


Spatiotemporal Variations in the Sensitivity of Vegetation Growth to Typical Climate Factors on the Qinghai–Tibet Plateau


Abstract
:1. Introduction
2. Materials and Methods
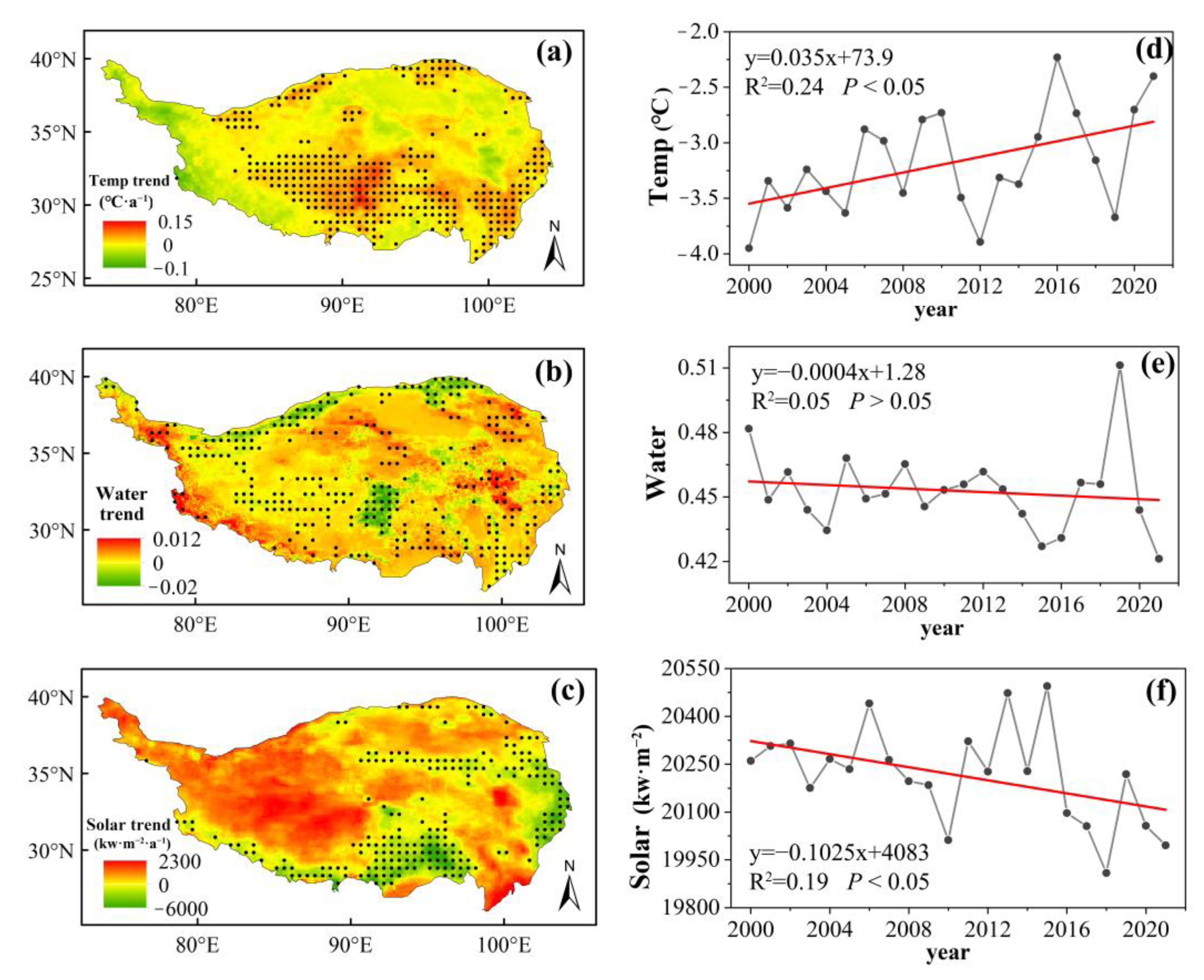
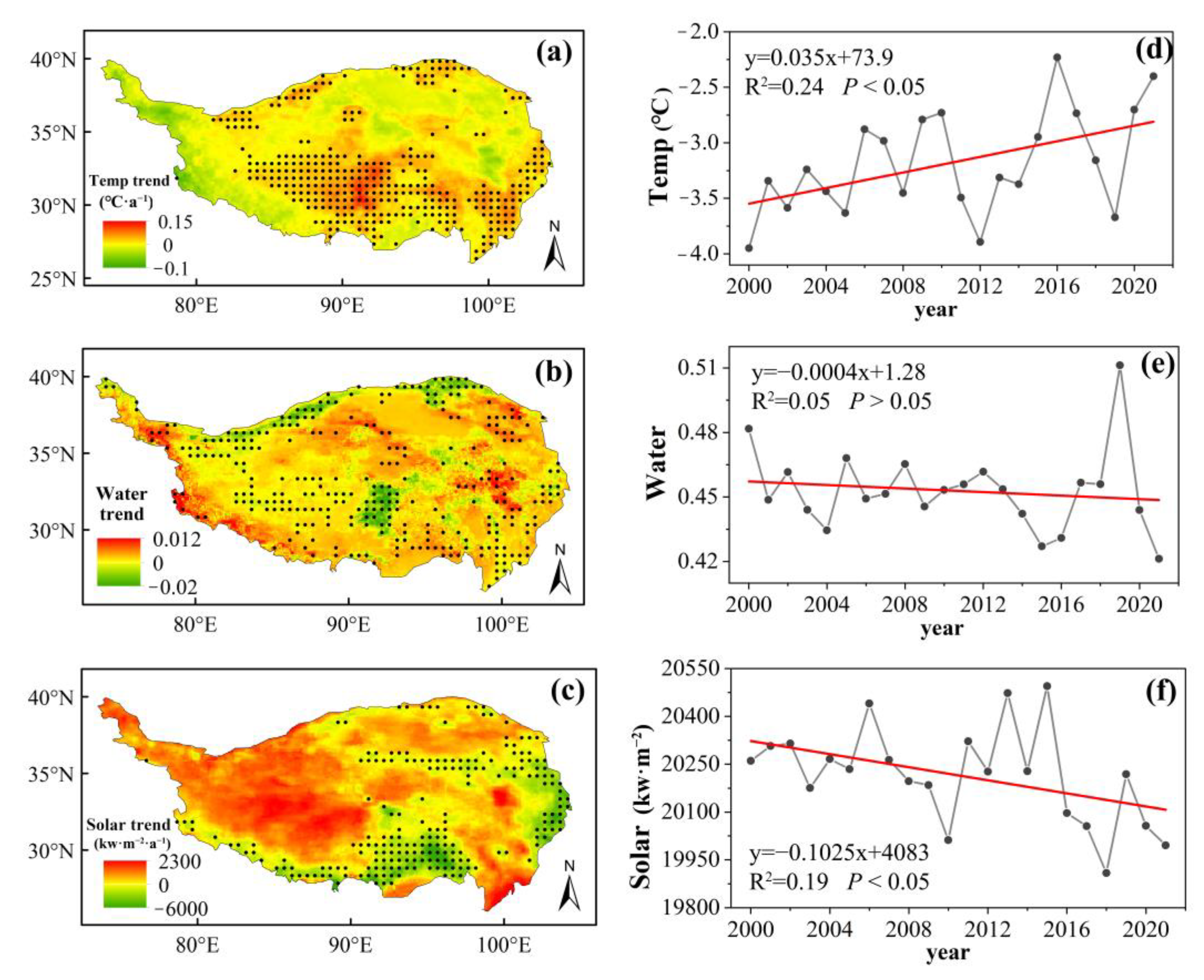
2.1. ERA5-Land Climate Data
2.2. Satellite-Based Vegetation Dynamic Data
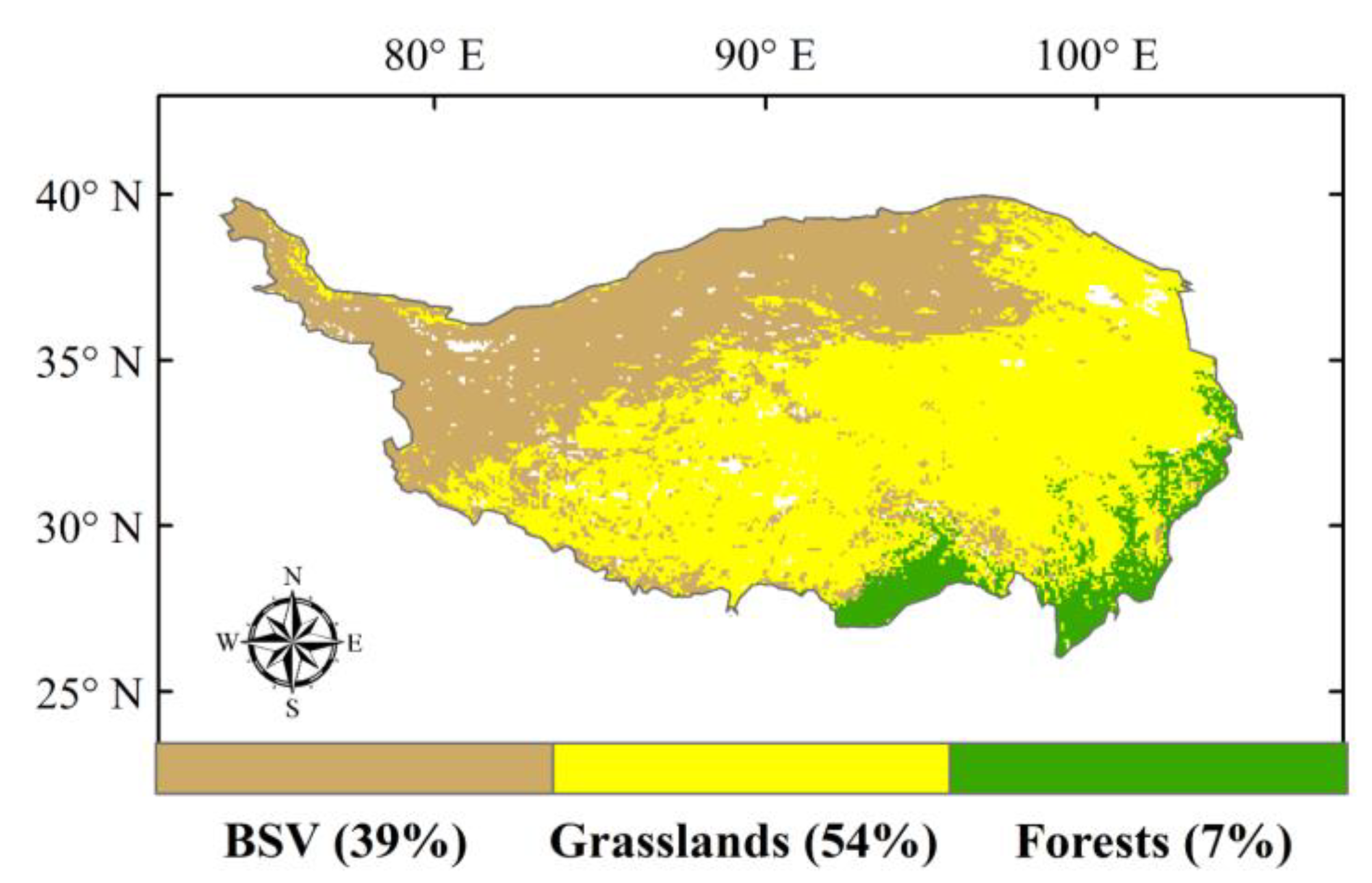
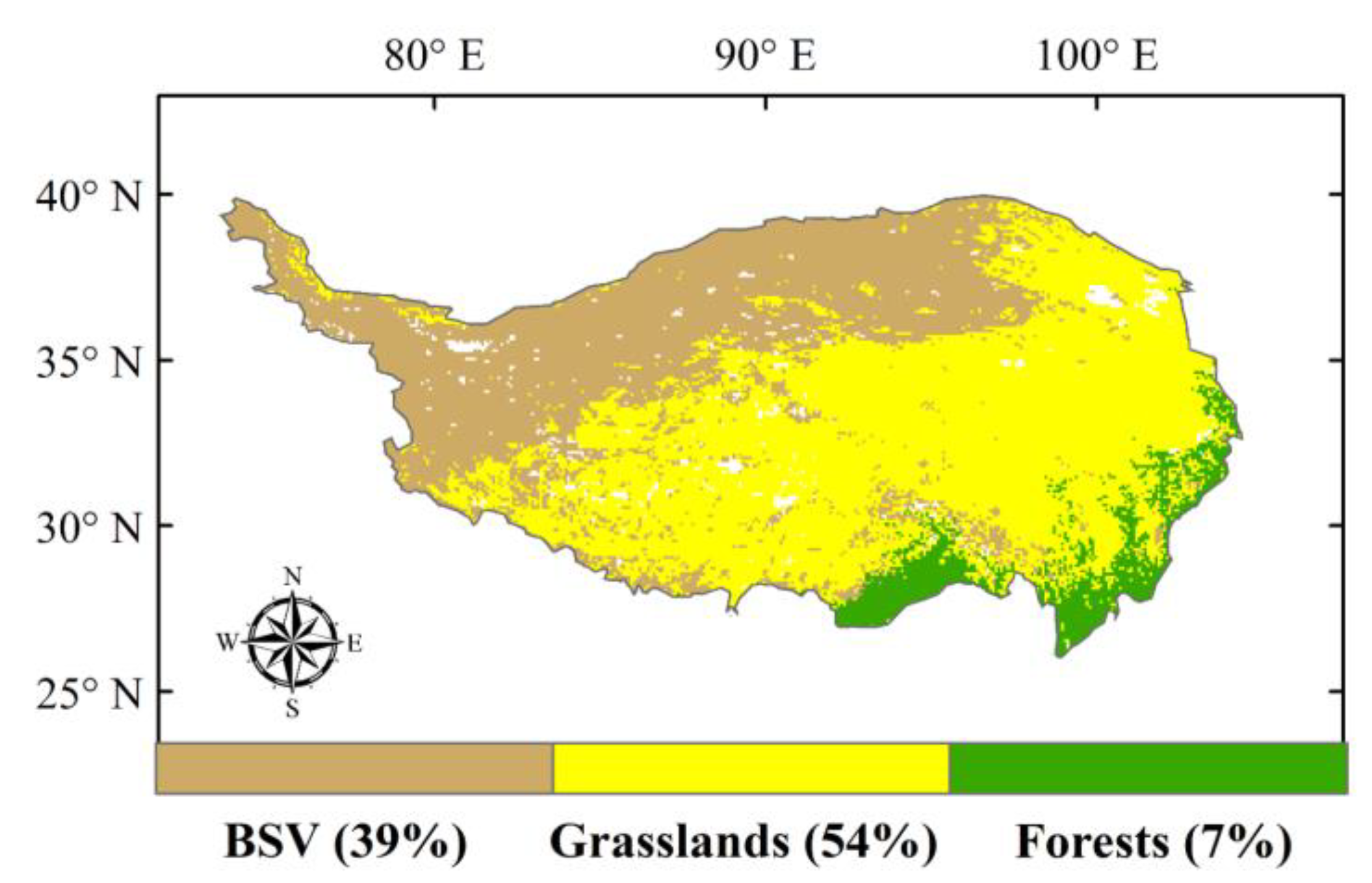
2.3. Land-Cover Data
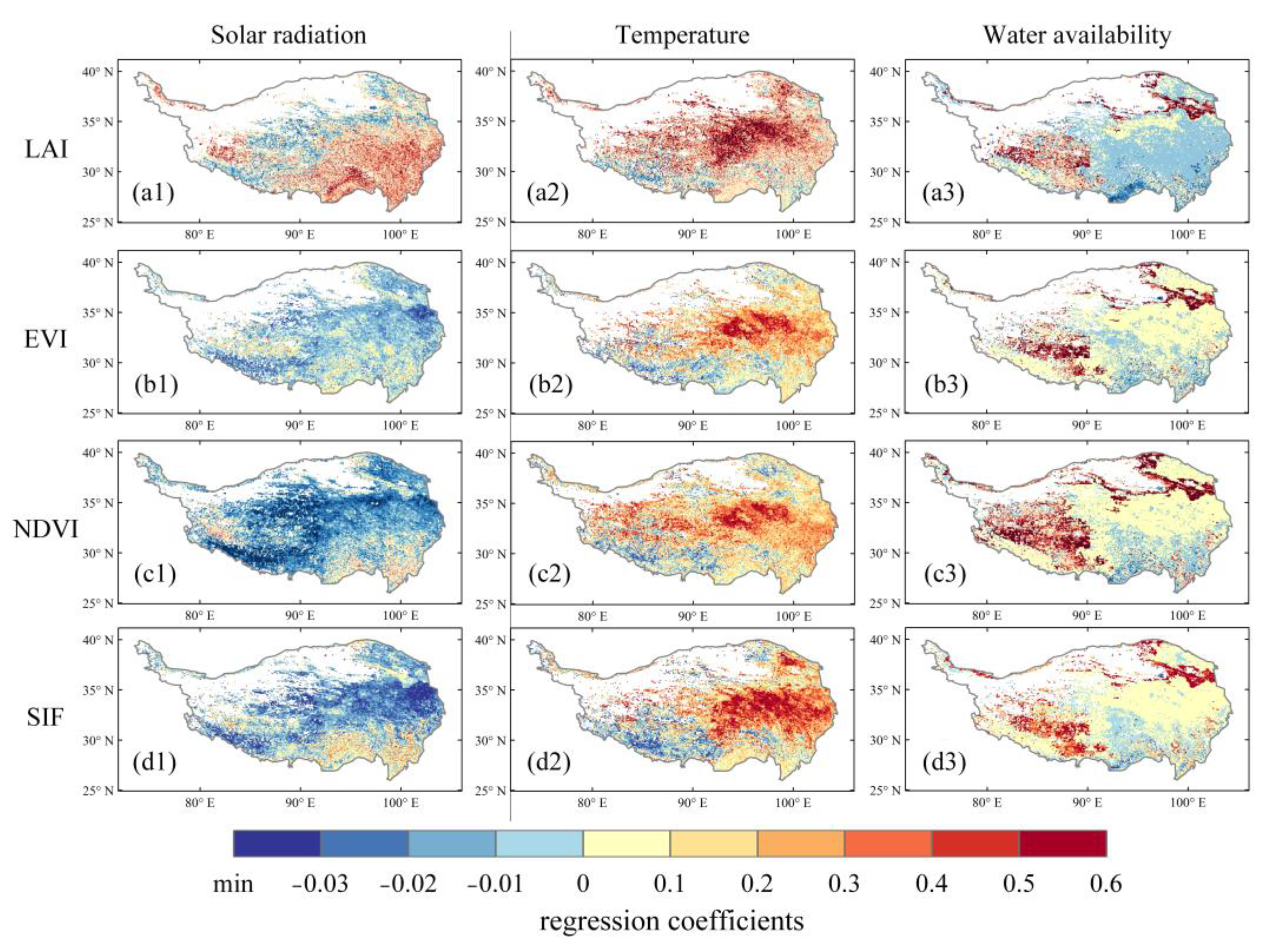
2.4. Vegetation Sensitivity in Response to Climate Change
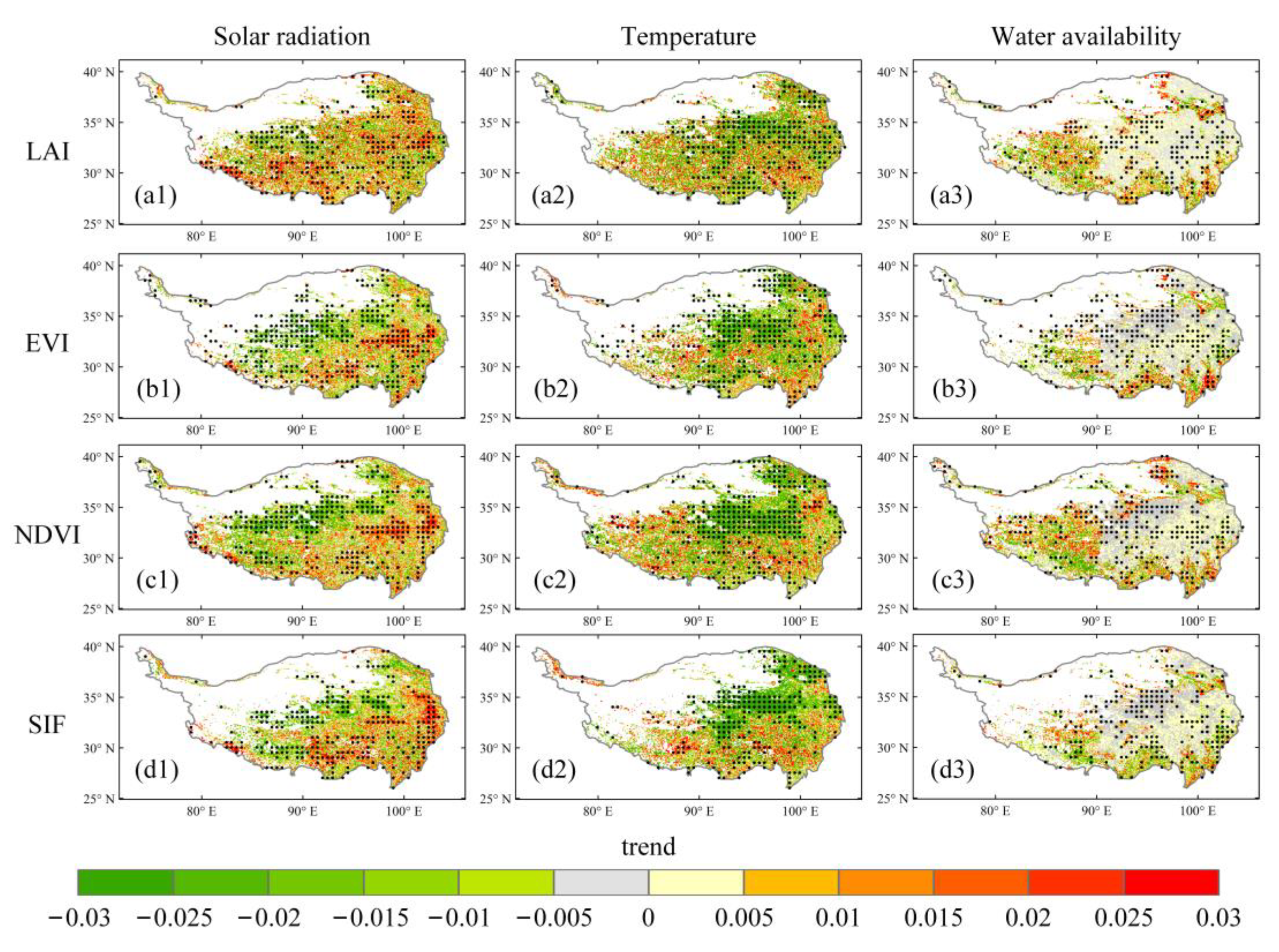
2.5. Trend Analysis with the Mann–Kendall Test
3. Results
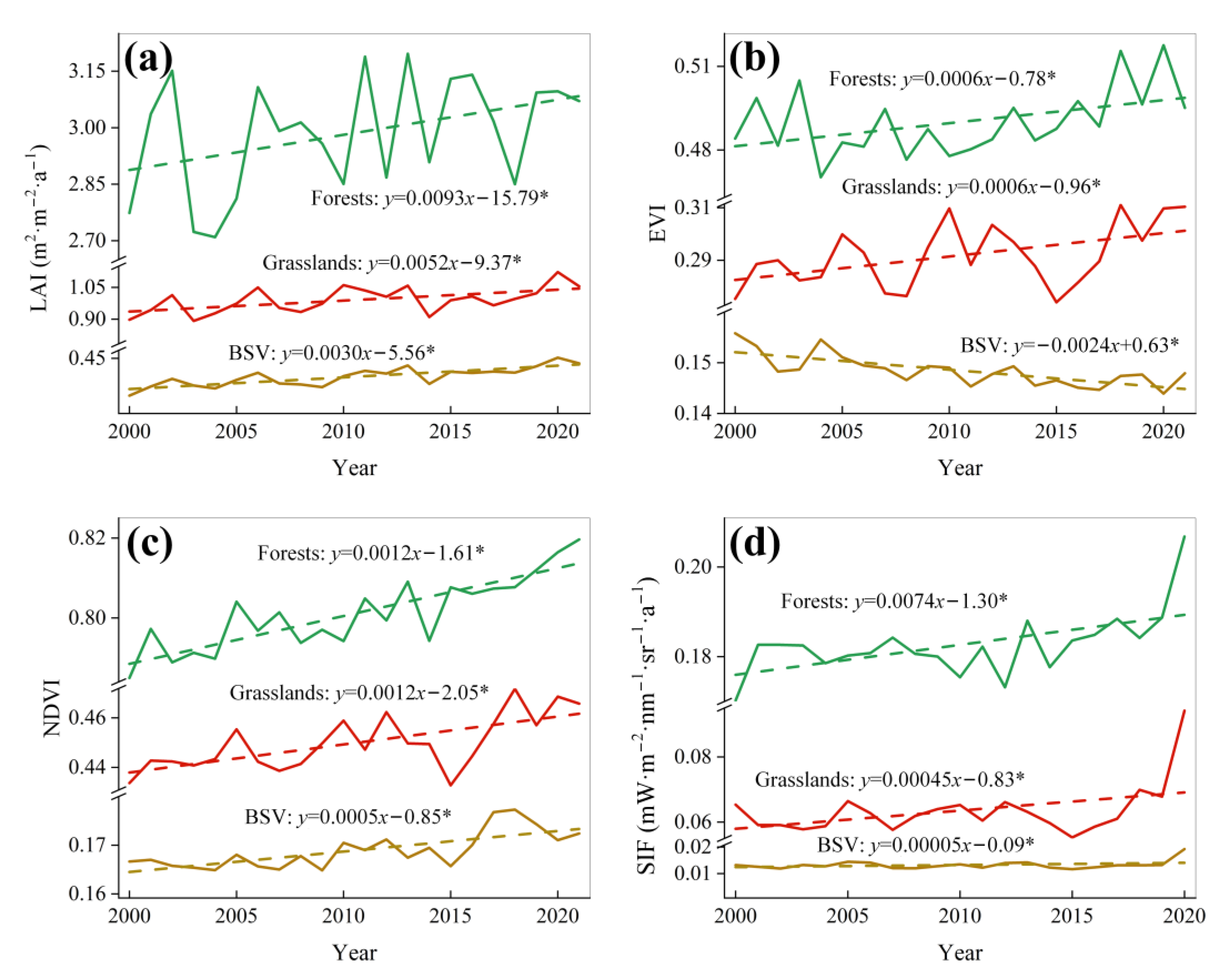
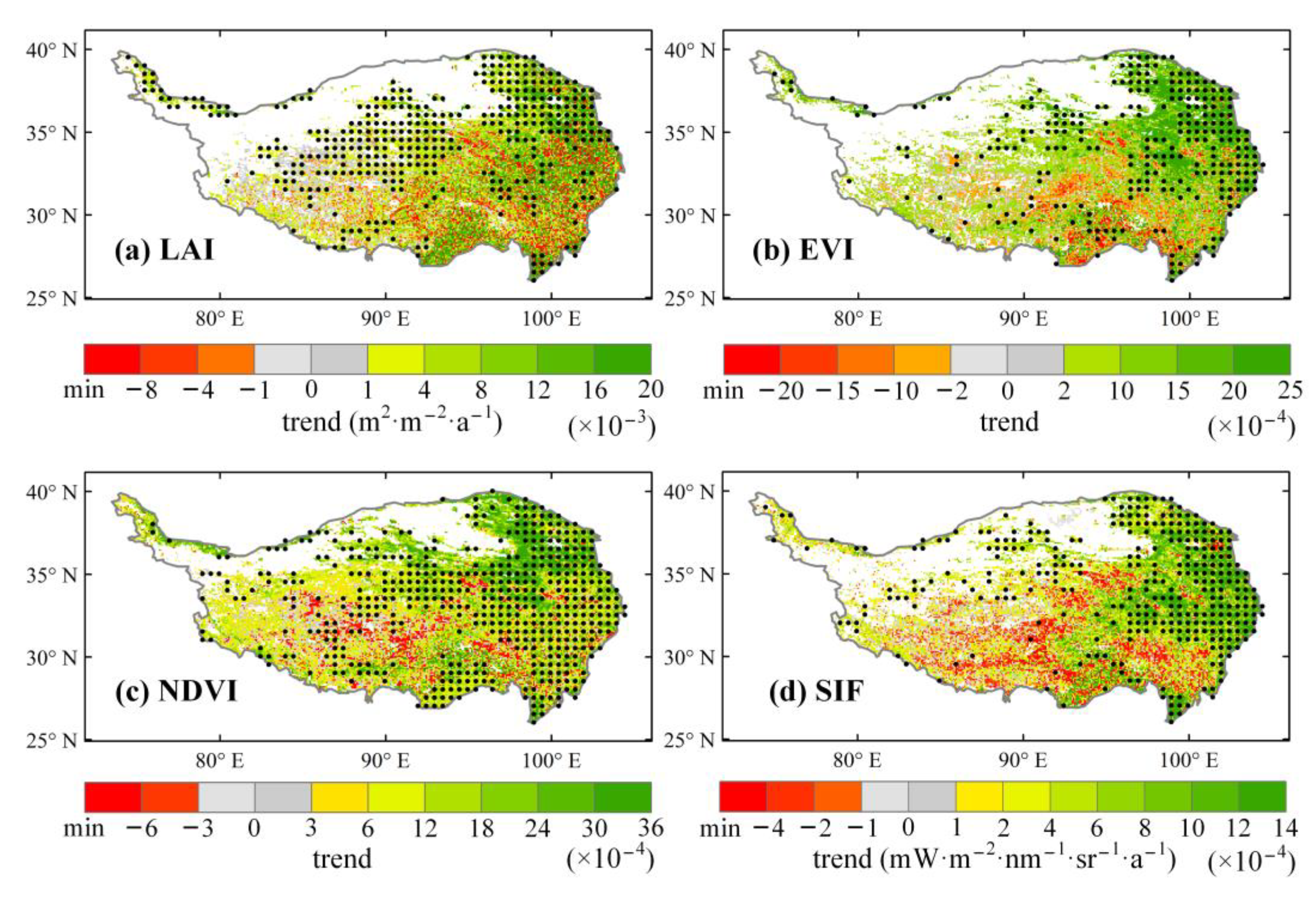
3.1. Interannual Vegetation Dynamics Based on LAI, EVI, NDVI, and SIF Products
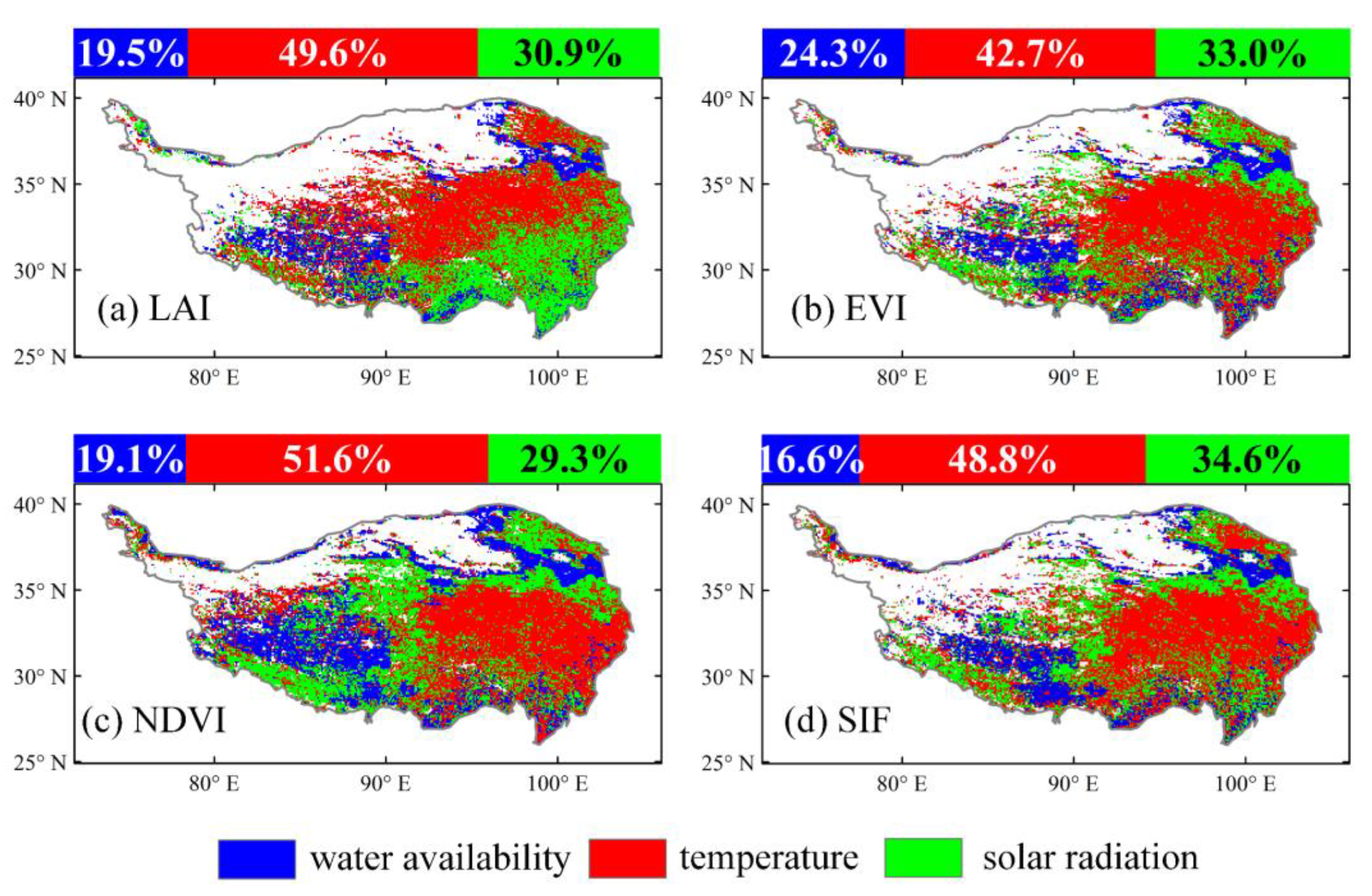
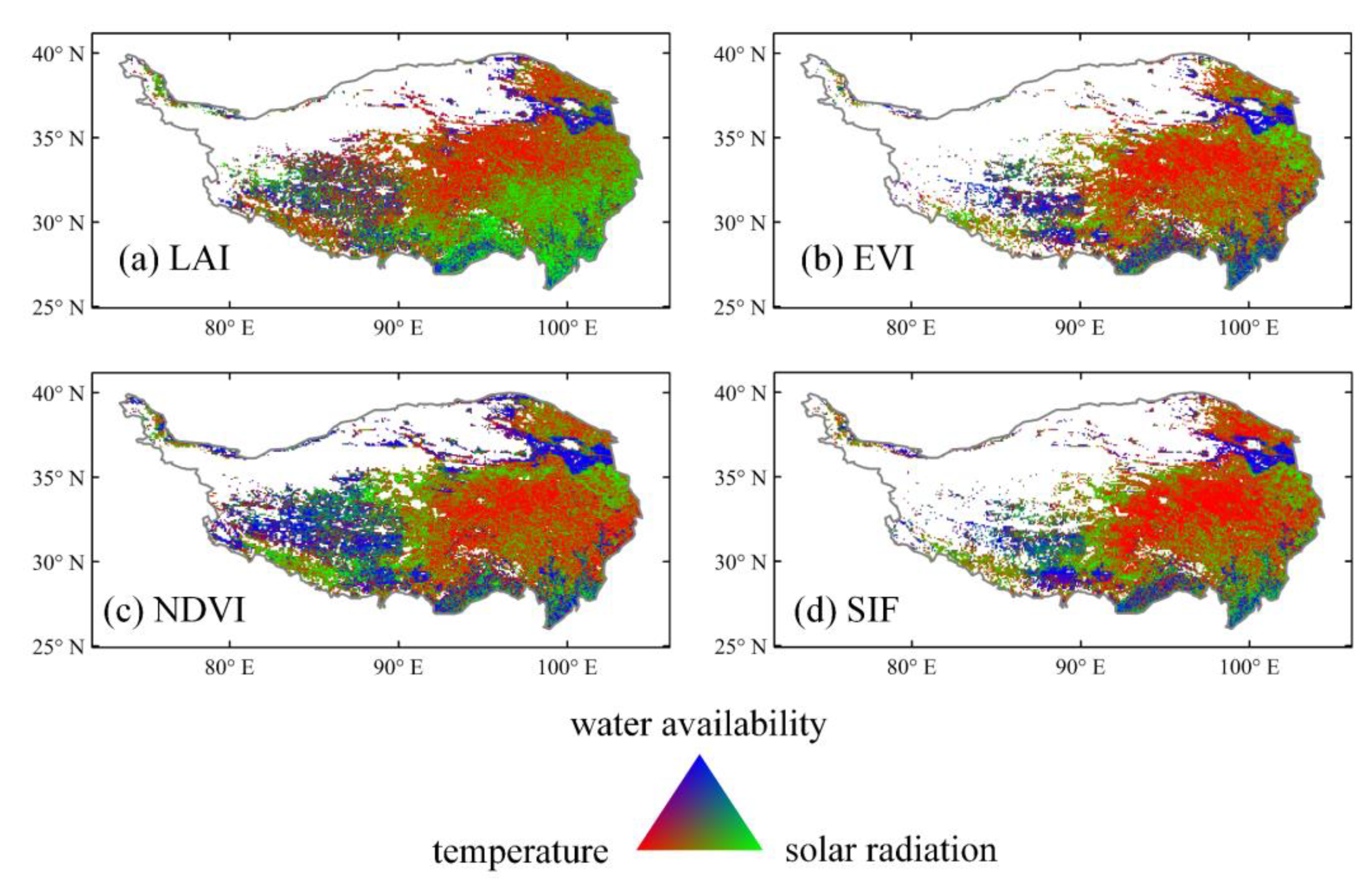
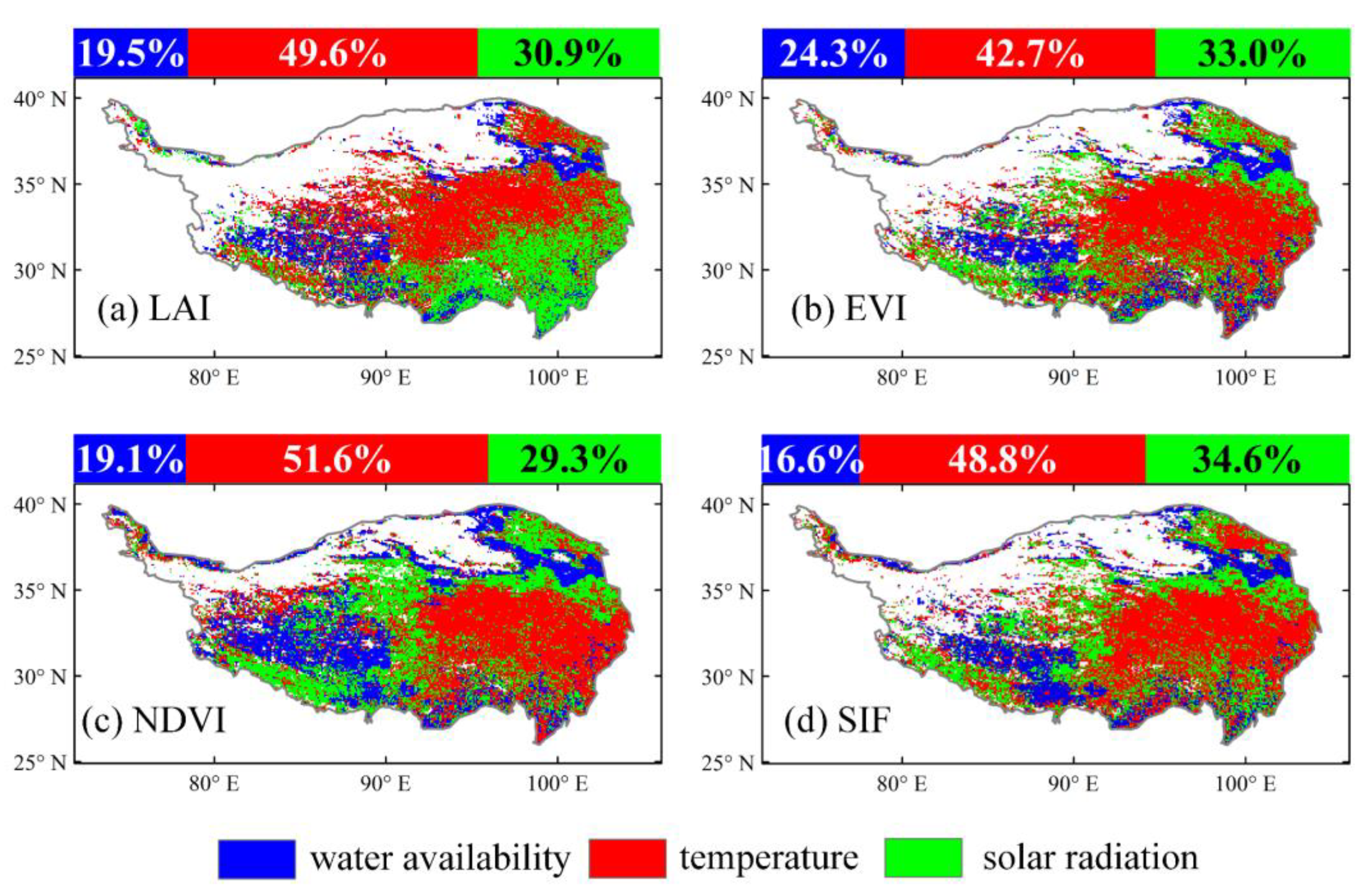
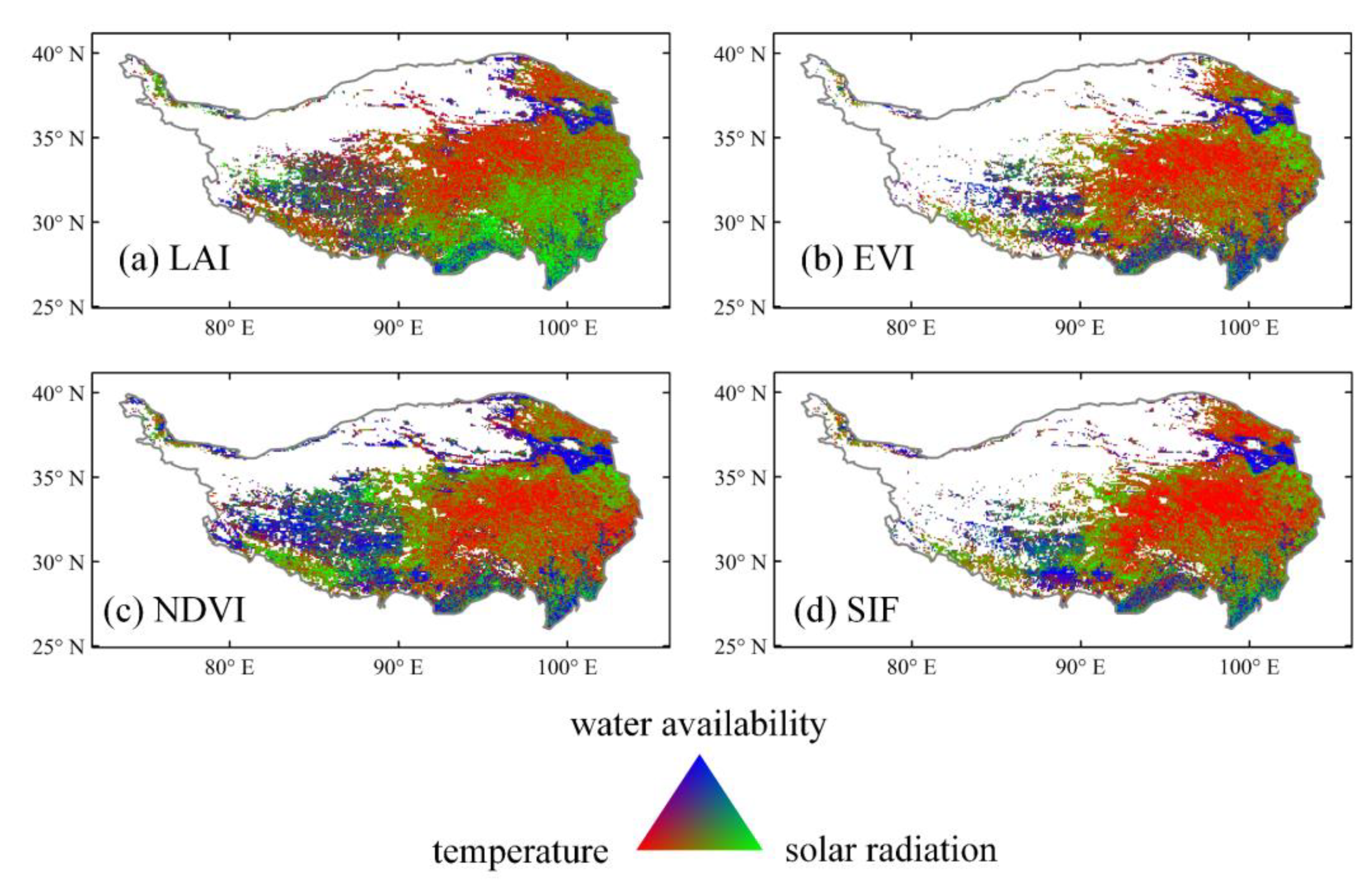
3.2. Time-Invariant Response of Vegetation Dynamics to Climate Changes
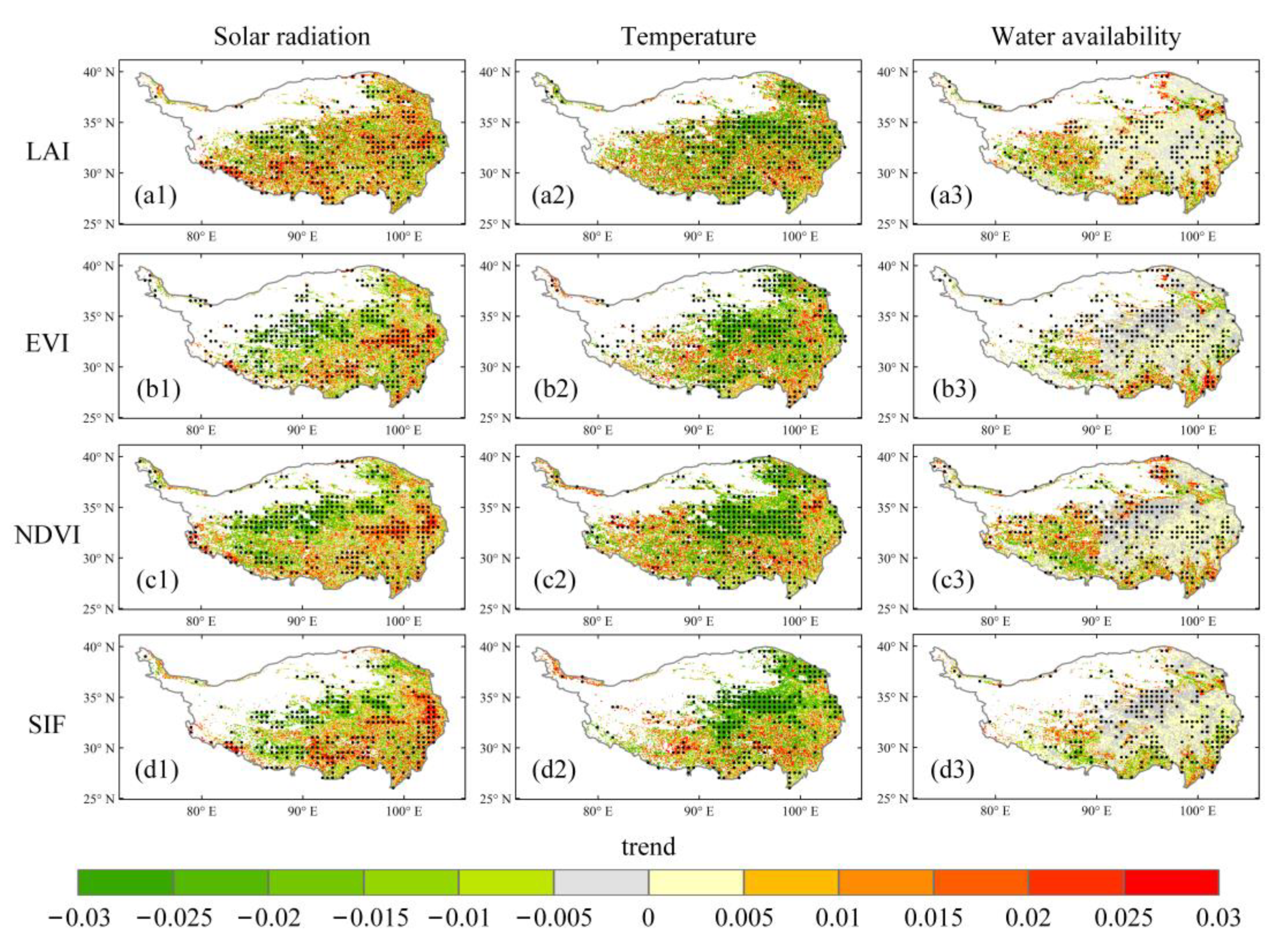
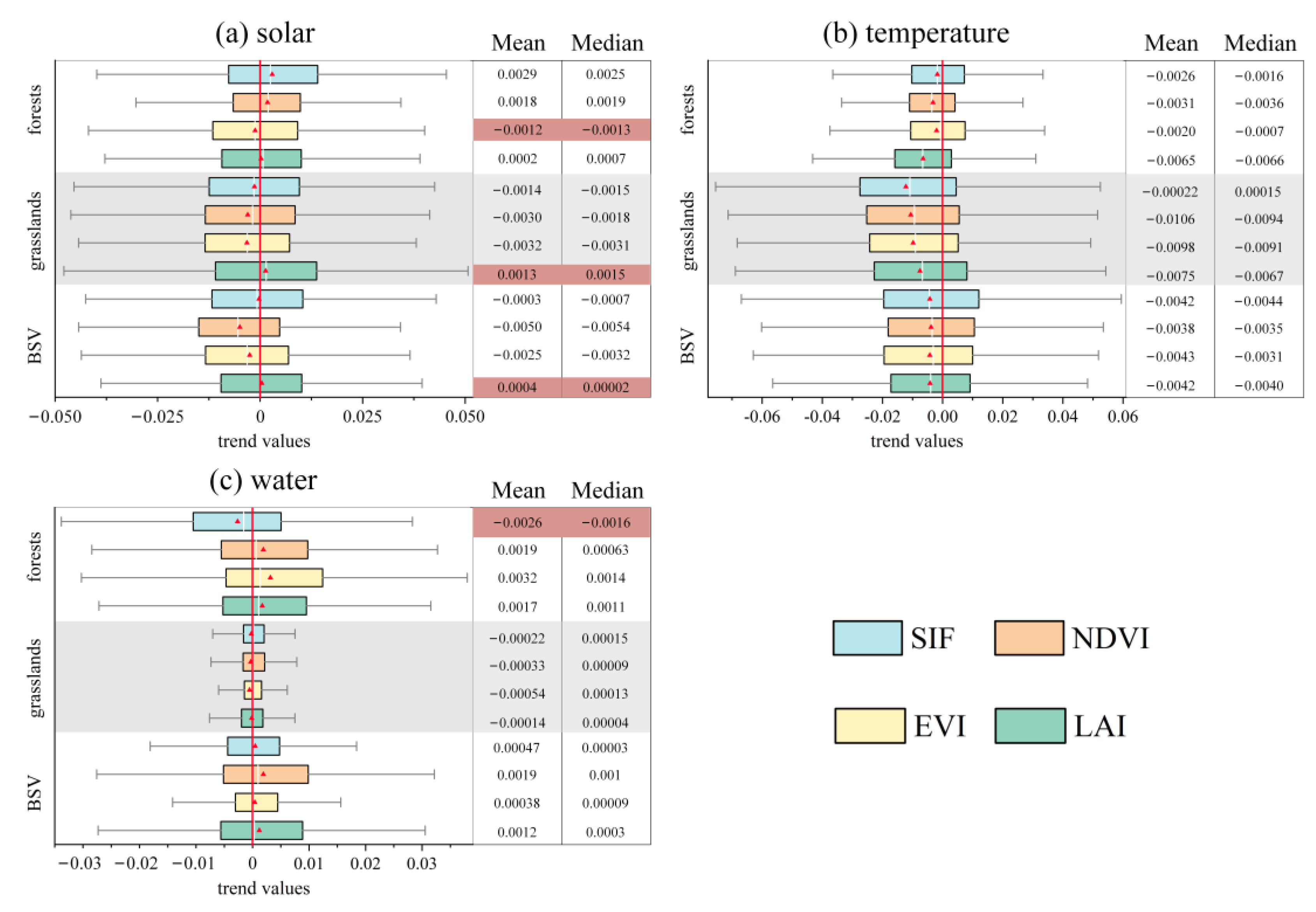
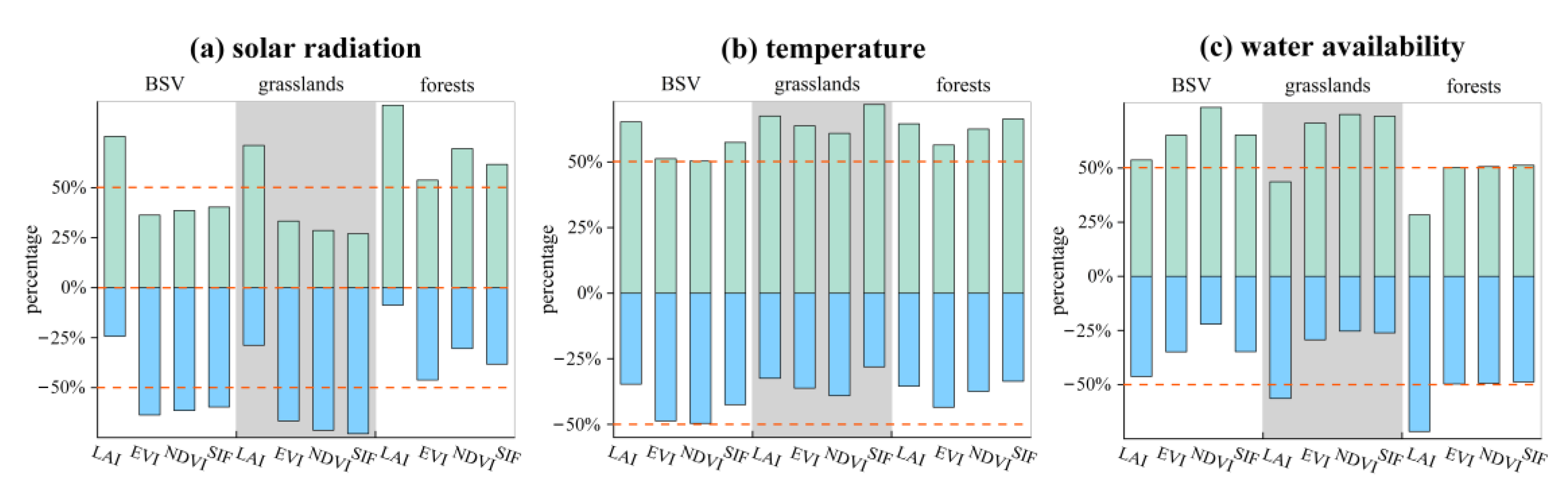
3.3. Time-Variant Response of Vegetation Dynamics to Climate Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Piao, S.; Cui, M.; Chen, A.; Wang, X.; Ciais, P.; Liu, J.; Tang, Y. Altitude and Temperature Dependence of Change in the Spring Vegetation Green-up Date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, Z.; Wang, S.; Hu, Y.; Xu, G.; Luo, C.; Chang, X.; Duan, J.; Lin, Q.; Xu, B.; et al. Response of Ecosystem Respiration to Warming and Grazing during the Growing Seasons in the Alpine Meadow on the Tibetan Plateau. Agric. For. Meteorol. 2011, 151, 792–802. [Google Scholar] [CrossRef]
- Yao, T.; Wu, F.; Ding, L.; Sun, J.; Zhu, L.; Piao, S.; Deng, T.; Ni, X.; Zheng, H.; Ouyang, H. Multispherical Interactions and Their Effects on the Tibetan Plateau’s Earth System: A Review of the Recent Researches. Natl. Sci. Rev. 2015, 2, 468–488. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation Impacts on Vegetation Spring Phenology on the Tibetan Plateau. Glob. Change Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Cao, J.; Hansen, J.; Yao, T.; Joswia, D.R.; Wang, N.; Wu, G.; Wang, M.; Zhao, H.; Yang, W.; et al. Black Soot and the Survival of Tibetan Glaciers. Proc. Natl. Acad. Sci. USA 2009, 106, 22114–22118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Guo, H.D.; Wang, C.Z.; Ji, L.; Li, J.; Wang, K.; Dai, L. The Long-Term Trends (1982–2006) in Vegetation Greenness of the Alpine Ecosystem in the Qinghai-Tibetan Plateau. Environ. Earth Sci. 2014, 72, 1827–1841. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Ogle, K. Tree Growth Sensitivity to Climate Is Temporally Variable. Ecol. Lett. 2020, 23, 1561–1572. [Google Scholar] [CrossRef]
- Xie, X.; He, B.; Guo, L.; Huang, L.; Hao, X.; Zhang, Y.; Liu, X.; Tang, R.; Wang, S. Revisiting Dry Season Vegetation Dynamics in the Amazon Rainforest Using Different Satellite Vegetation Datasets. Agric. For. Meteorol. 2022, 312, 108704. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Q. Assessing Ecological Vulnerability in Western China Based on Time-Integrated NDVI Data. J. Arid. Land 2016, 8, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Jiao, K.; Wu, S. Quantitative Assessment of Ecosystem Vulnerability to Climate Change: Methodology and Application in China. Environ. Res. Lett. 2018, 13, 094016. [Google Scholar] [CrossRef]
- Li, K.; Tong, Z.; Liu, X.; Zhang, J.; Tong, S. Quantitative Assessment and Driving Force Analysis of Vegetation Drought Risk to Climate Change:Methodology and Application in Northeast China. Agric. For. Meteorol. 2020, 282–283, 107865. [Google Scholar] [CrossRef]
- He, L.; Guo, J.; Yang, W.; Jiang, Q.; Chen, L.; Tang, K. Multifaceted Responses of Vegetation to Average and Extreme Climate Change over Global Drylands. Sci. Total Environ. 2023, 858, 159942. [Google Scholar] [CrossRef] [PubMed]
- Abel, C.; Horion, S.; Tagesson, T.; De Keersmaecker, W.; Seddon, A.W.R.; Abdi, A.M.; Fensholt, R. The Human–Environment Nexus and Vegetation–Rainfall Sensitivity in Tropical Drylands. Nat. Sustain. 2021, 4, 25–32. [Google Scholar] [CrossRef]
- Yuan, X.; Hamdi, R.; Ochege, F.U.; Kurban, A.; De Maeyer, P. The Sensitivity of Global Surface Air Temperature to Vegetation Greenness. Int. J. Climatol. 2020, 41, 483–496. [Google Scholar] [CrossRef]
- You, G.; Liu, B.; Zou, C.; Li, H.; McKenzie, S.; He, Y.; Gao, J.; Jia, X.; Altaf Arain, M.; Wang, S.; et al. Sensitivity of Vegetation Dynamics to Climate Variability in a Forest-Steppe Transition Ecozone, North-Eastern Inner Mongolia, China. Ecol. Indic. 2021, 120, 106833. [Google Scholar] [CrossRef]
- Zhang, Q.; Yuan, R.; Singh, V.P.; Xu, C.Y.; Fan, K.; Shen, Z.; Wang, G.; Zhao, J. Dynamic Vulnerability of Ecological Systems to Climate Changes across the Qinghai-Tibet Plateau, China. Ecol. Indic. 2022, 134, 108483. [Google Scholar] [CrossRef]
- Li, W.; Migliavacca, M.; Forkel, M.; Denissen, J.M.C.; Reichstein, M.; Yang, H.; Duveiller, G.; Weber, U.; Orth, R. Widespread Increasing Vegetation Sensitivity to Soil Moisture. Nat. Commun. 2022, 13, 3959. [Google Scholar] [CrossRef]
- Bao, Z.; Zhang, J.; Wang, G.; Guan, T.; Jin, J.; Liu, Y.; Li, M.; Ma, T. The Sensitivity of Vegetation Cover to Climate Change in Multiple Climatic Zones Using Machine Learning Algorithms. Ecol. Indic. 2021, 124, 107443. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, H.; Xu, C.; Wu, X.; Liang, B.; Cao, J.; Chen, D. Modeling Vegetation Greenness and Its Climate Sensitivity with Deep-Learning Technology. Ecol. Evol. 2021, 11, 7335–7345. [Google Scholar] [CrossRef]
- Chen, B.; Chen, H.; Li, M.; Fiedler, S.; Mărgărint, M.C.; Nowak, A.; Wesche, K.; Tietjen, B.; Wu, J. Climate Sensitivity of the Arid Scrublands on the Tibetan Plateau Mediated by Plant Nutrient Traits and Soil Nutrient Availability. Remote Sens. 2022, 14, 4601. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Y.; Wu, J.; Li, S.; Zhang, B.; Zu, J.; Zhang, H.; Ding, M.; Paudel, B. Increasing Sensitivity of Alpine Grasslands to Climate Variability along an Elevational Gradient on the Qinghai-Tibet Plateau. Sci. Total Environ. 2019, 678, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation Phenology on the Qinghai-Tibetan Plateau and Its Response to Climate Change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, W.; Xue, K.; Wang, S.; Zhang, L.; Hu, R.; Zeng, H.; Xu, X.; Li, Y.; Jiang, L.; et al. Grassland Changes and Adaptive Management on the Qinghai–Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 668–683. [Google Scholar] [CrossRef]
- Ma, M.; Yuan, W.; Dong, J.; Zhang, F.; Cai, W.; Li, H. Large-Scale Estimates of Gross Primary Production on the Qinghai-Tibet Plateau Based on Remote Sensing Data. Int. J. Digit. Earth 2018, 11, 1166–1183. [Google Scholar] [CrossRef]
- Sun, B.; Jiang, M.; Han, G.; Zhang, L.; Zhou, J.; Bian, C.; Du, Y.; Yan, L.; Xia, J. Experimental Warming Reduces Ecosystem Resistance and Resilience to Severe Flooding in a Wetland. Sci. Adv. 2022, 8, eabl9526. [Google Scholar] [CrossRef]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehman, A.; et al. Global Ecosystem Thresholds Driven by Aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Wigneron, J.P.; Fan, L.; Ciais, P.; Bastos, A.; Brandt, M.; Chave, J.; Saatchi, S.; Baccini, A.; Fensholt, R. Tropical Forests Did Not Recover from the Strong 2015–2016 El Niño Event. Sci. Adv. 2020, 6, eaay4603. [Google Scholar] [CrossRef] [Green Version]
- Armenteras, D.; Dávalos, L.M.; Barreto, J.S.; Miranda, A.; Hernández-Moreno, A.; Zamorano-Elgueta, C.; González-Delgado, T.M.; Meza-Elizalde, M.C.; Retana, J. Fire-Induced Loss of the World’s Most Biodiverse Forests in Latin America. Sci. Adv. 2021, 7, eabd3357. [Google Scholar] [CrossRef]
- Oliveira, B.F.; Moore, F.C.; Dong, X. Biodiversity Mediates Ecosystem Sensitivity to Climate Variability. Commun. Biol. 2022, 5, 628. [Google Scholar] [CrossRef]
- Macdougall, A.S.; McCann, K.S.; Gellner, G.; Turkington, R. Diversity Loss with Persistent Human Disturbance Increases Vulnerability to Ecosystem Collapse. Nature 2013, 494, 86–89. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, Y.; Wang, Y.; Fu, B. Greater Increases in China’s Dryland Ecosystem Vulnerability in Drier Conditions than in Wetter Conditions. J. Environ. Manag. 2021, 291, 112689. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Bao, A.; Liu, T.; Zheng, G.; Jiang, L.; Guo, H.; Jiang, P.; Yu, T.; De Maeyer, P. Assessing Vegetation Stability to Climate Variability in Central Asia. J. Environ. Manag. 2021, 298, 113330. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Liu, B.; Yuan, Y. Quantifying Vegetation Vulnerability to Climate Variability in China. Remote Sens. 2022, 14, 3491. [Google Scholar] [CrossRef]
- Zhang, Y.; Gentine, P.; Luo, X.; Lian, X.; Liu, Y.; Zhou, S.; Michalak, A.M.; Sun, W.; Fisher, J.B.; Piao, S.; et al. Increasing Sensitivity of Dryland Vegetation Greenness to Precipitation Due to Rising Atmospheric CO2. Nat. Commun. 2022, 13, 4875. [Google Scholar] [CrossRef]
- Hua, T.; Zhao, W.; Cherubini, F.; Hu, X.; Pereira, P. Sensitivity and Future Exposure of Ecosystem Services to Climate Change on the Tibetan Plateau of China. Landsc. Ecol. 2021, 36, 3451–3471. [Google Scholar] [CrossRef]
- Zeng, X.; Hu, Z.; Chen, A.; Yuan, W.; Hou, G.; Han, D.; Liang, M.; Di, K.; Cao, R.; Luo, D. The Global Decline in the Sensitivity of Vegetation Productivity to Precipitation from 2001 to 2018. Glob. Change Biol. 2022, 28, 6823–6833. [Google Scholar] [CrossRef]
- Jiang, P.; Ding, W.; Yuan, Y.; Ye, W.; Mu, Y. Interannual Variability of Vegetation Sensitivity to Climate in China. J. Environ. Manag. 2022, 301, 113768. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.; Berry, J.A.; et al. Recent Global Decline of CO2 Fertilization Effects on Vegetation Photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef]
- Wang, Q.; Ju, Q.; Wang, Y.; Fu, X.; Zhao, W.; Du, Y.; Jiang, P.; Hao, Z. Regional Patterns of Vegetation Dynamics and Their Sensitivity to Climate Variability in the Yangtze River Basin. Remote Sens. 2022, 14, 5623. [Google Scholar] [CrossRef]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Nicolas, J.; Peubey, C.; Radu, R.; Bonavita, M.; Dee, D.; et al. The ERA5 Global Reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of Global Terrestrial Ecosystems to Climate Variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Joiner, J.; Hamed Alemohammad, S.; Zhou, S.; Gentine, P. A Global Spatially Contiguous Solar-Induced Fluorescence (CSIF) Dataset Using Neural Networks. Biogeosciences 2018, 15, 5779–5800. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, A.; Buchmann, N.; Gharun, M. How Well Do Recently Reconstructed Solar-Induced Fluorescence Datasets Model Gross Primary Productivity? Remote Sens. Environ. 2022, 283, 113282. [Google Scholar] [CrossRef]
- Hu, J.; Liu, L.; Guo, J.; Du, S.; Liu, X. Upscaling Solar-Induced Chlorophyll Fluorescence from an Instantaneous to Daily Scale Gives an Improved Estimation of the Gross Primary Productivity. Remote Sens. 2018, 10, 1663. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Piao, S.; Knapp, A.K.; Wang, X.; Peng, S.; Yuan, W.; Running, S.; Mao, J.; Shi, X.; Ciais, P.; et al. Decoupling of Greenness and Gross Primary Productivity as Aridity Decreases. Remote Sens. Environ. 2022, 279, 113120. [Google Scholar] [CrossRef]
- Lahiri, S.N. On the Moving Block Bootstrap under Long Range Dependence. Stat. Probab. Lett. 1993, 18, 405–413. [Google Scholar] [CrossRef]
- Sen, P.K. Journal of the American Statistical Estimates of the Regression Coefficient Based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Mann, H.B. Non-Parametric Test Against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods. Biometrika 1957, 44, 298. [Google Scholar] [CrossRef]
- Chen, J.; Yan, F.; Lu, Q. Spatiotemporal Variation of Vegetation on the Qinghai-Tibet Plateau and the Influence of Climatic Factors and Human Activities on Vegetation Trend (2000–2019). Remote Sens. 2020, 12, 3150. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Chen, Y.; Mindje Kayumba, P.; Wang, X.; Liu, C.; Long, Y.; Sun, F. Biophysical Impacts of Vegetation Dynamics Largely Contribute to Climate Mitigation in High Mountain Asia. Agric. For. Meteorol. 2022, 327, 109233. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Sloan, L.C.; Snyder, M.A.; Bell, J.L.; Kaplan, J.; Shafer, S.L.; Bartlein, P.J. Vegetation Sensitivity to Global Anthropogenic Carbon Dioxide Emissions in a Topographically Complex Region. Glob. Biogeochem. Cycles 2003, 17, 1067. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Ding, Y.; Shi, H.; Cai, H.; Fu, Q.; Liu, S.; Li, T. Analysis and Prediction of Vegetation Dynamic Changes in China: Past, Present and Future. Ecol. Indic. 2020, 117, 106642. [Google Scholar] [CrossRef]
- Jiao, W.; Chang, Q.; Wang, L. The Sensitivity of Satellite Solar-Induced Chlorophyll Fluorescence to Meteorological Drought. Earth’s Future 2019, 7, 558–573. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Bai, X. Scenarios of Potential Vegetation Distribution in the Different Gradient Zones of Qinghai-Tibet Plateau under Future Climate Change. Sci. Total Environ. 2021, 796, 148918. [Google Scholar] [CrossRef]
- Potithep, S.; Nagai, S.; Nasahara, K.N.; Muraoka, H.; Suzuki, R. Two Separate Periods of the LAI-VIs Relationships Using in Situ Measurements in a Deciduous Broadleaf Forest. Agric. For. Meteorol. 2013, 169, 148–155. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Asrar, G.; Kanemasu, E.T. Intercepted Photosynthetically Active Radiation Estimated by Spectral Reflectance. Remote Sens. Environ. 1984, 14, 65–75. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, F.; Zhao, X. The Superiority of Solar-Induced Chlorophyll Fluorescence Sensitivity over Other Vegetation Indices to Drought. J. Arid. Environ. 2022, 204, 104787. [Google Scholar] [CrossRef]
- Arjasakusuma, S.; Mutaqin, B.W.; Sekaranom, A.B.; Marfai, M.A. Sensitivity of Remote Sensing-Based Vegetation Proxies to Climate and Sea Surface Temperature Variabilities in Australia and Parts of Southeast Asia. Int. J. Remote Sens. 2020, 41, 8631–8653. [Google Scholar] [CrossRef]
- Walther, S.; Voigt, M.; Thum, T.; Gonsamo, A.; Zhang, Y.; Köhler, P.; Jung, M.; Varlagin, A.; Guanter, L. Satellite Chlorophyll Fluorescence Measurements Reveal Large-Scale Decoupling of Photosynthesis and Greenness Dynamics in Boreal Evergreen Forests. Glob. Change Biol. 2016, 22, 2979–2996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, G.; Sun, W. Temperature Sensitivities of Vegetation Indices and Aboveground Biomass Are Primarily Linked with Warming Magnitude in High-Cold Grasslands. Sci. Total Environ. 2022, 843, 157002. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Wang, H.; Harrison, S.P.; Prentice, I.C.; Qiao, S.; Tan, S. Optimality Principles Explaining Divergent Responses of Alpine Vegetation to Environmental Change. Glob. Change Biol. 2022, 29, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Mi, Z.; Lin, L.; Wang, Y.; Zhang, Z.; Zhang, F.; Wang, H.; Liu, L.; Zhu, B.; Cao, G.; et al. Shifting Plant Species Composition in Response to Climate Change Stabilizes Grassland Primary Production. Proc. Natl. Acad. Sci. USA 2018, 115, 4051–4056. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhang, X.; Niu, B.; He, Y.; Wang, X.; Wu, J. Changes in Plant Species Richness Distribution in Tibetan Alpine Grasslands under Different Precipitation Scenarios. Glob. Ecol. Conserv. 2020, 21, e00848. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Niu, B.; Zheng, Y.; He, Y.; Cao, Y.; Feng, Y.; Wu, J. Divergent Climate Sensitivities of the Alpine Grasslands to Early Growing Season Precipitation on the Tibetan Plateau. Remote Sens. 2022, 14, 2484. [Google Scholar] [CrossRef]
- Sun, J.; Liu, B.; You, Y.; Li, W.; Liu, M.; Shang, H.; He, J.S. Solar Radiation Regulates the Leaf Nitrogen and Phosphorus Stoichiometry across Alpine Meadows of the Tibetan Plateau. Agric. For. Meteorol. 2019, 271, 92–101. [Google Scholar] [CrossRef]
- Fang, O.; Zhang, Q.B. Tree Resilience to Drought Increases in the Tibetan Plateau. Glob. Change Biol. 2019, 25, 245–253. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Xiong, J.; He, W.; Yong, Z.; Wang, X. Responses of the Remote Sensing Drought Index with Soil Information to Meteorological and Agricultural Droughts in Southeastern Tibet. Remote Sens. 2022, 14, 6125. [Google Scholar] [CrossRef]
- Chen, C.; He, B.; Yuan, W.; Guo, L.; Zhang, Y. Increasing Interannual Variability of Global Vegetation Greenness. Environ. Res. Lett. 2019, 14, 124005. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | Solar Radiation | Water Availability | ||||
|---|---|---|---|---|---|---|
| Trend | Sign | Trend | Sign | Trend | Sign | |
| Forests | Downward | Positive | Upward | Positive | Upward | No sign |
| Grasslands | Downward | Positive | Downward | Negative | No trend | Positive |
| BSVs | Downward | Positive | Downward | Negative | Upward | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, K.; Chen, J.; Yang, H.; Yang, Y.; Hu, Z. Spatiotemporal Variations in the Sensitivity of Vegetation Growth to Typical Climate Factors on the Qinghai–Tibet Plateau. Remote Sens. 2023, 15, 2355. https://doi.org/10.3390/rs15092355
Wu K, Chen J, Yang H, Yang Y, Hu Z. Spatiotemporal Variations in the Sensitivity of Vegetation Growth to Typical Climate Factors on the Qinghai–Tibet Plateau. Remote Sensing. 2023; 15(9):2355. https://doi.org/10.3390/rs15092355
Chicago/Turabian StyleWu, Kai, Jiahao Chen, Han Yang, Yue Yang, and Zhongmin Hu. 2023. "Spatiotemporal Variations in the Sensitivity of Vegetation Growth to Typical Climate Factors on the Qinghai–Tibet Plateau" Remote Sensing 15, no. 9: 2355. https://doi.org/10.3390/rs15092355
APA StyleWu, K., Chen, J., Yang, H., Yang, Y., & Hu, Z. (2023). Spatiotemporal Variations in the Sensitivity of Vegetation Growth to Typical Climate Factors on the Qinghai–Tibet Plateau. Remote Sensing, 15(9), 2355. https://doi.org/10.3390/rs15092355