote Sensing for the Detection of Soil-Borne Plant Parasitic Nematodes and Fungal Pathogens. In Precision Crop Protection–The Challenge and Use of Heterogeneity; Oerke, E.C., Gerhards, R., Menz, G., Sikora, R., Eds.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Polder, G.; Blok, P.M.; De Villiers, H.A.C.; Van Der Wolf, J.M.; Kamp, J. Potato Virus Y Detection in Seed Potatoes Using Deep Learning on Hyperspectral Images. Front. Plant Sci. 2019, 10, 209. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, R.; Tsuda, S.; Tamiya, S.; Itoh, A.; Nishiwaki, K.; Murakami, N.; Shibuya, Y.; Hirafuji, M.; Nuske, S. Field phenotyping system for the assessment of potato late blight resistance using RGB imagery from an unmanned aerial vehicle. Biosyst. Eng. 2016, 148, 1–10. [Google Scholar] [CrossRef]
- Siebring, J.; Valente, J.; Domingues Franceschini, M.H.; Kamp, J.; Kooistra, L. Object-Based Image Analysis Applied to Low Altitude Aerial Imagery for Potato Plant Trait Retrieval and Pathogen Detection. Sensors 2019, 19, 5477. [Google Scholar] [CrossRef] [Green Version]
- Overstreet, C.; McGawley, E.C.; Khalilian, A.; Kirkpatrick, T.L.; Monfort, W.S.; Henderson, W.; Mueller, J.D. Site specific nematode management-development and success in cotton production in the United States. J. Nematol. 2014, 46, 309–320. [Google Scholar]
- Ortiz, V.; Phelan, S.; Mullins, E. A temporal assessment of nematode community structure and diversity in the rhizosphere of cisgenic Phytophthora infestans-resistant potatoes. BMC Ecol. 2016, 16, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Joalland, S.; Screpanti, C.; Varella, H.V.; Reuther, M.; Schwind, M.; Lang, C.; Walter, A.; Liebisch, F. Aerial and Ground Based Sensing of Tolerance to Beet Cyst Nematode in Sugar Beet. Remote Sens. 2018, 10, 787. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.J.; Assis, G.A.; Guizilini, V.; Faria, E.R.; Souza, J.R. Segmenting and detecting nematode in coffee crops using aerial images. In Computer Vision Systems. ICVS 2019; Tzovaras, D., Giakoumis, D., Vincze, M., Argyros, A., Eds.; Springer: Cham, Switzerland, 2014; Volume 11754. [Google Scholar] [CrossRef]
- Arjoune, Y.; Sugunaraj, N.; Peri, S.; Nair, S.V.; Skurdal, A.; Ranganathan, P.; Johnson, B. Soybean cyst nematode detection and management: A review. Plant Methods 2022, 18, 110. [Google Scholar] [CrossRef]
- Shao, H.; Zhang, P.; Peng, D.; Huang, W.; Kong, L.; Li, C.; Liu, E.; Peng, H. Current advances in the identification ofplant nematode diseases: From lab assays to in-field diagnostics. Front. Plant Sci. 2023, 14, 1106784. [Google Scholar] [CrossRef]
- Teklu, M.G.; Schomaker, C.H.; Been, T.H. Relative susceptibilities of five fodder radish varieties (Raphanus sativus var. Oleiformis) to Meloidogyne chitwoodi. Nematology 2014, 16, 577–590. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS Third ERTS-1 Symposium; NASA: Washington, DC, USA, 1974; pp. 309–317.
- Gong, P.; Pu, R.; Biging, G.S.; Larrieu, M.R. Estimation of forest leaf area index using vegetation indices derived from Hyperion hyperspectral data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1355–1362. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Vina, A.; Ciganda, V.; Rundquist, D.C.; Arkebauer, T.J. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, L08403. [Google Scholar] [CrossRef] [Green Version]
- Pineda, M.; Barón, M.; Pérez-Bueno, M.-L. Thermal Imaging for Plant Stress Detection and Phenotyping. Remote Sens. 2021, 13, 68. [Google Scholar] [CrossRef]
- Shakeel, Q.; Bajwa, R.T.; Rashid, I.; Aslam, H.M.U.; Iftikhar, Y.; Mubeen, M.; Li, G.; Wu, M. Concept and application of infrared thermography for plant disease measurement. In Trends in Plant Disease Assessment; Ul Haq, I., Ijaz, S., Eds.; Springer: Singapore, 2022. [Google Scholar] [CrossRef]
- Melandri, G.; Prashar, A.; McCouch, S.R.; van der Linden, G.; Jones, H.C.; Kadam, N.; Jagadish, K.; Bouwmeester, H.; Ruyter-Spira, C. Association mapping and genetic dissection of drought-induced canopy temperature differences in rice. J. Exp. Bot. 2020, 71, 1614–1627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Niu, Y.; Zhang, H.; Han, W.; Li, G.; Tang, J.; Peng, X. Maize Canopy Temperature Extracted From UAV Thermal and RGB Imagery and Its Application in Water Stress Monitoring. Front. Plant Sci. 2019, 10, 1270. [Google Scholar] [CrossRef]
- Ludovisi, R.; Tauro, F.; Salvati, R.; Khoury, S.; Scarascia Mugnozzaa, G.; Harfouche, A. UAV-Based Thermal Imaging for High-Throughput Field Phenotyping of Black Poplar Response to Drought. Front. Plant Sci. 2017, 8, 1681. [Google Scholar] [CrossRef]
- Vannoppen, A.; Gobin, A. Estimating yield from NDVI, weather data, and soil water depletion for sugar beet and potato in Northern Belgium. Water 2022, 14, 1188. [Google Scholar] [CrossRef]
- Newton, I.H.; Islam, T.; Saiful, I. Yield Prediction Model for Potato Using Landsat Time Series Images Driven Vegetation Indices. Remote Sens. Earth Syst. Sci. 2018, 1, 29–38. [Google Scholar] [CrossRef]
- Sun, C.; Zhou, J.; Ma, Y.; Xu, Y.; Pan, B.; Zhang, Z. A review of remote sensing for potato traits characterization in precision agriculture. Front. Plant Sci. 2022, 13, 871859. [Google Scholar] [CrossRef]
- Plumblee, M.T.; Mueller, J.D. Implementing precision agriculture concepts and technologies into crop production and site-specific management of nematodes. In Integrated Nematode Management: State-of-the-Art and Visions for the Future; CABI Books; CABI International: Wallingford, UK, 2021. [Google Scholar] [CrossRef]
- Gorny, A.M.; Wang, X.; Hay, F.S.; Pethybridge, S.J. Development of a species-specific PCR for detection and quantification of Meloidogyne hapla in soil using the 16D10 root-knot nematode effector gene. Plant Dis. 2019, 103, 1902–1909. [Google Scholar] [CrossRef]
- Takemoto, S.; Niwa, S.; Okada, H. Effect of storage temperature on soil nematode community structures as revealed by PCR-DGGE. J. Nematol. 2010, 42, 324. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Question | Technology Utilized | References |
|---|---|---|
| Detecting potato Y-virus in seed potatoes | Hyperspectral images | [14] |
| Assessment of potato late blight disease | Red-Green-Blue (RGB) images | [15] |
| Detecting Erwinia bacteria and potato Y-virus | RGB very high-resolution imagery | [16] |
| Cultivars | m | T | Ymax | sem | seT | seYmax | R2 | df | LSDm | LSDT | LSDYmax |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Avarna | 0.40 | 0.62 | 73.92 | 0.09 | 0.31 | 11.92 | 0.48 | 21 | 0.26 | 1.63 | 25.78 |
| Fontane | 0.26 | 0.76 | 80.36 | 0.09 | 0.42 | 17.07 | 0.56 | 22 | 0.25 * | 1.68 | 35.64 |
| Sarion | 0.60 | 2.16 | 49.48 | 0.09 | 1.58 | 4.65 | 0.44 | 23 | - | - | - |
| Serresta | 0.31 | 1.33 | 58.51 | 0.09 | 0.62 | 7.48 | 0.59 | 22 | 0.26 * | 1.59 | 17.73 |
| Cultivars | acpt | Mcpt | seacpt | seMcpt | R2 | df | LSDacpt | LSDMcpt |
|---|---|---|---|---|---|---|---|---|
| Avarna | 47.12 | 29.99 | 12.1 | 0.62 | 0.45 | 20 | 0.963 | 0.071 |
| Fontane | 52.55 | 30.2 | 16.6 | 0.72 | 0.32 | 22 | 1.026 | 0.075 |
| Sarion | 64.29 | 30.4 | 27.1 | 0.86 | 0.2 | 23 | - | - |
| Seresta | 46.78 | 31.11 | 13.9 | 0.77 | 0.39 | 20 | 1.006 | 0.076 |
| Cultivars | Variable | m | T | Ymax | sem | seT | seYmax | R2 | N | df | LSDm | LSDT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Avarna | FTWYield | 0.40 | 0.62 | 73.92 | 0.09 | 0.31 | 11.92 | 0.48 | 24 | 21 | - | - |
| Avarna | WDVI | 0.69 | 0.75 | 0.72 | 0.07 | 0.38 | 0.07 | 0.46 | 23 | 20 | 0.24 * | 1.35 |
| Avarna | Chlorophyl | 0.60 | 0.62 | 26.31 | 0.08 | 0.32 | 3.29 | 0.45 | 23 | 20 | 0.26 | 1.37 |
| Avarna | NDRE | 0.95 | 1.05 | 0.59 | 0.04 | 1.11 | 0.02 | 0.06 | 23 | 20 | 0.20 * | 1.99 |
| Avarna | NDVI | 0.94 | 0.69 | 0.93 | 0.02 | 0.44 | 0.02 | 0.29 | 23 | 20 | 0.20 * | 1.51 |
| Fontane | FTWYield | 0.26 | 0.76 | 80.36 | 0.09 | 0.42 | 17.07 | 0.56 | 25 | 22 | - | - |
| Fontane | WDVI | 0.56 | 1.54 | 0.69 | 0.05 | 0.77 | 0.05 | 0.74 | 25 | 22 | 0.20 * | 1.42 |
| Fontane | Chlorophyl | 0.49 | 2.91 | 21.85 | 0.09 | 1.54 | 1.30 | 0.73 | 25 | 22 | 0.25 | 1.45 |
| Fontane | NDRE | 0.80 | 2.70 | 0.58 | 0.03 | 1.30 | 0.01 | 0.72 | 25 | 22 | 0.19 * | 1.40 |
| Fontane | NDVI | 0.86 | 3.65 | 0.92 | 0.03 | 2.09 | 0.01 | 0.70 | 25 | 22 | 0.18 * | 1.50 * |
| Sarion | FTWYield | 0.60 | 2.16 | 49.48 | 0.09 | 1.58 | 4.65 | 0.44 | 26 | 23 | - | - |
| Sarion | WDVI | 0.35 | 1.91 | 0.67 | 0.06 | 0.72 | 0.05 | 0.77 | 26 | 23 | 0.22 * | 1.51 |
| Sarion | Chlorophyl | 0.26 | 1.47 | 25.58 | 0.06 | 0.48 | 2.25 | 0.82 | 26 | 23 | 0.21 * | 1.47 |
| Sarion | NDRE | 0.62 | 3.26 | 0.60 | 0.04 | 0.99 | 0.02 | 0.88 | 26 | 23 | 0.19 | 1.45 |
| Sarion | NDVI | 0.68 | 3.28 | 0.93 | 0.03 | 1.03 | 0.02 | 0.87 | 26 | 23 | 0.19 | 1.45 |
| Serresta | FTWYield | 0.31 | 1.33 | 58.51 | 0.09 | 0.62 | 7.48 | 0.59 | 25 | 22 | - | - |
| Serresta | WDVI | 0.42 | 2.27 | 0.67 | 0.07 | 1.15 | 0.06 | 0.74 | 23 | 20 | 0.23 | 1.32 |
| Serresta | Chlorophyl | 0.31 | 3.17 | 22.90 | 0.11 | 1.86 | 2.33 | 0.71 | 23 | 20 | 0.29 | 1.42 |
| Serresta | NDRE | 0.62 | 5.62 | 0.58 | 0.11 | 4.20 | 0.02 | 0.74 | 23 | 20 | 0.29 * | 1.62 |
| Serresta | NDVI | 0.70 | 5.89 | 0.93 | 0.08 | 3.94 | 0.03 | 0.78 | 23 | 20 | 0.25 * | 1.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jindo, K.; Teklu, M.G.; van Boheeman, K.; Njehia, N.S.; Narabu, T.; Kempenaar, C.; Molendijk, L.P.G.; Schepel, E.; Been, T.H. Unmanned Aerial Vehicle (UAV) for Detection and Prediction of Damage Caused by Potato Cyst Nematode G. pallida on Selected Potato Cultivars. Remote Sens. 2023, 15, 1429. https://doi.org/10.3390/rs15051429
Jindo K, Teklu MG, van Boheeman K, Njehia NS, Narabu T, Kempenaar C, Molendijk LPG, Schepel E, Been TH. Unmanned Aerial Vehicle (UAV) for Detection and Prediction of Damage Caused by Potato Cyst Nematode G. pallida on Selected Potato Cultivars. Remote Sensing. 2023; 15(5):1429. https://doi.org/10.3390/rs15051429
Chicago/Turabian StyleJindo, Keiji, Misghina Goitom Teklu, Koen van Boheeman, Njane Stephen Njehia, Takashi Narabu, Corne Kempenaar, Leendert P. G. Molendijk, Egbert Schepel, and Thomas H. Been. 2023. "Unmanned Aerial Vehicle (UAV) for Detection and Prediction of Damage Caused by Potato Cyst Nematode G. pallida on Selected Potato Cultivars" Remote Sensing 15, no. 5: 1429. https://doi.org/10.3390/rs15051429
APA StyleJindo, K., Teklu, M. G., van Boheeman, K., Njehia, N. S., Narabu, T., Kempenaar, C., Molendijk, L. P. G., Schepel, E., & Been, T. H. (2023). Unmanned Aerial Vehicle (UAV) for Detection and Prediction of Damage Caused by Potato Cyst Nematode G. pallida on Selected Potato Cultivars. Remote Sensing, 15(5), 1429. https://doi.org/10.3390/rs15051429