
Impacts of Climate Variations and Human Activities on the Net Primary Productivity of Different Grassland Types in the Three-River Headwaters Region



Abstract
:1. Introduction
2. Materials and Methods
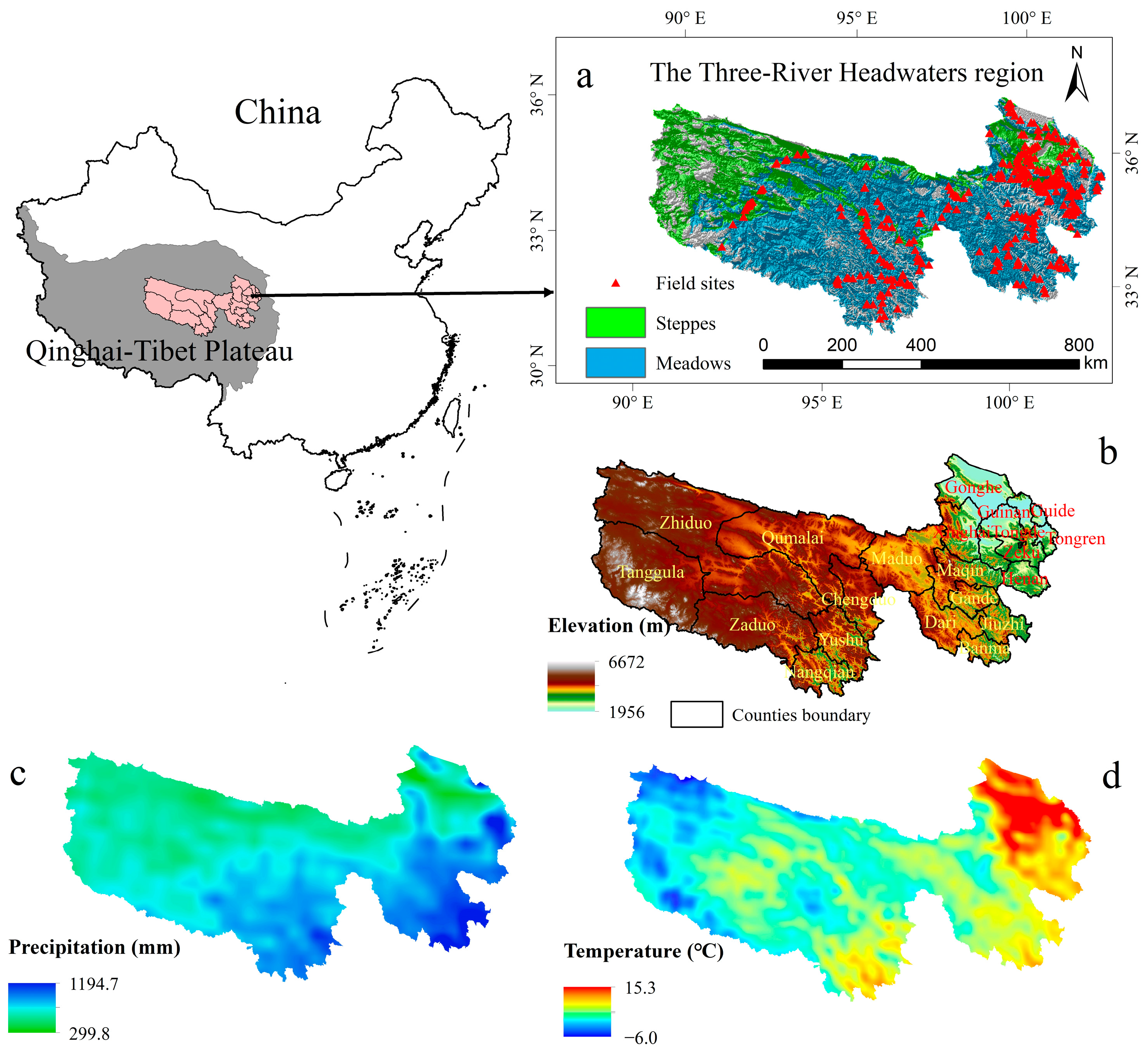
2.1. Study Area
2.2. Datasets and Preprocessing
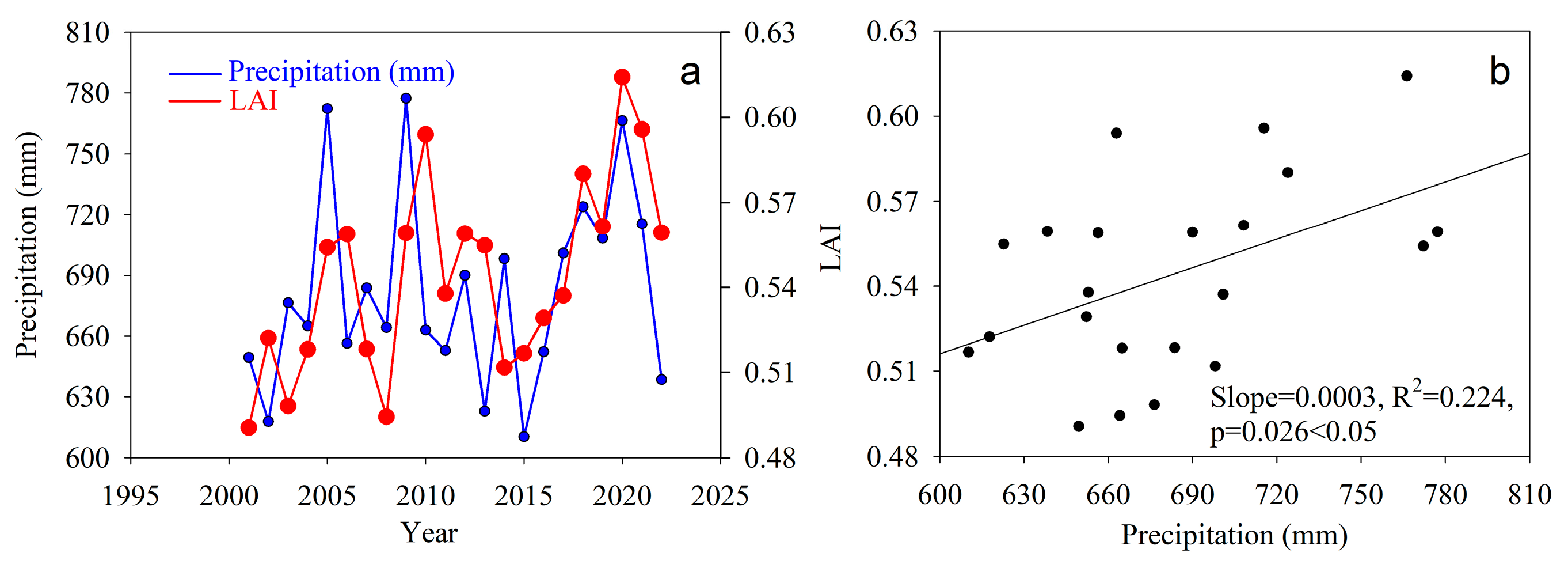
2.2.1. LAI Data
2.2.2. Grassland Types Data
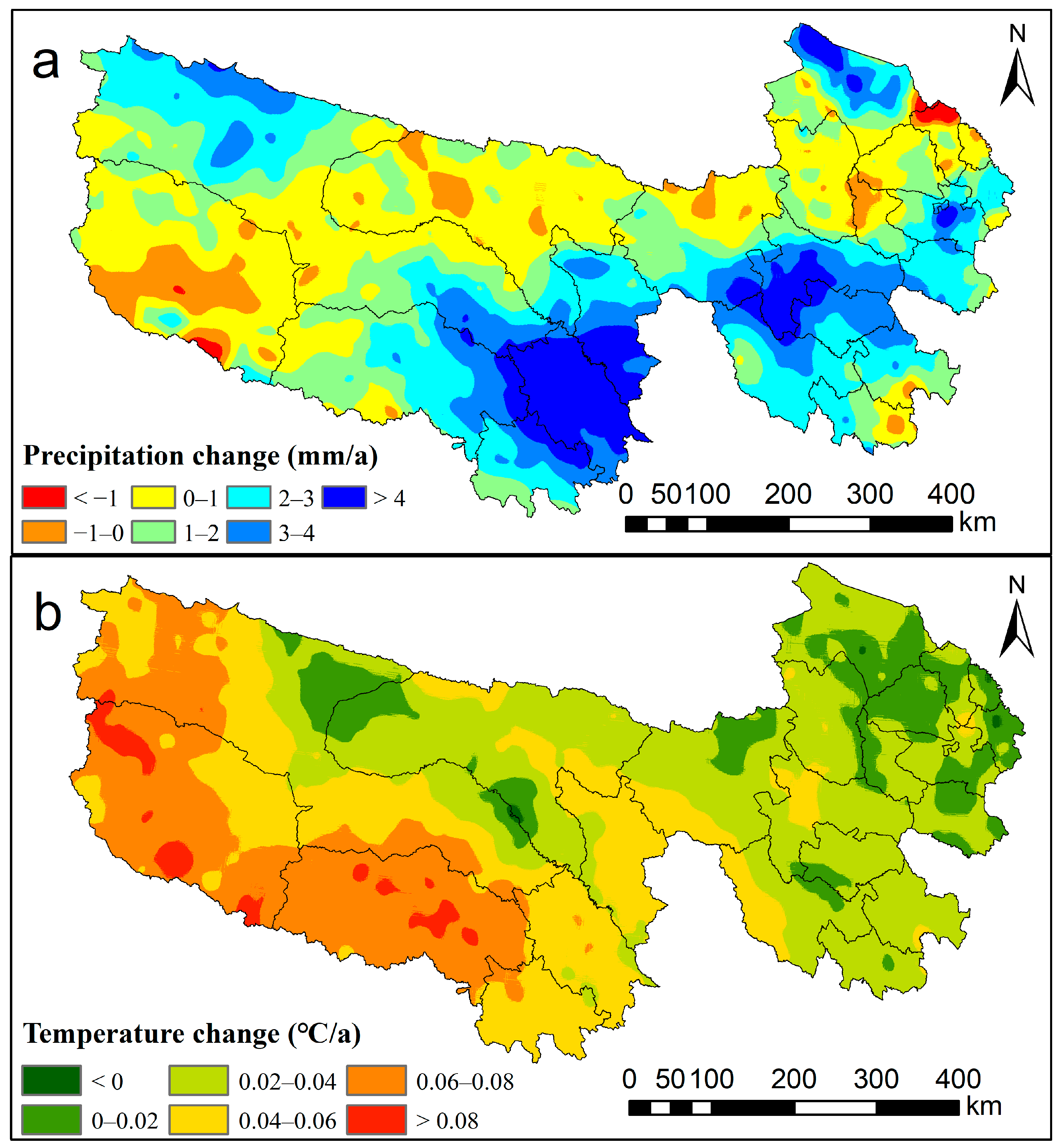
2.2.3. Climate Data
2.2.4. Field Survey Data
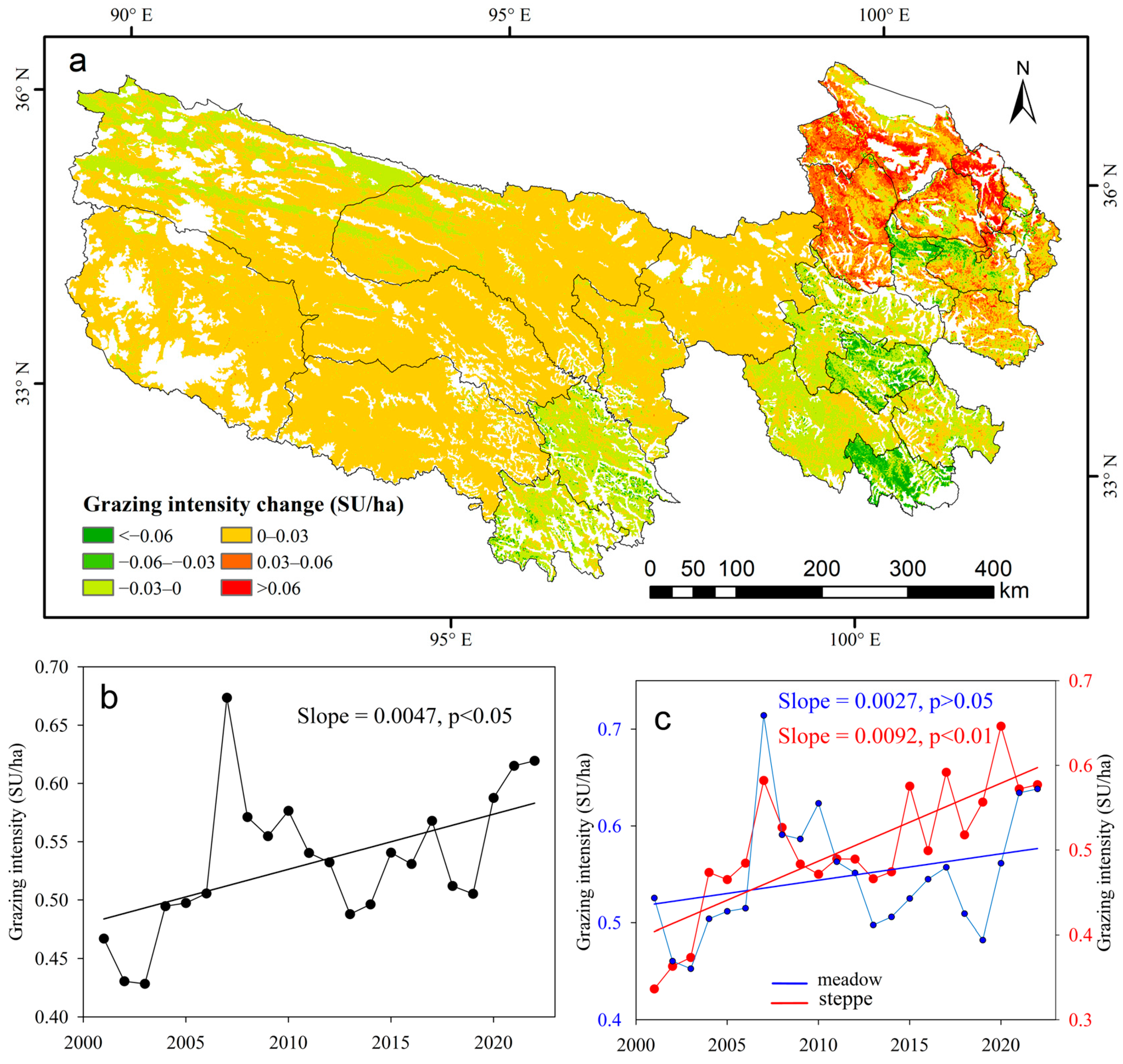
2.2.5. Grazing Intensity Data
2.3. Modeling Actual NPP, Climate NPP, and Human Activity NPP
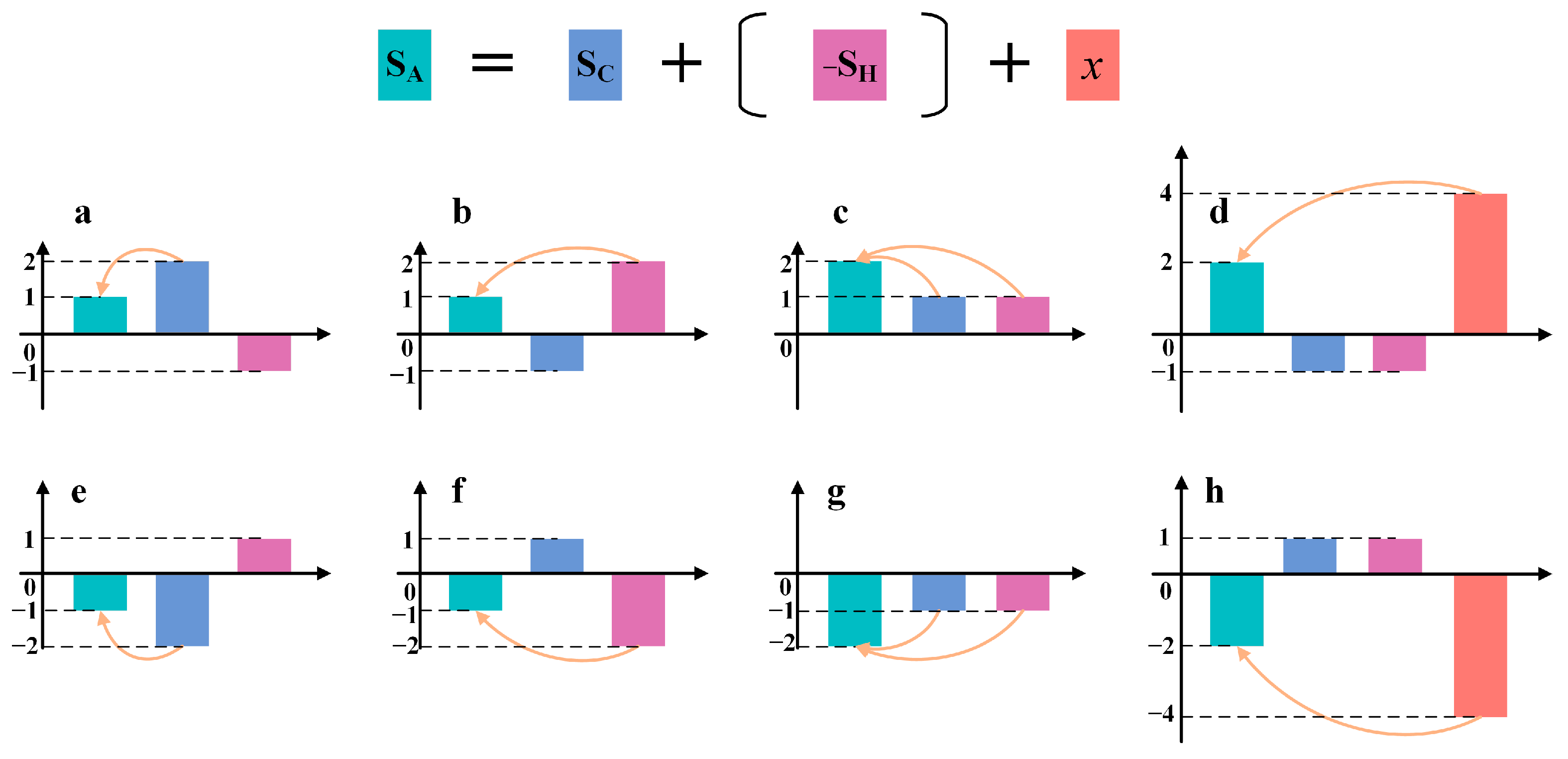
2.4. Attribution of Changes in NPP
2.4.1. NPP Dynamic Assessment
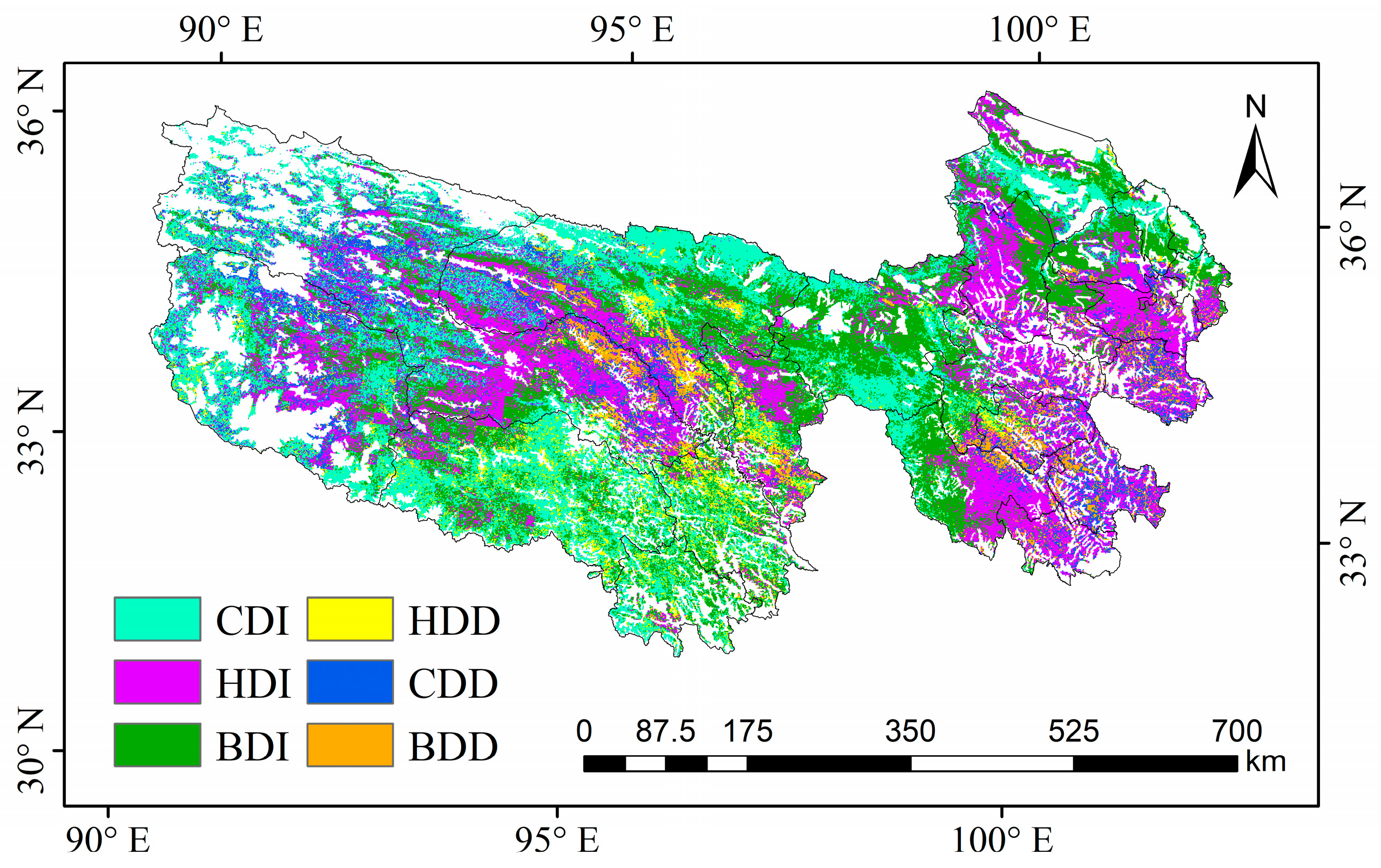
2.4.2. Establishing Scenarios
3. Results
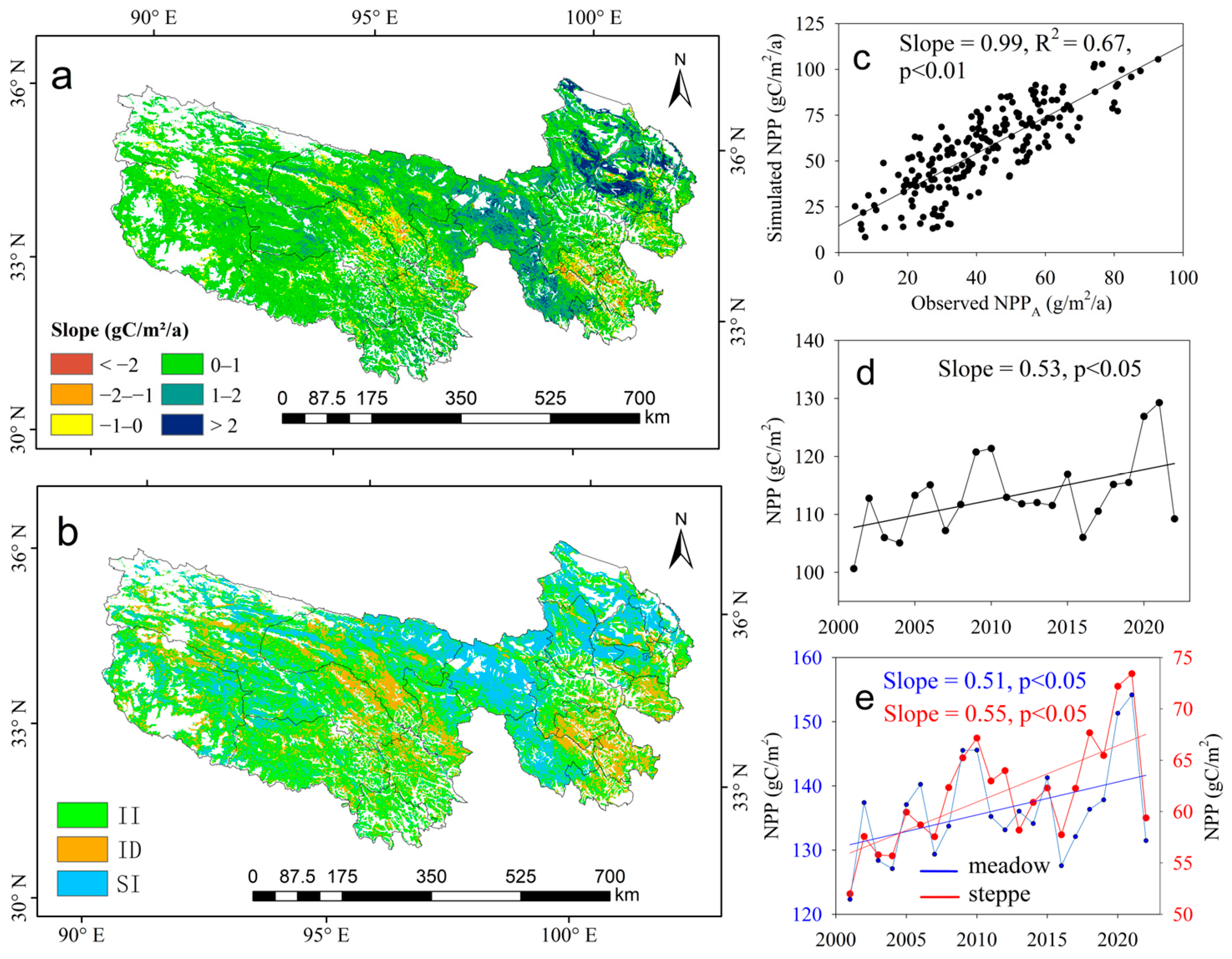
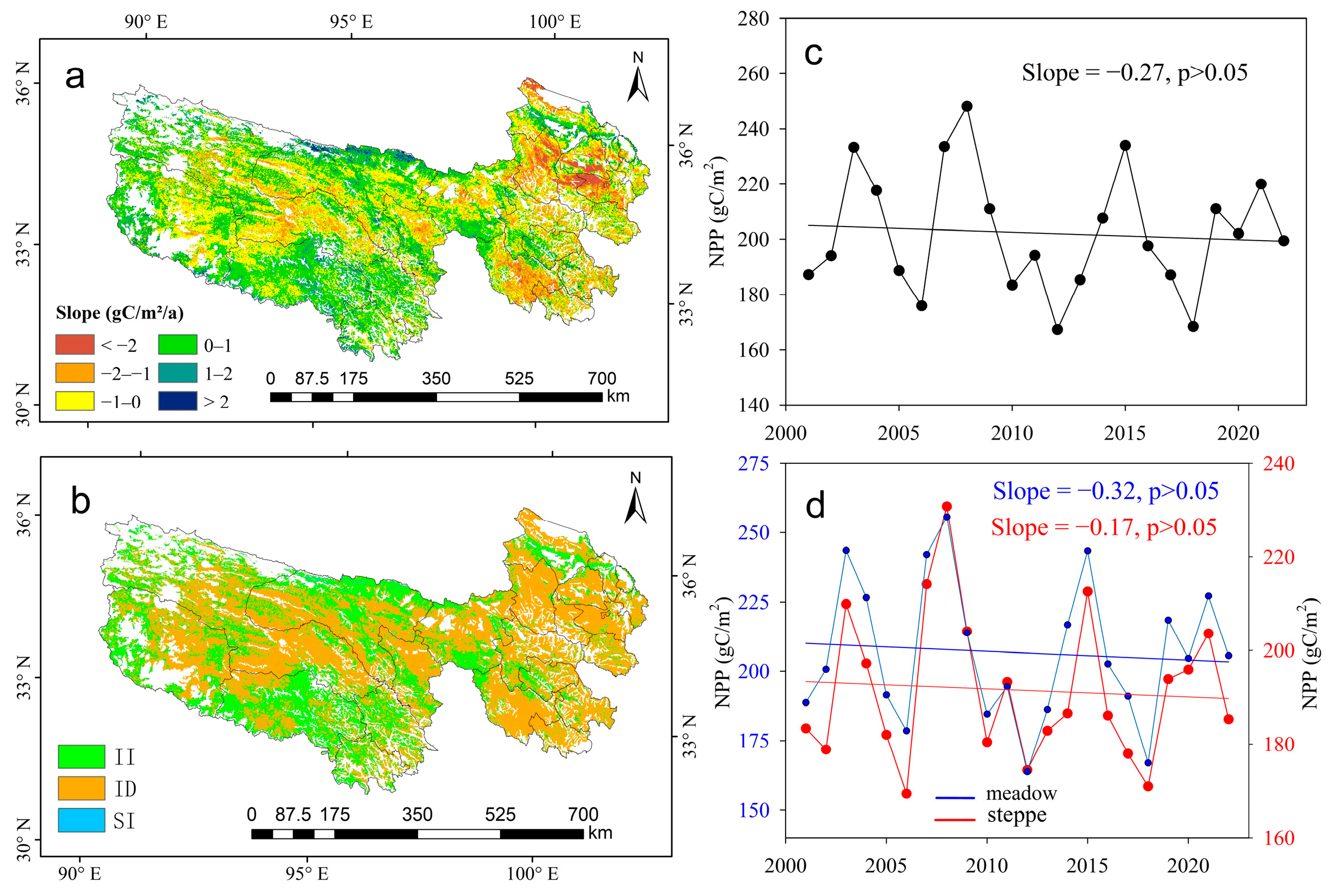
3.1. Trends of NPPA in Meadow and Steppe
3.2. Trends of NPPC in Meadow and Steppe
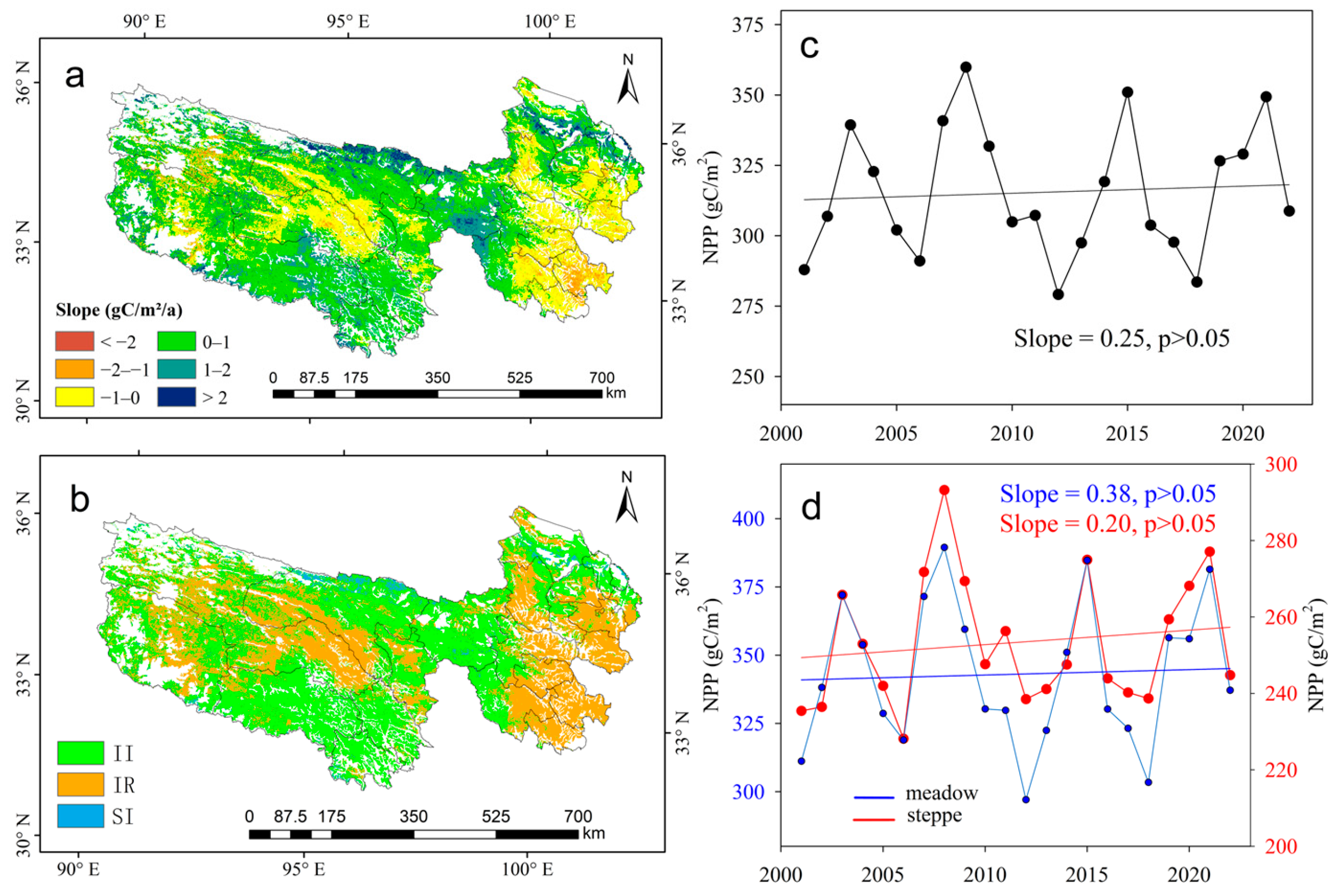
3.3. Trends of NPPH in Meadow and Steppe
3.4. Contribution of Climate and Human Activities on NPPA Dynamics
4. Discussion
4.1. Trends of NPP Are Increasing but Slowing Down
4.2. Climate and Human Activities on NPP Variations
4.3. Responses of Alpine Meadow and Steppe to Climate and Human Activity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominated Factor | CASA | DNN | Similarity |
|---|---|---|---|
| CDD | 262.42 | 252.68 | 96.29% |
| HDD | 166.23 | 163.38 | 98.29% |
| BDD | 130.39 | 129.77 | 99.52% |
| CDI | 821.91 | 794.72 | 96.69% |
| HDI | 786.41 | 790.89 | 99.43% |
| BDI | 780.48 | 816.38 | 95.60% |

References
- Wang, J.; Wang, K.; Zhang, M.; Zhang, C. Impacts of climate change and human activities on vegetation cover in hilly southern China. Ecol. Eng. 2015, 81, 451–461. [Google Scholar] [CrossRef]
- Rull, V.; González-Sampériz, P.; Pablo, J.; Mario, C.; Santiago, J.M. Vegetation changes in the southern Pyrenean flank during the last millennium in relation to climate and human activities: The Montcortès lacustrine record. J. Paleolimnol. 2010, 46, 387–404. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Yang, Y.; Zhou, W.; Gang, C.; Zhang, Y.; Li, J.; An, R.; Wang, K.; Odeh, I.; et al. Quantitative assess the driving forces on the grassland degradation in the Qinghai–Tibet Plateau, in China. Ecol. Inform. 2016, 33, 32–44. [Google Scholar] [CrossRef]
- Zheng, K.; Wei, J.Z.; Pei, J.Y.; Cheng, H.; Zhang, X.L.; Huang, F.Q.; Li, F.M.; Ye, J.S. Impacts of climate change and human activities on grassland vegetation variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef]
- Yuan, X.; Li, L.; Chen, X. Increased grass NDVI under contrasting trends of precipitation change over North China during 1982–2011. Remote Sens. Lett. 2015, 6, 69–77. [Google Scholar] [CrossRef]
- Suzuki, R.; Tanaka, S.; Yasunari, T. Relationships between meridional profiles of satellite-derived vegetation index (NDVI) and climate over Siberia. Int. J. Climatol. 2015, 20, 955–967. [Google Scholar] [CrossRef]
- Huang, S.; Zheng, X.; Ma, L.; Wang, H.; Huang, Q.; Leng, G.; Meng, E.; Guo, Y. Quantitative contribution of climate change and human activities to vegetation cover variations based on GA-SVM model. J. Hydrol. 2020, 584, 124687. [Google Scholar] [CrossRef]
- Li, S.; Yang, S.; Liu, X.; Liu, Y.; Shi, M. NDVI-Based Analysis on the Influence of Climate Change and Human Activities on Vegetation Restoration in the Shaanxi-Gansu-Ningxia Region, Central China. Remote Sens. 2015, 7, 11163–11182. [Google Scholar] [CrossRef]
- Wang, Z.; Song, K.; Hu, L. China’s Largest Scale Ecological Migration in the Three-River Headwater Region. AMBIO 2010, 39, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Li, Y.; Wan, Y.; Jiangcun, W.; Qin, X.; Wang, B. Significant Achievements in Protection and Restoration of Alpine Grassland Ecosystem in Northern Tibet, China. Restor. Ecol. 2010, 17, 320–323. [Google Scholar] [CrossRef]
- Caviezel, C.; Hunziker, M.; Schaffner, M.; Kuhn, N. Soil–vegetation interaction on slopes with bush encroachment in the central Alps—Adapting slope stability measurements to shifting process domains. Earth Surf. Process. Landf. 2014, 39, 509–521. [Google Scholar] [CrossRef]
- Wang, Z.; Chang, J.; Peng, S.; Piao, S.; Ciais, P.; Betts, R. Changes in productivity and carbon storage of grasslands in China under future global warming scenarios of 1.5 °C and 2 °C. J. Plant Ecol. 2019, 12, 804–814. [Google Scholar] [CrossRef]
- Li, X.L.; Gao, J.; Brierley, G.; Qiao, Y.M.; Zhang, J.; Yang, Y.W. Rangeland degradation on the Qinghai-Tibet Plateau: Implications for rehabilitation. Land Degrad. Dev. 2013, 24, 72–80. [Google Scholar] [CrossRef]
- Fan, J.W.; Shao, Q.Q.; Liu, J.Y.; Wang, J.B.; Harris, W.; Chen, Z.Q.; Zhong, H.P.; Xu, X.L.; Liu, R.G. Assessment of effects of climate change and grazing activity on grassland yield in the Three Rivers Headwaters Region of Qinghai–Tibet Plateau, China. Environ. Monit. Assess. 2010, 170, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xiang, R.; Tian, L.; Zhang, T. Effect of grazing exclusion on ecosystem respiration among three different alpine grasslands on the central Tibetan Plateau. Ecol. Eng. 2016, 94, 599–607. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, X.; Niu, B.; He, Y.; Wu, J. Changes in plant species richness distribution in Tibetan alpine grasslands under different precipitation scenarios. Glob. Ecol. Conserv. 2020, 21, e00848. [Google Scholar]
- Zou, S.; Lv, F. Two Special Vegetation Types on the Qinghai Tibet Plateau: Alpine Grassland and Alpine Meadow. Geogr. Teach. 2016, 2, 4–7. [Google Scholar]
- Schloss, A.L.; Kicklighter, D.W.; Kaduk, J.; Wittenberg, U. Comparing global models of terrestrial net primary productivity (NPP): Comparison of NPP to climate and the Normalized Difference Vegetation Index (NDVI). Glob. Change Biol. 1999, 5, 25–34. [Google Scholar] [CrossRef]
- Zhao, C.; Ma, Y.; Shang, W.; Chen, X.; Rong, J.; Tian, Y.; Yuan, Q. RESTREND-based assessment of factors affecting vegetation dynamics on the Mongolian Plateau. Ecol. Model. 2021, 440, 109415. [Google Scholar] [CrossRef]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A.; Klooster, S.A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Lieth, H. Primary production: Terrestrial ecosystems. Hum. Ecol. 1973, 1, 303–332. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Wang, Z.; Chen, Y.; Gang, C.; An, R.; Li, J. Vegetation dynamics and its driving forces from climate change and human activities in the Three-River Source Region, China from 1982 to 2012. Sci. Total Environ. 2016, 563–564, 210–220. [Google Scholar] [CrossRef]
- Gang, C.; Zhou, W.; Chen, Y.; Wang, Z.; Sun, Z.; Li, J.; Qi, J.; Odeh, I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Li, J.; Peng, S.; Zhi, L. Detecting and attributing vegetation changes on China’s Loess Plateau. Agric. For. Meteorol. 2017, 247, 260–270. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Li, J.; Hua, T. Roles of climate changes and human interventions in land degradation: A case study by net primary productivity analysis in China’s Shiyanghe Basin. Environ. Earth Sci. 2011, 64, 2183–2193. [Google Scholar] [CrossRef]
- Ruimy, A.; Kergoat, A.; Bondeau, A. Comparing global models of terrestrial net primary productivity (NPP): Analysis of differences in light absorption and light-use efficiency. Glob. Change Biol. 1999, 5 (Suppl. 1), 56–64. [Google Scholar] [CrossRef]
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens. 1986, 7, 1417–1434. [Google Scholar] [CrossRef]
- Zhang, X. Vegetation of China and Its Geographic Pattern: Illustration of the Vegetation Map of the People’s Republic of China (1:1,000,000); Geological Publishing House: Beijing, China, 2007. [Google Scholar]
- Sun, Y.; Chang, J.; Fang, J. Above-and belowground net-primary productivity: A field-based global database of grasslands. Ecol. Ecol. Soc. Am. 2023, 104, e3904. [Google Scholar] [CrossRef]
- Xia, J.; Ma, M.; Liang, T.; Wu, C.; Yang, Y.; Zhang, L.; Zhang, Y.; Yuan, W. Estimates of grassland biomass and turnover time on the Tibetan Plateau. Environ. Res. Lett. 2018, 13, 014020. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ma, W.; Guo, D.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar] [CrossRef]
- He, L.; Li, Z.-L.; Wang, X.; Xie, Y.; Ye, J.-S. Lagged precipitation effect on plant productivity is influenced collectively by climate and edaphic factors in drylands. Sci. Total Environ. 2021, 755, 142506. [Google Scholar] [CrossRef]
- Zhu, W.; Pan, Y.; Zhang, J. Estimation of net primary productivity of chinese terrestrial vegetation based on remote sensing. J. Plant Ecol. 2007, 31, 412–424. [Google Scholar]
- Zhu, W.; Pan, Y.; Hao, H.E. Simulation of maximum light use efficiency for some typical vegetation types in China. Chin. Sci. Bull. 2006, 51, 457–463. [Google Scholar] [CrossRef]
- Han, Z.; Song, W.; Deng, X.; Xu, X. Grassland ecosystem responses to climate change and human activities within the Three-River Headwaters region of China. Sci. Rep. 2018, 8, 9079. [Google Scholar] [CrossRef] [PubMed]
- Maeda, E.E. Downscaling MODIS LST in the East African mountains using elevation gradient and land-cover information. Int. J. Remote Sens. 2014, 35, 3094–3108. [Google Scholar] [CrossRef]
- Quinonez-Pinon, R.; Mendoza-Duran, A.; Valeo, C. Design of an environmental monitoring program using NDVI and cumulative effects assessment. Int. J. Remote Sens. 2007, 28, 1643–1664. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, Y.; Zhu, J.; Liu, Y.; Zu, J.; Zhang, J. The Influences of Climate Change and Human Activities on Vegetation Dynamics in the Qinghai-Tibet Plateau. Remote Sens. 2016, 8, 876. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, W.; Fu, C.; Wu, L. The influence of vegetation cover on summer precipitation in China: A statistical analysis of NDVI and climate data. Adv. Atmos. Sci. 2003, 20, 1002–1006. [Google Scholar]
- Ge, W.; Lin, H. The Vegetation NDVI Variation Trend in Qinghai-Tibet Plateau and Its Response to Climate Change. In Proceedings of the Remote Sensing, Environment and Transportation Engineering (RSETE), 2012 2nd International Conference on IEEE, Nanjing, China, 1–3 June 2012. [Google Scholar]
- Zhang, Z.; Han, W.; Wang, J.; Cai, Y.; Gang, L.I.; Cai, Z.; Zhang, C.; Sheng, H.; Zhao, C. Characteristics of Climate Change and its Response in the Qinghai-Tibet Plateau during 1971–2010. J. Landsc. Res. 2016, 8, 41–45,52. [Google Scholar]
- Cao, J.; Wu, E.; Wu, S.; Fan, R.; Xu, L.; Ning, K.; Li, Y.; Lu, R.; Xu, X.; Zhang, J.; et al. Spatiotemporal Dynamics of Ecological Condition in Qinghai-Tibet Plateau Based on Remotely Sensed Ecological Index. Remote Sens. 2022, 14, 4234. [Google Scholar] [CrossRef]
- Zheng, Y.; Han, J.; Huang, Y.; Fassnacht, S.R.; Shuai, X.; Lv, E.; Min, C. Vegetation response to climate conditions based on NDVI simulations using stepwise cluster analysis for the Three-River Headwaters region of China. Ecol. Indic. 2017, 92, 18–29. [Google Scholar] [CrossRef]
- Li, M.; Zhang, X.; He, Y.; Niu, B.; Wu, J. Assessment of the vulnerability of alpine grasslands on the Qinghai-Tibetan Plateau. PeerJ 2020, 8, e8513. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Xue, X.; Wang, T.; Kang, W.; Liao, J.; Liu, S. Spatial and temporal differences in alpine meadow, alpine steppe and all vegetation of the Qinghai-Tibetan Plateau and their responses to climate change. Remote Sens. 2021, 13, 669. [Google Scholar] [CrossRef]
- State Council of the People’s Republic of China. Ecological Protection and Construction of Qinghai Three RiversNature Reserve Master Plan; State Council of the People’s Republic of China: Beijing, China, 2005; p. 54. (In Chinese) [Google Scholar]
- Zeng, C.; Zhang, F.; Wang, Q.; Chen, Y.; Joswiak, D. Impact of alpine meadow degradation on soil hydraulic properties over the Qinghai-Tibetan Plateau. J. Hydrol. 2013, 478, 148–156. [Google Scholar] [CrossRef]
- Wu, H.; Lü, L.; Zhang, Y.; Xu, C.; Yu, Q. Sediment addition and legume cultivation result in sustainable, long-term increases in ecosystem functions of sandy grasslands. Land Degrad. Dev. 2019, 30, 1667–1676. [Google Scholar] [CrossRef]
- Wen, L.; Jinlan, W.; Xiaojiao, Z.; Shangli, S.; Wenxia, C. Effect of degradation and rebuilding of artificial grasslands on soil respiration and carbon and nitrogen pools on an alpine meadow of the Qinghai-Tibetan Plateau. Ecol. Eng. 2018, 111, 134–142. [Google Scholar] [CrossRef]
- Mackenzie, C.A.; Salerno, J.; Hartter, J.; Chapman, C.A.; Drake, M. Changing perceptions of protected area benefits and problems around Kibale National Park, Uganda. J. Environ. Manag. 2017, 200, 217–228. [Google Scholar] [CrossRef]
- Kim, Y.; Wang, G. Soil moisture-vegetation-precipitation feedback over North America: Its sensitivity to soil moisture climatology. J. Geophys. Res. Atmos. 2012, 117, 1–18. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, Y.; Yang, X.; Zhang, H.; Guo, Z.; Hu, G. Indirect effects of warming via phenology on reproductive success of alpine plants. J. Ecol. 2024, 113, 180–193. [Google Scholar] [CrossRef]
- Zhang, X. Plant Community and Soil Change Characteristics of Artificial Grassland in Black Soil Beach Based on Different Restoration Measures; Qinghai University: Xining, China, 2021. [Google Scholar]

| Dominated Factor | Area (102 km2) | NPP Change (gC/m2/a) | Total NPP (GgC) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Steppes | Meadows | Total | Steppes | Meadows | Total | Steppes | Meadows | |
| CDD 1 | 262.42 | 89.72 | 172.70 | −0.27 | −0.19 | −0.31 | −7.10 | −1.68 | −5.41 |
| HDD 2 | 166.23 | 23.99 | 142.24 | −0.37 | −0.22 | −0.39 | −6.08 | −0.52 | −5.56 |
| BDD 3 | 130.39 | 8.72 | 121.67 | −1.05 | −0.62 | −1.08 | −13.64 | −0.54 | −13.09 |
| CDI 4 | 821.91 | 330.29 | 491.62 | 0.51 | 0.49 | 0.53 | 41.98 | 16.03 | 25.95 |
| HDI 5 | 786.41 | 182.90 | 603.51 | 0.67 | 0.59 | 0.70 | 52.69 | 10.72 | 41.97 |
| BDI 6 | 780.48 | 194.60 | 585.88 | 1.16 | 1.40 | 1.08 | 90.59 | 27.34 | 63.26 |
| Dominated Factor | Area (102 km2) | Dominated Factor | Area (102 km2) | ||||
|---|---|---|---|---|---|---|---|
| Total | Steppes | Meadows | Total | Steppes | Meadows | ||
| CDI1-1 | 19.67 | 9.13 | 10.54 | HDI4-1 | 59.91 | 16.63 | 43.29 |
| CDD1-2 | 2.60 | 1.24 | 1.36 | HDD4-1 | 12.88 | 1.99 | 10.89 |
| CDI2-1 | 17.50 | 5.50 | 11.99 | HDI4-2 | 20.20 | 1.64 | 18.56 |
| CDD2-2 | 7.90 | 2.62 | 5.27 | HDD4-2 | 3.58 | 0.22 | 3.36 |
| CDI3-2 | 15.59 | 8.84 | 6.74 | BDI1-1 | 47.71 | 16.70 | 31.01 |
| CDD3-2 | 4.34 | 2.70 | 1.64 | BDD1-2 | 10.54 | 0.84 | 9.69 |
| CDI3-1 | 28.61 | 9.22 | 19.39 | BDI2-1 | 30.40 | 3.66 | 26.74 |
| CDD3-1 | 11.03 | 2.28 | 8.75 | BDD2-2 | 2.34 | 0.03 | 2.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, K.; Liu, X.; Zou, X.; Wang, Z. Impacts of Climate Variations and Human Activities on the Net Primary Productivity of Different Grassland Types in the Three-River Headwaters Region. Remote Sens. 2025, 17, 471. https://doi.org/10.3390/rs17030471
Zheng K, Liu X, Zou X, Wang Z. Impacts of Climate Variations and Human Activities on the Net Primary Productivity of Different Grassland Types in the Three-River Headwaters Region. Remote Sensing. 2025; 17(3):471. https://doi.org/10.3390/rs17030471
Chicago/Turabian StyleZheng, Kai, Xiang Liu, Xiaoyu Zou, and Zhaoqi Wang. 2025. "Impacts of Climate Variations and Human Activities on the Net Primary Productivity of Different Grassland Types in the Three-River Headwaters Region" Remote Sensing 17, no. 3: 471. https://doi.org/10.3390/rs17030471
APA StyleZheng, K., Liu, X., Zou, X., & Wang, Z. (2025). Impacts of Climate Variations and Human Activities on the Net Primary Productivity of Different Grassland Types in the Three-River Headwaters Region. Remote Sensing, 17(3), 471. https://doi.org/10.3390/rs17030471