

Early Detection of Drought Stress in Durum Wheat Using Hyperspectral Imaging and Photosystem Sensing

 ,
,  , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
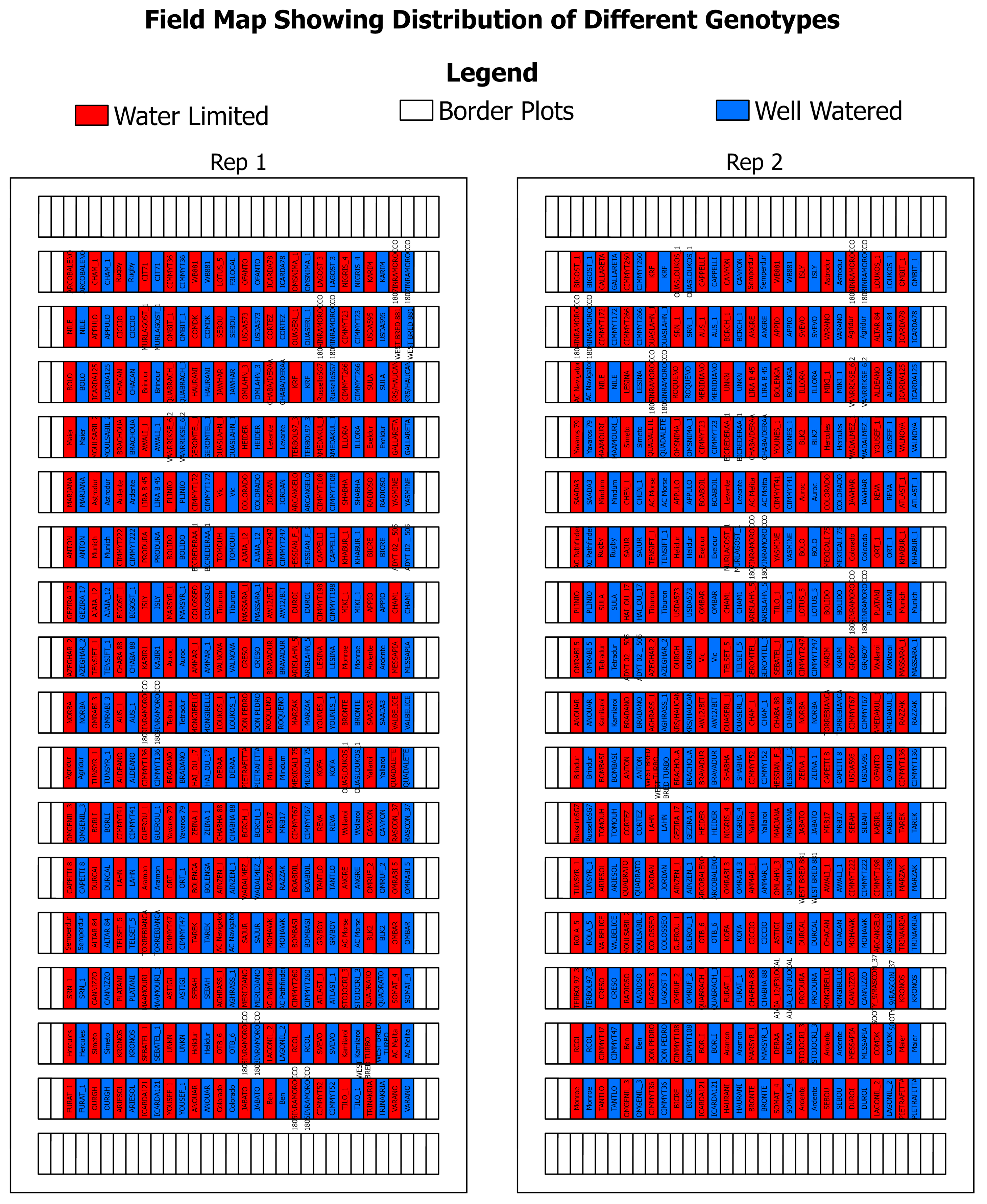
2. Experimental Setup and Data Collection
2.1. Gantry Hyperspectral System
2.2. Plant Physiological Data
3. Methods
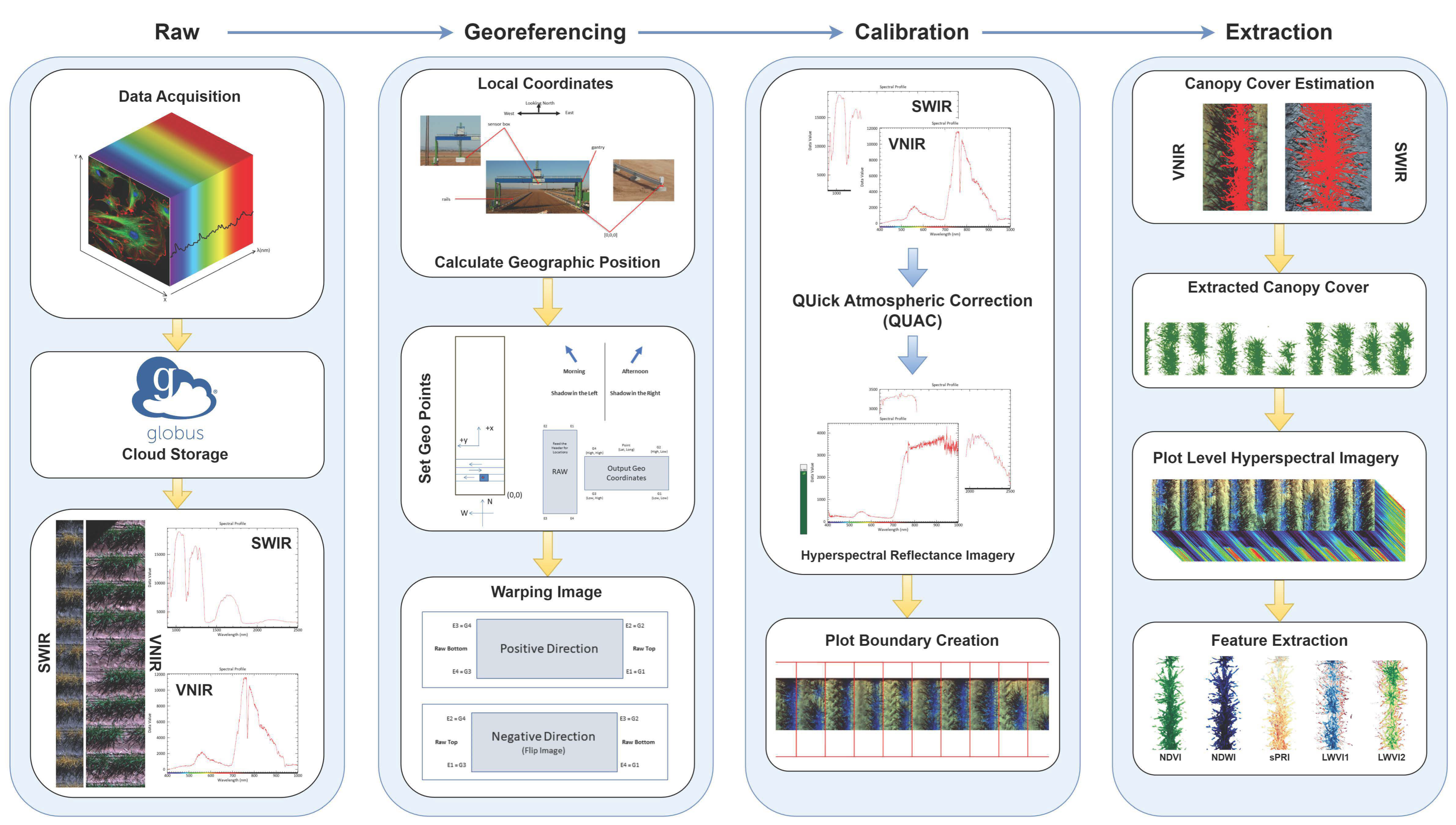
3.1. Hyperspectral Image Acquisition, Georeferencing, and Calibration
3.2. Plot Segmentation and Canopy Cover Estimation
3.3. Feature (Index) Extraction
4. Results
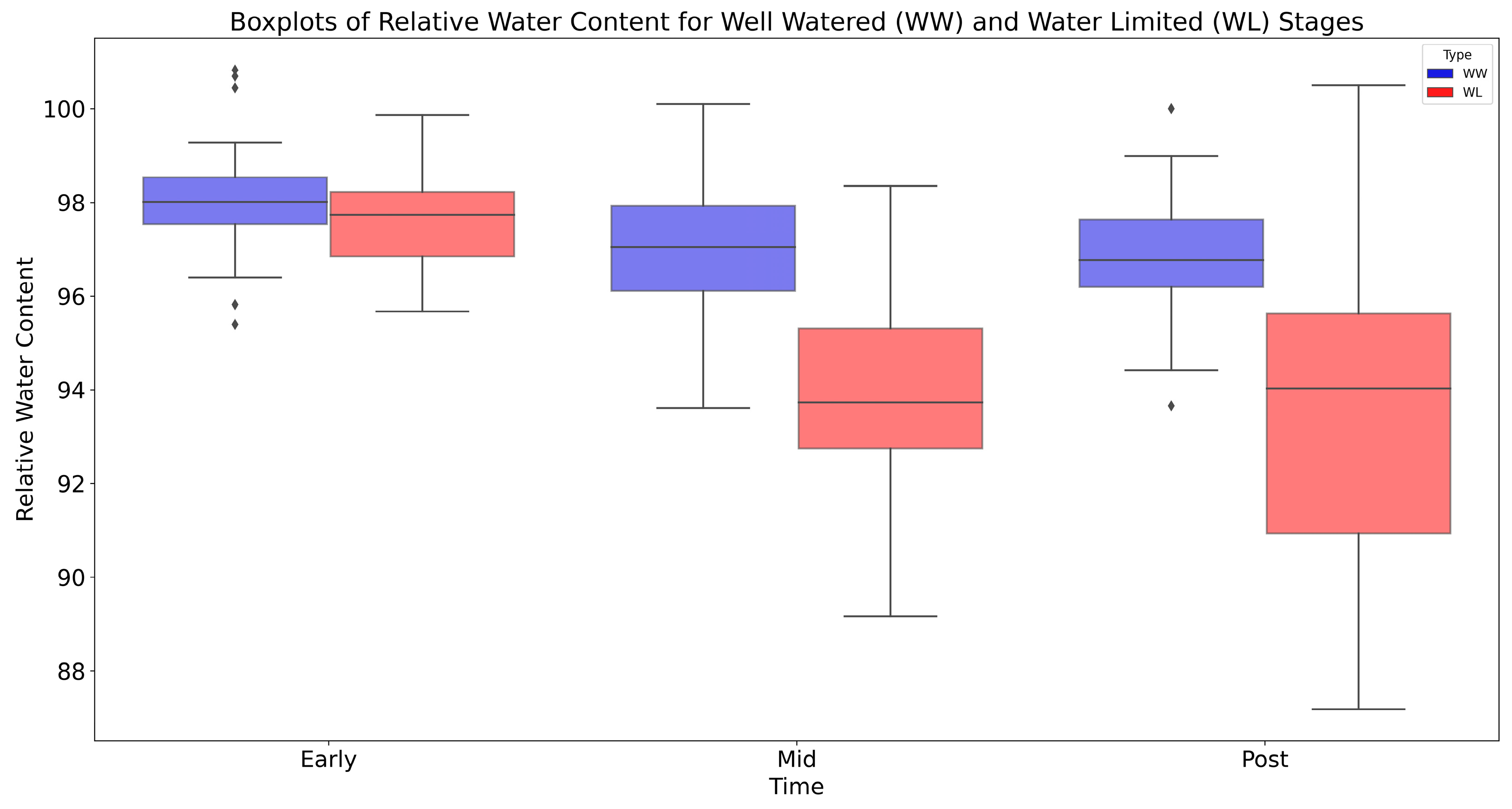
4.1. Changes in Relative Water Content (RWC)
4.2. Statistical Analysis
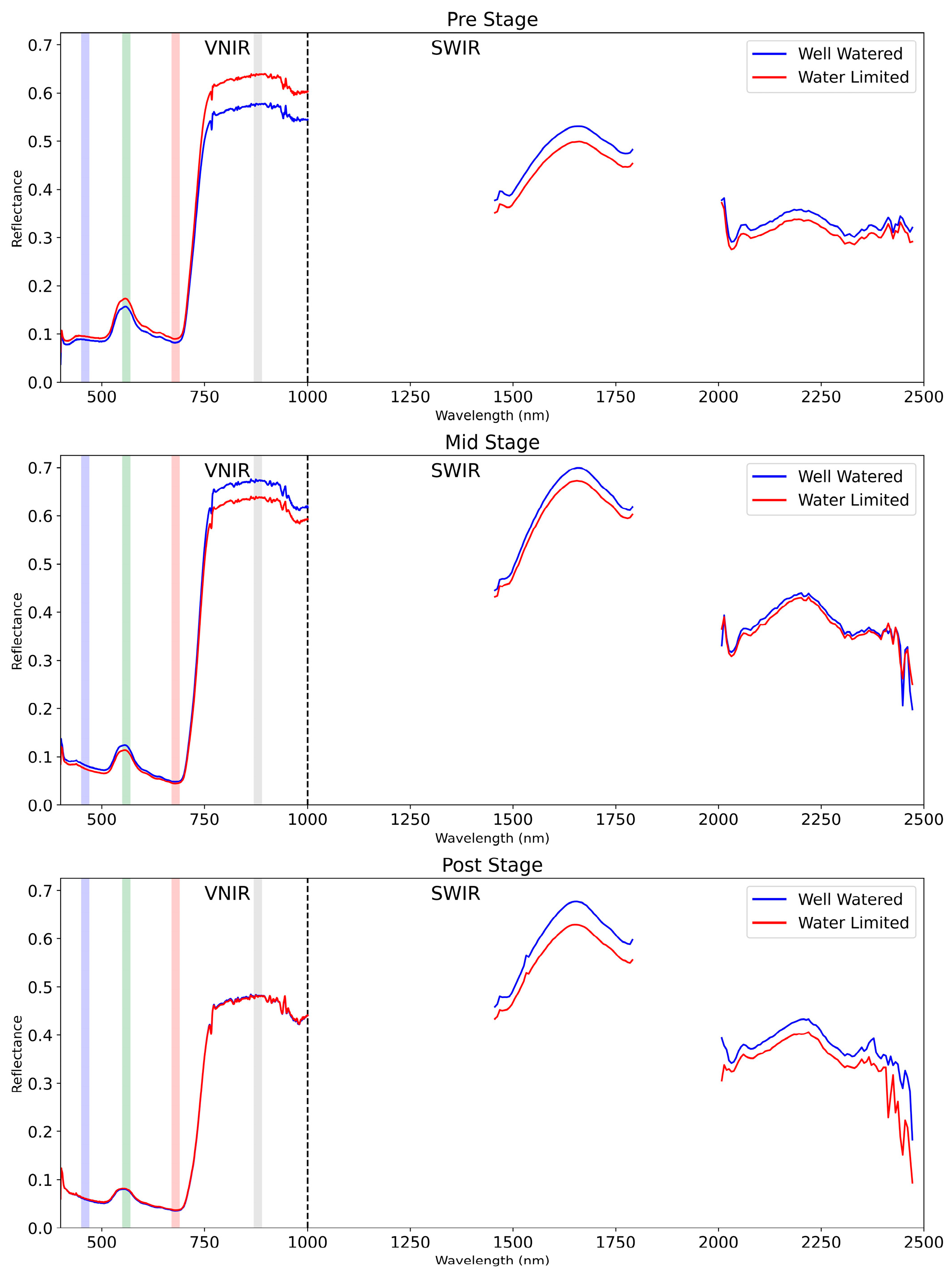
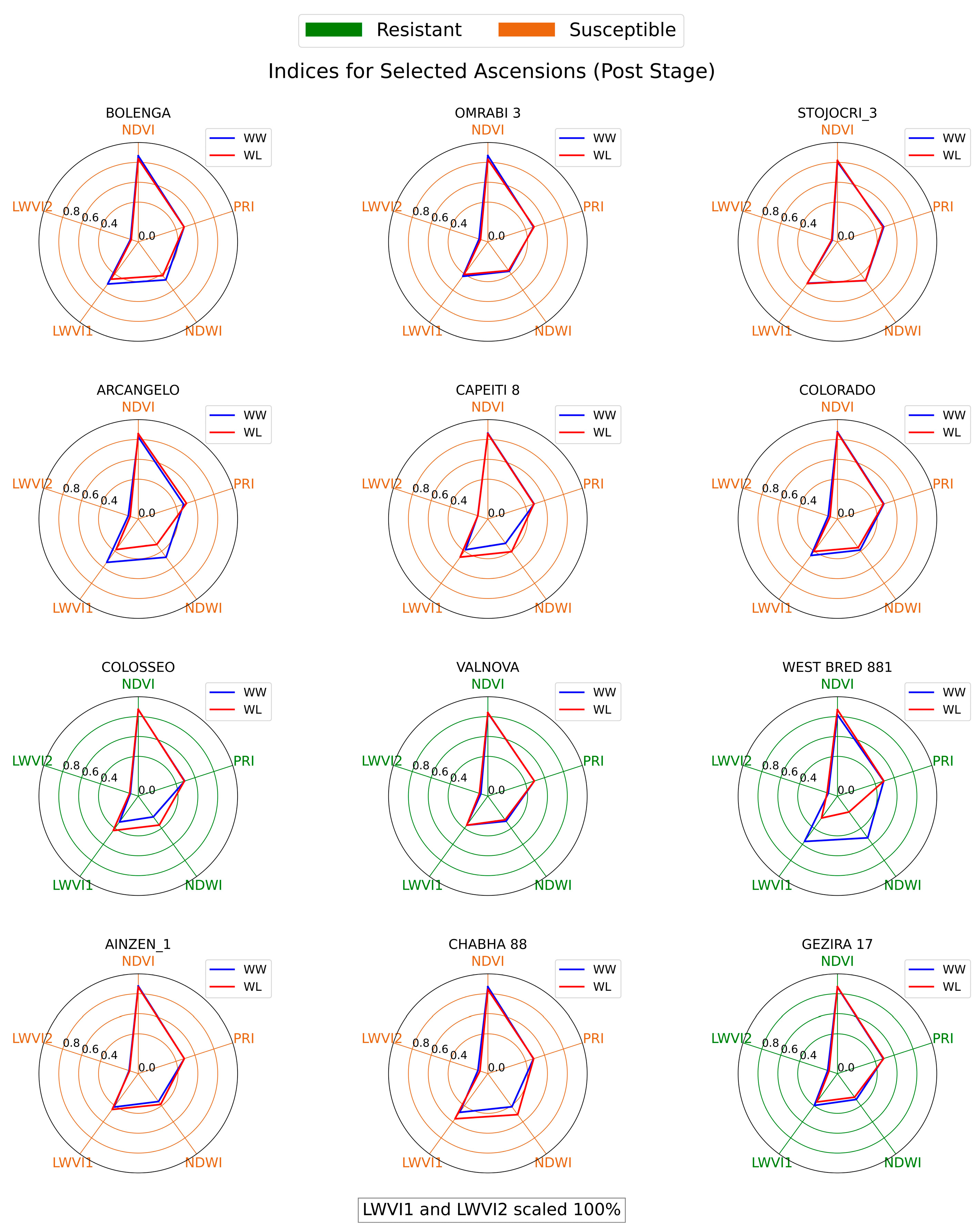
4.3. Spectral Vegetation and Water Indices
5. Discussion
5.1. Changes in Relative Water Content (RWC)
5.2. Analysis of Spectral Vegetation and Water Indices
5.3. Genotypic Responses across Different Drought Stages
6. Conclusions
- NDVI is effective for assessing plant health, but LWVI, especially LWVI2, is more sensitive to early water stress, making it ideal for early-stage drought-stress analysis.
- Early signs of water stress, noticeable in declines in NDWI and LWVI indices even in well-watered conditions, can predict genotypes’ late-season drought tolerance.
- Comparisons of spectral data from hyperspectral sensors with ground truth measurements identified distinct spectral signatures of varying water-stress levels, enhancing understanding of genotypic drought resistance or susceptibility.
- LWVI2, in conjunction with ground truth RWC data, emerged as the most reliable method for evaluating drought tolerance, aligning well with physiological responses, and effectively distinguishing between drought-resistant and drought-prone genotypes.
- Hyperspectral imaging’s effectiveness in capturing spatial and temporal variability in drought response confirms its potential for the early detection and management of drought stress in agriculture.
- Genotypes like COLOSSEO, VALNOVA, and GEZIRA 17 demonstrated strong drought resistance, with their spectral indices under water-limited conditions closely resembling those in well-watered conditions, indicating robust drought tolerance.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhai, S.; Song, G.; Qin, Y.; Ye, X.; Lee, J. Modeling the Impacts of Climate Change and Technical Progress on the Wheat Yield in Inland China: An Autoregressive Distributed Lag Approach. PLoS ONE 2017, 12, e0184474. [Google Scholar] [CrossRef] [PubMed]
- Kusunose, Y.; Rossi, J.J.; Van Sanford, D.A.; Alderman, P.D.; Anderson, J.A.; Chai, Y.; Gerullis, M.K.; Jagadish, S.V.K.; Paul, P.A.; Tack, J.B.; et al. Sustaining productivity gains in the face of climate change: A research agenda for US wheat. Glob. Change Biol. 2022, 29, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Bronson, K.F.; White, J.W.; Conley, M.M.; Hunsaker, D.J.; Thorp, K.R.; French, A.N.; Mackey, B.E.; Holland, K.H. Active Optical Sensors in Irrigated Durum Wheat: Nitrogen and Water Effects. Agron. J. 2017, 109, 1060–1071. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture. Small Grains Annual Summary; USDA Economics, Statistics and Market Information System: Washington, DC, USA, 2022. [Google Scholar]
- Porporato, A.; Laio, F.; Ridolfi, L.; Rodriguez-Iturbe, I. Plants in water-controlled ecosystems: Active role in hydrologic processes and response to water stress: III. Vegetation water stress. Adv. Water Resour. 2001, 24, 725–744. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Fereres, E.; Acevedo, E.; Henderson, D.W. Water Stress and Dynamics of Growth and Yield of Crop Plants. In Water and Plant Life: Problems and Modern Approaches; Lange, O.L., Kappen, L., Schulze, E.D., Eds.; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 1976; pp. 281–305. [Google Scholar]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; OsÓRio, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Alqudah, A.M.; Dawood, M.G.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed]
- Sall, A.A.; Chiari, T.; Legesse, W.; Seid-Ahmed, K.; Ortiz, R.; Ginkel, M.V.; Bassi, F.M. Durum Wheat (Triticum Durum Desf.): Origin, Cultivation and Potential Expansion in Sub-Saharan Africa. Agronomy 2019, 9, 263. [Google Scholar] [CrossRef]
- Hebrard, A.; Oulahna, D.; Galet, L.; Cuq, B.; Abecassis, J.; Fages, J. Hydration Properties of Durum Wheat Semolina: Influence of Particle Size and Temperature. Powder Technol. 2003, 130, 211–218. [Google Scholar] [CrossRef]
- Liu, H.; Searle, I.R.; Mather, D.E.; Able, A.J.; Able, J.A. Morphological, Physiological and Yield Responses of Durum Wheat to Pre-Anthesis Water-Deficit Stress Are Genotype-Dependent. Crop Pasture Sci. 2015, 66, 1024–1038. [Google Scholar] [CrossRef]
- Pascale, S.D.; Costa, L.; Vallone, S.; Barbieri, G.; Maggio, A. Increasing Water Use Efficiency in Vegetable Crop Production: From Plant to Irrigation Systems Efficiency. Horttechnology 2011, 21, 301–308. [Google Scholar] [CrossRef]
- Jones, H.G.; Vaughan, R.A. Remote Sensing of Vegetation: Principles, Techniques, and Applications, 1st ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2010. [Google Scholar]
- Blackburn, G.L. Hyperspectral Remote Sensing of Plant Pigments. J. Exp. Bot. 2006, 58, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Im, J.; Jensen, J.T. Hyperspectral Remote Sensing of Vegetation. Geogr. Compass 2008, 2, 1943–1961. [Google Scholar] [CrossRef]
- Wong, C.Y.S.; Gilbert, M.J.; Pierce, M.A.; Parker, T.; Palkovic, A.; Gepts, P.; Magney, T.S.; Buckley, T.N. Hyperspectral Remote Sensing for Phenotyping the Physiological Drought Response of Common and Tepary Bean. Plant Phenomics 2023, 16, 0021. [Google Scholar] [CrossRef] [PubMed]
- Mertens, S.; Verbraeken, L.; Sprenger, H.; Demuynck, K.; Maleux, K.; Cannoot, B.; Block, J.D.; Maere, S.; Nelissen, H.; Bonaventure, G.; et al. Proximal Hyperspectral Imaging Detects Diurnal and Drought-Induced Changes in Maize Physiology. Front. Plant Sci. 2021, 12, 640914. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, H.; Wang, X.F.; Liu, C.T.; Wang, Y.; Huang, P.M. A Hyperspectral Remote Sensing Fusion Technology Based on Spectral Normalization of Gf and Zy Series Satellites. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2022, 43, 509–514. [Google Scholar] [CrossRef]
- Xu, N.X.; Tian, J.; Tian, Q.J.; Xu, K.J.; Tang, S.F. Analysis of Vegetation Red Edge with Different Illuminated/Shaded Canopy Proportions and to Construct Normalized Difference Canopy Shadow Index. Remote Sens. 2019, 11, 1192. [Google Scholar] [CrossRef]
- Loggenberg, K.; Strever, A.; Greyling, B.; Poona, N. Modelling Water Stress in a Shiraz Vineyard Using Hyperspectral Imaging and Machine Learning. Remote Sens. 2018, 10, 202. [Google Scholar] [CrossRef]
- Maimaitiyiming, M.; Ghulam, A.; Bozzolo, A.; Wilkins, J.L.; Kwasniewski, M.T. Early Detection of Plant Physiological Responses to Different Levels of Water Stress Using Reflectance Spectroscopy. Remote Sens. 2017, 9, 745. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Rueda, C.A.; Ustin, S.L. Water content estimation in vegetation with MODIS reflectance data and model inversion methods. Remote Sens. Environ. 2003, 85, 109–124. [Google Scholar] [CrossRef]
- Feret, J.B.; Francois, C.; Gitelson, A.; Asner, G.P.; Barry, K.M.; Panigada, C.; Richardson, A.D.; Jacquemoud, S. Optimizing spectral indices and chemometric analysis of leaf chemical properties using radiative transfer modeling. Remote Sens. Environ. 2011, 115, 2742–2750. [Google Scholar] [CrossRef]
- Stimson, H.C.; Breshears, D.D.; Ustin, S.L.; Kefauver, S.C. Spectral sensing of foliar water conditions in two co-occurring conifer species: Pinus edulis and Juniperus monosperma. Remote Sens. Environ. 2005, 96, 108–118. [Google Scholar] [CrossRef]
- Imanishi, J.; Sugimoto, K.; Morimoto, Y. Detecting drought status and LAI of two Quercus species canopies using derivative spectra. Comput. Electron. Agric. 2004, 43, 109–129. [Google Scholar] [CrossRef]
- Elvanidi, A.; Katsoulas, N.; Ferentinos, K.P.; Bartzanas, T.; Kittas, C. Hyperspectral machine vision as a tool for water stress severity assessment in soilless tomato crop. Biosyst. Eng. 2018, 165, 25–35. [Google Scholar] [CrossRef]
- Krishna, G.; Sahoo, R.N.; Singh, P.; Bajpai, V.; Patra, H.; Kumar, S.; Dandapani, R.; Gupta, V.K.; Viswanathan, C.; Ahmad, T.; et al. Comparison of various modelling approaches for water deficit stress monitoring in rice crop through hyperspectral remote sensing. Agric. Water Manag. 2019, 213, 231–244. [Google Scholar] [CrossRef]
- Gerhards, M.; Schlerf, M.; Mallick, K.; Udelhoven, T. Challenges and Future Perspectives of Multi-/Hyperspectral Thermal Infrared Remote Sensing for Crop Water-Stress Detection: A Review. Remote Sens. 2019, 11, 1240. [Google Scholar] [CrossRef]
- Kyratzis, A.C.; Skarlatos, D.P.; Menexes, G.C.; Vamvakousis, V.F.; Katsiotis, A. Assessment of Vegetation Indices Derived by UAV Imagery for Durum Wheat Phenotyping under a Water Limited and Heat Stressed Mediterranean Environment. Front. Plant Sci. 2017, 8, 1114. [Google Scholar] [CrossRef] [PubMed]
- Boulet, G.; Mougenot, B.; Lhomme, J.P.; Fanise, P.; Lili-Chabaane, Z.; Olioso, A.; Bahir, M.; Rivalland, V.; Jarlan, L.; Merlin, O.; et al. The SPARSE model for the prediction of water stress and evapotranspiration components from thermal infra-red data and its evaluation over irrigated and rainfed wheat. Hydrol. Earth Syst. Sci. 2015, 19, 4653–4672. [Google Scholar] [CrossRef]
- Jackson, R.D.; Reginato, R.J.; Idso, S.B. Wheat canopy temperature: A practical tool for evaluating water requirements. Water Resour. Res. 1977, 13, 651–656. [Google Scholar] [CrossRef]
- Becker, E.; Schmidhalter, U. Evaluation of Yield and Drought Using Active and Passive Spectral Sensing Systems at the Reproductive Stage in Wheat. Front. Plant Sci. 2017, 8, 379. [Google Scholar] [CrossRef]
- Gao, Y.; Walker, J.P.; Allahmoradi, M.; Monerris, A.; Ryu, D.; Jackson, T.J. Optical Sensing of Vegetation Water Content: A Synthesis Study. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 1456–1464. [Google Scholar] [CrossRef]
- Sow, M.; Mbow, C.; Hély, C.; Fensholt, R.; Sambou, B. Estimation of Herbaceous Fuel Moisture Content Using Vegetation Indices and Land Surface Temperature From MODIS Data. Remote Sens. 2013, 5, 2617–2638. [Google Scholar] [CrossRef]
- Ghulam, A.; Li, Z.-L.; Qin, Q.; Tong, Q.; Wang, J.; Kasimu, A.; Zhu, L. A Method for Canopy Water Content Estimation for Highly Vegetated Surfaces-Shortwave Infrared Perpendicular Water Stress Index. Sci. China Earth Sci. 2007, 50, 1359–1368. [Google Scholar] [CrossRef]
- van Eeuwijk, F.A.; Bustos-Korts, D.; Millet, E.J.; Boer, M.P.; Kruijer, W.; Thompson, A.; Malosetti, M.; Iwata, H.; Quiroz, R.; Kuppe, C.; et al. Modelling strategies for assessing and increasing the effectiveness of new phenotyping techniques in plant breeding. Plant Sci. 2019, 282, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Meacham-Hensold, K.; Guan, K.Y.; Bernacchi, C.J. Hyperspectral Leaf Reflectance as Proxy for Photosynthetic Capacities: An Ensemble Approach Based on Multiple Machine Learning Algorithms. Front. Plant Sci. 2019, 10, 730. [Google Scholar] [CrossRef] [PubMed]
- TERRA-REF Sensing Platforms. Available online: https://terraref.org/data/sensing-components (accessed on 13 September 2023).
- Sagan, V.; Maimaitijiang, M.; Paheding, S.; Bhadra, S.; Gosselin, N.; Burnette, M.; Demieville, J.; Hartling, S.; LeBauer, D.; Newcomb, M.; et al. Data-Driven Artificial Intelligence for Calibration of Hyperspectral Big Data. IEEE Trans. Geosci. Remote Sens. 2022, 60, 1–20. [Google Scholar] [CrossRef]
- Burnette, M.; Kooper, R.; Maloney, J.D.; Rohde, G.S.; Terstriep, J.A.; Willis, C.; Fahlgren, N.; Mockler, T.; Newcomb, M.; Sagan, V.; et al. TERRA-REF Data Processing Infrastructure. In Proceedings of the Practice and Experience on Advanced Research Computing, Pittsburgh, PA, USA, 22–26 July 2018; p. 27. [Google Scholar]
- Condorelli, G.E.; Newcomb, M.; Groli, E.L.; Maccaferri, M.; Forestan, C.; Babaeian, E.; Tuller, M.; White, J.W.; Ward, R.; Mockler, T.; et al. Genome Wide Association Study Uncovers the QTLome for Osmotic Adjustment and Related Drought Adaptive Traits in Durum Wheat. Genes 2022, 13, 293. [Google Scholar] [CrossRef] [PubMed]
- LeBauer, D.; Burnette, M.; Demieville, J.; Fahlgren, N.; French, A.N.; Garnett, R.; Hu, Z.; Huynh, K.; Kooper, R.; Li, Z.; et al. TERRA-REF, An Open Reference Data Set from High Resolution Genomics, Phenomics, and Imaging Sensors. 2020. Available online: https://datadryad.org/stash/dataset/doi:10.5061/dryad.4b8gtht99 (accessed on 22 November 2023).
- Bernardo, N.; Watanabe, F.; Rodrigues, T.; Alcântara, E. Atmospheric correction issues for retrieving total suspended matter concentrations in inland waters using OLI/Landsat-8 image. Adv. Space Res. 2017, 59, 2335–2348. [Google Scholar] [CrossRef]
- Maimaitijiang, M.; Sagan, V.; Bhadra, S.; Nguyen, C.; Mockler, T.C.; Shakoor, N. A fully automated and fast approach for canopy cover estimation using super high-resolution remote sensing imagery. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2021, V-3-2021, 219–226. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Maimaitiyiming, M.; Miller, A.J.; Ghulam, A. Discriminating Spectral Signatures Among and Within Two Closely Related Grapevine Species. Photogramm. Eng. Remote Sens. 2016, 82, 51–62. [Google Scholar] [CrossRef]
- Gao, B.-c. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Galvão, L.S.; Formaggio, A.R.; Tisot, D.A. Discrimination of sugarcane varieties in Southeastern Brazil with EO-1 Hyperion data. Remote Sens. Environ. 2005, 94, 523–534. [Google Scholar] [CrossRef]
- Hallett, P.D.; Gordon, D.C.; Bengough, A.G. Plant influence on rhizosphere hydraulic properties: Direct measurements using a miniaturized infiltrometer. New Phytol. 2003, 157, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Guizani, A.; Askri, H.; Amenta, M.L.; Defez, R.; Babay, E.; Bianco, C.; Rapaná, N.; Finetti-Sialer, M.; Gharbi, F. Drought responsiveness in six wheat genotypes: Identification of stress resistance indicators. Front. Plant Sci. 2023, 14, 1232583. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Anwar, S.; Ali, A.; Ullah, Z.; Binjawhar, D.N.; Sher, H.; Abdel-Hameed, U.K.; Khan, M.A.; Majeed, K.; Jaremko, M. Biochemical and phenological characterization of diverse wheats and their association with drought tolerance genes. BMC Plant Biol. 2023, 23, 326. [Google Scholar] [CrossRef]
- Islam, M.A.; Shorna, M.N.A.; Islam, S.; Biswas, S.; Biswas, J.; Islam, S.; Dutta, A.K.; Uddin, M.S.; Zaman, S.; Akhtar-E-Ekram, M.; et al. Hydrogen-rich water: A key player in boosting wheat (Triticum aestivum L.) seedling growth and drought resilience. Sci. Rep. 2023, 13, 22521. [Google Scholar] [CrossRef]
- Faghani, E.; Gharechahi, J.; Komatsu, S.; Mirzaei, M.; Khavarinejad, R.A.; Najafi, F.; Farsad, L.K.; Salekdeh, G.H. Comparative physiology and proteomic analysis of two wheat genotypes contrasting in drought tolerance. J. Proteom. 2015, 114, 1–15. [Google Scholar] [CrossRef]
- Elhag, M.; Bahrawi, J.A. Soil salinity mapping and hydrological drought indices assessment in arid environments based on remote sensing techniques. Geosci. Instrum. Methods Data Syst. 2017, 6, 149–158. [Google Scholar] [CrossRef]
- Li, M.; Chu, R.H.; Yu, Q.; Islam, A.M.T.; Chou, S.R.; Shen, S.H. Evaluating Structural, Chlorophyll-Based and Photochemical Indices to Detect Summer Maize Responses to Continuous Water Stress. Water 2018, 10, 500. [Google Scholar] [CrossRef]
- D’Odorico, P.; Schönbeck, L.; Vitali, V.; Meusburger, K.; Schaub, M.; Ginzler, C.; Zweifel, R.; Velasco, V.M.E.; Gisler, J.; Gessler, A.; et al. Drone-based physiological index reveals long-term acclimation and drought stress responses in trees. Plant Cell Environ. 2021, 44, 3552–3570. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Brown, J.F.; Verdin, J.P.; Wardlow, B. A five-year analysis of MODIS NDVI and NDWI for grassland drought assessment over the central Great Plains of the United States. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef]
- Sarlikioti, V.; Driever, S.M.; Marcelis, L.F.M. Photochemical reflectance index as a mean of monitoring early water stress. Ann. Appl. Biol. 2010, 157, 81–89. [Google Scholar] [CrossRef]
- Vélez, S.; Martínez-Peña, R.; Castrillo, D. Beyond Vegetation: A Review Unveiling Additional Insights into Agriculture and Forestry through the Application of Vegetation Indices. Multidiscip. Sci. J. 2023, 6, 421–436. [Google Scholar] [CrossRef]
- Yue, B.; Xue, W.; Xiong, L.; Yu, X.; Luo, L.; Cui, K.; Jin, D.; Xing, Y.; Zhang, Q. Genetic Basis of Drought Resistance at Reproductive Stage in Rice: Separation of Drought Tolerance from Drought Avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Sensor | Features |
|---|---|---|
| Hyperspectral | Headwall VNIR | Spectral Range: 380–1000 nm Spatial Resolution: 0.66 nm |
| Headwall SWIR | Spectral Range: 950–2500 nm Spatial Resolution: 5.8 nm | |
| Fluorescence Intensity | LemnaTec PSII (Manta G-235b) | Camera Resolution: 1936 × 1216 Center Wavelength: 710 nm |
| Moisture | Acclima TDR-310H (SDI-12) | Incident Wave Amplitude: 400 mV Volumetric Water Content: 0% to 100% |
| Sensors | Remote Sensing | Proximal Sensors | |
|---|---|---|---|
| Hyperspectral Images | RWC | ||
| Stages | Pre-Stress | 28 February (Both) | - |
| Early Drought | - | 17 March (Both) | |
| Mid Drought | 22 March (Rep 1) 23 March (Rep 2) | 26 March (Both) | |
| Post Treatment | 7 April (Rep 1) 8 April (Rep 2) | 7 April (Rep 1) 8 April (Rep 2) |
| Genotype | Origin | Plot ID (in Field) | Drought Resistance (2019) |
|---|---|---|---|
| BOLENGA | IRTA-SPAIN | DP_033 | Susceptible |
| OMRABI 3 | ICARDA | DP_070 | Susceptible |
| STOJOCRI_3 | ICARDA | DP_074 | Susceptible |
| ARCANGELO | ITALY | DP_079 | Susceptible |
| CAPEITI 8 | ITALY | DP_083 | Susceptible |
| COLORADO | DESERT | DP_086 | Susceptible |
| COLOSSEO | ITALY | DP_087 | Resistant |
| VALNOVA | ITALY | DP_114 | Resistant |
| WEST BRED 881 | DESERT | DP_116 | Resistant |
| AINZEN_1 | ICARDA | DP_119 | Susceptible |
| CHABHA 88 | ICARDA | DP_135 | Susceptible |
| GEZIRA 17 | ICARDA | DP_140 | Resistant |
| Spectral Index | Equation (in nm) | Reference |
|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | (R750 − R705)/(R750 + R705) | [46] |
| Photochemical Reflective Index (PRI) | (R531 − R570)/(R531 + R570) | [47] |
| Normalized Difference Water Index (NDWI) | (R864 − R1245)/(R864 + R1245) | [48] |
| Leaf Water Vegetation Index (LWVI-1) | (R1094 − R893)/(R1094 + R893) | [49] |
| Leaf Water Vegetation Index (LWVI-2) | (R1094 − R1205)/(R1094 + R1205) | [49] |
| Test Name | Early Stage | Mid Stage | Post Stage | |||
|---|---|---|---|---|---|---|
| F-Statistics | p-Value | F-Statistics | p-Value | F-Statistics | p-Value | |
| O’Brien | 2.94 | 0.09 | 38.59 | * | 17.43 | * |
| Brown–Forsythe | 0.23 | 0.63 | 1.43 | 0.23 | 9.97 | * |
| Levene | 0.23 | 0.63 | 1.43 | 0.23 | 9.97 | * |
| Bartlett | 1.08 | 0.30 | 1.67 | 0.19 | 14.28 | * |
| F-Test (ANOVA) | 2.94 | 0.09 | 38.58 | * | 17.43 | * |
| Stage | Treatment | Mean | Std Error | Lower 95% | Upper 95% |
|---|---|---|---|---|---|
| Early Stage | WW | 98.10 | 0.27 | 97.53 | 98.68 |
| WL | 97.49 | 0.22 | 97.03 | 97.95 | |
| Mid Stage | WW | 96.90 | 0.30 | 96.25 | 97.54 |
| WL | 93.71 | 0.40 | 92.87 | 94.56 | |
| Post Stage | WW | 96.87 | 0.29 | 96.25 | 97.50 |
| WL | 93.72 | 0.69 | 92.28 | 95.15 |
| Stage | Early Stage | Mid Stage | Post Stage |
|---|---|---|---|
| Difference | 0.61 | 3.18 | 3.15 |
| Std Error Difference | 0.35 | 0.51 | 0.75 |
| Upper CL Diff | 1.31 | 4.18 | 4.64 |
| Lower CL Diff | −0.08 | 2.17 | 1.67 |
| Confidence Level | 0.95 | 0.95 | 0.95 |
| t-Ratio | 1.71 | 6.21 | 4.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, B.; Sagan, V.; Haireti, A.; Newcomb, M.; Tuberosa, R.; LeBauer, D.; Shakoor, N. Early Detection of Drought Stress in Durum Wheat Using Hyperspectral Imaging and Photosystem Sensing. Remote Sens. 2024, 16, 155. https://doi.org/10.3390/rs16010155
Roy B, Sagan V, Haireti A, Newcomb M, Tuberosa R, LeBauer D, Shakoor N. Early Detection of Drought Stress in Durum Wheat Using Hyperspectral Imaging and Photosystem Sensing. Remote Sensing. 2024; 16(1):155. https://doi.org/10.3390/rs16010155
Chicago/Turabian StyleRoy, Bishal, Vasit Sagan, Alifu Haireti, Maria Newcomb, Roberto Tuberosa, David LeBauer, and Nadia Shakoor. 2024. "Early Detection of Drought Stress in Durum Wheat Using Hyperspectral Imaging and Photosystem Sensing" Remote Sensing 16, no. 1: 155. https://doi.org/10.3390/rs16010155
APA StyleRoy, B., Sagan, V., Haireti, A., Newcomb, M., Tuberosa, R., LeBauer, D., & Shakoor, N. (2024). Early Detection of Drought Stress in Durum Wheat Using Hyperspectral Imaging and Photosystem Sensing. Remote Sensing, 16(1), 155. https://doi.org/10.3390/rs16010155