Evaluation of Multiple Spring Phenological Indicators of Yearly GPP and NEP at Three Canadian Forest Sites



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Flux Data
2.3. Satellite Data
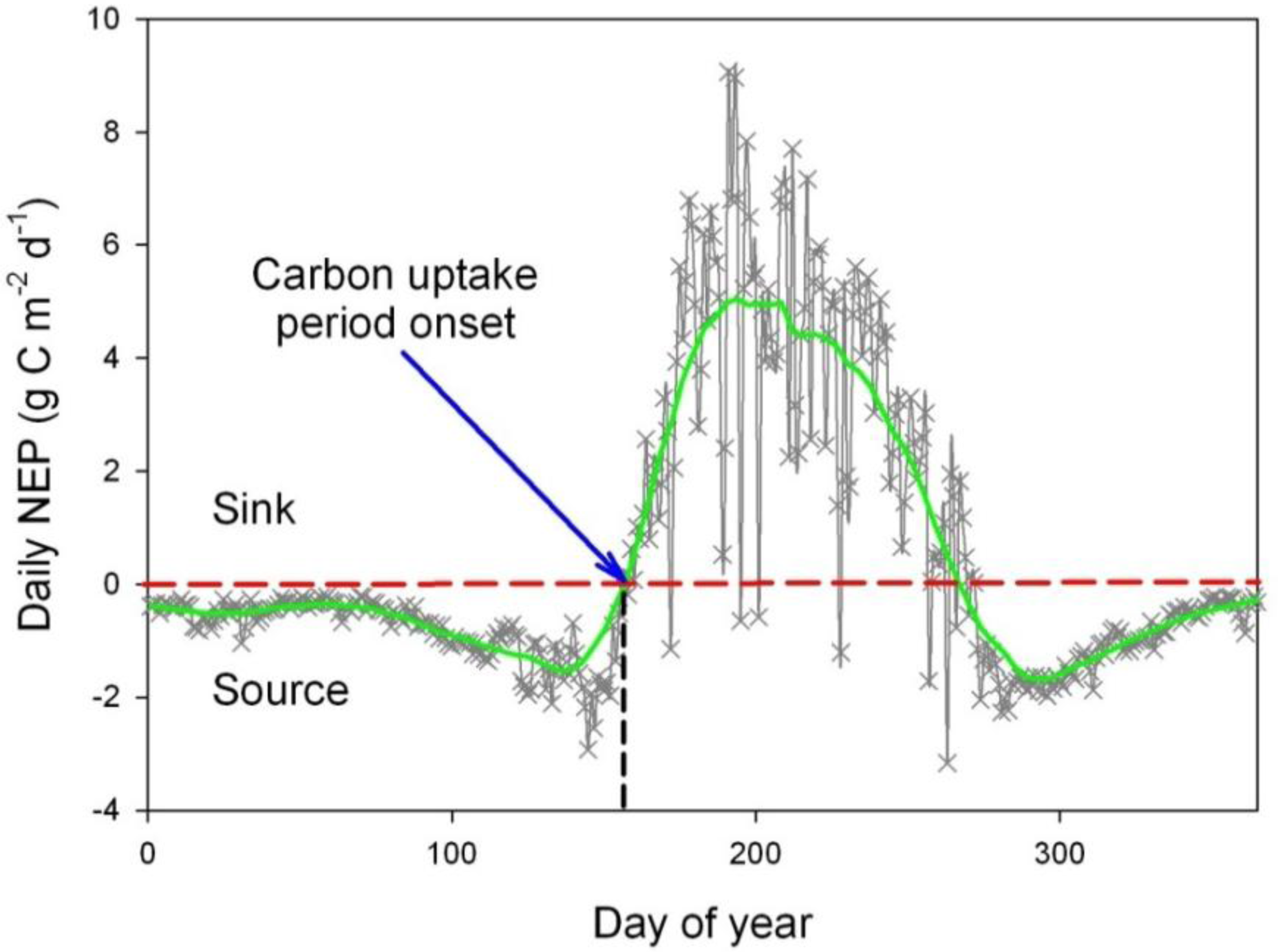
2.4. Spring Phenological Indicators
3. Results
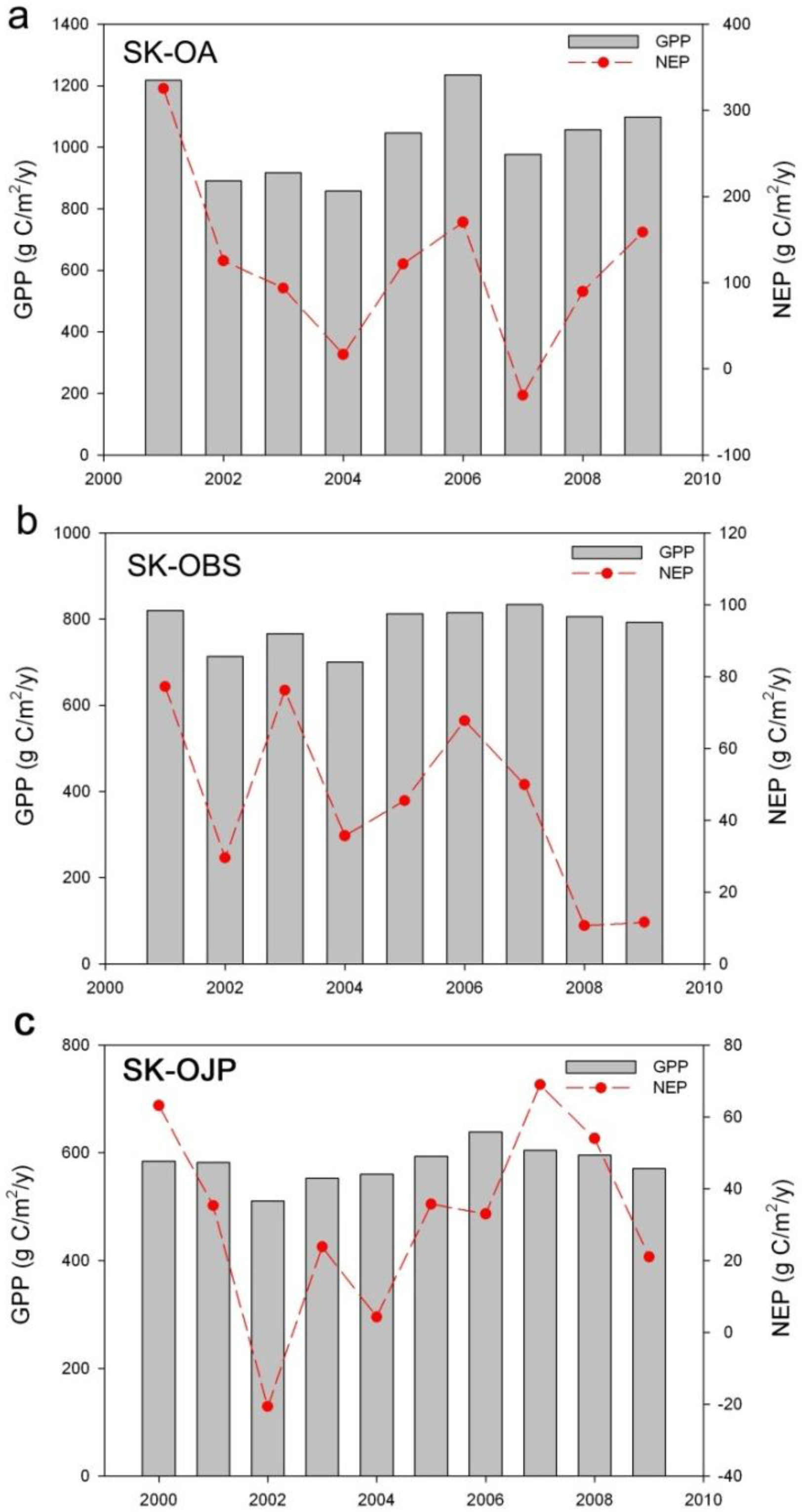
3.1. Descriptions of Annual C Fluxes
3.2. Spring Phenological Indicators across Sites and Intercorrelations among These Indicators
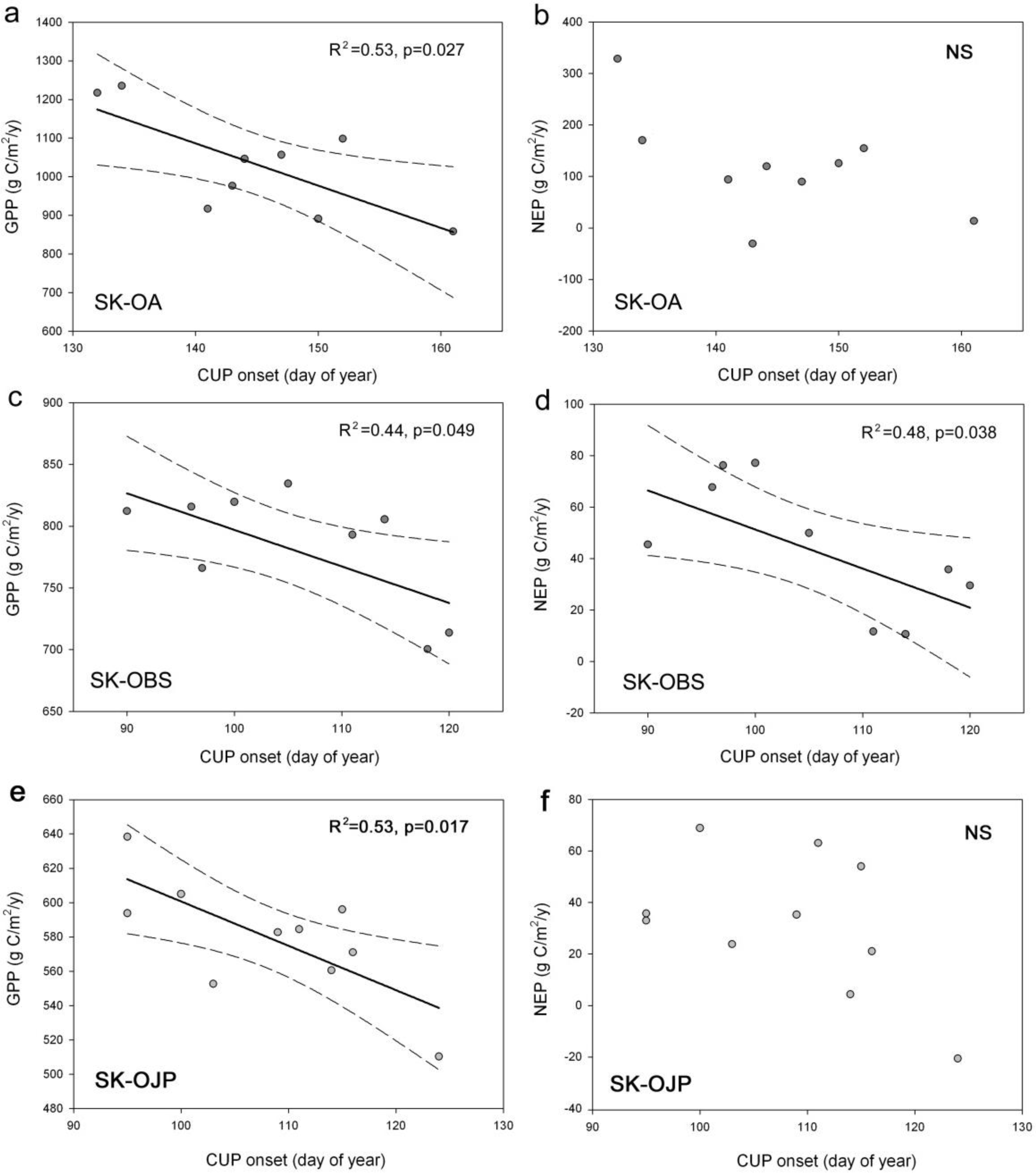
3.3. Relationships between CUP Onset and C Fluxes
3.4. Relationships between Spring Temperature and C Fluxes
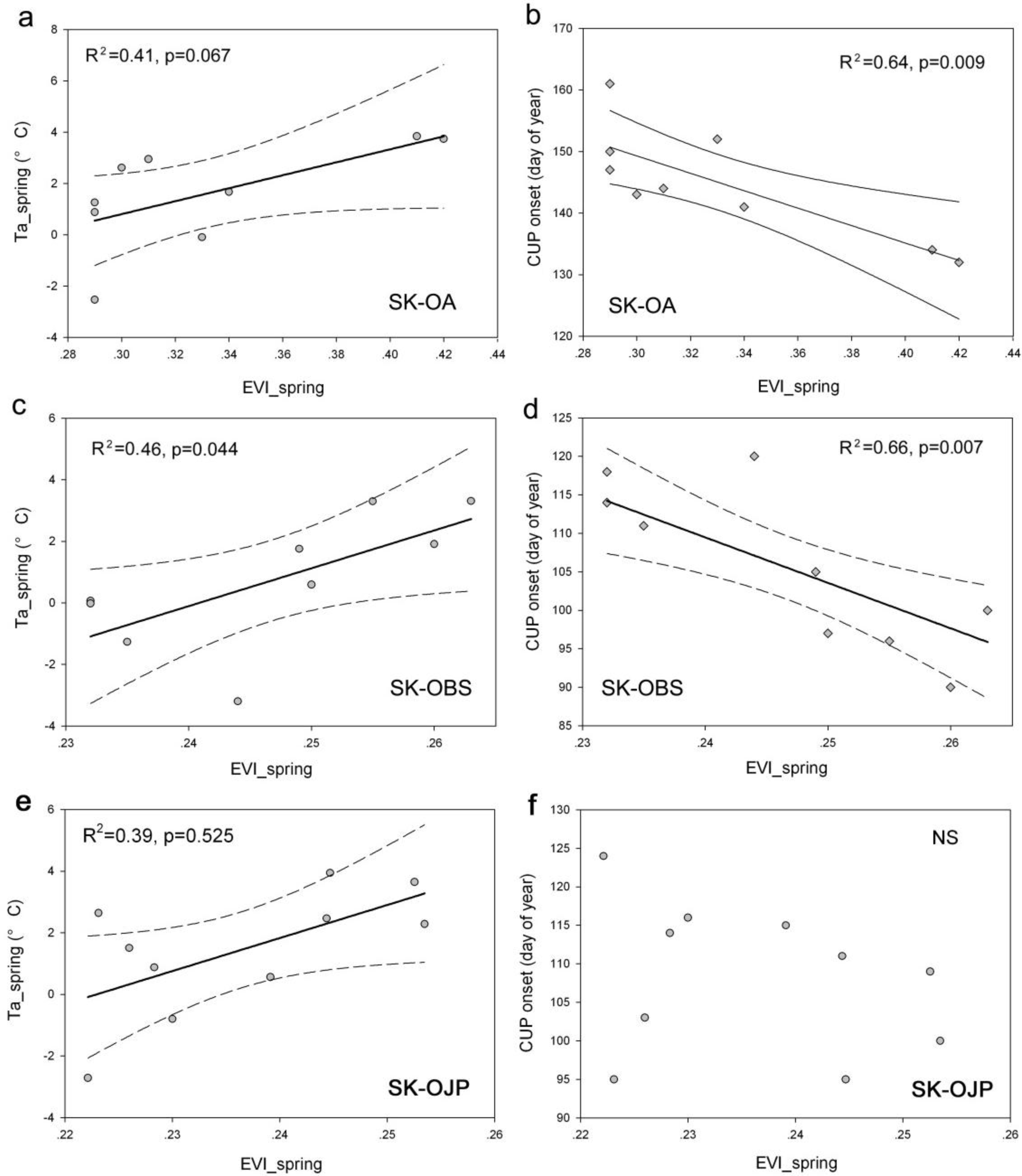
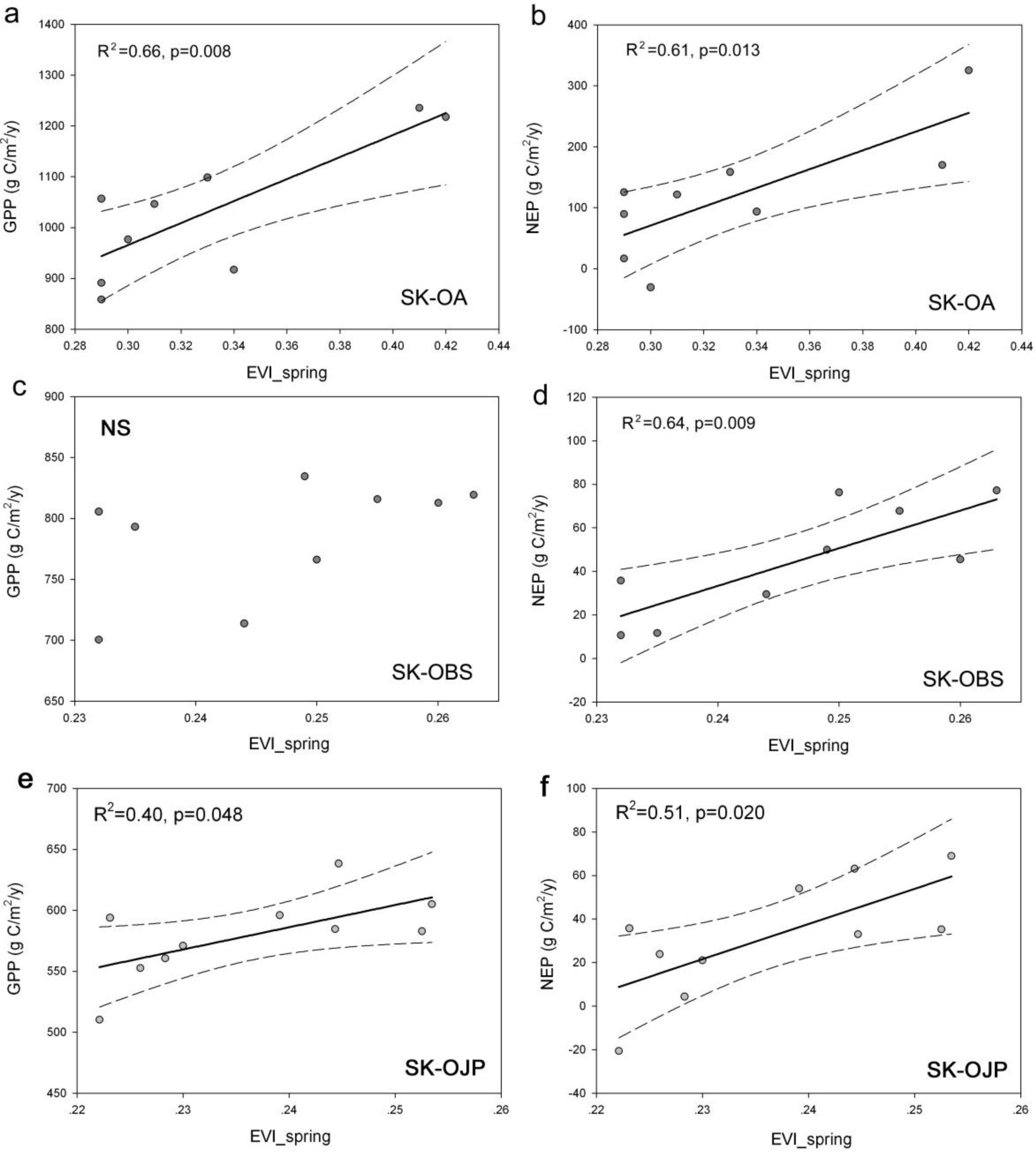
3.5. Relationships between Spring EVI and C Fluxes
4. Discussion
4.1. Comparison of the Spring Phenological Indicators
4.2. Impact of Plant Functional Type
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B-Biol. Sci 2010, 365, 3227–3246. [Google Scholar] [Green Version]
- Wu, C.; Chen, J.M.; Gonsamo, A.; Price, D.T.; Black, T.A.; Kurz, W.A. Interannual variability of carbon sequestration is determined by the lag between ends of net uptake and photosynthesis: Evidence from long records of two contrasting forest stands. Agric. For. Meteorol 2012, 164, 29–38. [Google Scholar]
- Gonsamo, A.; Chen, J.; Wu, C.; Dragoni, D. Predicting deciduous forest carbon uptake phenology by upscaling FLUXNET measurements using remote sensing data. Agric. For. Meteorol 2012, 165, 127–135. [Google Scholar]
- Garrity, S.R.; Maurer, K.D.; Mueller, K.L.; Vogel, C.S.; Curtis, P.S. A comparison of multiple phenology data sources for estimating seasonal transitions in deciduous forest carbon exchange. Agric. For. Meteorol 2011, 151, 1741–1752. [Google Scholar]
- Wu, C.; Chen, J.M.; Black, T.A.; Price, D.T.; Kurz, W.A.; Desai, A.R.; Gonsamo, A.; Jassal, R.S.; Gough, C.M.; Bohrer, G.; et al. Interannual variability of net ecosystem productivity in forests is explained by carbon flux phenology in autumn. Glob. Ecol. Biogeogr 2013, 22, 994–1006. [Google Scholar]
- Xiao, J.; Zhuang, Q.; Baldocchi, D.D.; Law, B.E.; Richardson, A.D.; Chen, J.; Oren, R.; Starr, G.; Noormets, A.; Ma, S.; et al. Estimation of net ecosystem carbon exchange of the conterminous United States by combining MODIS and AmeriFlux data. Agric. For. Meteorol 2008, 148, 1827–1847. [Google Scholar]
- Yuan, W.P.; Luo, Y.Q.; Richardson, A.D.; Oren, R.; Luyssaert, S.; Janssens, I.A.; Ceulemans, R.; Zhou, X.H.; Grunwald, T.; Aubinet, M.; et al. Latitudinal patterns of magnitude and interannual variability in net ecosystem exchange regulated by biological and environmental variables. Glob. Chang. Biol 2009, 15, 2905–2920. [Google Scholar]
- White, M.A.; Running, S.W.; Thornton, P.E. The impact of growing-season length variability on carbon assimilation and evapotranspiration over 88 years in the eastern US deciduous forest. Int. J. Biometeorol 1998, 42, 139–145. [Google Scholar]
- White, M.A.; Nemani, R.R. Canopy duration has little influence on annual carbon storage in the deciduous broad leaf forest. Glob. Chang. Biol 2003, 9, 967–972. [Google Scholar]
- Wu, C.; Gonsamo, A.; Chen, J.M.; Kurz, W.A.; Price, D.T.; Lafleur, P.M.; Jassal, R.S.; Dragoni, D.; Bohrer, G.; Gough, C.M.; et al. Interannual and spatial impacts of phenological transitions, growing season length, and spring and autumn temperatures on carbon sequestration: A North America flux data synthesis. Glob. Planet. Chang 2012, 92, 179–190. [Google Scholar]
- Thomas, C.K.; Law, B.E.; Irvine, J.; Martin, J.G.; Pettijohn, J.C.; Davis, K.J. Seasonal hydrology explains interannual and seasonal variation in carbon and water exchange in a semiarid mature ponderosa pine forest in central Oregon. J. Geophys. Res 2009, 114. [Google Scholar] [CrossRef]
- Medvigy, D.; Wofsy, S.C.; Munger, J.W.; Hollinger, D.Y.; Moorcroft, P.R. Mechanistic scaling of ecosystem function and dynamics in space and time: The ecosystem demography model version 2. J. Geophys. Res 2009, 114. [Google Scholar] [CrossRef]
- Chen, J.; Chen, W.; Liu, J.; Cihlar, J.; Gray, S. Annual carbon balance of Canada’s forests during 1895–1996. Glob. Biogeochem. Cy 2000, 14, 839–849. [Google Scholar]
- Black, T.A.; Chen, W.J.; Barr, A.G.; Arain, M.A.; Chen, Z.; Nesic, Z.; Hogg, E.H.; Neumann, H.H.; Yang, P.C. Increased carbon sequestration by a boreal deciduous forest in years with a warm spring. Geophys. Res. Lett 2000, 27, 1271–1274. [Google Scholar]
- Morgenstern, K.; Black, T.A.; Humphreys, E.R.; Griffis, T.J.; Drewitt, G.B.; Cai, T.; Nesic, Z.; Spittlehouse, D.L.; Livingston, N.J. Sensitivity and uncertainty of the carbon balance of a Pacific Northwest Douglas-fir forest during an El Nino LaNina cycle. Agric. For. Meteorol 2004, 123, 201–219. [Google Scholar]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar]
- Baldocchi, D.B. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol 2003, 9, 479–492. [Google Scholar]
- Churkina, G.; Schimel, D.; Braswell, B.; Xiao, X. Spatial analysis of growing season length control over net ecosystem exchange. Glob. Chang. Biol 2005, 11, 1777–1787. [Google Scholar]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cy 2007, 21. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ 2003, 84, 471–475. [Google Scholar]
- Liang, L.; Schwartz, M.D.; Fei, S. Validating satellite phenology through intensive ground observation and landscape scaling in a mixed seasonal forest. Remote Sens. Environ 2011, 115, 143–157. [Google Scholar]
- Nagai, S.; Nasahara, K.N.; Muraoka, H.; Akiyama, T.; Tsuchida, S. Field experiments to test the use of the normalized-difference vegetation index for phenology detection. Agric. For. Meteorol 2010, 150, 152–160. [Google Scholar] [Green Version]
- Dash, J.; Jeganathan, C.; Atkinson, P.M. The use of MERIS terrestrial chlorophyll index to study spatio-temporal variation in vegetation phenology over India. Remote Sens. Environ 2010, 114, 1388–1402. [Google Scholar]
- Fisher, J.I.; Mustard, J.F. Cross-scalar satellite phenology from ground, Landsat, and MODIS data. Remote Sens. Environ 2007, 109, 261–273. [Google Scholar]
- Ganguly, S.; Friedl, M.A.; Tan, B.; Zhang, X.; Verma, M. Land surface phenology from MODIS: Characterization of the collection 5 global land cover dynamics product. Remote Sens. Environ 2010, 114, 1805–1816. [Google Scholar]
- Barr, A.G.; Black, T.A.; Hogg, E.H.; Kljun, N.; Morgenstern, K.; Nesic, Z. Interannual variability in the leaf area index of a boreal aspen-hazelnut forest in relation to net ecosystem production. Agric. For. Meteorol 2004, 126, 237–255. [Google Scholar]
- Barr, A.G.; Black, T.A.; Hogg, E.H.; Griffis, T.J.; Morgenstern, K.; Kljun, N.; Theede, A.; Nesic, Z. Climatic controls on the carbon and water balances of a boreal aspen forest, 1994–2003. Glob. Chang. Biol 2007, 13, 561–576. [Google Scholar]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol 2002, 113, 97–120. [Google Scholar]
- DAAC, MODIS Land Product Subsets. Available online: http://www.modis.ornl.gov/modis/index.cfm (accessed on 4 March 2014).
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ 2002, 83, 195–213. [Google Scholar]
- Sims, D.A.; Rahman, A.F.; Cordova, V.D.; El-Masri, B.Z.; Baldocchi, D.D.; Bolstad, P.V.; Flanagan, L.B.; Goldstein, A.H.; Hollinger, D.Y.; Misson, L.; et al. A new model of gross primary productivity for North American ecosystems based solely on the enhanced vegetation index and land surface temperature from MODIS. Remote Sens. Environ 2008, 112, 1633–1646. [Google Scholar]
- Wu, C.; Chen, J.M.; Huang, N. Predicting gross primary production from the enhanced vegetation index and photosynthetically active radiation: Evaluation and calibration. Remote Sens. Environ 2011, 115, 3424–3435. [Google Scholar]
- Chen, B.; Coops, N.C.; Fu, D.; Margolis, H.A.; Amiro, B.D.; Barr, A.G.; Black, T.A.; Arain, M.A.; Bourque, C.P.A.; Flanagan, L.B.; et al. Assessing eddy-covariance flux tower location bias across the Fluxnet-Canada research network based on remote sensing and footprint modeling. Agric. For. Meteorol 2011, 151, 87–100. [Google Scholar]
- Wu, C.; Chen, J.M.; Desai, A.R.; Hollinger, D.Y.; Arain, M.A.; Margolis, H.A.; Gough, C.M.; Staebler, R.M. Remote sensing of canopy light use efficiency in temperate and boreal forests of North America using MODIS imagery. Remote Sens. Environ 2012, 118, 60–72. [Google Scholar]
- Welp, L.R.; Randerson, J.T.; Liu, H.P. The sensitivity of carbon fluxes to spring warming and summer drought depends on plant functional type in boreal forest ecosystems. Agric. For. Meteorol 2007, 147, 172–185. [Google Scholar]
- Wan, Z. New refinements and validation of the MODIS land-surface temperature/emissivity products. Remote Sens. Environ 2008, 112, 59–74. [Google Scholar]
- Pouliot, D.; Latifovic, R.; Fernandes, R.; Olthof, I. Evaluation of compositing period and AVHRR and MERIS combination for improvement of spring phenology detection in deciduous forests. Remote Sens. Environ 2011, 115, 158–166. [Google Scholar]
- Gurung, R.B.; Breidt, F.J.; Dutin, A.; Ogle, S.M. Predicting Enhanced Vegetation Index (EVI) curves for ecosystem modeling applications. Remote Sens. Environ 2009, 113, 2186–2193. [Google Scholar]
- Wu, C.; Gough, C.M.; Gonsamo, A.; Chen, J.M. Evidence of autumn phenology control on annual net ecosystem productivity in two temperate deciduous forests. Ecol. Eng 2013, 60, 88–95. [Google Scholar]
- Euskirchen, E.S.; McGuire, A.D.; Kicklighter, D.W.; Davidm, W.; Zhuang, Q.; Clein, J.S.; Dargaville, R.J.; Dye, D.G.; Kimball, J.S.; McDonald, K.C.; et al. Importance of recent shifts in soil thermal dynamics on growing season length, productivity, and carbon sequestration in terrestrial high-latitude ecosystems. Glob. Chang. Biol 2006, 12, 731–750. [Google Scholar]
- Griffis, T.J.; Black, T.A.; Morgenstern, K.; Barr, A.G.; Nesic, Z.; Drewitt, G.B.; Gaumont-Guay, D.; McCaughey, J.H. Ecophysiological controls on the carbon balance of three southern boreal forests. Agric. For. Meteorol 2003, 117, 53–71. [Google Scholar]
- Makkonen, K.; Helmisaari, H.S. Fine root biomass and production in Scots pine stands in relation to stand age. Tree Physiol 2001, 21, 193–198. [Google Scholar]
- Saurette, D.D.; Chang, S.X.; Thomas, B.R. Autotrophic and heterotrophic respiration rates across a chronosequence of hybrid poplar plantations in northern Alberta. Can. J. Soil Sci 2008, 88, 261–272. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | CUP Onset (DOY) | Ta_Spring (°C) | EVI_Spring | |
|---|---|---|---|---|
| SK-OA | Minimum | 132 | −2.5 | 0.291 |
| Maximum | 161 | 3.8 | 0.422 | |
| Average | 145 | 1.6 | 0.331 | |
| SD | 9.0 | 2.0 | 0.051 | |
| SK-OBS | Minimum | 90 | −3.2 | 0.232 |
| Maximum | 120 | 3.3 | 0.263 | |
| Average | 106 | 0.7 | 0.245 | |
| SD | 10.6 | 2.1 | 0.012 | |
| SK-OJP | Minimum | 95 | −2.7 | 0.225 |
| Maximum | 124 | 3.9 | 0.254 | |
| Average | 108 | 1.4 | 0.237 | |
| SD | 9.7 | 2.0 | 0.011 | |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Q.; Zhang, L.; Wu, T.; Cen, Y.; Huang, C.; Tong, Q. Evaluation of Multiple Spring Phenological Indicators of Yearly GPP and NEP at Three Canadian Forest Sites. Remote Sens. 2014, 6, 1991-2007. https://doi.org/10.3390/rs6031991
Wang Q, Zhang L, Wu T, Cen Y, Huang C, Tong Q. Evaluation of Multiple Spring Phenological Indicators of Yearly GPP and NEP at Three Canadian Forest Sites. Remote Sensing. 2014; 6(3):1991-2007. https://doi.org/10.3390/rs6031991
Chicago/Turabian StyleWang, Qian, Lifu Zhang, Taixia Wu, Yi Cen, Changping Huang, and Qingxi Tong. 2014. "Evaluation of Multiple Spring Phenological Indicators of Yearly GPP and NEP at Three Canadian Forest Sites" Remote Sensing 6, no. 3: 1991-2007. https://doi.org/10.3390/rs6031991
APA StyleWang, Q., Zhang, L., Wu, T., Cen, Y., Huang, C., & Tong, Q. (2014). Evaluation of Multiple Spring Phenological Indicators of Yearly GPP and NEP at Three Canadian Forest Sites. Remote Sensing, 6(3), 1991-2007. https://doi.org/10.3390/rs6031991