
Combining Sentinel-2 Data and Risk Maps to Detect Trees Predisposed to and Attacked by European Spruce Bark Beetle





Abstract
:1. Introduction
- (1)
- How does detection accuracy change with time after bark beetle swarming?
- (2)
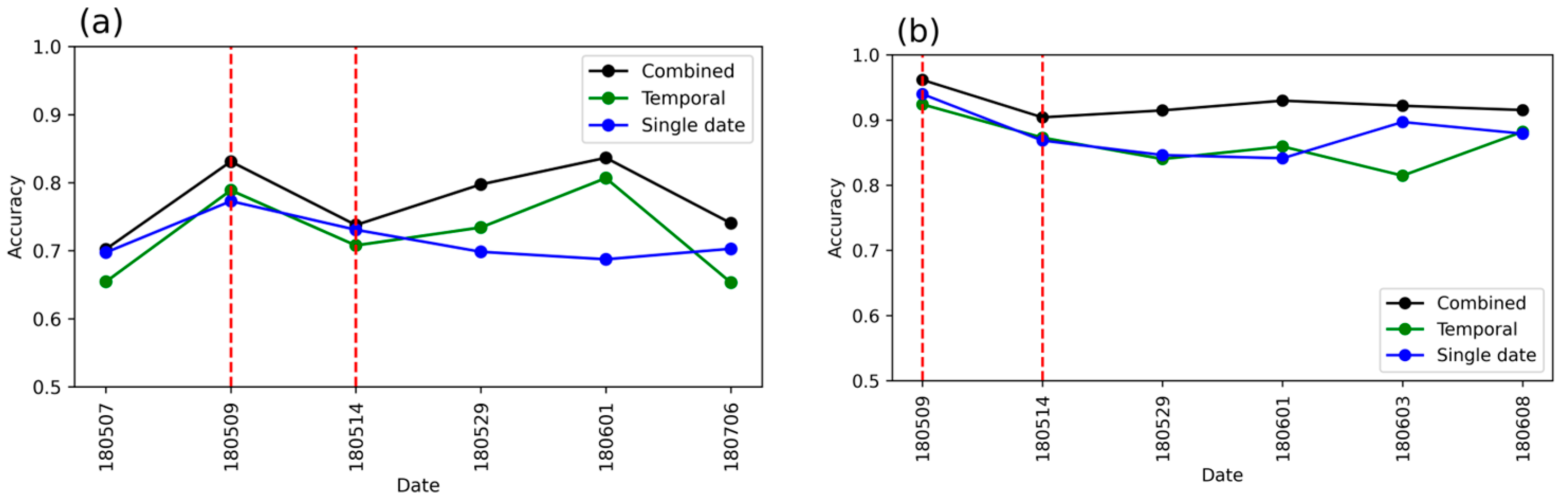
- Does accuracy increase when single-date and temporal features are combined?
- (3)
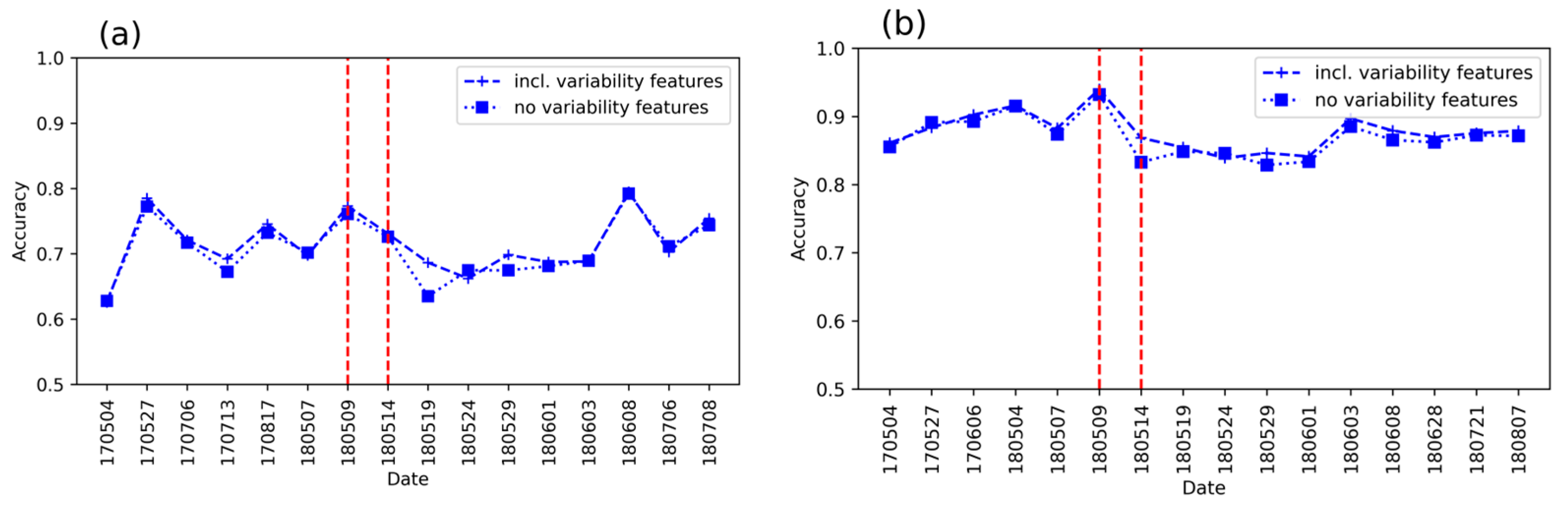
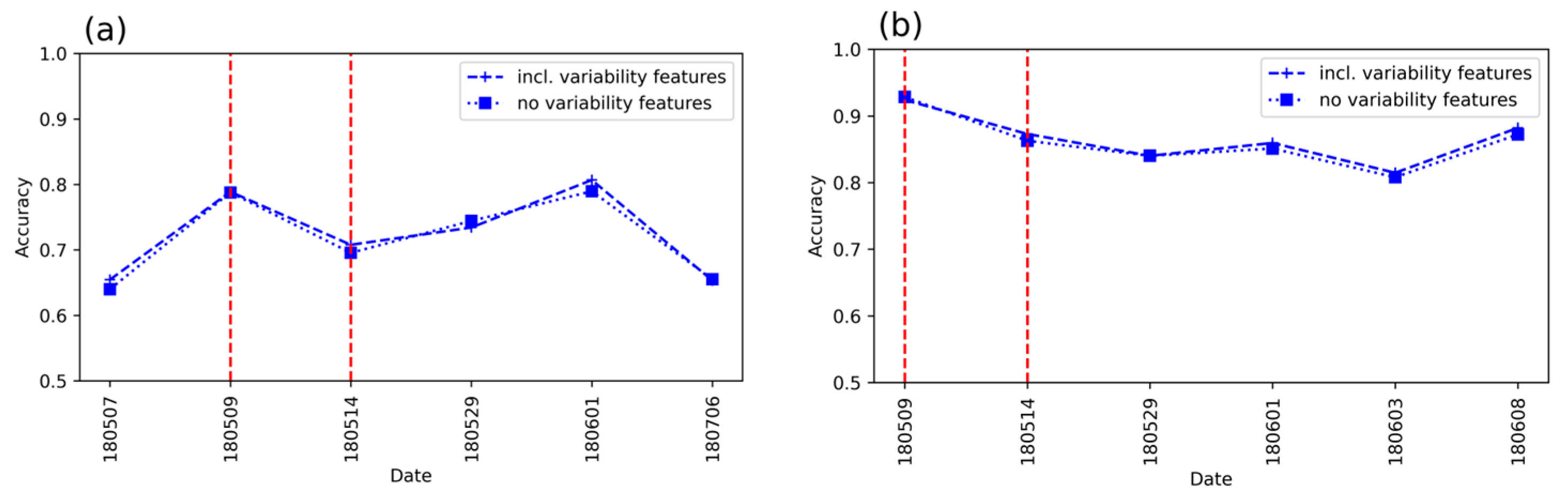
- Does the inclusion of spatial variability increase detection accuracy?
- (4)
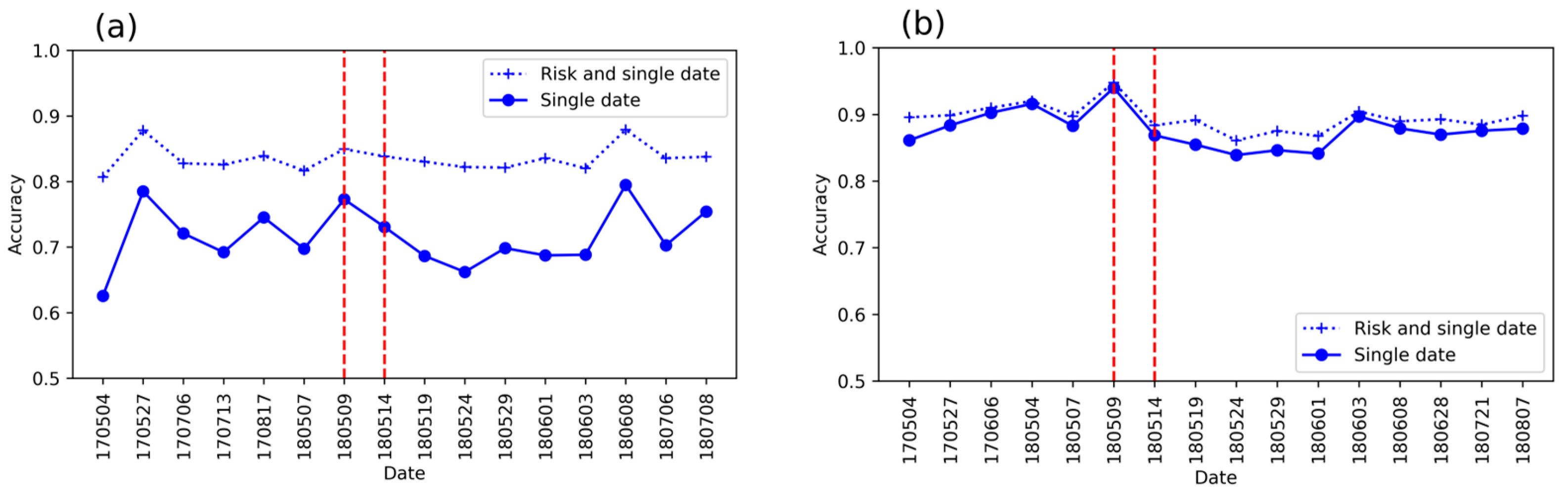
- Does the inclusion of a risk map increase detection accuracy?
- (5)
- Which wavelength bands are more important for detection of trees predisposed to and attacked by bark beetle?
2. Materials and Methods
2.1. Study Area

2.2. Data and Pre-Processing
2.2.1. Sentinel-2 Data
2.2.2. Bark Beetle Damage Data
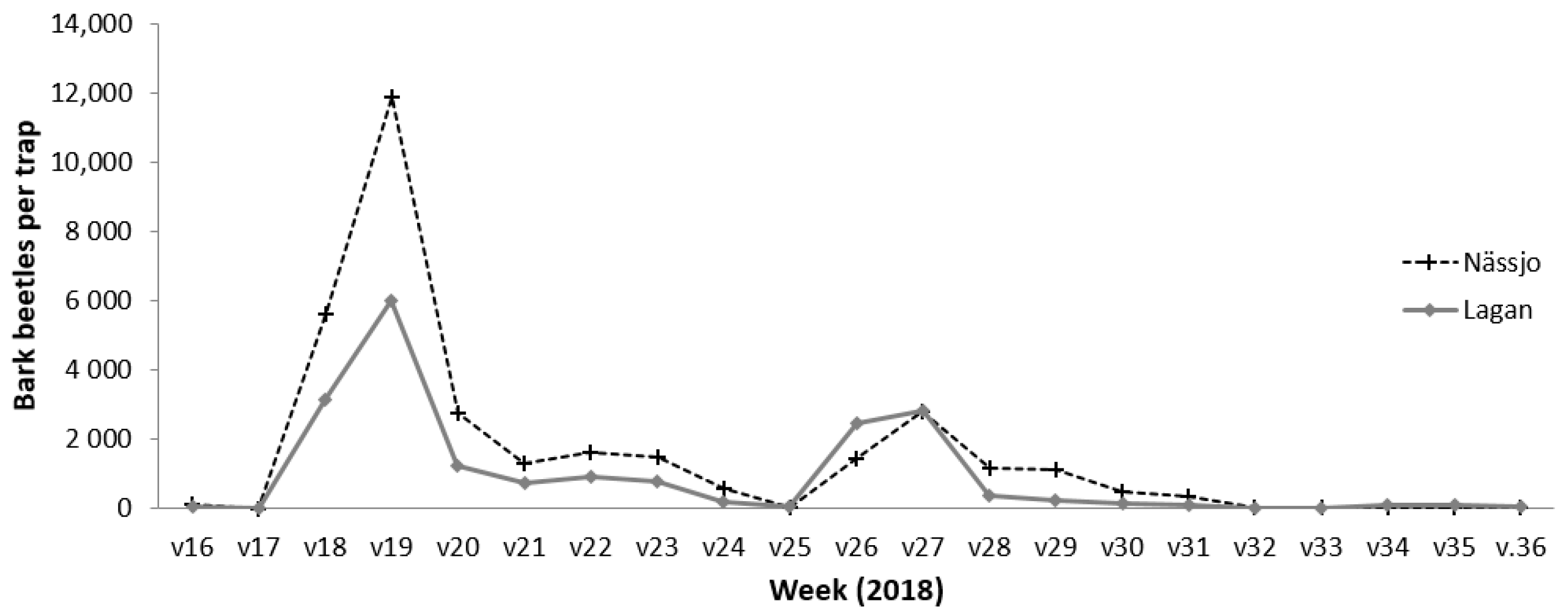
2.2.3. Bark Beetle Swarming Data
2.2.4. Bark Beetle Risk Map
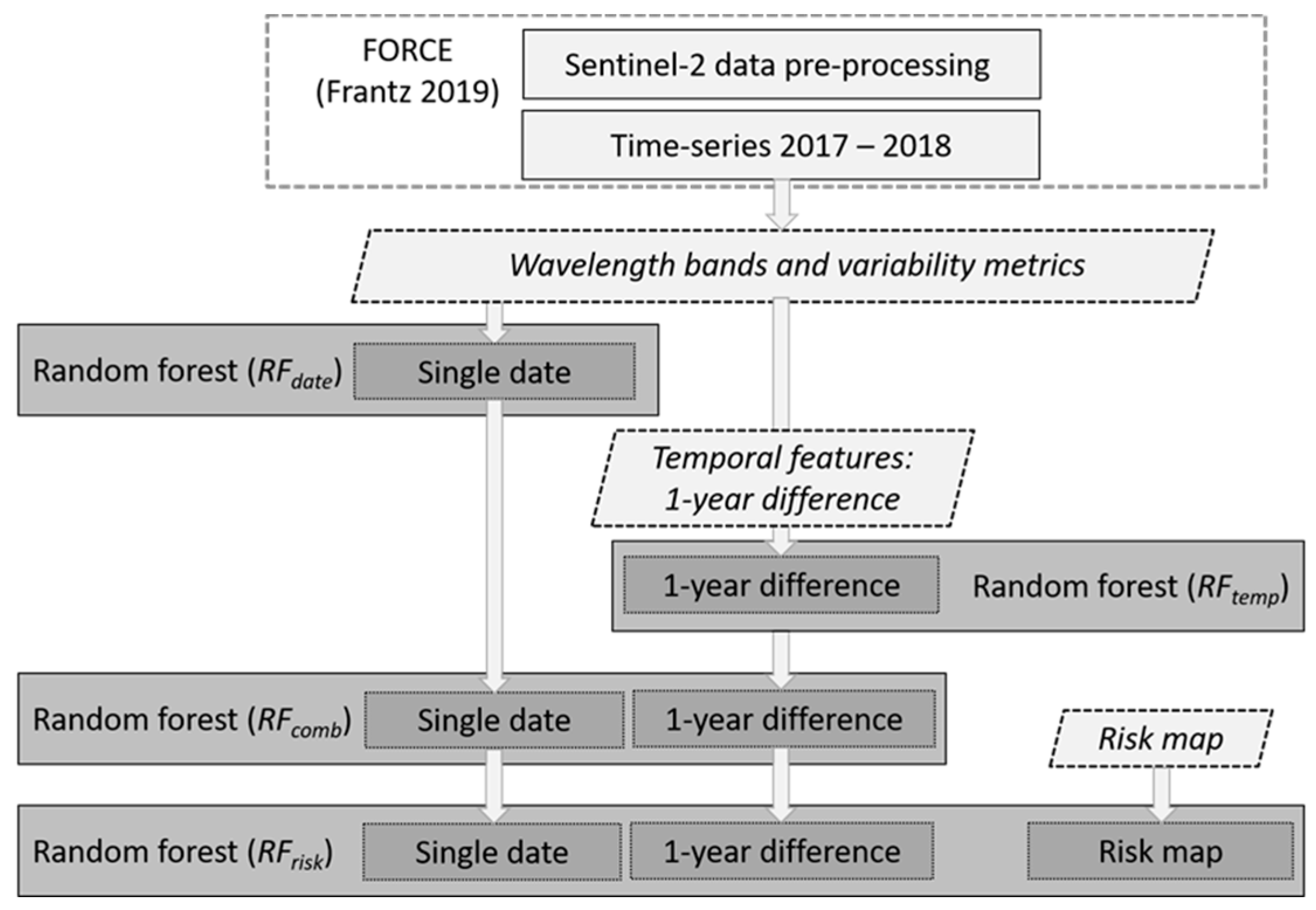
2.3. Bark Beetle Attack Detection with Random Forest
- models including only data from a single date (RFdate)
- models with only temporal features (RFtemp)
- models including both single-date and temporal features (RFcomb)
- models that in addition to single-date and temporal features also include the risk map (RFrisk)
2.3.1. Sentinel-2-Derived Features for the Random Forest Models
- Wavelength bands
- Variability metrics
- Temporal features
2.3.2. Random Forest
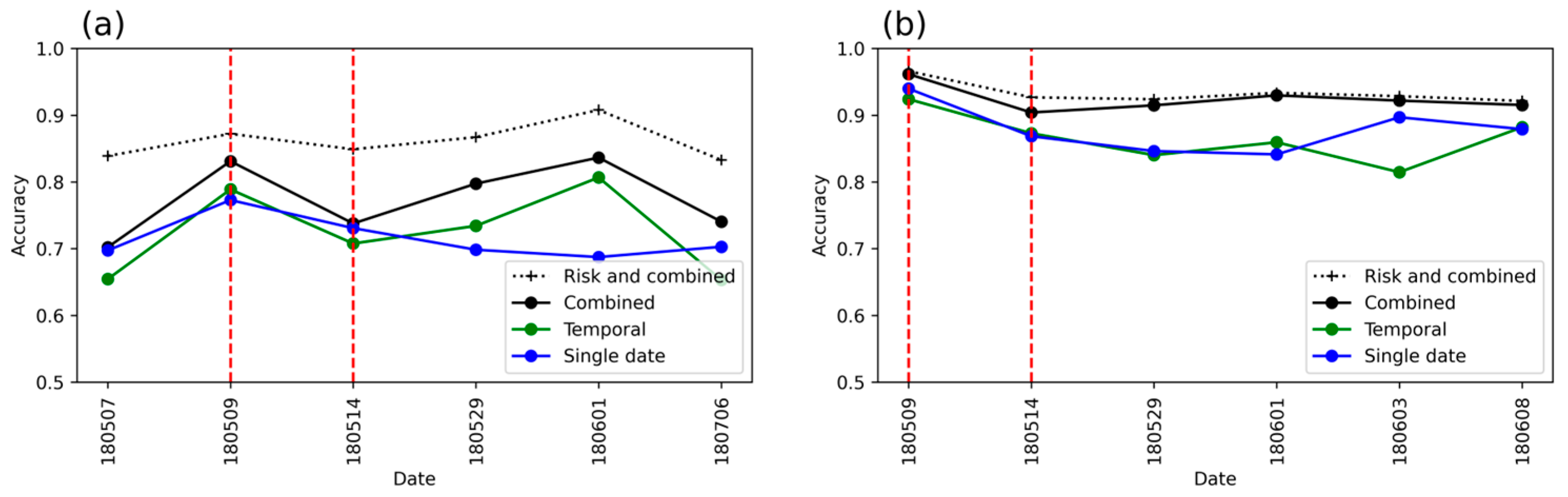
3. Results
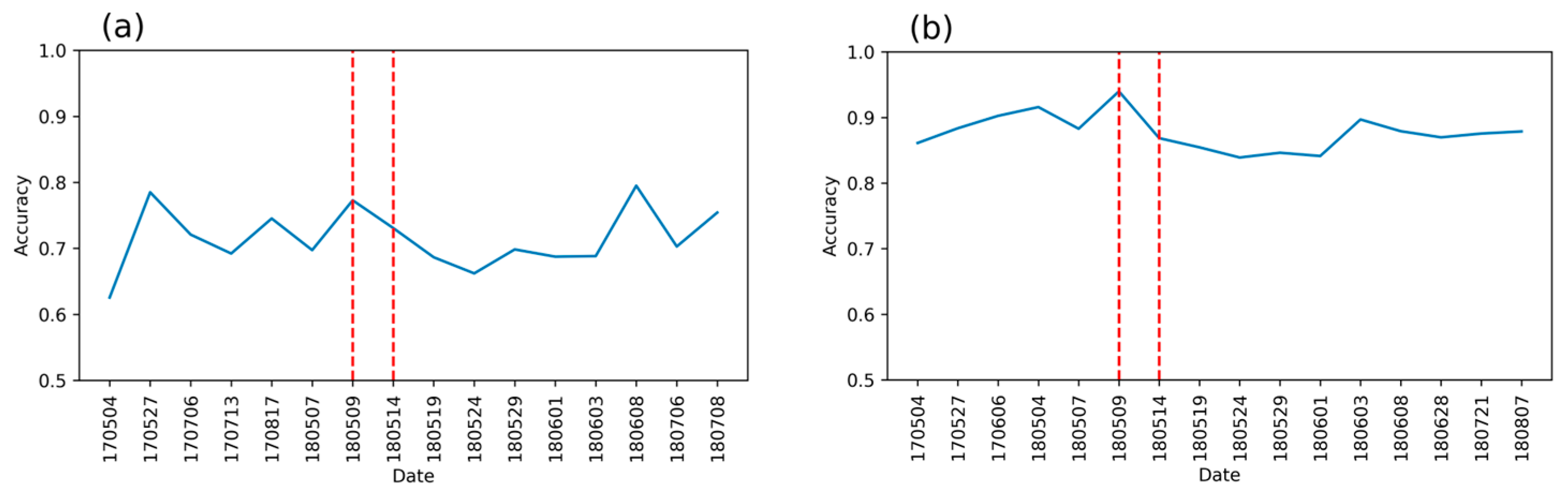
3.1. Detection Accuracy for Single-Date (RFdate) and Temporal (RFtemp) Models
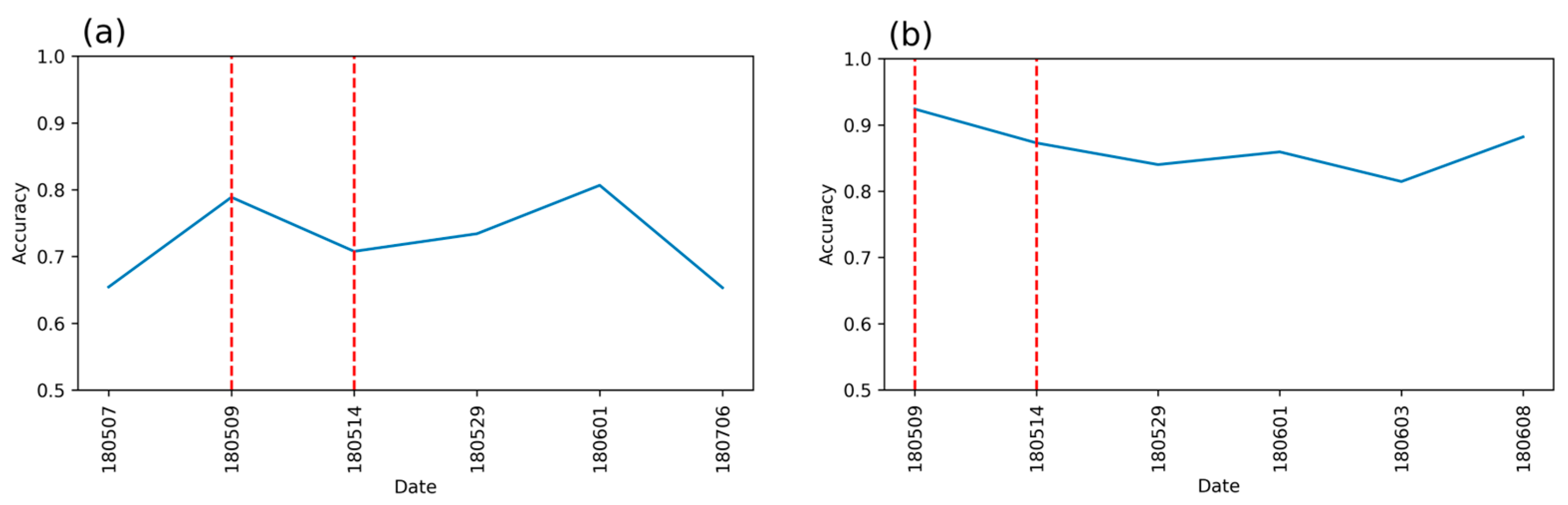
3.2. Detection Accuracy with Single-Date and Temporal Features Combined (RFcomb)
3.3. Detection Accuracy with and Without the Variability Metrics
3.4. Detection Accuracy with the Risk Map Included
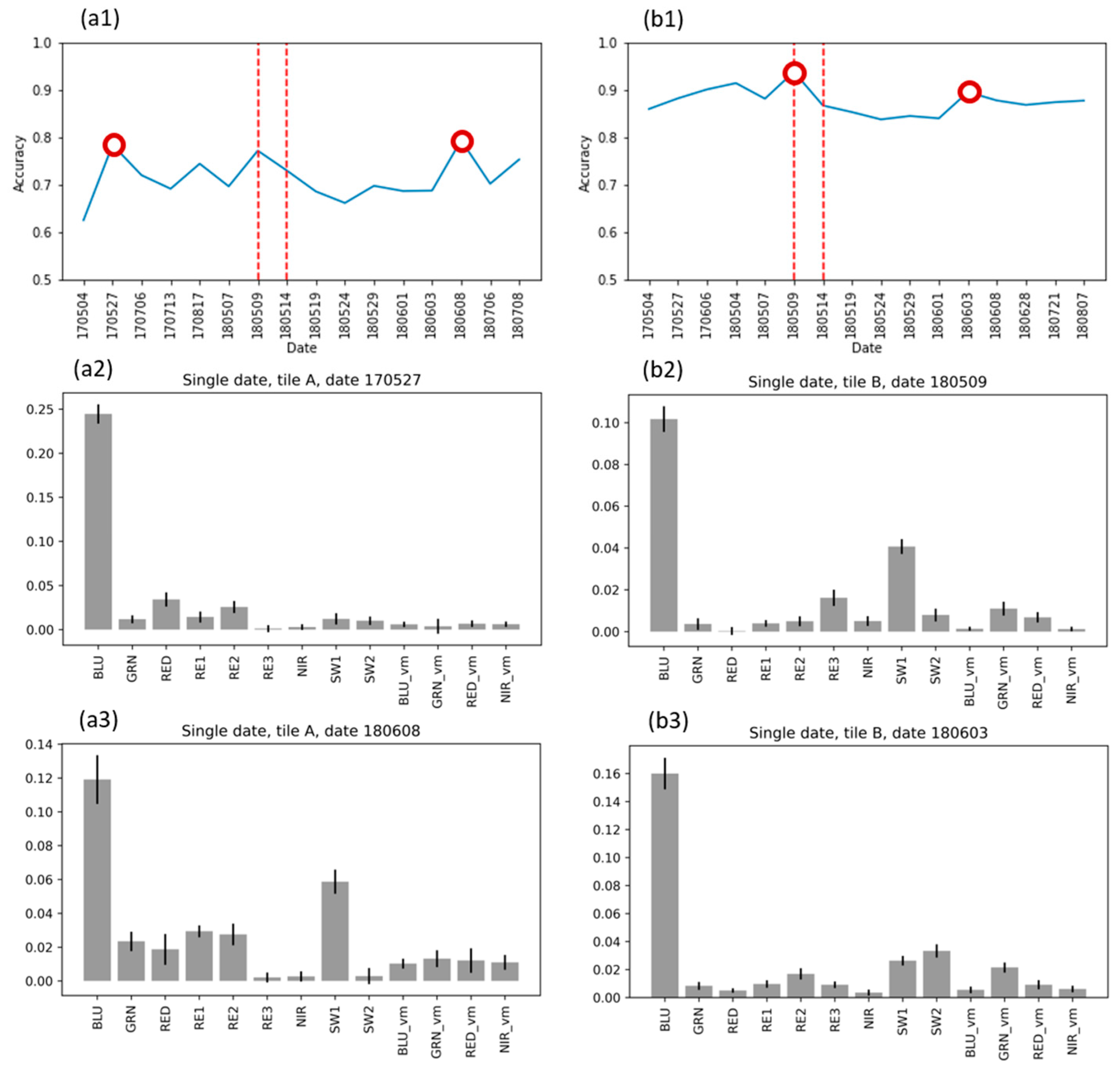
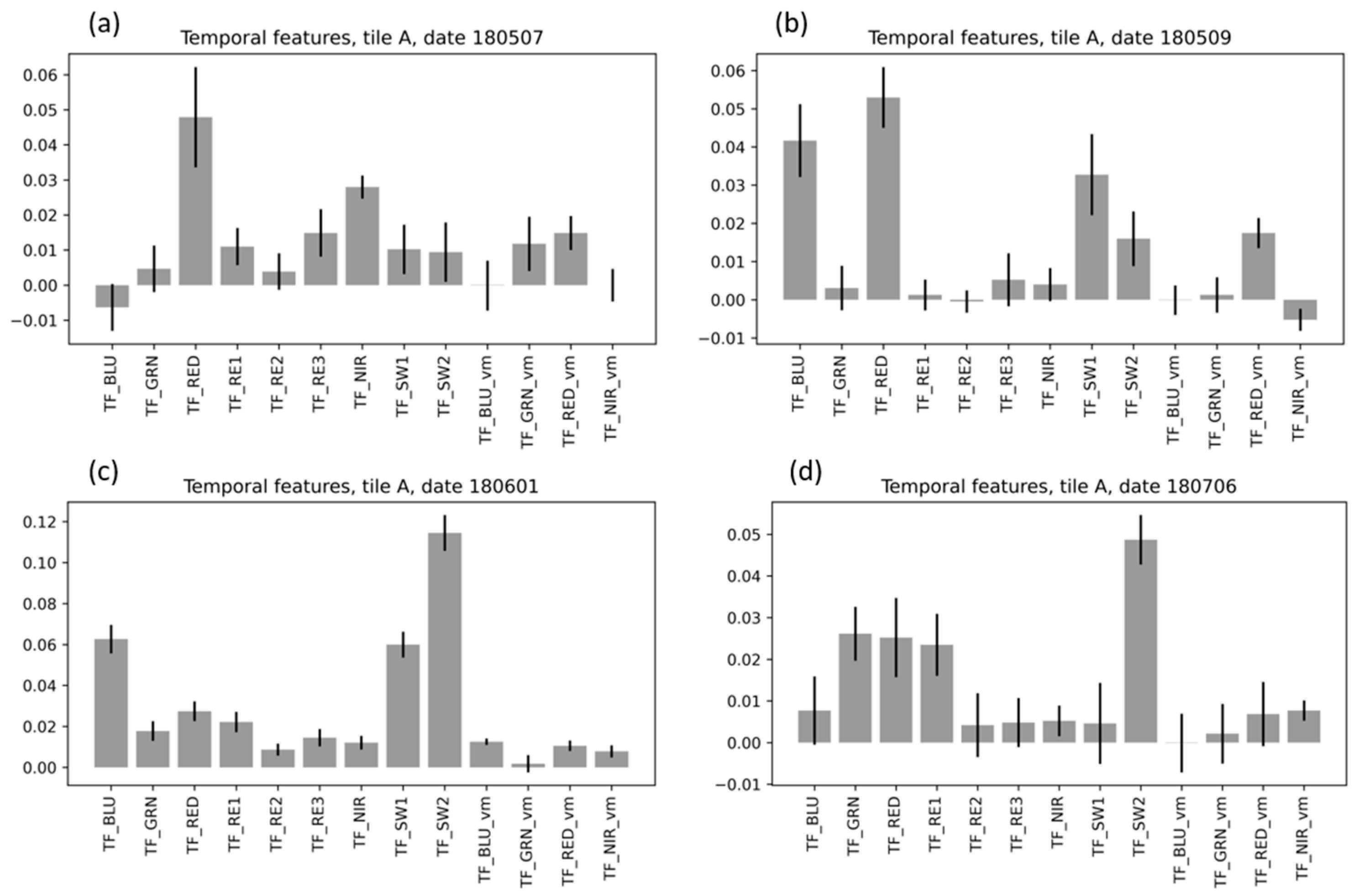
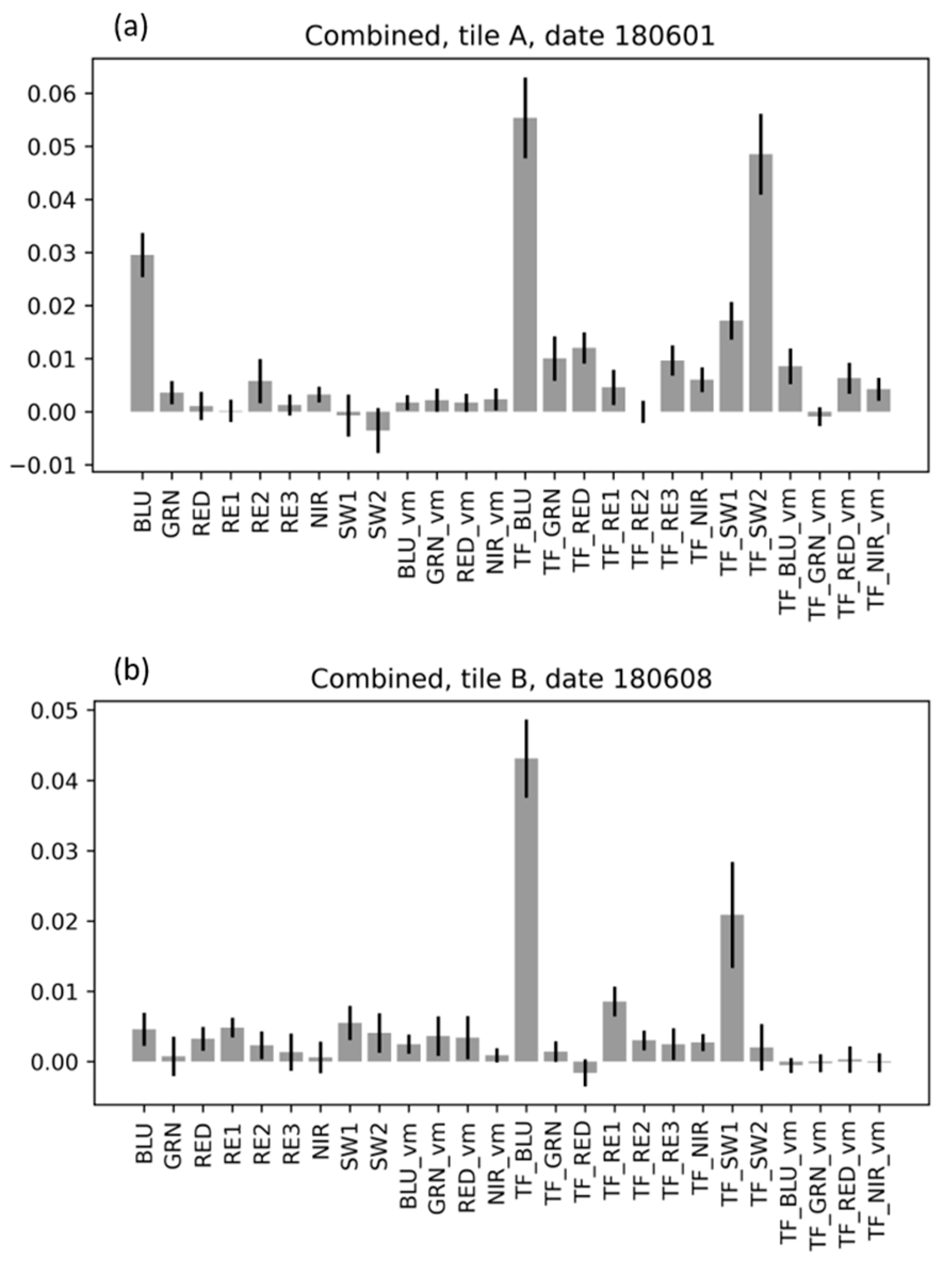
3.5. Analysis of Feature Importance
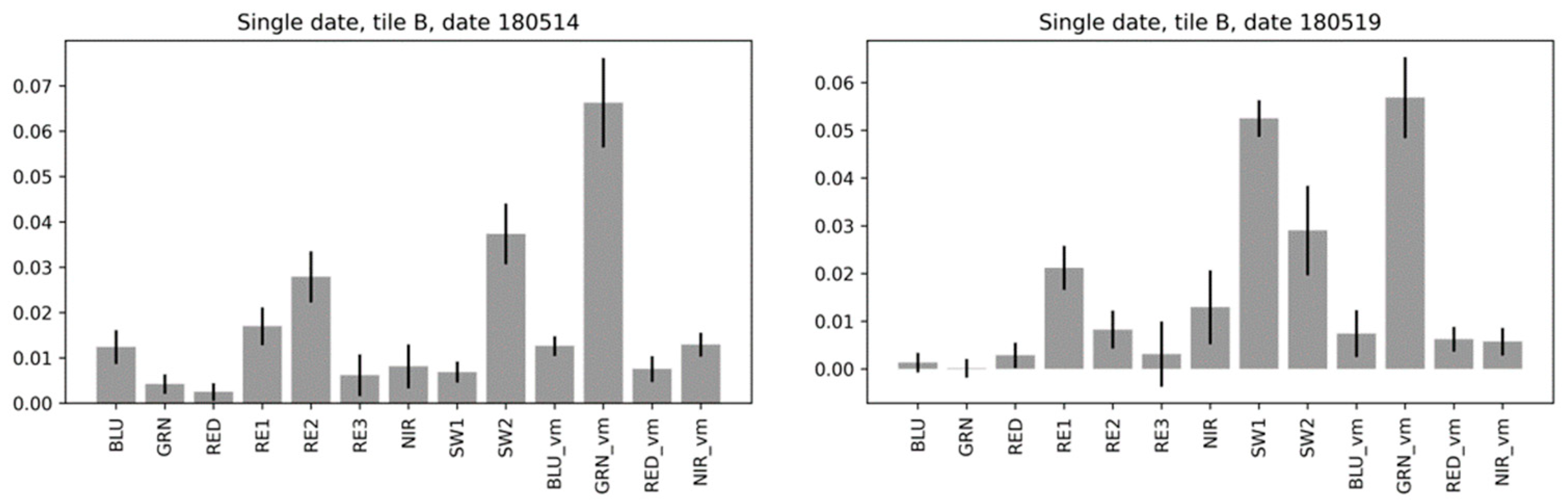
3.5.1. Feature Importance for Single-Date Random Forest Models
3.5.2. Feature Importance for Temporal Random Forest Models
3.5.3. Feature Importance for the Combined Random Forest Models
3.5.4. Feature Importance for the Variability Metrics and Risk Map
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant increase in natural disturbance impacts on European forests since 1950. Glob. Chang. Biol. 2023, 29, 1359–1376. [Google Scholar] [CrossRef] [PubMed]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Jactel, H.; Petit, J.; Desprez-Loustau, M.-L.; Delzon, S.; Piou, D.; Battisti, A.; Koricheva, J. Drought effects on damage by forest insects and pathogens: A meta-analysis. Glob. Chang. Biol. 2012, 18, 267–276. [Google Scholar] [CrossRef]
- Netherer, S.; Panassiti, B.; Pennerstorfer, J.; Matthews, B. Acute Drought Is an Important Driver of Bark Beetle Infestation in Austrian Norway Spruce Stands. Front. For. Glob. Chang. 2019, 2, 39. [Google Scholar] [CrossRef]
- Marini, L.; Lindelöw, Å.; Jönsson, A.M.; Wulff, S.; Schroeder, L.M. Population dynamics of the spruce bark beetle: A long-term study. Oikos 2013, 122, 1768–1776. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Schroeder, L.M.; Lagergren, F.; Anderbrant, O.; Smith, B. Guess the impact of Ips typographus—An ecosystem modelling approach for simulating spruce bark beetle outbreaks. Agric. For. Meteorol. 2012, 166–167, 188–200. [Google Scholar] [CrossRef]
- Bentz, B.J.; Jönsson, A.M.; Schroeder, M.; Weed, A.; Wilcke, R.A.I.; Larsson, K. Ips typographus and Dendroctonus ponderosae Models Project Thermal Suitability for Intra- and Inter-Continental Establishment in a Changing Climate. Front. For. Glob. Chang. 2019, 2, 1. [Google Scholar] [CrossRef]
- Müller, M.; Olsson, P.O.; Eklundh, L.; Jamali, S.; Ardö, J. Features predisposing forest to bark beetle outbreaks and their dynamics during drought. For. Ecol. Manag. 2022, 523, 120480. [Google Scholar] [CrossRef]
- Kärvemo, S.; Huo, L.; Öhrn, P.; Lindberg, E.; Persson, H.J. Different triggers, different stories: Bark-beetle infestation patterns after storm and drought-induced outbreaks. For. Ecol. Manag. 2023, 545, 121255. [Google Scholar] [CrossRef]
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Leckie, D.G.; Carroll, A.L. Surveying mountain pine beetle damage of forests: A review of remote sensing opportunities. For. Ecol. Manag. 2006, 221, 27–41. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M. Sentinel-2 accurately maps green-attack stage of European spruce bark beetle (Ips typographus, L.) compared with Landsat-8. Remote Sens. Ecol. Conserv. 2018, 5, 87–106. [Google Scholar] [CrossRef]
- König, S.; Thonfeld, F.; Förster, M.; Dubovyk, O.; Heurich, M. Assessing Combinations of Landsat, Sentinel-2 and Sentinel-1 Time series for Detecting Bark Beetle Infestations. GIScience Remote Sens. 2023, 60, 2226515. [Google Scholar] [CrossRef]
- Huo, L.; Lindberg, E.; Bohlin, J.; Persson, H.J. Assessing the detectability of European spruce bark beetle green attack in multispectral drone images with high spatial- and temporal resolutions. Remote Sens. Environ. 2023, 287, 113484. [Google Scholar] [CrossRef]
- Bárta, V.; Lukeš, P.; Homolová, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Dalponte, M.; Solano-Correa, Y.T.; Frizzera, L.; Gianelle, D. Mapping a European Spruce Bark Beetle Outbreak Using Sentinel-2 Remote Sensing Data. Remote Sens. 2022, 14, 3135. [Google Scholar] [CrossRef]
- Candotti, A.; De Giglio, M.; Dubbini, M.; Tomelleri, E. A Sentinel-2 Based Multi-Temporal Monitoring Framework for Wind and Bark Beetle Detection and Damage Mapping. Remote Sens. 2022, 14, 6105. [Google Scholar] [CrossRef]
- Jamali, S.; Olsson, P.-O.; Ghorbanian, A.; Müller, M. Examining the potential for early detection of spruce bark beetle attacks using multi-temporal Sentinel-2 and harvester data. ISPRS J. Photogramm. Remote Sens. 2023, 205, 352–366. [Google Scholar] [CrossRef]
- Olsson, P.O.; Bergman, H.; Piltz, K. Exploring the potential to use in-between pixel variability for early detection of bark beetle attacked trees. AGILE GIScience Ser. 2023, 4, 35. [Google Scholar] [CrossRef]
- Jamali, S.; Olsson, P.O.; Müller, M.; Ghorbanian, A. Kernel-Based Early Detection of Forest Bark Beetle Attack Using Vegetation Indices Time Series of Sentinel-2. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2024, 17, 12868–12877. [Google Scholar] [CrossRef]
- Kautz, M.; Feurer, J.; Adler, P. Early detection of bark beetle (Ips typographus) infestations by remote sensing—A critical review of recent research. For. Ecol. Manag. 2024, 556, 121595. [Google Scholar] [CrossRef]
- Lindbladh, M.; Axelsson, A.-L.; Hultberg, T.; Brunet, J.; Felton, A. From broadleaves to spruce—The borealization of southern Sweden. Scand. J. For. Res. 2014, 29, 686–696. [Google Scholar] [CrossRef]
- SMHI. Månads-, Årstids- och Årskartor. 2024. Available online: https://www.smhi.se/data/meteorologi/kartor/normal/arstidsmedeltemperatur-normal/sommar (accessed on 20 August 2024).
- Schroeder, M. Granbarkborrens Förökningsframgång i Dödade Träd Under Sommaren 2022 i Sydöstra Småland, Värmland och Uppland/Västmanland. Swedish University of Agricultural Sciences. 2023. Available online: https://www.slu.se/globalassets/ew/org/inst/ekol/rapporter/granbarkborrens-forokningsframgang-2022.pdf (accessed on 29 December 2023).
- Lantmäteriet. GSD-Fastighetskartan Vektor Version 7.5.5. 2021. Available online: https://www.lantmateriet.se/sv/kartor/ (accessed on 5 October 2021).
- OpenStreetMap Contributors. OpenStreetMap’s Standard Tile Layer. 2024. Available online: https://tile.openstreetmap.org/ (accessed on 16 April 2024).
- Frantz, D. FORCE—Landsat + Sentinel-2 Analysis Ready Data and Beyond. Remote Sens. 2019, 11, 1124. [Google Scholar] [CrossRef]
- Tanré, D.; Herman, M.; Deschamps, P.Y.; de Leffe, A. Atmospheric modeling for space measurements of ground reflectances, including bidirectional properties. Appl. Opt. 1979, 18, 3587–3594. [Google Scholar] [CrossRef]
- Tanré, D.; Deroo, C.; Duhaut, P.; Herman, M.; Morcrette, J.J.; Perbos, J.; Deschamps, P.Y. Technical note Description of a computer code to simulate the satellite signal in the solar spectrum: The 5S code. Int. J. Remote Sens. 1990, 11, 659–668. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, S.; Woodcock, C.E. Improvement and expansion of the Fmask algorithm: Cloud, cloud shadow, and snow detection for Landsats 4–7, 8, and Sentinel 2 images. Remote Sens. Environ. 2015, 159, 269–277. [Google Scholar] [CrossRef]
- Zhu, Z.; Woodcock, C.E. Object-based cloud and cloud shadow detection in Landsat imagery. Remote Sens. Environ. 2012, 118, 83–94. [Google Scholar] [CrossRef]
- Frantz, D.; Haß, E.; Uhl, A.; Stoffels, J.; Hill, J. Improvement of the Fmask algorithm for Sentinel-2 images: Separating clouds from bright surfaces based on parallax effects. Remote Remote Sens. Environ. 2018, 215, 471–481. [Google Scholar] [CrossRef]
- Royer, A.; Charbonneau, L.; Teillet, P.M. Interannual landsat-MSS reflectance variation in an urbanized temperate zone. Remote Sens. Environ. 1988, 24, 423–446. [Google Scholar] [CrossRef]
- Hill, J. High Precision Land Cover Mapping and Inventory with Multi-Temporal Earth Observation Satellite Data: The Ardèche Experiment. Ph.D. Thesis, Trier University, Trier, Germany, 1993. [Google Scholar]
- Sobolev, V.V.; Haar, D. Light Scattering in Planetary Atmospheres: International Series of Monographs in Natural Philosophy; Elsevier Science: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Frantz, D.; Stellmes, M.; Hostert, P. A Global MODIS Water Vapor Database for the Operational Atmospheric Correction of Historic and Recent Landsat Imagery. Remote Sens. 2019, 11, 257. [Google Scholar] [CrossRef]
- Roy, D.P.; Li, J.; Zhang, H.K.; Yan, L.; Huang, H.; Li, Z. Examination of Sentinel-2A multi-spectral instrument (MSI) reflectance anisotropy and the suitability of a general method to normalize MSI reflectance to nadir BRDF adjusted reflectance. Remote Sens. Environ. 2017, 199, 25–38. [Google Scholar] [CrossRef]
- Roy, D.P.; Li, Z.; Zhang, H.K. Adjustment of Sentinel-2 Multi-Spectral Instrument (MSI) Red-Edge Band Reflectance to Nadir BRDF Adjusted Reflectance (NBAR) and Quantification of Red-Edge Band BRDF Effects. Remote Sens. 2017, 9, 1325. [Google Scholar] [CrossRef]
- Roy, D.P.; Zhang, H.K.; Ju, J.; Gomez-Dans, J.L.; Lewis, P.E.; Schaaf, C.B.; Sun, Q.; Li, J.; Huang, H.; Kovalskyy, V. A general method to normalize Landsat reflectance data to nadir BRDF adjusted reflectance. Remote Sens. Environ. 2016, 176, 255–271. [Google Scholar] [CrossRef]
- Rufin, P.; Frantz, D.; Yan, L.; Hostert, P. Operational Coregistration of the Sentinel-2A/B Image Archive Using Multitemporal Landsat Spectral Averages. IEEE Geosci. Remote Sens. Lett. 2021, 18, 712–716. [Google Scholar] [CrossRef]
- FORCE. FORCE Time Series Analysis. 2023. Available online: https://force-eo.readthedocs.io/en/latest/components/higher-level/tsa/index.html (accessed on 2 January 2024).
- SMHI. April 2018—Årets Första Vårmånad. 2018. Available online: https://www.smhi.se/klimat/klimatet-da-och-nu/manadens-vader-och-vatten-sverige/manadens-vader-i-sverige/april-2018-arets-forsta-varmanad-1.133032 (accessed on 3 January 2023).
- Naturvårdsverket. National Land Cover Database (NMD). 2021. Available online: https://www.naturvardsverket.se/en/services-and-permits/maps-and-map-services/national-land-cover-database/ (accessed on 20 August 2021).
- Lantmäteriet. GSD-Terrängkartan, Vektor. Version 5.10. 2021. Available online: https://www.lantmateriet.se/globalassets/geodata/geodataprodukter/produktlista/terrshmi.pdf (accessed on 5 January 2021).
- Skogsstyrelsen. Svarmningsövervakning. 2023. Available online: https://www.skogsstyrelsen.se/statistik/statistik-efter-amne/svarmningsovervakning/ (accessed on 20 August 2024).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Altmann, A.; Toloşi, L.; Sander, O.; Lengauer, T. Permutation importance: A corrected feature importance measure. Bioinformatics 2010, 26, 1340–1347. [Google Scholar] [CrossRef]
- Junttila, S.; Näsi, R.; Koivumäki, N.; Imangholiloo, M.; Saarinen, N.; Raisio, J.; Holopainen, M.; Hyyppä, H.; Hyyppä, J.; Lyytikäinen-Saarenmaa, P.; et al. Multispectral Imagery Provides Benefits for Mapping Spruce Tree Decline Due to Bark Beetle Infestation When Acquired Late in the Season. Remote Sens. 2022, 14, 909. [Google Scholar] [CrossRef]
- Trubin, A.; Kozhoridze, G.; Zabihi, K.; Modlinger, R.; Singh, V.V.; Surový, P.; Jakuš, R. Detection of susceptible Norway spruce to bark beetle attack using PlanetScope multispectral imagery. Front. For. Glob. Chang. 2023, 6, 1130721. [Google Scholar] [CrossRef]
- Vicca, S.; Balzarolo, M.; Filella, I.; Granier, A.; Herbst, M.; Knohl, A.; Longdoz, B.; Mund, M.; Nagy, Z.; Pintér, K.; et al. Remotely-sensed detection of effects of extreme droughts on gross primary production. Sci. Rep. 2016, 6, 28269. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, F.E.; Latifi, H.; Ghosh, A.; Joshi, P.K.; Koch, B. Assessing the potential of hyperspectral imagery to map bark beetle-induced tree mortality. Remote Sens. Environ. 2014, 140, 533–548. [Google Scholar] [CrossRef]
- Abdullah, H.; Darvishzadeh, R.; Skidmore, A.K.; Groen, T.A.; Heurich, M. European spruce bark beetle (Ips typographus, L.) green attack affects foliar reflectance and biochemical properties. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 199–209. [Google Scholar] [CrossRef]
- Huo, L.; Lindberg, E.; Fransson, J.E.S.; Persson, H.J. Comparing Spectral Differences Between Healthy and Early Infested Spruce Forests Caused by Bark Beetle Attacks using Satellite Images. In Proceedings of the IGARSS 2022—2022 IEEE International Geoscience and Remote Sensing Symposium, Kuala Lumpur, Malaysia, 17–22 July 2022; pp. 7709–7712. [Google Scholar]
- Huo, L.; Koivumäki, N.; Oliveira, R.A.; Hakala, T.; Markelin, L.; Näsi, R.; Suomalainen, J.; Polvivaara, A.; Junttila, S.; Honkavaara, E. Bark beetle pre-emergence detection using multi-temporal hyperspectral drone images: Green shoulder indices can indicate subtle tree vitality decline. ISPRS J. Photogramm. Remote Sens. 2024, 216, 200–216. [Google Scholar] [CrossRef]
- Zhen, Z.; Chen, S.; Yin, T.; Gastellu-Etchegorry, J.-P. Globally quantitative analysis of the impact of atmosphere and spectral response function on 2-band enhanced vegetation index (EVI2) over Sentinel-2 and Landsat-8. ISPRS J. Photogramm. Remote Sens. 2023, 205, 206–226. [Google Scholar] [CrossRef]
- Olsson, P.-O.; Lindström, J.; Eklundh, L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI. Remote Sens. Environ. 2016, 181, 42–53. [Google Scholar] [CrossRef]
- Georganos, S.; Grippa, T.; Niang Gadiaga, A.; Linard, C.; Lennert, M.; Vanhuysse, S.; Mboga, N.; Wolff, E.; Kalogirou, S. Geographical random forests: A spatial extension of the random forest algorithm to address spatial heterogeneity in remote sensing and population modelling. Geocarto Int. 2021, 36, 121–136. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Features | |||||
|---|---|---|---|---|---|
| Random Forest Model | Single Date | 1-Year Difference | Variability | Risk | Total |
| Single date (RFdate) | 9 | 4 | 13 | ||
| 1-year diff. (RFtemp) | 9 | 4 | 13 | ||
| Combined (RFcomb) | 9 | 9 | 8 | 26 | |
| Risk (RFrisk) | 9 | 9 | 8 | 1 | 27 |
| Number of RF Models 2017 | Number of RF Models 2018 | ||
|---|---|---|---|
| Tile | Single Date | Single Date | 1-Year Difference |
| A (north) | 5 | 11 | 6 |
| B (south) | 3 | 13 | 6 |
| Combined—Single Date | Combined—Temporal | |||
|---|---|---|---|---|
| Tile | Mean Diff. | Max Diff. | Mean Diff. | Max Diff. |
| Tile A (north) | 5.9% | 14.9% | 4.6% | 8.8% |
| Tile B (south) | 5.0% | 8.7% | 5.9% | 10.7% |
| Single-Date Models | Temporal Models | |||
|---|---|---|---|---|
| Tile | Mean Diff. | Max Diff. | Mean Diff. | Max Diff. |
| Tile A (north) | 0.8% | 5.1% | 0.8% | 1.7% |
| Tile B (south) | 0.8% | 3.6% | 0.5% | 1.0% |
| Single-Date Models | Combined Models | |||
|---|---|---|---|---|
| Tile | Mean Diff. | Max Diff. | Mean Diff. | Max Diff. |
| Tile A (north) | 12.0% | 18.1% | 8.7% | 13.7% |
| Tile B (south) | 1.8% | 3.7% | 0.9% | 2.3% |
| Feature Labels | |||||
|---|---|---|---|---|---|
| Spatial | Variability Metrics | ||||
| Sentinel-2 Band | Resolution (m) | Single Date | 1-Year Diff. | Single Date | 1-Year Diff. |
| Band 2, Blue | 10 | BLU | TF_BLU | BLU_vm | TF_BLU_vm |
| Band 3, Green | 10 | GRN | TF_GRN | GRN_vm | TF_GRN_vm |
| Band 4, Red | 10 | RED | TF_RED | RED_vm | TF_RED_vm |
| Band 5, red-edge1 | 20 | RE1 | TF_RE1 | ||
| Band 6, red-edge2 | 20 | RE2 | TF_RE2 | ||
| Band 7, red-edge3 | 20 | RE3 | TF_RE3 | ||
| Band 8, NIR | 10 | NIR | TF_NIR | NIR_vm | TF_NIR_vm |
| Band 11, SWIR1 | 20 | SW1 | TF_SW1 | ||
| Band 12, SWIR2 | 20 | SW2 | TF_SW2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olsson, P.-O.; Zhao, P.; Müller, M.; Mansourian, A.; Ardö, J. Combining Sentinel-2 Data and Risk Maps to Detect Trees Predisposed to and Attacked by European Spruce Bark Beetle. Remote Sens. 2024, 16, 4166. https://doi.org/10.3390/rs16224166
Olsson P-O, Zhao P, Müller M, Mansourian A, Ardö J. Combining Sentinel-2 Data and Risk Maps to Detect Trees Predisposed to and Attacked by European Spruce Bark Beetle. Remote Sensing. 2024; 16(22):4166. https://doi.org/10.3390/rs16224166
Chicago/Turabian StyleOlsson, Per-Ola, Pengxiang Zhao, Mitro Müller, Ali Mansourian, and Jonas Ardö. 2024. "Combining Sentinel-2 Data and Risk Maps to Detect Trees Predisposed to and Attacked by European Spruce Bark Beetle" Remote Sensing 16, no. 22: 4166. https://doi.org/10.3390/rs16224166
APA StyleOlsson, P.-O., Zhao, P., Müller, M., Mansourian, A., & Ardö, J. (2024). Combining Sentinel-2 Data and Risk Maps to Detect Trees Predisposed to and Attacked by European Spruce Bark Beetle. Remote Sensing, 16(22), 4166. https://doi.org/10.3390/rs16224166