1. Introduction
Although the exact area or extent is unclear, current trends suggest that UK forests and woodlands are subject to a greater threat from exotic phytopathogens than ever previously experienced [
1,
2]. One particular invasive phytopathogen,
Phytophthora ramorum, has caused large-scale infections of larch (
Larix spp.) trees in UK forestry, particularly across Southwest England, South Wales and Southwest Scotland [
3]. The nature of the infection and its foliar symptoms include discolouration and defoliation [
4], which are currently being used for manual aerial detection by tree-health surveyors during helicopter surveys. This highlights the potential application of remotely sensed datasets for
P. ramorum assessment in larch across the UK [
5].
In recent decades, airborne laser scanning (ALS), also referred to as lidar, has been increasingly applied in forestry [
6,
7,
8]. The three-dimensional nature of ALS data provides structural information on topography, canopy height, tree density and crown dimensions, which can be used to determine biophysical parameters and inform forest inventories [
9,
10,
11]. The high resolution and accuracy associated with ALS enable the extraction of forest parameters associated with individual tree crowns (ITCs) within the forest canopy [
12]. The ability to conduct crown-based analysis of remotely sensed data for forest environments provides the opportunity for the detailed study of forest condition and dynamics [
13].
A range of algorithms can be applied for ITC delineation from ALS data, which typically exploit structural differences present among tree tops, canopy boundaries and canopy spaces [
14]. These algorithms include the region growing [
15,
16] and watershed segmentations [
14] which often require the prior identification of treetops as seed inputs. From ALS data, treetops are typically located via the detection and filtering of local maxima [
17,
18], in CHMs these represent points where neighbouring pixels present equal or lower values in height [
19]. The performance of a particular ITC delineation algorithm is dependent on the characteristics of both tree canopies and input datasets, and the subsequent suitability of methods for a specific study should be considered [
20,
21,
22].
In the case of ITC delineation, ALS datasets are typically analysed in raster format as a canopy height model (CHM) [
23]. A CHM represents the canopy surface elevation and is computed by the subtraction of the digital terrain model (DTM) from the digital surface model (DSM) [
24,
25]. The selected pixel size for the CHM, can affect the potential performance of the ITC detection [
14,
26]. In cases where the resolution is too coarse, the ability to distinguish between the boundaries of neighbouring tree crowns can be lost where pixels contain multiple overlapping tree crowns. Conversely, when spatial resolution is too fine, excessive intra-crown height variability may cause an over-segmentation of canopies. Pouliot et al. [
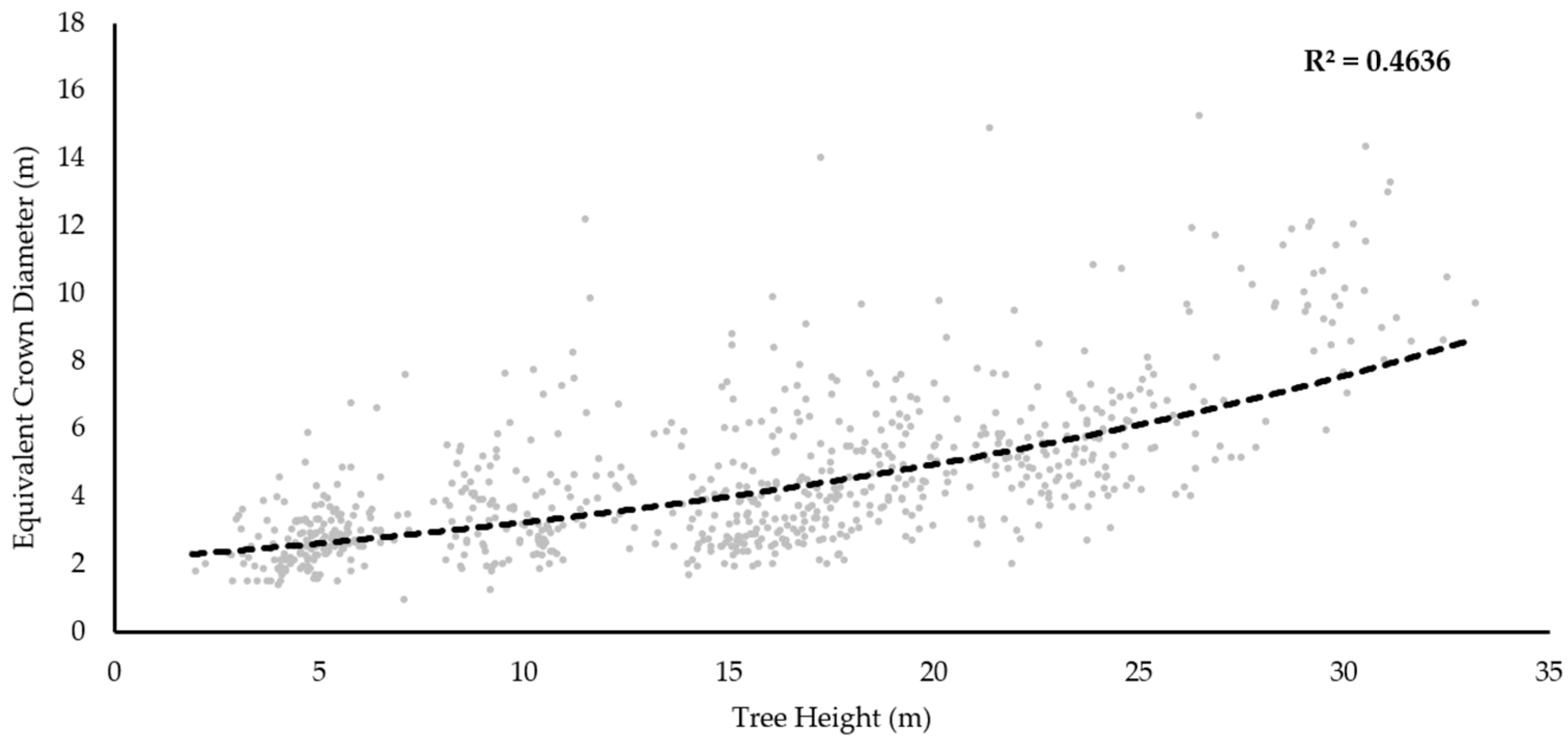
20] described the ratio between crown diameter and pixel size in order to address this point. The optimum ratio will vary in accordance with the sensitivity of segmentation algorithms to intra-crown irregularity and the distinction of crown boundaries, but can provide a useful tool in considering the influence of pixel size on ITC delineation success. Nevertheless, the insufficient availability of data regarding the relationship between pixel size and tree crown dimensions makes it difficult to determine specific recommendations for selection [
27]. In this paper we will consider the implications of varying CHM pixel size on the performance of ITC delineation across a range of crown dimensions.
Canopy height anomalies that are present in CHMs are known as data pits and can influence the accuracy of ITC detection [
25,
26]. Despite the uncertainty surrounding the specific source of these pits, cited causal factors have been acknowledged during the acquisition and processing of ALS datasets [
25,
28], these include: penetration of laser beams through the canopy, merging of ALS flight lines [
29], classification of ground and non-ground points [
30] and interpolation of point clouds to raster datasets [
31]. To address the problem, many studies have used image smoothing techniques such as Gaussian filtering, to remove intra-canopy elevation artefacts [
12,
19,
21]. More recent solutions have introduced the use of pit-filling algorithms [
25,
32] and the application of pit-free CHM generation methodologies [
26].
In forests subject to defoliation and dieback as a result of phytopathogens and pests, canopy structures are typically more complex and exhibit larger elevation irregularities across the canopy surface [
33,
34], causing an increased presence of data pits. Such characteristics can be useful in disease detection [
35,
36], but may also provide an added complication with regard to the isolation of ITCs for the assessment of crown deterioration. Consequently, the methodologies employed for ITC delineation in canopies affected by disease require consideration of data pits and their implications for segmentation accuracy.
In the case of phytopathological assessments from remotely sensed datasets, the application of an ITC approach presents several advantages for identifying areas that require phytosanitary interventions [
37]. In the early stages of the disease establishment in the forest stand, the isolation of ITCs, which initially succumb to infection, can enable a rapid response to pests and pathogens, which present new risks to forest areas [
38]. In studies of diseased forest landscapes, crown-based approaches can also facilitate the detailed study of heterogeneous patterns of infection [
39]. In addition, the use of ITC delineation techniques alongside species identification [
40,
41] can facilitate a targeted assessment of susceptible tree species [
39]. This combined approach also presents the potential for the identification of disease resistant individuals, which may prove particularly useful with regard to the breeding of resistant genotypes and the development of resilience in forest stock [
42]. Resultantly, in the case of
P. ramorum, the application of an ITC-based approach could therefore facilitate a detailed assessment of infected forested areas.
To address the difficulties in conducting ITC segmentation in partially or wholly defoliated forest canopies, this study aimed to determine the best method for the extraction of ITCs within P. ramorum infected larch forests using ALS data. In order to identify the most suitable approach the separate influences of segmentation method, CHM resolution and pit-removal were all considered.
4. Discussion
The absence of an optimal method or input CHM for ITC delineation across the study plots highlights the difficulties of utilising a single algorithm and input dataset to assess forest environments comprising of mixed stand ages and species [
12,
47]. Nevertheless, the results shown here allow us to infer several key points informing the selection of the most appropriate segmentation approach and CHM input for canopies subject to
P. ramorum infection.
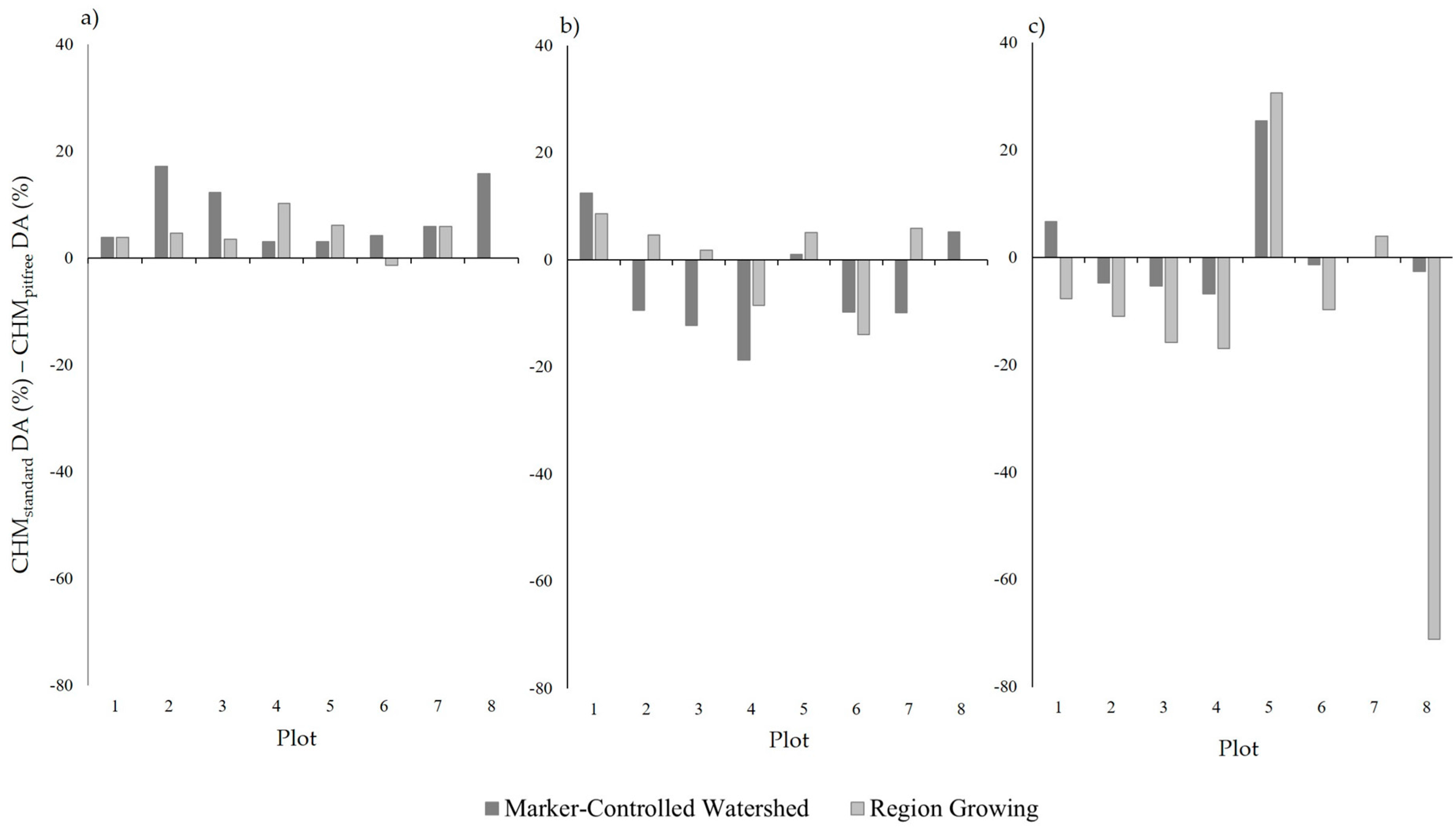
Neither CHM generation method (standard/pit-free) exhibited a consistently stronger performance, with each outperforming the other for four of the eight sample plots. This is different from the results observed by Khosravipour et al. [
26], who documented consistently improved tree top detection accuracy for all pit-free CHM inputs at pixel sizes of 0.15 m and 0.5 m. In our case, it is likely that the weaker performance of the pit-free CHMs in four of the sample plots may stem from the low-pass filtering used to smooth the canopy surface for improved local maxima detection. This was not applied in the study by Khosravipour et al. [
26], who instead extracted local maxima with an established variable window for coniferous forests [
51]. In addition, Khosravipour et al. [
26] assessed the accuracy of treetop detection, rather than the segmentation on ITCs. Nevertheless, in the case of the Plots 2, 3 and 4 which were subject to moderate and heavy
P. ramorum infection, the pit-free CHMs out performed those generated using the standard CHM. This improved performance is likely to be a result of the data pit filling during the CHM generation, providing a more complete canopy for image segmentation (
Figure 3).
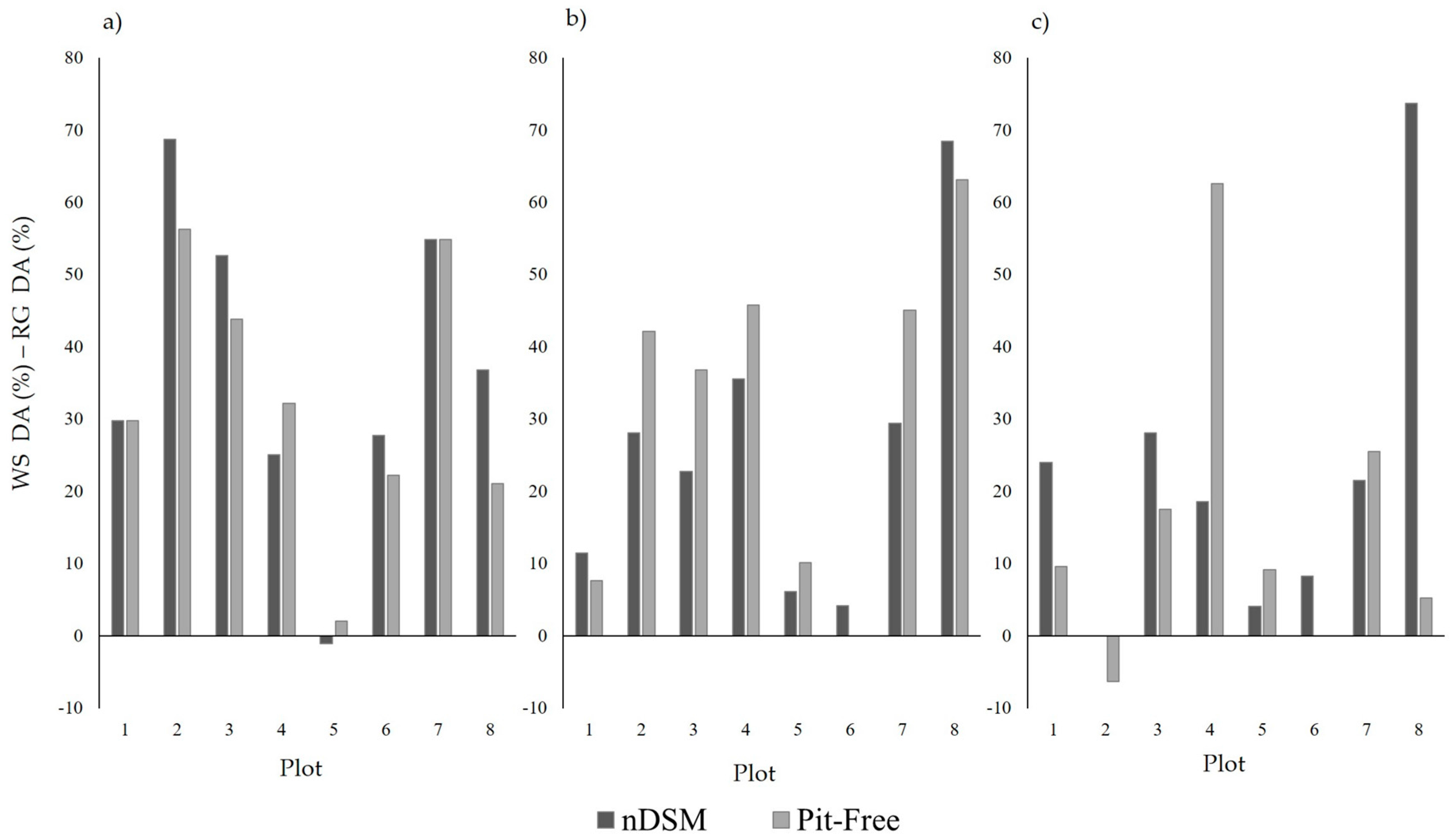
Between the two segmentation algorithms, the marker-controlled watershed demonstrated a superior performance (
p < 0.10) compared to the region growing segmentation, for both CHM generation methods and all CHM pixel sizes tested. Since both algorithms were provided with the same seed points, the difference in performance is due to their ability to delineate crown boundaries in the CHMs. In the case of the region growing segmentation, boundaries are delineated when a specified threshold value is reached. The poor overall performance of this particular segmentation algorithm is likely to have resulted from the inability of the region growing threshold to accommodate the variation in crown size and characteristics across the canopy, as a result of mixed species and tree ages [
50]. Although the region growing algorithm has previously produced satisfactory ITC delineations [
27,
72,
73,
74], studies have also noted difficulties and poor performance associated with the algorithm when applied in forest stands comprising of mixed species, multiple canopy layers and high tree densities [
27,
73,
74]. In comparison, the delineation of crown boundaries by the watershed segmentation directly utilises the height information for the CHMs when delineating watershed boundaries, providing a more informed segmentation [
58]. Studies from an array of forest types have noted successful isolation or segmentation of ITCs resulting from the marker-controlled watershed segmentation, examples include: commercially thinned conifer forest (75.6%) [
58], savanna woodland (64.1%) [
14] and eucalypts forest [
75]. Nevertheless, the delineation accuracies achieved by segmentation algorithms can vary due to a combination of factors including: the effectiveness of the algorithm, forest characteristics and ALS data acquisition and properties [
65]. Consequently, the difference between the performance of the marker-controlled watershed and region growing algorithms may change for segmentations performed for other forest environments and ALS datasets.
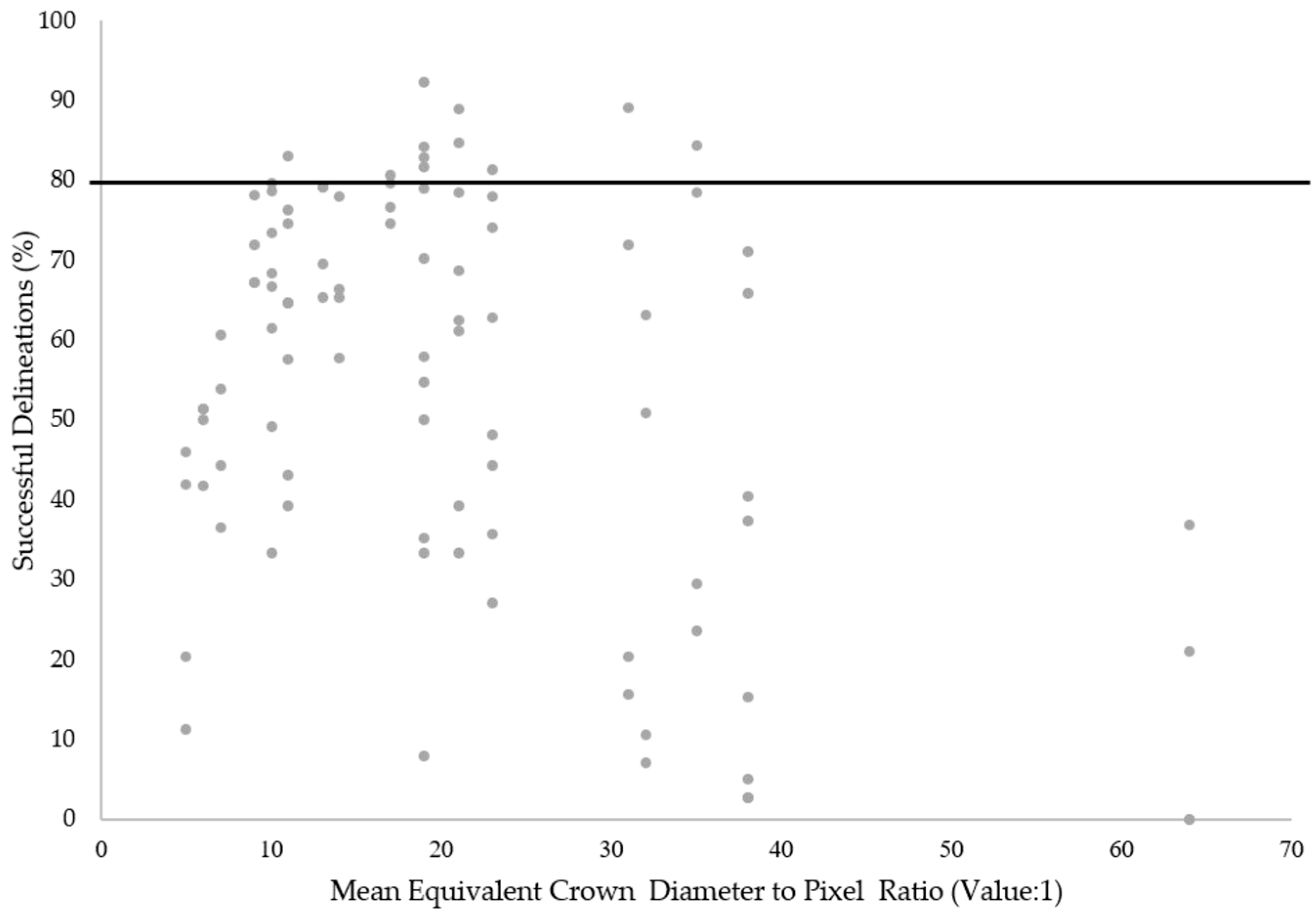
The results from the study also highlighted that CHM resolution used in the ITC segmentation strongly influences segmentation accuracy. Relationships between the maximum canopy height and optimum CHM pixel size were not unexpected given the strong influence of tree height on crown diameter [
14]. The suitability of certain pixel sizes for particular forest canopies has previously been explored in the literature [
20]. For example, with regard to crown diameter to pixel ratios, Pouliot et al. [
20] suggested a lower and upper limit of 3:1 and 19:1 respectively. In comparison, the ratios (mean equivalent crown diameter) that produced the best tree crown delineations across the sample plots were slightly higher, between 10:1 and 35:1. This may be as a result of reduced intra-canopy variation within the CHMs caused by the image smoothing filters applied [
12,
19]. In addition, these examples from Pouliot et al. [
20] were also suggested for imagery rather than CHMs produced from ALS. Previous studies have also considered the impact of CHM resolution on segmentation accuracy, Stereńczak et al. [
76] for example, noted no significant difference between the ITC segmentation results from the 0.25 m and 0.5 m resolution CHMs, however a significant reduction in the percentage of recognised ITCs was noted for the 1.0 m CHM. The results from previous research, in addition to those obtained in this study highlight the importance of selecting a suitable CHM pixel size for ITC segmentation. Nevertheless, in many cases the selected resolution of the CHMs utilised for ITC segmentations is governed by ALS point density [
26].
In relation to each of the CHM resolutions, several observed relationships require further discussion. Firstly, the 0.25 m CHM resolution performed consistently well across all sample plots, especially for the marker-controlled watershed segmentations, suggesting that this pixel size may be most suitable for the two study areas as a whole. This implies that at the 0.25 m resolution, enough detail is provided for the majority of ITCs to delineate boundaries without high levels intra-canopy variation resulting in over segmentations. However, in the cases of plots which exhibited a large maximum tree height (>30 m), the application of a lower resolution CHM (0.5 m) typically facilitated a greater percentage of successful ITC delineations. Again, the level of intra-crown variation provided in the CHM is likely to be the causal factor for this observation. With regard to the higher resolution CHMs (0.15 m) tested, a relationship between pixel size suitability and maximum tree height was also evident. In this instance, plots characterised by a small maximum tree height (<20 m) in general produced higher percentages of successful ITC delineations.
Nevertheless, it is important to note, that while these criteria explain the characteristic relationship between pixel size, maximum tree height and successful ITC delineation across the sample plots, variability in the observed trend was also evident in the dataset. This suggests that other plot characteristics such as variation in tree size and tree density may also influence the suitability of a particular CHM pixel size for ITC segmentation [
50]. Hence, it should be acknowledged that, while crown diameter should be recognised as a dominant variable influencing the suitability of pixel sizes for ITC delineation, consideration should also be given to other forest characteristics. In addition, it should also be recognised that the level of intra-canopy variation is also controlled by the filtering of the CHMs before segmentation [
14]. Consequently, relationships between the performance of CHMs at different resolutions and tree height may be altered for CHMs subject to varying degrees of smoothing.
The research demonstrates that ITCs within larch stands affected by
P. ramorum can be successfully delineated (>70%) using a pit-free CHM generation methodology, a marker-controlled watershed segmentation and the selection of an appropriate CHM pixel size. Nevertheless, whilst these methods provide successful results at the selected study sites, further testing would be required to consider the performance these methods for defoliated canopies of other tree species in different forest environments. In addition, preliminary testing was used to identify the most suitable parameters in the filtering and segmentation processes, however such parameters can significantly influence final segmentation results and may not be best suited for other forest environments and ALS datasets [
14,
72]. Furthermore, an additional limitation to the study can also be noted with regard to the use of two separate study areas for the comparison between healthy and infected stands. Although larch dominated plots were selected to best match with regard to tree height parameters, variations in tree density and species composition may have also influenced the performance of ITC segmentations across the two sites.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}