Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data


Abstract
1. Introduction
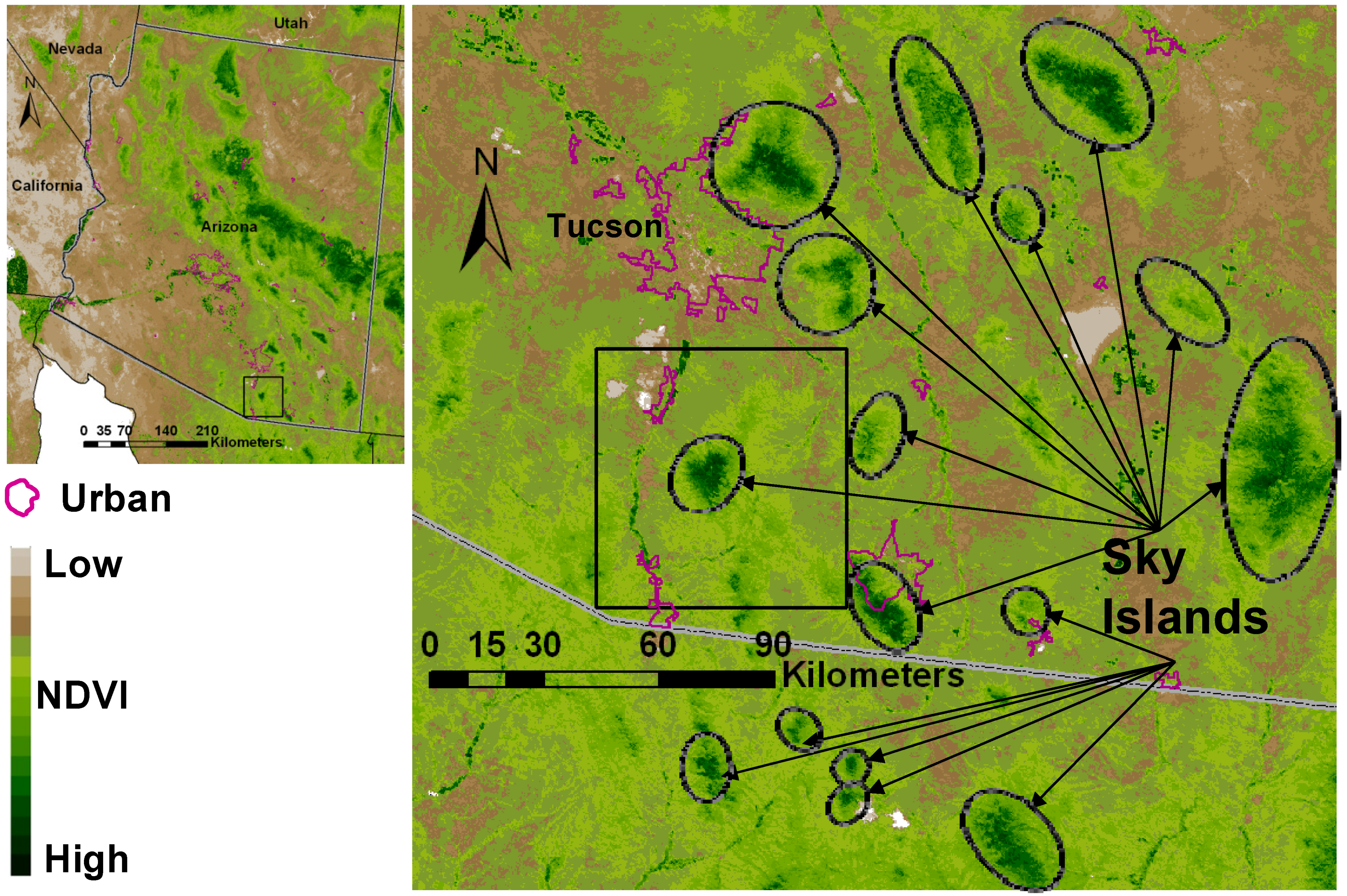
1.1. Nature of the Study Area
1.2. Study Objectives
2. Data and Methods
2.1. Study Area
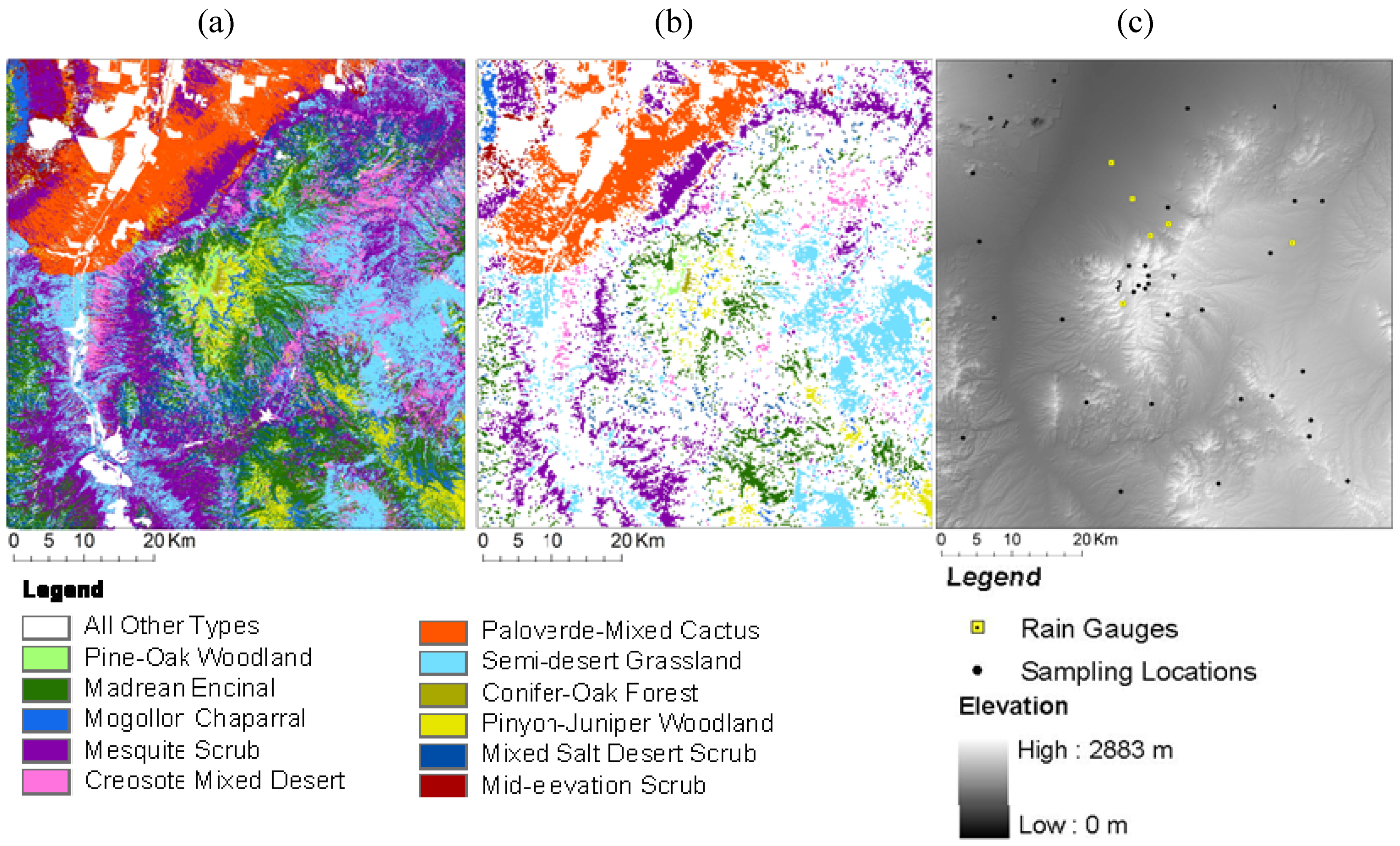
2.2. Research Design: Sampling Land Cover, NDVI and Climate Time Series Data

2.3. SWReGAP Landcover and DEM

2.4. MODIS NDVI Time Series
2.5. Precipitation and Temperature

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecoregion and natural landcover type | Elevation (m) | Tmin (°C) | Tmax (°C) | Annual Precip. (mm) | Precip. (mm) [djfm] | Precip. (mm) [amj] | Precip. (mm) [jas] | Precip. (mm) [on] | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sonoran Paloverde-Mixed Cacti Desert Scrub | 966 | 1.2 | 37.7 | 318.1 | 73.2 | 26.9 | 206.8 | 24.6 | |||
| Apacherian-Chihuahuan Mesquite Upland Scrub | 1,144 | −0.1 | 36.1 | 359.1 | 82.1 | 28.3 | 236.4 | 27.1 | |||
| Sonoran Mid-Elevation Desert Scrub | 1,151 | 1.8 | 36.3 | 361.6 | 80.7 | 26.9 | 241.7 | 28.6 | |||
| Chihuahuan Creosotebush, Mixed Desert and Thorn Scrub | 1,333 | −0.4 | 35.0 | 394.6 | 91.4 | 30.5 | 258.3 | 29.9 | |||
| Apacherian-Chihuahuan Piedmont Semi-Desert Grassland and Steppe | 1,358 | −1.3 | 34.8 | 393.7 | 90.5 | 30.4 | 259.9 | 28.2 | |||
| Chihuahuan Mixed Salt Desert Scrub | 1,377 | −1.1 | 34.5 | 413.7 | 94.2 | 33.0 | 271.8 | 28.9 | |||
| Madrean Encinal | 1,488 | −1.0 | 32.7 | 490.3 | 117.1 | 36.1 | 318.4 | 40.5 | |||
| Madrean Pinyon-Juniper Woodland | 1,689 | −2.4 | 33.0 | 454.2 | 105.2 | 35.9 | 297.2 | 34.8 | |||
| Mogollon Chaparral | 1,749 | −3.3 | 33.1 | 369.6 | 82.8 | 33.3 | 242.5 | 24.2 | |||
| Madrean Pine-Oak Forest and Woodland | 2,204 | −1.5 | 31.8 | 645.2 | 170.4 | 42.5 | 402.6 | 69.4 | |||
| Madrean Upper Montane Conifer-Oak Forest and Woodland | 2,627 | −1.6 | 31.6 | 702.6 | 185.7 | 45.2 | 437.6 | 79.8 | |||
2.6. Derivation of Phenological Metrics

2.7. Data Analysis
3. Results and Discussion
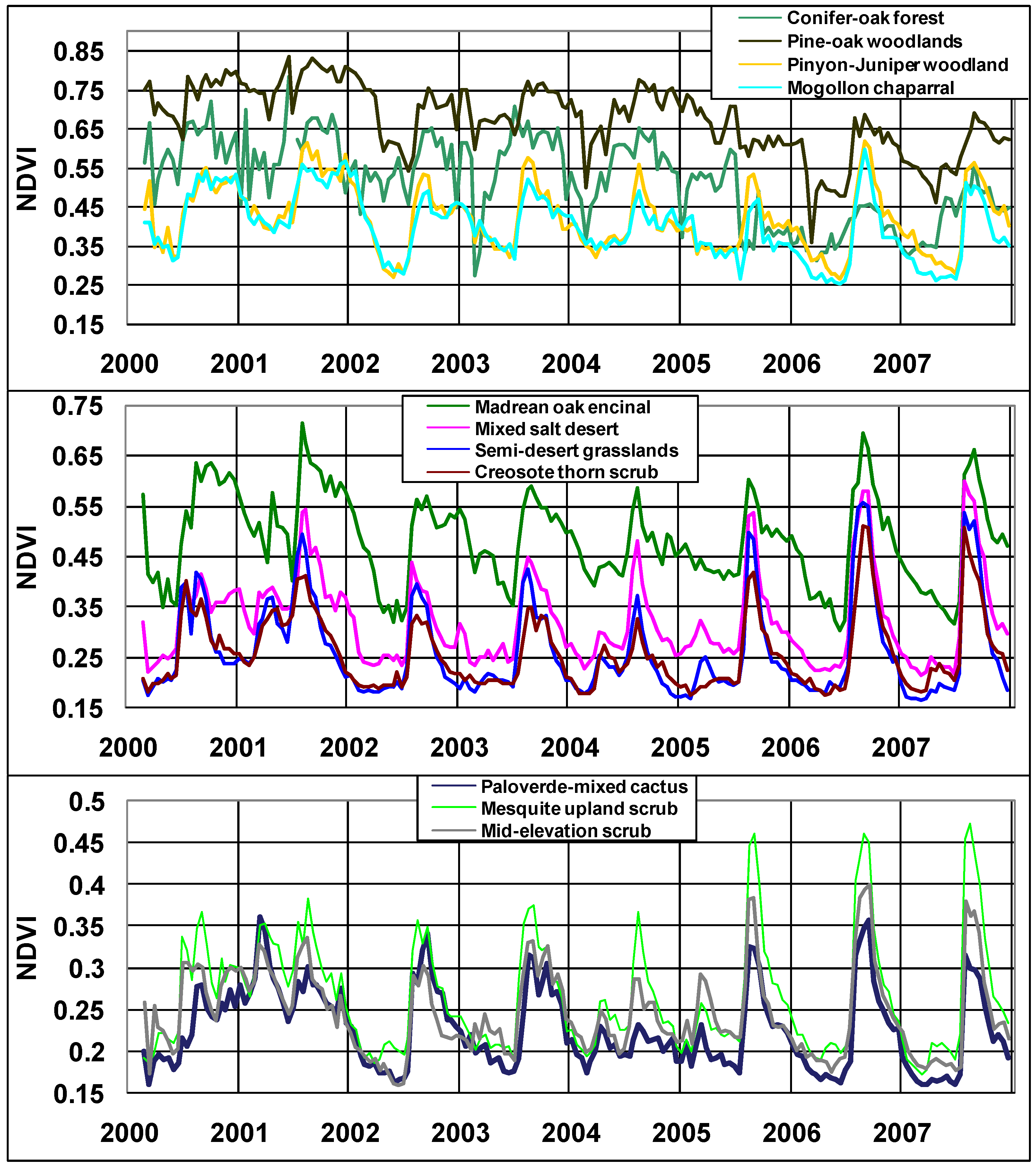
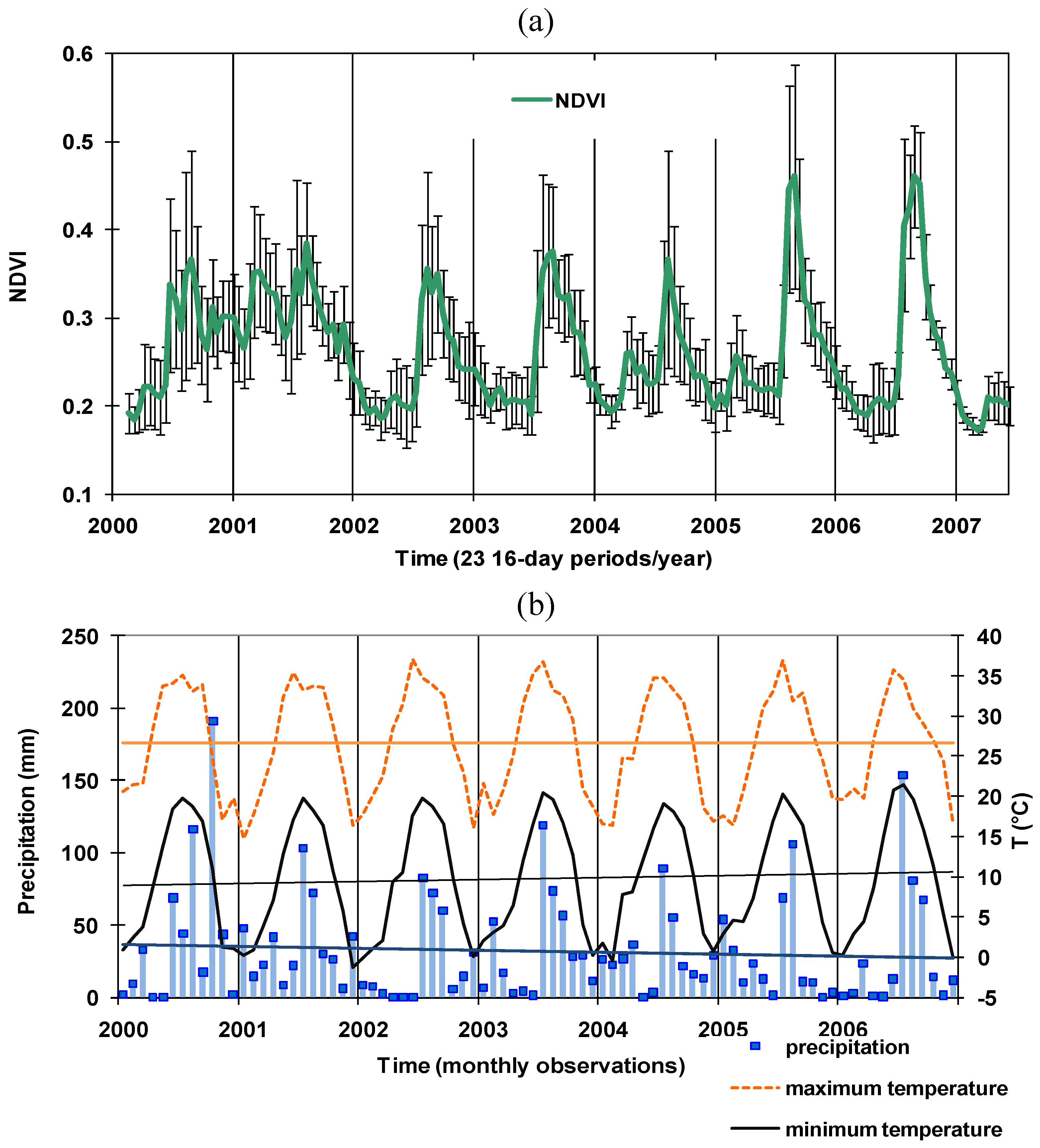
3.1. NDVI and Climate Time Series Patterns among Vegetation Types



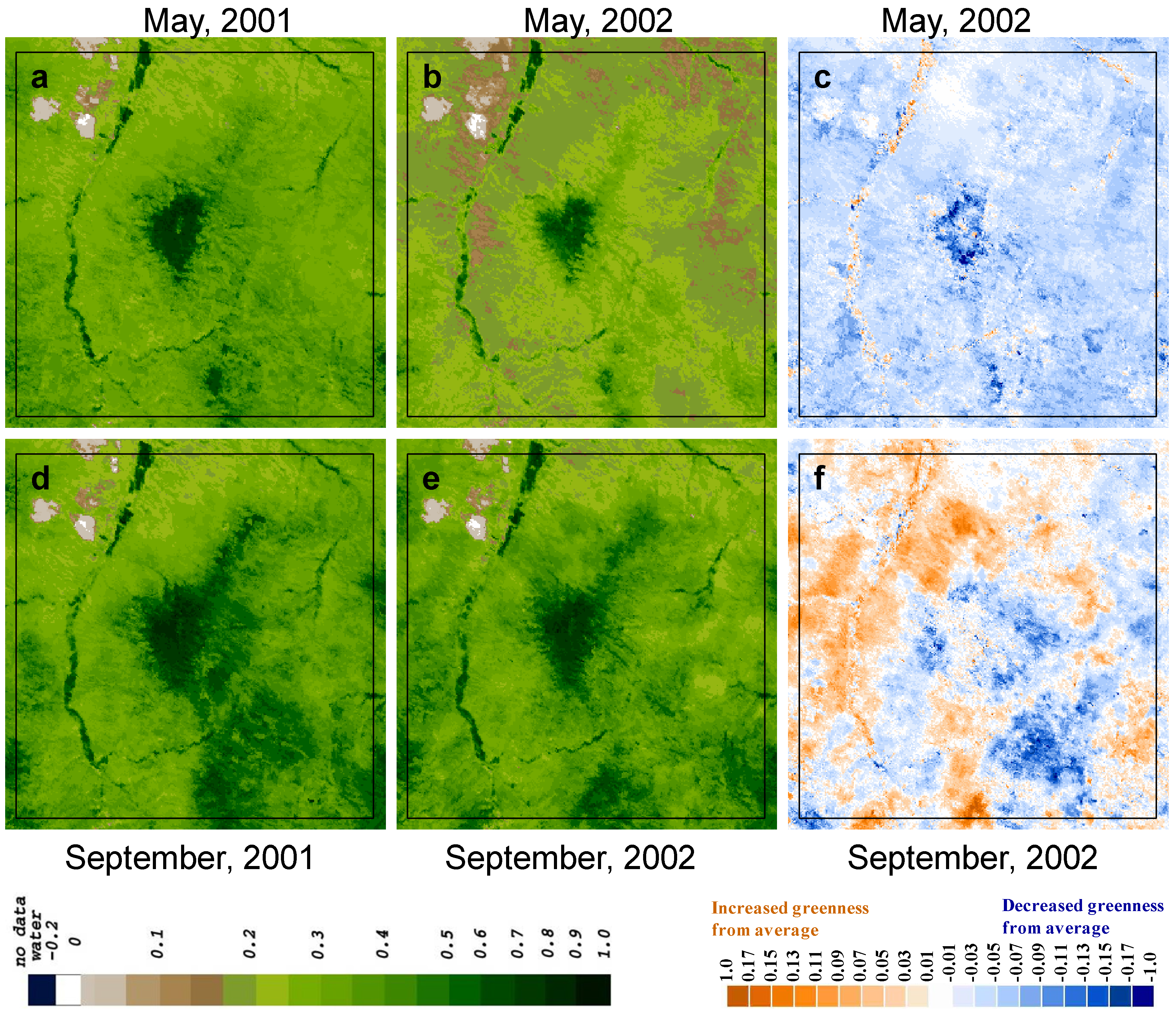
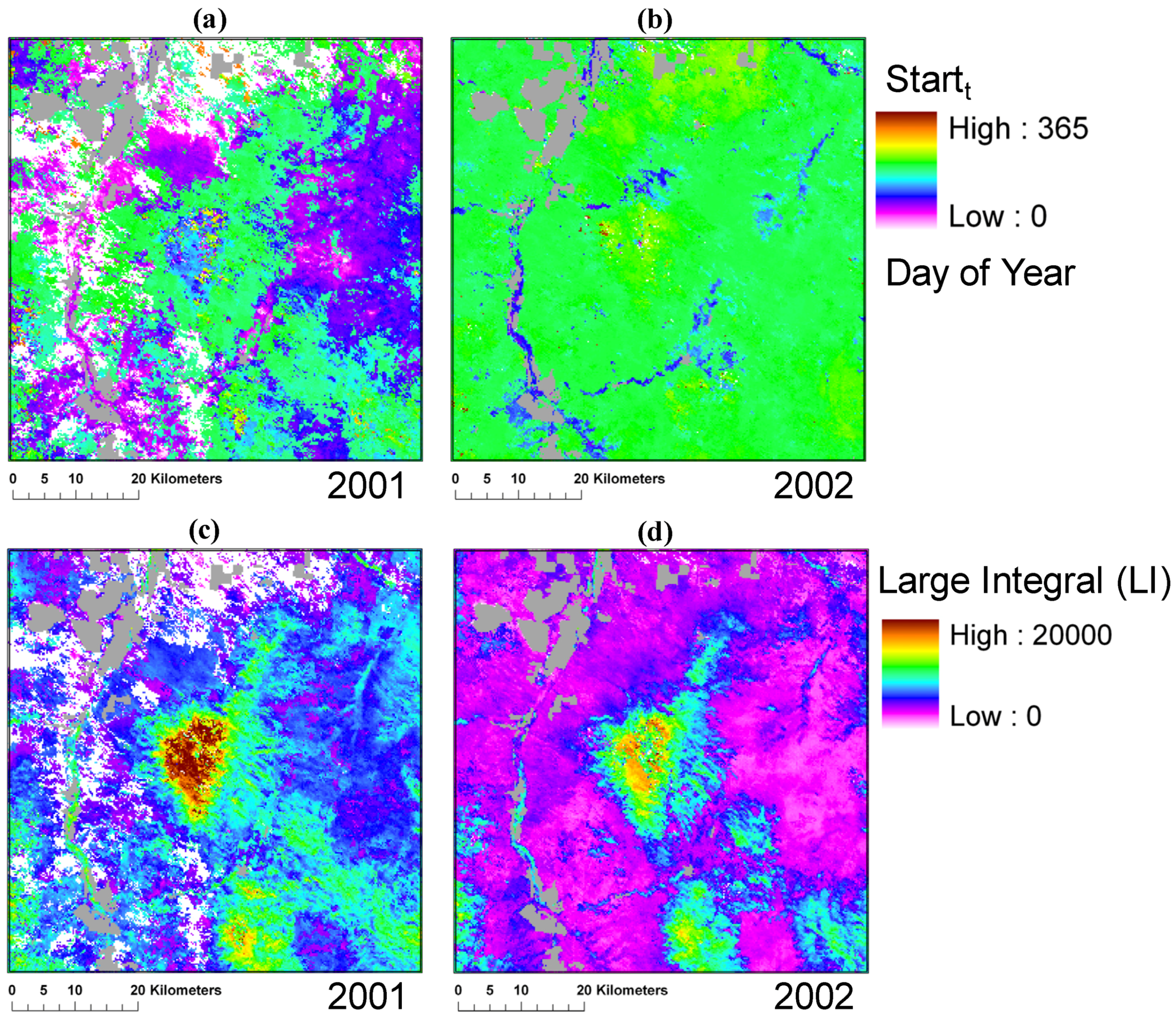
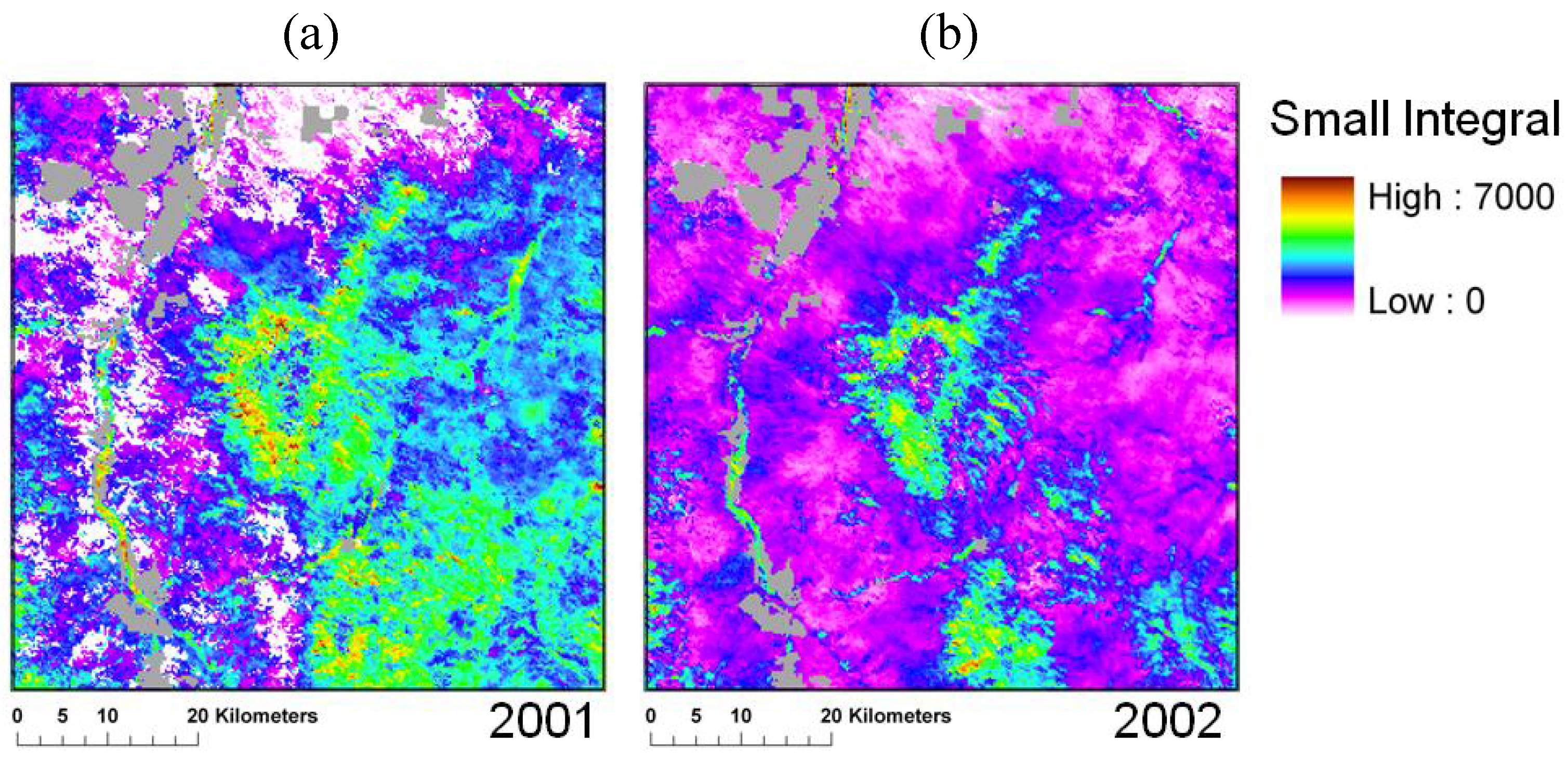
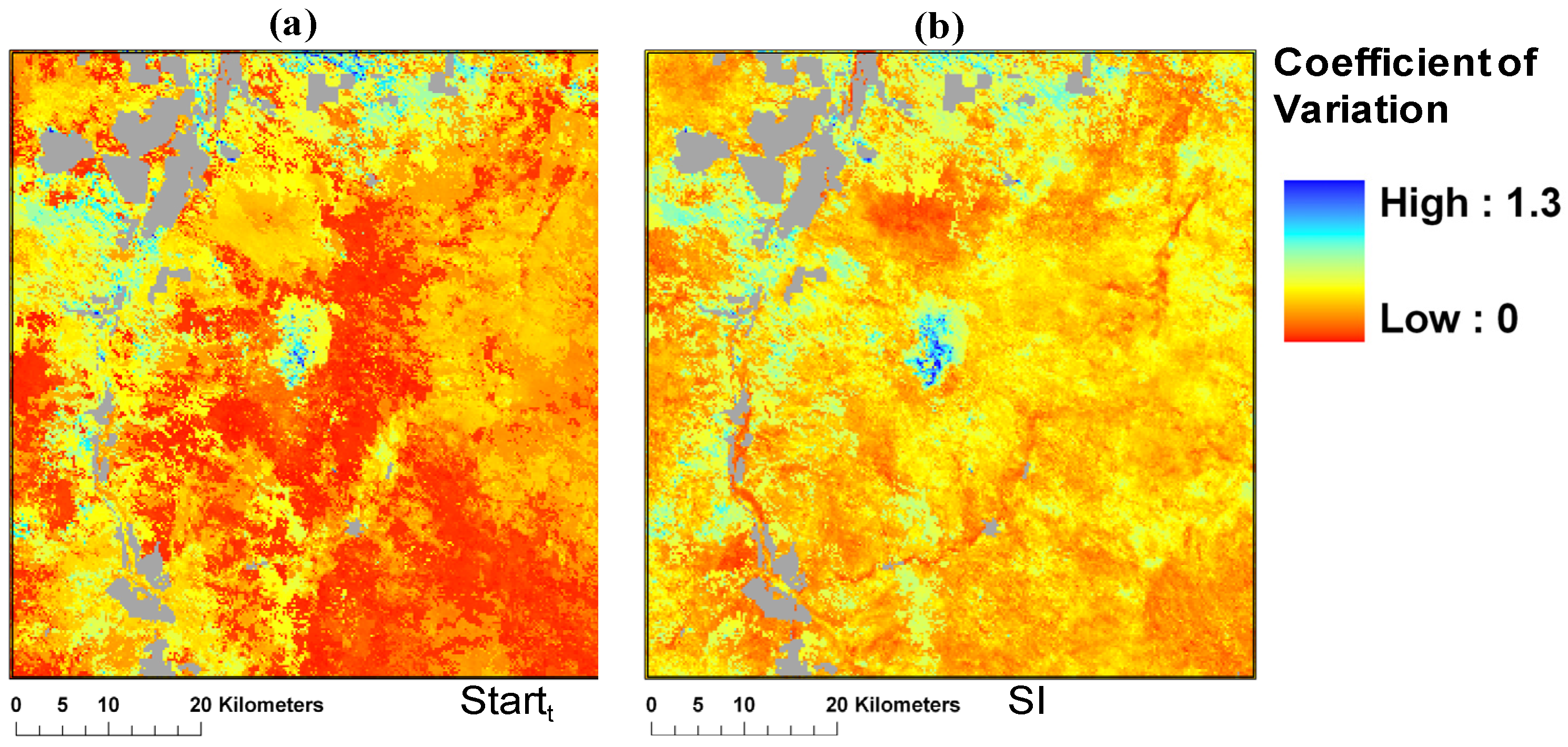
3.2. Spatio-Temporal Patterns of Phenometrics



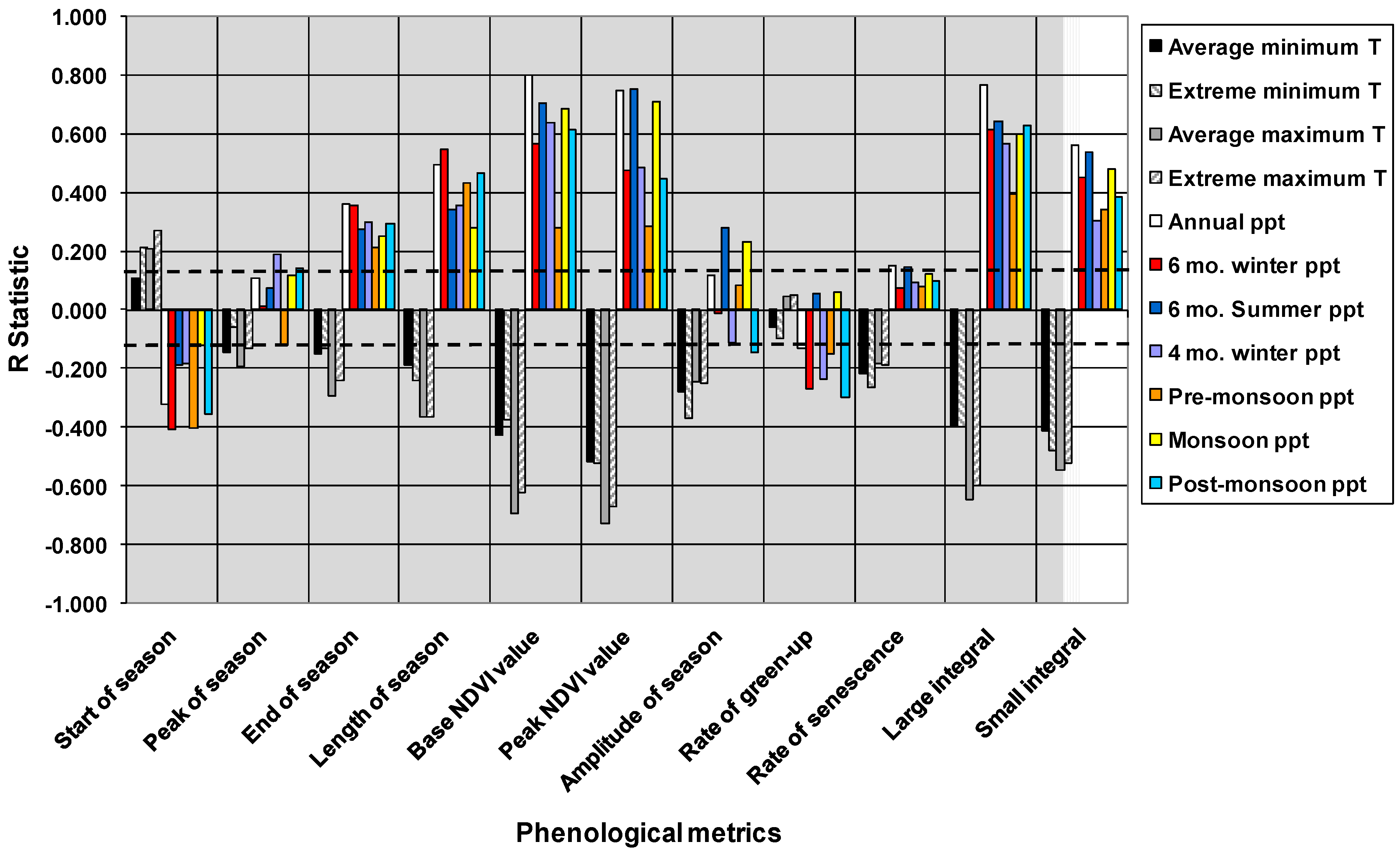
3.3. Phenometrics and Climate across Vegetation Communities

| Pheno-metric | Extreme minimum temp. | Extreme maximum temp. | Annual precip. | Winter precip.* | Pre-monsoon precip.** | Monsoon precip. | Post-monsoon precip.* | Whole model |
|---|---|---|---|---|---|---|---|---|
| Start of season | β = -2.25e-2 | β = 1.35 | β = 2.81e-2 | F3,234 = 24.6 | ||||
| t234 = -7.85 | t234 = 2.84 | t234 = 6.46 | p < 0.0001 | |||||
| p < 0.0001 | p = 0.0050 | p < 0.0001 | R2 = 0.230 | |||||
| Peak of season | β = -2.02e-1 | β = 7.26e-1 | β = -2.14e-1 | F3,234 = 6.7 | ||||
| t234 = -2.53 | t234 = 2.24 | t234 = -3.57 | p = 0.002 | |||||
| p = 0.0122 | p = 0.0261 | p = 0.0004 | R2 = 0.067 | |||||
| End of season | β = 1.00e-2 | F1,236 = 34.8 | ||||||
| t236 = 5.90 | p < 0.0001 | |||||||
| p < 0.0001 | R2 = 0.125 | |||||||
| Length of season | β = 3.10e-2 | β = -3.09e-2 | F2,235 = 59.4 | |||||
| t235 = 10.27 | t235 = -5.74 | p < 0.0001 | ||||||
| p < 0.0001 | p < 0.0001 | R2 = 0.330 | ||||||
| Base NDVI* | β = -7.45e-2 | β = 4.17e-3 | β = -7.44e-2 | β = -3.31e-3 | β = -1.57e-1 | F5,232 = 127.9 | ||
| t232 = -8.48 | t232 = 11.41 | t232 = -8.08 | t232 = -6.53 | t232 = -4.84 | p < 0.0001 | |||
| p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | R2 = 0.744 | |||
| Peak NDVI | β = -1.68e-2 | β = -1.65e-2 | β = 9.78e-4 | β = -1.11e-2 | β = -8.10e-2 | F5,232 = 114.9 | ||
| t232 = -4.35 | t232 = -4.18 | t232 = 13.86 | t232 = -5.15 | t232 = -7.06 | p < 0.0001 | |||
| p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | R2 = 0.706 | |||
| Amplitude of season | β = -1.54e-2 | β = -2.87e-4 | β = 9.44e-3 | β = 7.71e-4 | β = -3.74e-2 | F4,233 = 24.2 | ||
| t233 = -5.49 | t233 = -2.29 | t233 = 3.15 | t233 = 4.67 | t233 = -4.58 | p < 0.0001 | |||
| p < 0.0001 | p = 0.0230 | p = 0.0019 | p < 0.0001 | p < 0.0001 | R2 = 0.328 | |||
| Rate of Greenup | β = -2.99e-4 | β = 5.32e-3 | β = 4.95e-4 | F3,234 = 20.4 | ||||
| t234 = -7.31 | t234 = 4.39 | t234 = 7.37 | p < 0.0001 | |||||
| p < 0.0001 | p < 0.0001 | p < 0.0001 | R2 = 0.197 | |||||
| Rate of Senescence* | β = -9.64e-2 | F1,236 = 22.4 | ||||||
| t236 = -4.74 | p < 0.0001 | |||||||
| p < 0.0001 | R2 = 0.083 | |||||||
| Small Integral* | β = 9.49e-2 | β = 1.98e-3 | β = 1.68e-1 | F3,234 = 62.47 | ||||
| t234 = -6.89 | t234 = 8.03 | t234 = -3.65 | p < 0.0001 | |||||
| p < 0.0001 | p < 0.0001 | p = 0.0003 | R2 = 0.438 | |||||
| Large Integral* | β = -4.11e-2 | β = -3.54e-2 | β = 2.35e-3 | F2,234 = 127.6 | ||||
| t234 = -2.76 | t234 = -2.35 | t234 = 11.07 | p < 0.0001 | |||||
| p = 0.0063 | p = 0.0197 | p < 0.0001 | R2 = 0.616 |
3.4. Phenometrics and Climate by Vegetation Type
| Phenometric | Climate Metric | R | p-value | n | |
| Base NDVI value | Winter (6 mo) precipitation | 0.62 | 0.0005 | 27 | |
| Base NDVI value | Annual precipitation | 0.50 | 0.0076 | 27 | |
| End of season | Monsoon precipitation | −0.51 | 0.0061 | 27 | |
| Large integral | Winter (6 mo) precipitation | 0.63 | 0.0004 | 27 | |
| Large integral | Pre-monsoon precipitation | 0.61 | 0.0006 | 27 | |
| Length of season | Pre-monsoon precipitation | 0.64 | 0.0003 | 27 | |
| Peak NDVI value | Winter (6 mo) precipitation | 0.67 | 0.0001 | 27 | |
| Peak NDVI value | Annual precipitation | 0.52 | 0.0053 | 27 | |
| Rate of senescence | Winter(6 mo) precipitation | −0.54 | 0.0037 | 27 | |
| Small integral | Pre-monsoon precipitation | 0.55 | 0.0029 | 27 | |
| Start of season | Post-monsoon precipitation | −0.53 | 0.0044 | 27 | |
| Start of season | Pre-monsoon precipitation | −0.64 | 0.0004 | 27 |
| Phenometric | Climate Metric | R | p-value | n | |
|---|---|---|---|---|---|
| Amplitude of season | Summer precipitation | 0.59 | 0.0028 | 23 | |
| Base NDVI value | Annual precipitation | 0.74 | 0.0001 | 23 | |
| Base NDVI value | Winter (6 mo) precipitation | 0.58 | 0.0041 | 23 | |
| Base NDVI value | Average maximum temperature | −0.60 | 0.0025 | 23 | |
| End of season | Average minimum temperature | 0.64 | 0.0010 | 23 | |
| End of season | Average maximum temperature | 0.64 | 0.0011 | 23 | |
| End of season | Winter (6 mo) precipitation | −0.59 | 0.0030 | 23 | |
| Large integral | Annual precipitation | 0.73 | 0.0001 | 23 | |
| Large integral | Summer precipitation | 0.70 | 0.0002 | 23 | |
| Large integral | Pre-monsoon precipitation | 0.58 | 0.0034 | 23 | |
| Large integral | Extreme maximum temperature | −0.54 | 0.0072 | 23 | |
| Large integral | Extreme minimum temperature | −0.63 | 0.0013 | 23 | |
| Length of season | Pre-monsoon precipitation | 0.62 | 0.0015 | 23 | |
| Peak NDVI value | Summer precipitation | 0.64 | 0.0010 | 23 | |
| Peak of season | Average maximum temperature | 0.54 | 0.0080 | 23 | |
| Peak of season | Pre-monsoon precipitation | −0.60 | 0.0027 | 23 | |
| Peak of season | Annual precipitation | −0.72 | 0.0001 | 23 | |
| Peak of season | Winter(6 mo) precipitation | −0.74 | 0.0000 | 23 | |
| Small integral | Summer precipitation | 0.75 | 0.0000 | 23 | |
| Small integral | Annual precipitation | 0.59 | 0.0031 | 23 | |
| Small integral | Monsoon precipitation | 0.56 | 0.0055 | 23 | |
| Start of season | Extreme maximum temperature | 0.59 | 0.0030 | 23 | |
| Start of season | Extreme minimum temperature | 0.55 | 0.0067 | 23 | |
| Start of season | Winter (6 mo) precipitation | −0.56 | 0.0054 | 23 | |
| Start of season | Pre-monsoon precipitation | −0.61 | 0.0022 | 23 |
4. Conclusions
Acknowledgements
References
- Reed, B.C.; Brown, J.F.; Vanderzee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Schwartz, M.D. Monitoring global change with phenology: the case of the spring green wave. Int. J. Biometeorol. 1994, 38, 18–22. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Nemani, R.; Keeling, C.; Hashimoto, H.; Jolly, W.; Piper, S.; Tucker, C.; Myneni, R.; Running, S. Climate driven increases in global terrestrial net primary production from 1982–1999. Science 2003, 300, 1562–1563. [Google Scholar] [CrossRef] [PubMed]
- Goward, S.N.; Tucker, C.J.; Dye, D.G. North American vegetation patterns observed with the NOAA-7 advanced very high resolution radiometer. Plant Ecol. 1985, 64, 3–14. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Envion. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Colwell, J.E. Vegetation Canopy Reflectance. Remote Sens. Environ. 1974, 3, 175–183. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981–1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- White, M.A.; Hoffman, F.; Hargrove, W.W.; Nemani, R.R. A global framework for monitoring phenological responses to climate change. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef]
- Gao, Y.; Mas, J.F.; Navarrete, A. The improvement of an object-oriented classification using multi-temporal MODIS EVI satellite data. Int. J. Dig. Earth 2009, 2, 219–236. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O'Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; Brown, J.F.; de Wit, A.; Schaepman, M.; Lin, X.M.; Dettinger, M.; Bailey, A.S.; Kimball, J.; Schwartz, M.D.; Baldocchi, D.D.; Lee, J.T.; Lauenroth, W.K. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Change Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D. Monitoring the effects of forest restoration treatments on post-fire vegetation recovery with MODIS multitemporal data. Sensors 2008, 8, 2017–2042. [Google Scholar] [CrossRef]
- Hwang, T.; Kangw, S.; Kim, J.; Kim, Y.; Lee, D.; Band, L. Evaluating drought effect on MODIS Gross Primary Production (GPP) with an eco-hydrological model in the mountainous forest, East Asia. Glob. Change Biol. 2008, 14, 1037–1056. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; Anderson, J.J.; Myers, O.B.; Meyer, C.W. Regional vegetation die-off in response to global-change-type drought. Proc. Nat. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- White, M.A.; Nemani, R.R.; Thornton, P.E.; Running, S.W. Satellite evidence of phenological differences between urbanized and rural areas of the eastern United States deciduous broadleaf forest. Ecosystems 2002, 5, 260–273. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Reed, B.C.; White, M.; Brown, J.F. Remote Sensing Phenology; Kluwer Academic Publishers: Dordrecht, the Netherlands, 2003; pp. 365–381. [Google Scholar]
- Jönsson, P.; Eklundh, L. Seasonality extraction and noise removal by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remot. Sen. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT–a program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Warshall, P. The Madrean sky island archipelago: a planetary overview. In Biodiversity and management of the Madrean Archipelago: the Sky Islands of Southwestern United States and Northwestern Mexico; DeBano, F.L., Ffolliott, P.F., Ortega-Rubio, A., Gottfried, G.J., Hamre, R.H., Edminster, C.B., Eds.; General Technical Report RM-GTR-264; US Department of Agriculture, Forest Service: Fort Collins, CO, USA, 1994; pp. 408–415. [Google Scholar]
- Coblentz, D.D.; Riitters, K.H. Topographic controls on the regional-scale biodiversity of the south-western USA. J Biogeogr. 2004, 31, 1125–1138. [Google Scholar] [CrossRef]
- Madrean Pine-Oak Woodlands—Biodiversity Hotspots. Conservation International. Available online: http://www.biodiversityhotspots.org/xp/hotspots/pine_oak/Pages/biodiversity.aspx (accessed on October 12, 2009).
- Swetnam, T.W.; Baisan, C.H. Historical Fire Regime Patterns in the Southwestern United States since AD 1700; USDA Forest Service: Fort Collins, CO, USA, 1996; pp. 11–32. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Impacts, Adaptation and Vulnerability: Summary for Policymakers; IPCC Climate Change 2007; Intergovernmental Panel on Climate Change: Brussels, Belgium, 2007; pp. 1–23.
- NEON Report on first Workshop on the National Ecological Observatory Network (NEON); Archbold Biological Station: Lake Placid, FL, USA, 2000.
- DeLong, J.P.; Cox, S.W.; Cox, N.S. A comparison of avian use of high- and low-elevation sites during autumn migration in central New Mexico. J. Field Ornithol. 2005, 76, 326–333. [Google Scholar] [CrossRef]
- Alerstam, T.; Hedenstrom, A. The development of bird migration theory. J. Avian Biology 1998, 29, 343–369. [Google Scholar] [CrossRef]
- Brown-Mitic, C.; Shuttleworth, W.J.; Chawn Harlow, R.; Petti, J.; Burke, E.; Bales, R. Seasonal water dynamics of a sky island subalpine forest in semi-arid southwestern United States. J. Arid Environ. 2007, 69, 237–258. [Google Scholar] [CrossRef]
- Marshall, J.T. Birds of the Pine-Oak Woodland in Southern Arizona and Adjacent Mexico; Cooper Ornithological Society: Berkeley, CA, USA, 1957; p. 125. [Google Scholar]
- Whittaker, R.H.; Niering, W.A. Vegetation of Santa Catalina Mountains, Arizona. 5. Biomass, production, and diversity along the elevation gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef]
- Merriam, C.H.; Steineger, L. Results of a Biological Survey of the San Francisco Mountain Region and the Desert of the Little Colorado, Arizona; North American Fauna Report 3; US Department of Agriculture, Division of Ornithology and Mammalogy: Washington, DC, USA, 1890; pp. 1–136. [Google Scholar]
- Moulin, S.; Kergoat, L.; Viovy, N.; Dedieu, G. Global-scale assessment of vegetation phenology using NOAA/AVHRR satellite measurements. J. Climate 1997, 10, 1154–1170. [Google Scholar] [CrossRef]
- Justice, C.O.; Vermote, E.; Townshend, J.R.G.; Defries, R.; Roy, D.P.; Hall, D.K.; Salomonson, V.V.; Privette, J.L.; Riggs, G.; Strahler, A.; Lucht, W.; Myneni, R.B.; Knyazikhin, Y.; Running, S.W.; Nemani, R.R.; Wan, Z.M.; Huete, A.R.; van Leeuwen, W.; Wolfe, R.E.; Giglio, L.; Muller, J.P.; Lewis, P.; Barnsley, M.J. The Moderate Resolution Imaging Spectroradiometer (MODIS): Land remote sensing for global change research. IEEE Trans. Geosci. Remote Sens. 1998, 36, 1228–1249. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Casady, G.M.; van Leeuwen, W.J.D.; Marsh, S.E. Evaluation post-wildfire vegetation regeneration as a response to multiple environmental determinants. Environ. Model. Assess. 2010, in press. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D.; Casady, G.M.; Neary, D.G.; Bautista, S.; Alloza, J.A.; Carmel, Y.; Wittenberg, L.; Malkinson, D.; Orr, B.J. Monitoring post-wildfire vegetation response with remotely sensed time-series data in Spain, USA and Israel. Int. J. Wildland Fire 2010, in press. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Global Biogeochem. Cycle. 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Bradley, B.A.; Jacob, R.W.; Hermance, J.F.; Mustard, J.F. A curve fitting procedure to derive inter-annual phenologies from time series of noisy satellite NDVI data. Remote Sens. Environ. 2007, 106, 137–145. [Google Scholar] [CrossRef]
- Morisette, J.T.; Richardson, A.D.; Knapp, A.K.; Fisher, J.I.; Graham, E.A.; Abatzoglou, J.; Wilson, B.E.; Breshears, D.D.; Henebry, G.M.; Hanes, J.M.; Liang, L. Tracking the rhythm of the seasons in the face of global change: phenological research in the 21st century. Front. Ecol. Environ. 2009, 7, 253–260. [Google Scholar] [CrossRef]
- Crimmins, T.M.; Crimmins, M.A.; Bertelsen, C.D. Flowering range changes across an elevation gradient in response to warming summer temperatures. Glob. Change Biol. 2009, 15, 1141–1152. [Google Scholar] [CrossRef]
- Lowry, J.; Ramsey, R.D.; Thomas, K.; Schrupp, D.; Sajwaj, T.; Kirby, J.; Waller, E.; Schrader, S.; Falzarano, S.; Langs, L.; Manis, G.; Wallace, C.; Schulz, K.; Comer, P.; Pohs, K.; Rieth, W.; Velasquez, C.; Wolk, B.; Kepner, W.; Boykin, K.; O'Brien, L.; Bradford, D.; Thompson, B.; Prior-Magee, J. Mapping moderate-scale land-cover over very large geographic areas within a collaborative framework: a case study of the Southwest Regional Gap Analysis Project (SWReGAP). Remote Sens. Environ. 2007, 108, 59–73. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W.J.D. A comparison of Vegetation Indices over a Global Set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- DiLuzio, M.; Johnson, G.L.; Daly, C.; Eischeid, J.K.; Arnold, J.G. Constructing retrospective gridded daily precipitation and temperature datasets for the conterminous United States. J. Appl. Meteorol. Climatol. 2008, 47, 475–497. [Google Scholar] [CrossRef]
- Crimmins, M.A.; Comrie, A.C. Interactions between antecedent climate and wildfire variability across south-eastern Arizona. Int. J. Wildland Fire 2004, 13, 455–466. [Google Scholar] [CrossRef]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-surface remote sensing of spatial and temporal variation in canopy phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Theresa, M.C.; Michael, A.C.; Bertelsen, C.D. Flowering range changes across an elevation gradient in response to warming summer temperatures. Glob. Change Biol. 2009, 15, 1141–1152. [Google Scholar]
- DeFries, R.; Hansen, M.; Townshend, J. Global discrimination of land cover types from metrics derived from AVHRR pathfinder data. Remote Sens. Environ. 1995, 54, 209–222. [Google Scholar] [CrossRef]
- Weiss, J.L.; Gutzler, D.S.; Coonrod, J.E.A.; Dahm, C.N. Long-term vegetation monitoring with NDVI in a diverse semi-arid setting, central New Mexico, USA. J. Arid Environ. 2004, 58, 249–272. [Google Scholar] [CrossRef]
- Schultz, P.A.; Halpert, M.S. Global correlation of temperature, NDVI and precipitation. Adv. Space Res. 1993, 13, 277–280. [Google Scholar] [CrossRef]
- Linderman, M.; Rowhani, P.; Benz, D.; Serneels, S.; Lambin, E.F. Land-cover change and vegetation dynamics across Africa. J. Geophys. Res.-Atmos. 2005, 110, D12. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Noy-Meir, I. Desert ecosystems: environment and producers. Ann. Rev. Ecol. Syst. 1973, 4, 25–52. [Google Scholar] [CrossRef]
- Sankoh, A.J.; Huque, M.F.; Dubey, S.D. Some comments on frequently used multiple endpoint adjustments methods in clinical trials. Stat. Med. 1997, 16, 2529–2542. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jonsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982-2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Loik, M.E.; Breshears, D.D.; Lauenroth, W.K.; Belnap, J. A multi-scale perspective of water pulses in dryland ecosystems: climatology and ecohydrology of the western USA. Oecologia 2004, 141, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Archibald, S.; Scholes, R.J. Leaf green-up in a semi-arid African savanna–separating tree and grass responses to environmental cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernandez, R.J. Modifying the ‘pulse-reserve’ paradigm for deserts of North America: precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Ignace, D.D.; Huxman, T.E.; Weltzin, J.F.; Williams, D.G. Leaf gas exchange and water status responses of a native and non-native grass to precipitation across contrasting soil surfaces in the Sonoran Desert. Oecologia 2007, 152, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Reed, B.; White, M.A. Assessing satellite-derived start-of-season (SOS) measures in the conterminous USA. Int. J. Climatol. 2002, 22, 1793–1805. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.; Schaaf, C. Sensitivity of vegetation phenology detection to the temporal resolution of satellite data. Int. J. Remote Sens. 2009, 30, 2061–2074. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Knight, J.F.; Lunetta, R.S.; Ediriwickrema, J.; Khorrarn, S. Regional scale land cover characterization using MODIS-NDVI 250 m multi-temporal imagery: a phenology-based approach. GISci. Remote Sens. 2006, 43, 1–23. [Google Scholar] [CrossRef]
- Loveland, T.R.; Merchant, J.W.; Brown, J.F.; Ohlen, D.O.; Reed, B.C.; Olson, P.; Hutchinson, J. Seasonal land-cover regions of the United States. Ann. Assn. Amer. Geogr. 1995, 85, 339–355. [Google Scholar] [CrossRef]
Appendix A
| Veetation community | Phenometric | Climate variable | Correlation (R) | p-value | n | Elevation (m) |
| Conifer-oak forest | Peak NDVI value | Winter precipitation (6 mo) | 0.90 | 0.0000 | 15 | 2626 |
| Conifer-oak forest | Base NDVI value | Winter precipitation (6 mo) | 0.87 | 0.0000 | 15 | 2626 |
| Conifer-oak forest | Peak NDVI value | Average maximum temperature | 0.84 | 0.0001 | 15 | 2626 |
| Conifer-oak forest | Peak NDVI value | Extreme maximum temperature | 0.80 | 0.0003 | 15 | 2626 |
| Conifer-oak forest | Small integral | Winter precipitation (4 mo) | 0.74 | 0.0017 | 15 | 2626 |
| Conifer-oak forest | Small integral | Average minimum temperature | 0.73 | 0.0020 | 15 | 2626 |
| Conifer-oak forest | Base NDVI value | Average maximum temperature | 0.71 | 0.0029 | 15 | 2626 |
| Conifer-oak forest | Base NDVI value | Extreme maximum temperature | 0.69 | 0.0041 | 15 | 2626 |
| Conifer-oak forest | Peak NDVI value | Average minimum temperature | 0.66 | 0.0074 | 15 | 2626 |
| Pine-oak woodlands | Peak NDVI value | Winter precipitation (6 mo) | 0.67 | 0.0001 | 27 | 2203 |
| Pine-oak woodlands | Large integral | Winter precipitation (6 mo) | 0.63 | 0.0004 | 27 | 2203 |
| Pine-oak woodlands | Start of season | Pre-monsoon precipitation | -0.64 | 0.0004 | 27 | 2203 |
| Pine-oak woodlands | Base NDVI value | Winter precipitation (6 mo) | 0.62 | 0.0005 | 27 | 2203 |
| Pine-oak woodlands | Large integral | Pre-monsoon precipitation | 0.61 | 0.0006 | 27 | 2203 |
| Pine-oak woodlands | Small integral | Pre-monsoon precipitation | 0.55 | 0.0029 | 27 | 2203 |
| Pine-oak woodlands | Rate of senescence | Winter precipitation (6 mo) | -0.54 | 0.0037 | 27 | 2203 |
| Pine-oak woodlands | Start of season | Post-monsoon precipitation | -0.53 | 0.0044 | 27 | 2203 |
| Pine-oak woodlands | Rate of green-up | Winter precipitation (4 mo) | -0.53 | 0.0047 | 27 | 2203 |
| Pine-oak woodlands | Peak NDVI value | Annual precipitation | 0.52 | 0.0053 | 27 | 2203 |
| Pine-oak woodlands | End of season | Monsoon precipitation | -0.51 | 0.0061 | 27 | 2203 |
| Pine-oak woodlands | Base NDVI value | Annual precipitation | 0.50 | 0.0076 | 27 | 2203 |
| Mogollon chaparral | Rate of green-up | Annual precipitation | 0.76 | 0.0040 | 12 | 1748 |
| Mogollon chaparral | Rate of senescence | Extreme minimum temperature | 0.76 | 0.0041 | 12 | 1748 |
| Mogollon chaparral | Start of season | Annual precipitation | -0.74 | 0.0057 | 12 | 1748 |
| Pinyon-Juniper | Peak NDVI value | Summer precipitation | 0.82 | 0.0000 | 18 | 1688 |
| Pinyon-Juniper | Peak NDVI value | Annual precipitation | 0.80 | 0.0001 | 18 | 1688 |
| Pinyon-Juniper | Base NDVI value | Annual precipitation | 0.80 | 0.0001 | 18 | 1688 |
| Pinyon-Juniper | Peak NDVI value | Monsoon precipitation | 0.78 | 0.0001 | 18 | 1688 |
| Pinyon-Juniper | Small integral | Summer precipitation | 0.66 | 0.0026 | 18 | 1688 |
| Pinyon-Juniper | Base NDVI value | Summer precipitation | 0.66 | 0.0027 | 18 | 1688 |
| Pinyon-Juniper | Large integral | Annual precipitation | 0.65 | 0.0038 | 18 | 1688 |
| Pinyon-Juniper | Rate of green-up | Winter precipitation (4 mo) | -0.63 | 0.0055 | 18 | 1688 |
| Pinyon-Juniper | Large integral | Summer precipitation | 0.62 | 0.0061 | 18 | 1688 |
| Pinyon-Juniper | Peak NDVI value | Post-monsoon precipitation | 0.62 | 0.0063 | 18 | 1688 |
| Pinyon-Juniper | Base NDVI value | Monsoon precipitation | 0.61 | 0.0069 | 18 | 1688 |
| Pinyon-Juniper | Base NDVI value | Post-monsoon precipitation | 0.61 | 0.0071 | 18 | 1688 |
| Madrean oak encinal | Peak NDVI value | Summer precipitation | 0.63 | 0.0009 | 24 | 1487 |
| Madrean oak encinal | Amplitude of season | Winter precipitation (4 mo) | -0.59 | 0.0022 | 24 | 1487 |
| Madrean oak encinal | Small integral | Summer precipitation | 0.54 | 0.0062 | 24 | 1487 |
| Madrean oak encinal | Peak NDVI value | Annual precipitation | 0.53 | 0.0074 | 24 | 1487 |
| Mixed salt desert | Small integral | Summer precipitation | 0.68 | 0.0003 | 24 | 1376 |
| Mixed salt desert | Base NDVI value | Extreme minimum temperature | -0.67 | 0.0004 | 24 | 1376 |
| Mixed salt desert | Base NDVI value | Winter precipitation (6 mo) | 0.63 | 0.0010 | 24 | 1376 |
| Mixed salt desert | Amplitude of season | Summer precipitation | 0.62 | 0.0014 | 24 | 1376 |
| Mixed salt desert | Base NDVI value | Average minimum temperature | -0.61 | 0.0015 | 24 | 1376 |
| Mixed salt desert | Peak NDVI value | Summer precipitation | 0.59 | 0.0022 | 24 | 1376 |
| Semi-desert grassland | Small integral | Summer precipitation | 0.75 | 0.0000 | 23 | 1357 |
| Semi-desert grassland | Peak of season | Winter precipitation (6 mo) | -0.74 | 0.0000 | 23 | 1357 |
| Semi-desert grassland | Base NDVI value | Annual precipitation | 0.74 | 0.0001 | 23 | 1357 |
| Semi-desert grassland | Large integral | Annual precipitation | 0.73 | 0.0001 | 23 | 1357 |
| Semi-desert grassland | Peak of season | Annual precipitation | -0.72 | 0.0001 | 23 | 1357 |
| Semi-desert grassland | Large integral | Summer precipitation | 0.70 | 0.0002 | 23 | 1357 |
| Semi-desert grassland | Peak NDVI value | Summer precipitation | 0.64 | 0.0010 | 23 | 1357 |
| Semi-desert grassland | End of season | Average minimum temperature | 0.64 | 0.0010 | 23 | 1357 |
| Semi-desert grassland | End of season | Average maximum temperature | 0.64 | 0.0011 | 23 | 1357 |
| Semi-desert grassland | Large integral | Extreme minimum temperature | -0.63 | 0.0013 | 23 | 1357 |
| Semi-desert grassland | Length of season | Pre-monsoon precipitation | 0.62 | 0.0015 | 23 | 1357 |
| Semi-desert grassland | Start of season | Pre-monsoon precipitation | -0.61 | 0.0022 | 23 | 1357 |
| Semi-desert grassland | Base NDVI value | Average maximum temperature | -0.60 | 0.0025 | 23 | 1357 |
| Semi-desert grassland | Peak of season | Pre-monsoon precipitation | -0.60 | 0.0027 | 23 | 1357 |
| Semi-desert grassland | Amplitude of season | Summer precipitation | 0.59 | 0.0028 | 23 | 1357 |
| Semi-desert grassland | Start of season | Extreme maximum temperature | 0.59 | 0.0030 | 23 | 1357 |
| Semi-desert grassland | End of season | Winter precipitation (6 mo) | -0.59 | 0.0030 | 23 | 1357 |
| Semi-desert grassland | Small integral | Annual precipitation | 0.59 | 0.0031 | 23 | 1357 |
| Semi-desert grassland | Large integral | Pre-monsoon precipitation | 0.58 | 0.0034 | 23 | 1357 |
| Semi-desert grassland | Base NDVI value | Winter precipitation (6 mo) | 0.58 | 0.0041 | 23 | 1357 |
| Semi-desert grassland | Start of season | Winter precipitation (6 mo) | -0.56 | 0.0054 | 23 | 1357 |
| Semi-desert grassland | Small integral | Monsoon precipitation | 0.56 | 0.0055 | 23 | 1357 |
| Semi-desert grassland | Start of season | Extreme minimum temperature | 0.55 | 0.0067 | 23 | 1357 |
| Semi-desert grassland | Large integral | Extreme maximum temperature | -0.54 | 0.0072 | 23 | 1357 |
| Semi-desert grassland | Peak of season | Average maximum temperature | 0.54 | 0.0080 | 23 | 1357 |
| Creosote thorn scrub | Peak of season | Average maximum temperature | 0.78 | 0.0001 | 18 | 1332 |
| Creosote thorn scrub | Start of season | Average maximum temperature | 0.71 | 0.0010 | 18 | 1332 |
| Creosote thorn scrub | Large integral | Pre-monsoon precipitation | 0.70 | 0.0011 | 18 | 1332 |
| Creosote thorn scrub | Start of season | Extreme maximum temperature | 0.70 | 0.0013 | 18 | 1332 |
| Creosote thorn scrub | Length of season | Winter precipitation (6 mo) | 0.68 | 0.0019 | 18 | 1332 |
| Creosote thorn scrub | Start of season | Pre-monsoon precipitation | -0.67 | 0.0024 | 18 | 1332 |
| Creosote thorn scrub | Peak of season | Extreme maximum temperature | 0.66 | 0.0029 | 18 | 1332 |
| Creosote thorn scrub | Rate of senescence | Winter precipitation (6 mo) | -0.66 | 0.0030 | 18 | 1332 |
| Creosote thorn scrub | Rate of green-up | Monsoon precipitation | 0.64 | 0.0039 | 18 | 1332 |
| Creosote thorn scrub | Length of season | Pre-monsoon precipitation | 0.64 | 0.0041 | 18 | 1332 |
| Creosote thorn scrub | Start of season | Winter precipitation (6 mo) | -0.63 | 0.0053 | 18 | 1332 |
| Creosote thorn scrub | Rate of green-up | Winter precipitation (6 mo) | -0.63 | 0.0053 | 18 | 1332 |
| Creosote thorn scrub | Rate of senescence | Winter precipitation (4mo) | -0.62 | 0.0060 | 18 | 1332 |
| Mid-elevation scrub | Amplitude of season | Post-monsoon precipitation | -0.73 | 0.0005 | 18 | 1150 |
| Mid-elevation scrub | Length of season | Extreme minimum temperature | -0.72 | 0.0008 | 18 | 1150 |
| Mid-elevation scrub | Large integral | Extreme minimum temperature | -0.69 | 0.0017 | 18 | 1150 |
| Mid-elevation scrub | End of season | Extreme minimum temperature | -0.68 | 0.0018 | 18 | 1150 |
| Mid-elevation scrub | End of season | Average minimum temperature | -0.63 | 0.0047 | 18 | 1150 |
| Mid-elevation scrub | Length of season | Average minimum temperature | -0.62 | 0.0065 | 18 | 1150 |
| Mid-elevation scrub | Base NDVI value | Pre-monsoon precipitation | 0.61 | 0.0069 | 18 | 1150 |
| Mesquite upland scrub | Peak NDVI value | Average minimum temperature | -0.59 | 0.0001 | 36 | 1143 |
| Mesquite upland scrub | Peak NDVI value | Extreme minimum temperature | -0.62 | 0.0001 | 36 | 1143 |
| Mesquite upland scrub | Small integral | Summer precipitation | 0.58 | 0.0002 | 36 | 1143 |
| Mesquite upland scrub | Start of season | Winter precipitation (6 mo) | -0.56 | 0.0004 | 36 | 1143 |
| Mesquite upland scrub | Rate of green-up | Extreme minimum temperature | -0.56 | 0.0004 | 36 | 1143 |
| Mesquite upland scrub | Peak of season | Pre-monsoon precipitation | -0.53 | 0.0009 | 36 | 1143 |
| Mesquite upland scrub | Rate of green-up | Average minimum temperature | -0.53 | 0.0010 | 36 | 1143 |
| Mesquite upland scrub | Amplitude of season | Extreme minimum temperature | -0.52 | 0.0013 | 36 | 1143 |
| Mesquite upland scrub | Start of season | Pre-monsoon precipitation | -0.52 | 0.0013 | 36 | 1143 |
| Mesquite upland scrub | Base NDVI value | Winter precipitation (6 mo) | 0.51 | 0.0014 | 36 | 1143 |
| Mesquite upland scrub | Base NDVI value | Average minimum temperature | -0.51 | 0.0016 | 36 | 1143 |
| Mesquite upland scrub | Rate of green-up | Summer precipitation | 0.50 | 0.0017 | 36 | 1143 |
| Mesquite upland scrub | Amplitude of season | Summer precipitation | 0.50 | 0.0020 | 36 | 1143 |
| Mesquite upland scrub | End of season | Extreme minimum temperature | 0.50 | 0.0021 | 36 | 1143 |
| Mesquite upland scrub | Rate of green-up | Monsoon precipitation | 0.50 | 0.0021 | 36 | 1143 |
| Mesquite upland scrub | Base NDVI value | Extreme minimum temperature | -0.49 | 0.0024 | 36 | 1143 |
| Mesquite upland scrub | End of season | Average minimum temperature | 0.49 | 0.0025 | 36 | 1143 |
| Mesquite upland scrub | Length of season | Winter precipitation | 0.49 | 0.0025 | 36 | 1143 |
| Mesquite upland scrub | Amplitude of season | Average minimum temperature | -0.49 | 0.0026 | 36 | 1143 |
| Mesquite upland scrub | Amplitude of season | Monsoon precipitation | 0.48 | 0.0028 | 36 | 1143 |
| Mesquite upland scrub | Peak NDVI value | Summer precipitation | 0.47 | 0.0035 | 36 | 1143 |
| Mesquite upland scrub | End of season | Extreme maximum temperature | 0.46 | 0.0048 | 36 | 1143 |
| Mesquite upland scrub | Rate of senescence | Monsoon precipitation | 0.46 | 0.0050 | 36 | 1143 |
| Mesquite upland scrub | Small integral | Annual precipitation | 0.46 | 0.0052 | 36 | 1143 |
| Mesquite upland scrub | Rate of senescence | Summer precipitation | 0.45 | 0.0053 | 36 | 1143 |
| Mesquite upland scrub | Peak of season | Winter precipitation (6 mo) | -0.44 | 0.0069 | 36 | 1143 |
| Mesquite upland scrub | Large integral | Winter precipitation (6 mo) | 0.44 | 0.0073 | 36 | 1143 |
| Paloverde-mixed cactus | Base NDVI value | Winter precipitation (6 mo) | 0.79 | 0.0000 | 23 | 965 |
| Paloverde-mixed cactus | Small integral | Average maximum temperature | 0.63 | 0.0013 | 23 | 965 |
| Paloverde-mixed cactus | Base NDVI value | Pre-monsoon precipitation | 0.61 | 0.0019 | 23 | 965 |
| Paloverde-mixed cactus | Length of season | Winter precipitation (6 mo) | 0.59 | 0.0028 | 23 | 965 |
| Paloverde-mixed cactus | Small integral | Summer precipitation | 0.59 | 0.0029 | 23 | 965 |
| Paloverde-mixed cactus | Base NDVI value | Annual precipitation | 0.57 | 0.0045 | 23 | 965 |
| Paloverde-mixed cactus | Start of season | Annual precipitation | -0.57 | 0.0049 | 23 | 965 |
| Paloverde-mixed cactus | End of season | Average minimum temperature | 0.54 | 0.0076 | 23 | 965 |
| Paloverde-mixed cactus | Amplitude of season | Monsoon precipitation | 0.54 | 0.0081 | 23 | 965 |
Appendix B
| Annual precipitation | Summer precipitation | Post-monsoon precipitation | Monsoon precipitation | Pre-monsoon precipitation | Winter precipitation (6 mo) | Winter precipitation (4 mo) | Extreme minimum temperature | Average minimum temperature | Extreme maximum temperature | Average maximum temperature | Total | |
| Base NDVI value | 8 | 2 | 7 | 2 | 4 | 3 | 6 | 3 | 5 | 4 | 5 | 49 |
| Peak NDVI value | 2 | 8 | 4 | 5 | 1 | 3 | 2 | 4 | 3 | 3 | 35 | |
| Amplitude of season | 4 | 6 | 6 | 3 | 4 | 3 | 4 | 2 | 2 | 34 | ||
| Start of season | 6 | 1 | 5 | 2 | 5 | 2 | 3 | 2 | 4 | 3 | 33 | |
| Large integral | 6 | 4 | 7 | 1 | 6 | 3 | 2 | 1 | 1 | 31 | ||
| Small integral | 7 | 5 | 8 | 2 | 1 | 3 | 1 | 2 | 29 | |||
| Peak of season | 3 | 2 | 4 | 1 | 3 | 1 | 3 | 3 | 2 | 3 | 2 | 27 |
| Rate of senescence | 9 | 1 | 2 | 1 | 5 | 1 | 2 | 3 | 1 | 1 | 26 | |
| Length of season | 6 | 1 | 1 | 5 | 2 | 3 | 2 | 3 | 1 | 24 | ||
| Rate of green-up | 1 | 5 | 1 | 2 | 2 | 1 | 3 | 3 | 1 | 1 | 20 | |
| End of season | 1 | 2 | 1 | 4 | 6 | 2 | 2 | 18 | ||||
| Total | 45 | 36 | 34 | 30 | 29 | 20 | 18 | 28 | 26 | 22 | 20 | 326 |
© 2010 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Van Leeuwen, W.J.D.; Davison, J.E.; Casady, G.M.; Marsh, S.E. Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data. Remote Sens. 2010, 2, 388-415. https://doi.org/10.3390/rs2020388
Van Leeuwen WJD, Davison JE, Casady GM, Marsh SE. Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data. Remote Sensing. 2010; 2(2):388-415. https://doi.org/10.3390/rs2020388
Chicago/Turabian StyleVan Leeuwen, Willem J.D., Jennifer E. Davison, Grant M. Casady, and Stuart E. Marsh. 2010. "Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data" Remote Sensing 2, no. 2: 388-415. https://doi.org/10.3390/rs2020388
APA StyleVan Leeuwen, W. J. D., Davison, J. E., Casady, G. M., & Marsh, S. E. (2010). Phenological Characterization of Desert Sky Island Vegetation Communities with Remotely Sensed and Climate Time Series Data. Remote Sensing, 2(2), 388-415. https://doi.org/10.3390/rs2020388