Dietary Fatty Acids Alter Lipid Profiles and Induce Myocardial Dysfunction without Causing Metabolic Disorders in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Statistical Analysis
3. Results
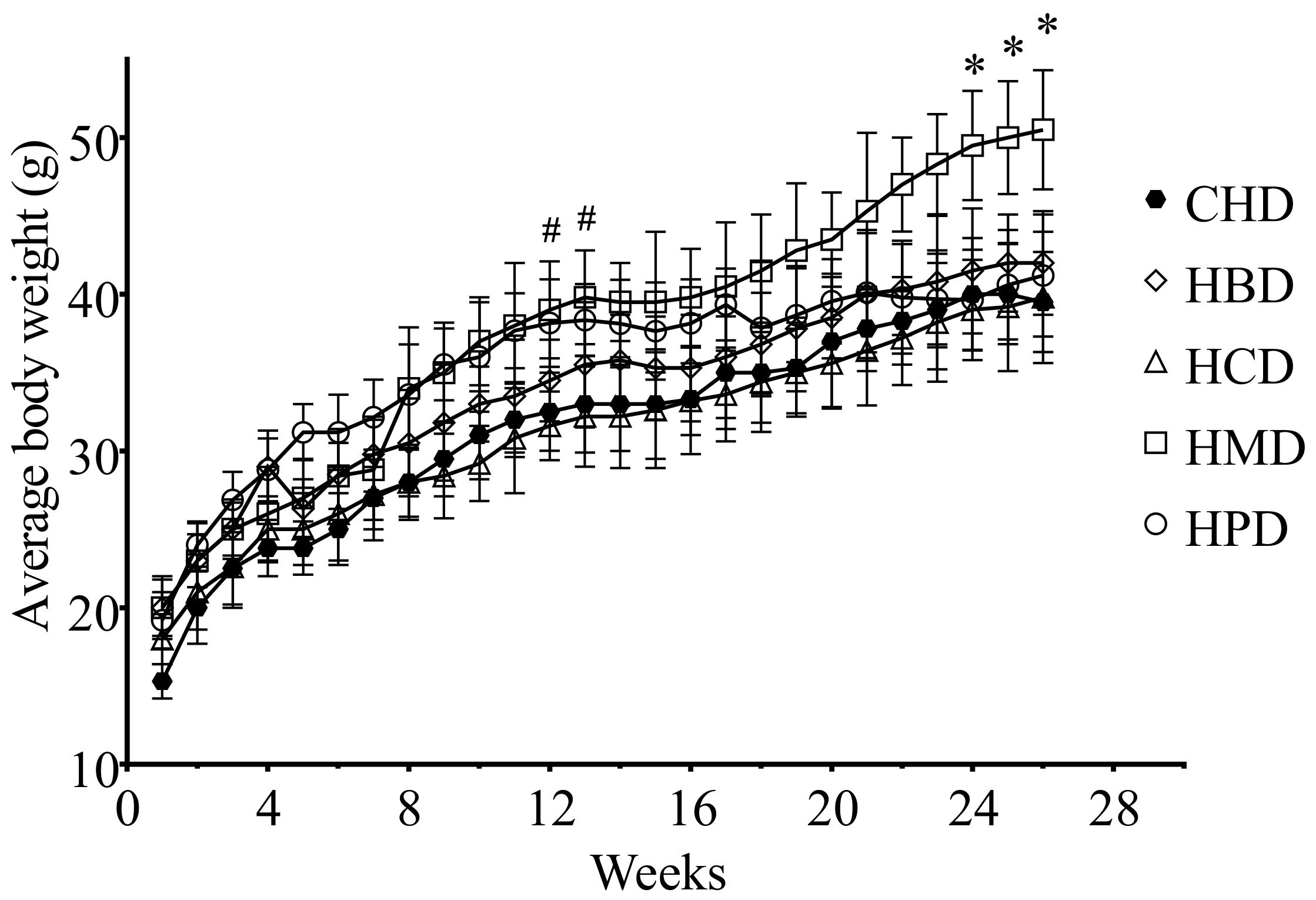
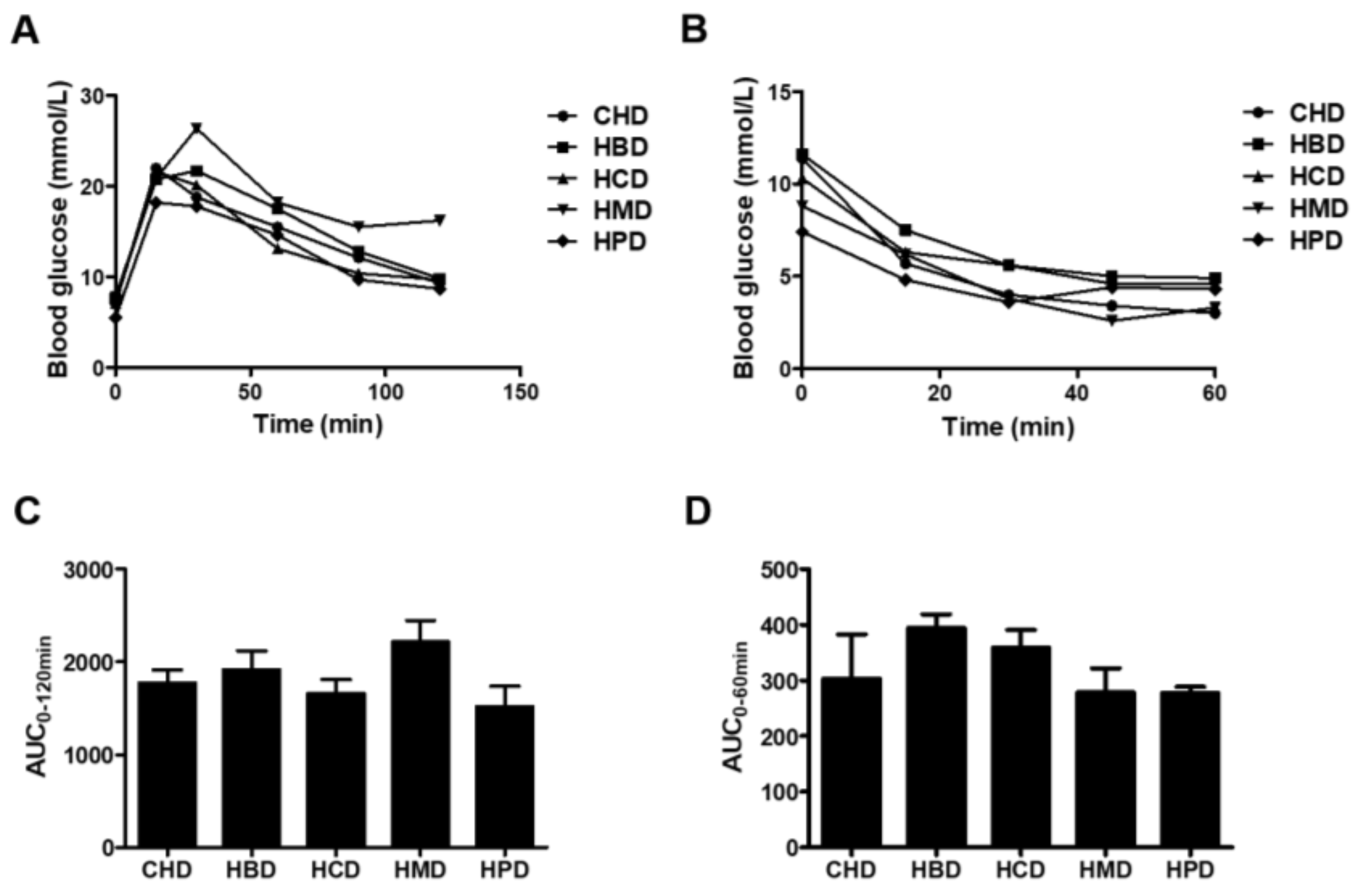
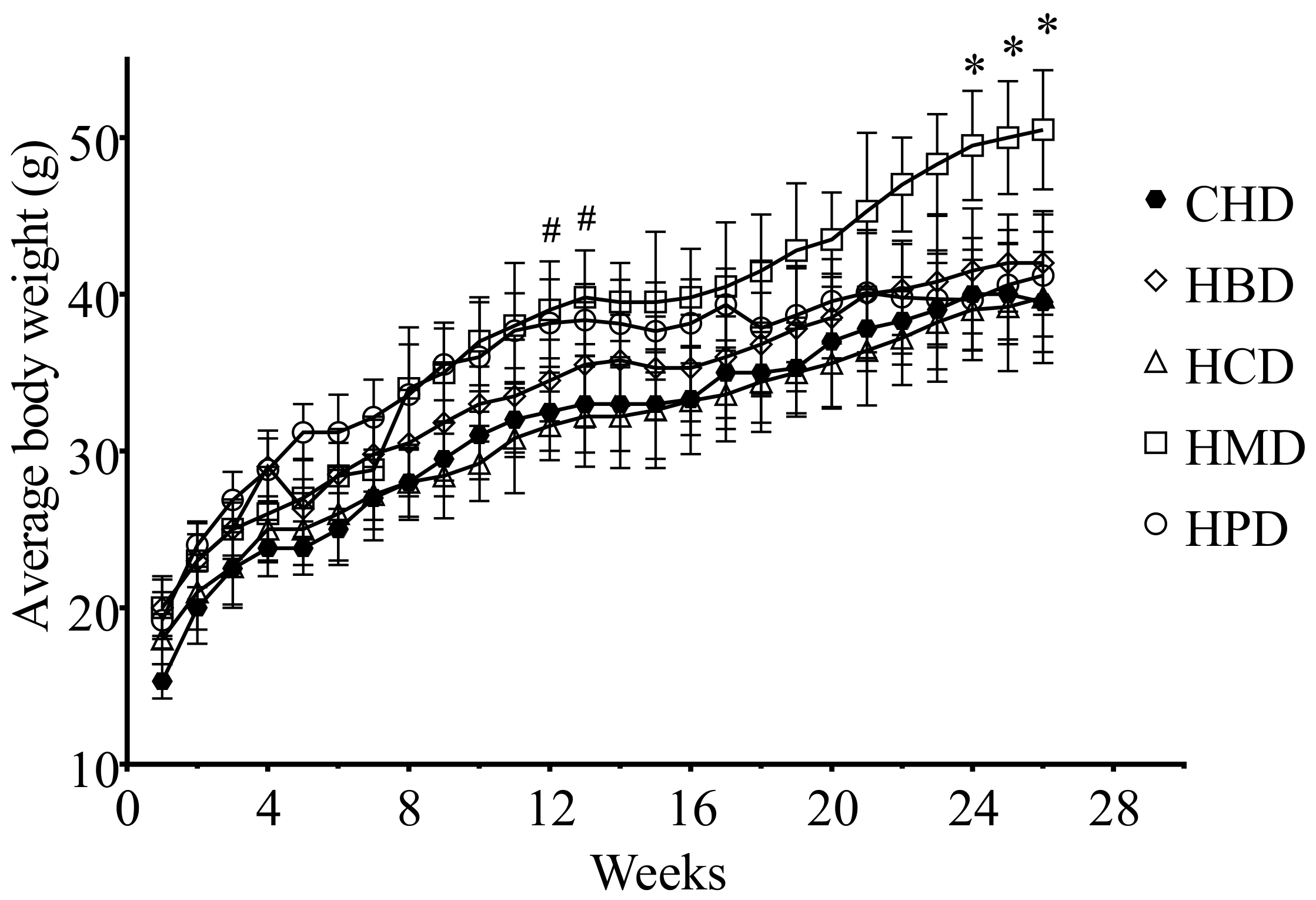
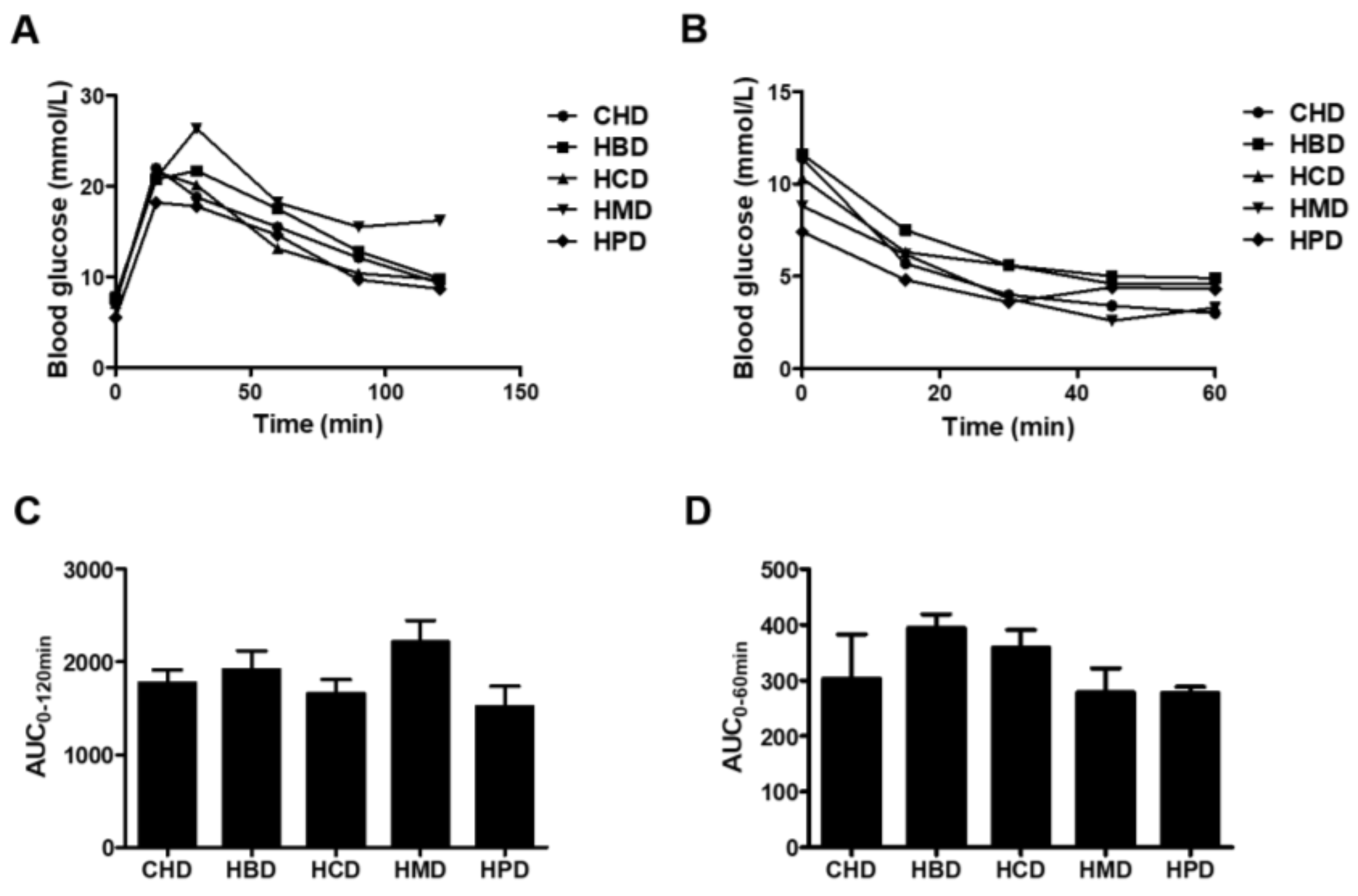
3.1. Metabolic Characteristicsin Mice Fed High SFA Diets
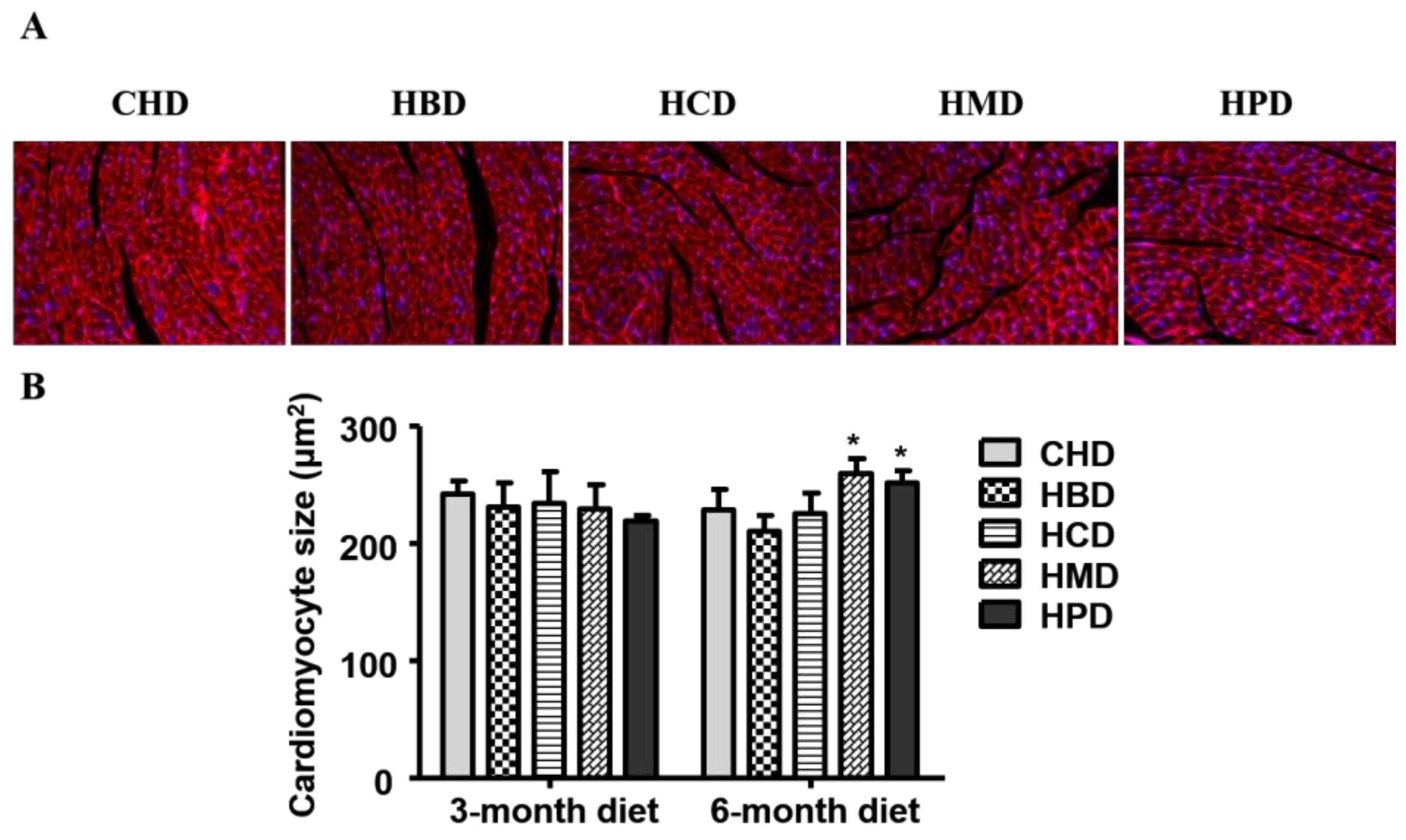
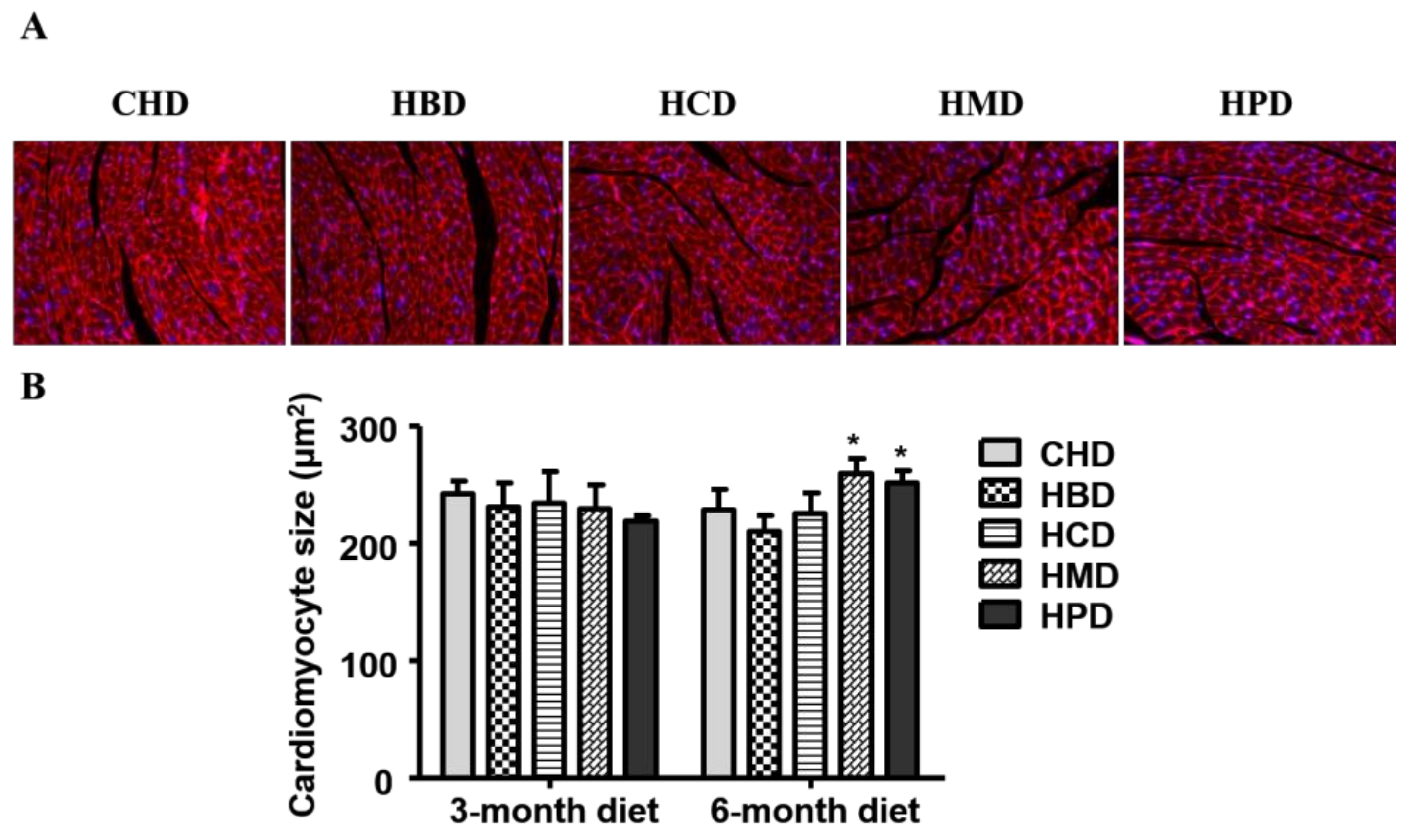
3.2. Cardiomyocyte Cross-Sectional Area in Mice Fed High SFA Diets
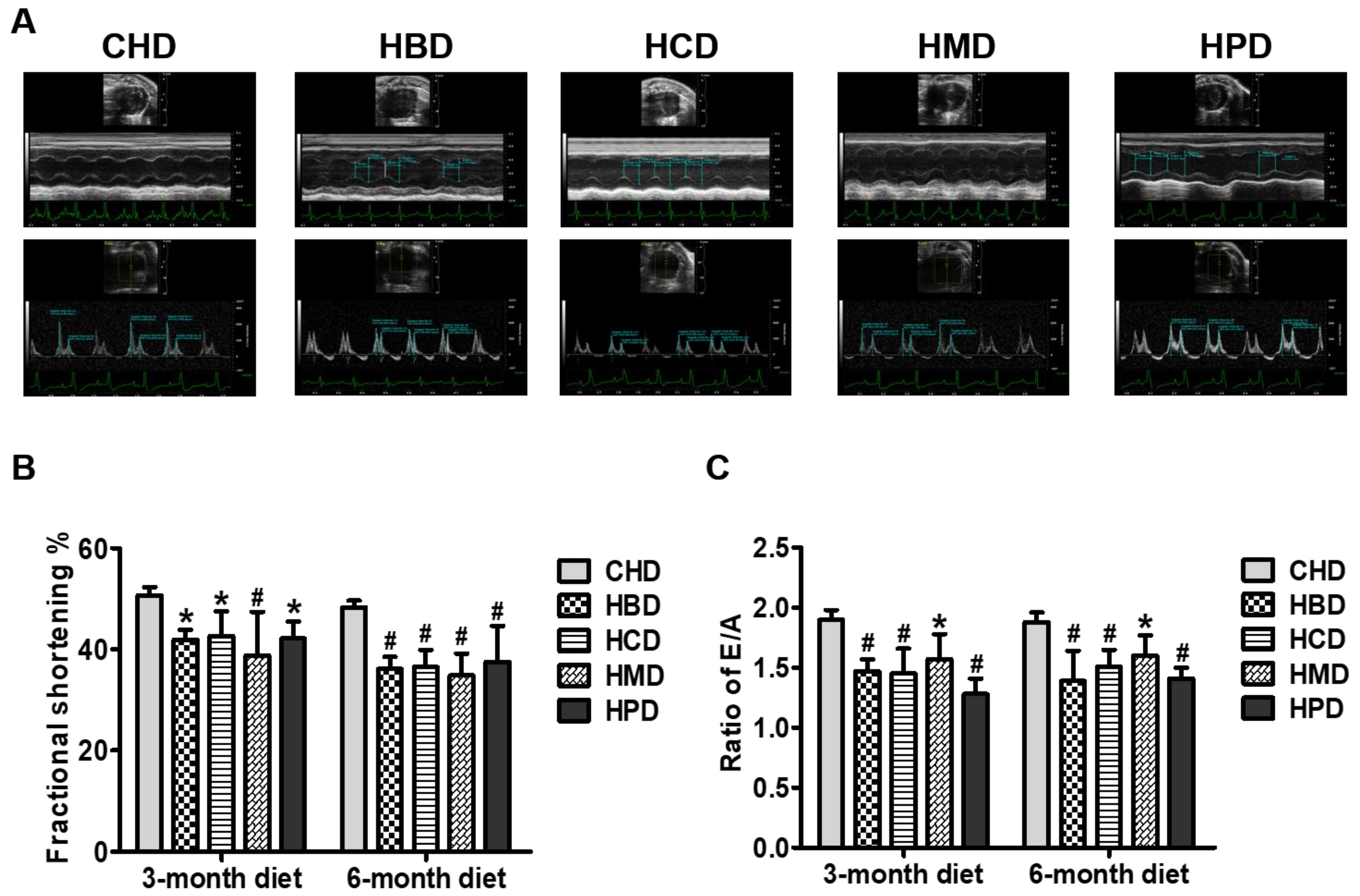
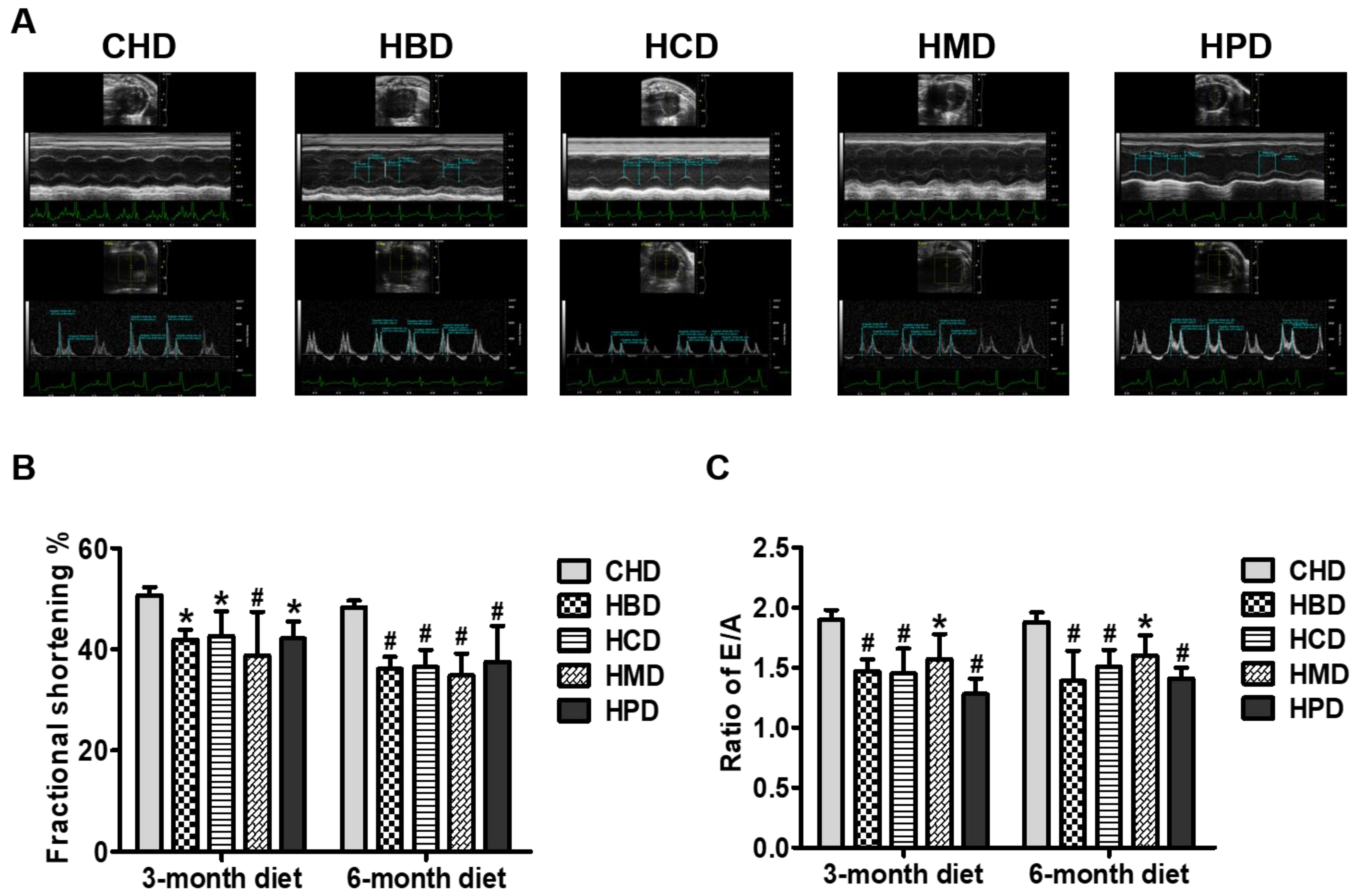
3.3. Cardiac Function in Mice Fed High SFA Diets
3.4. Fatty Acid Profile of Serum from Mice Fed High-Fat Diets
3.5. Fatty Acid Profile of Neutral Lipids and Phospholipids in Heart Tissues from Mice Fed High SFA Diets
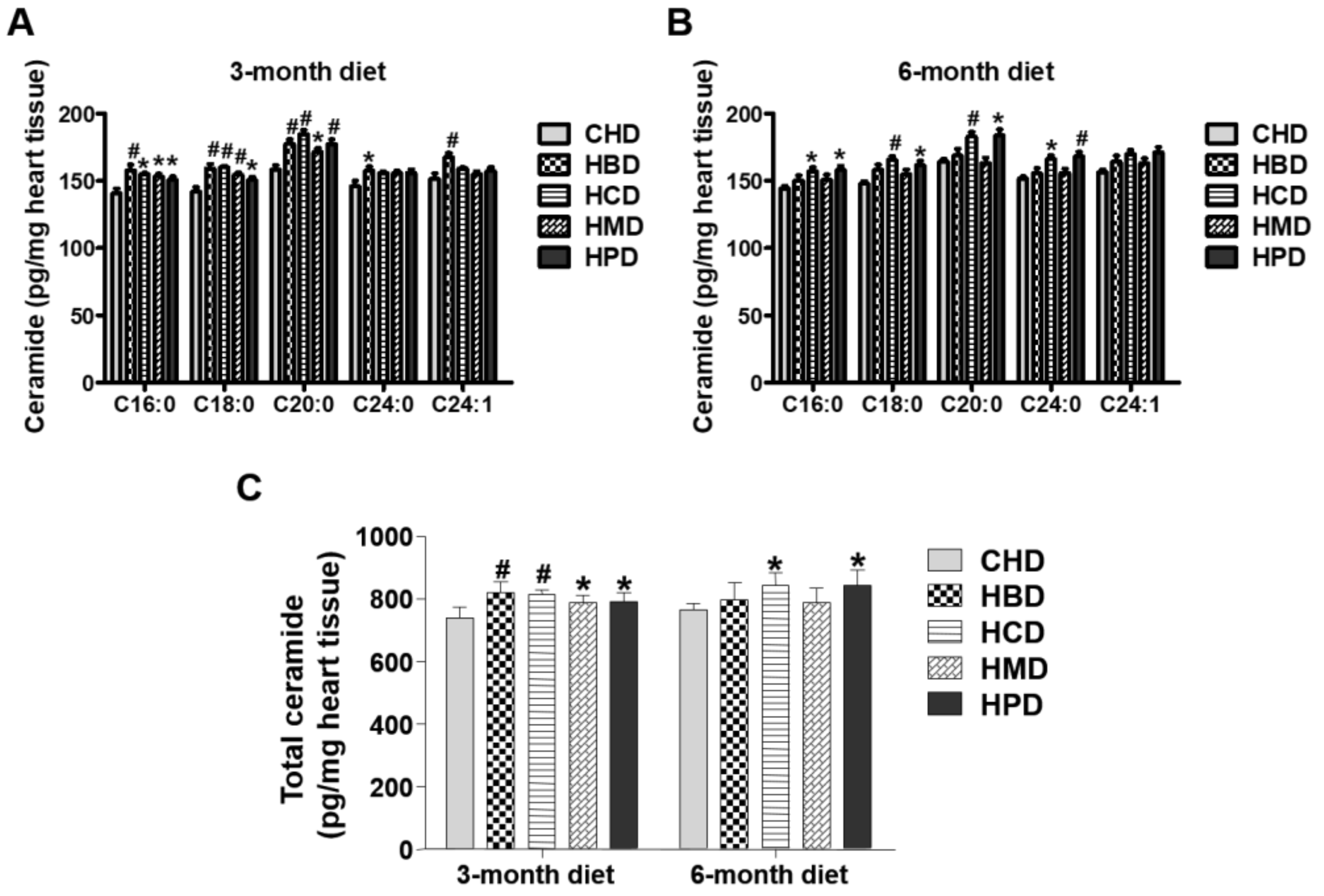
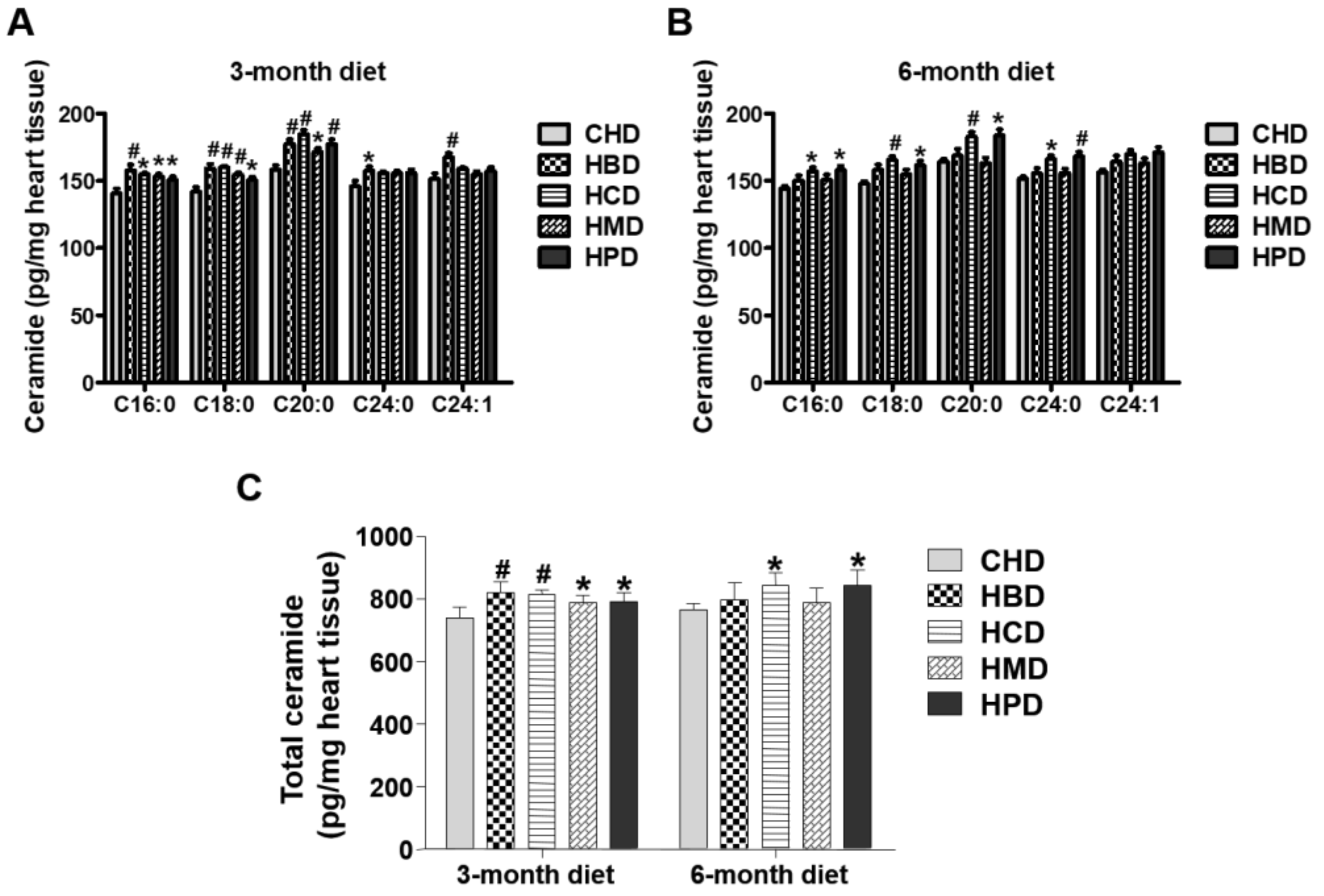
3.6. Ceramide Content in Heart Tissues from Mice Fed High SFA Diets
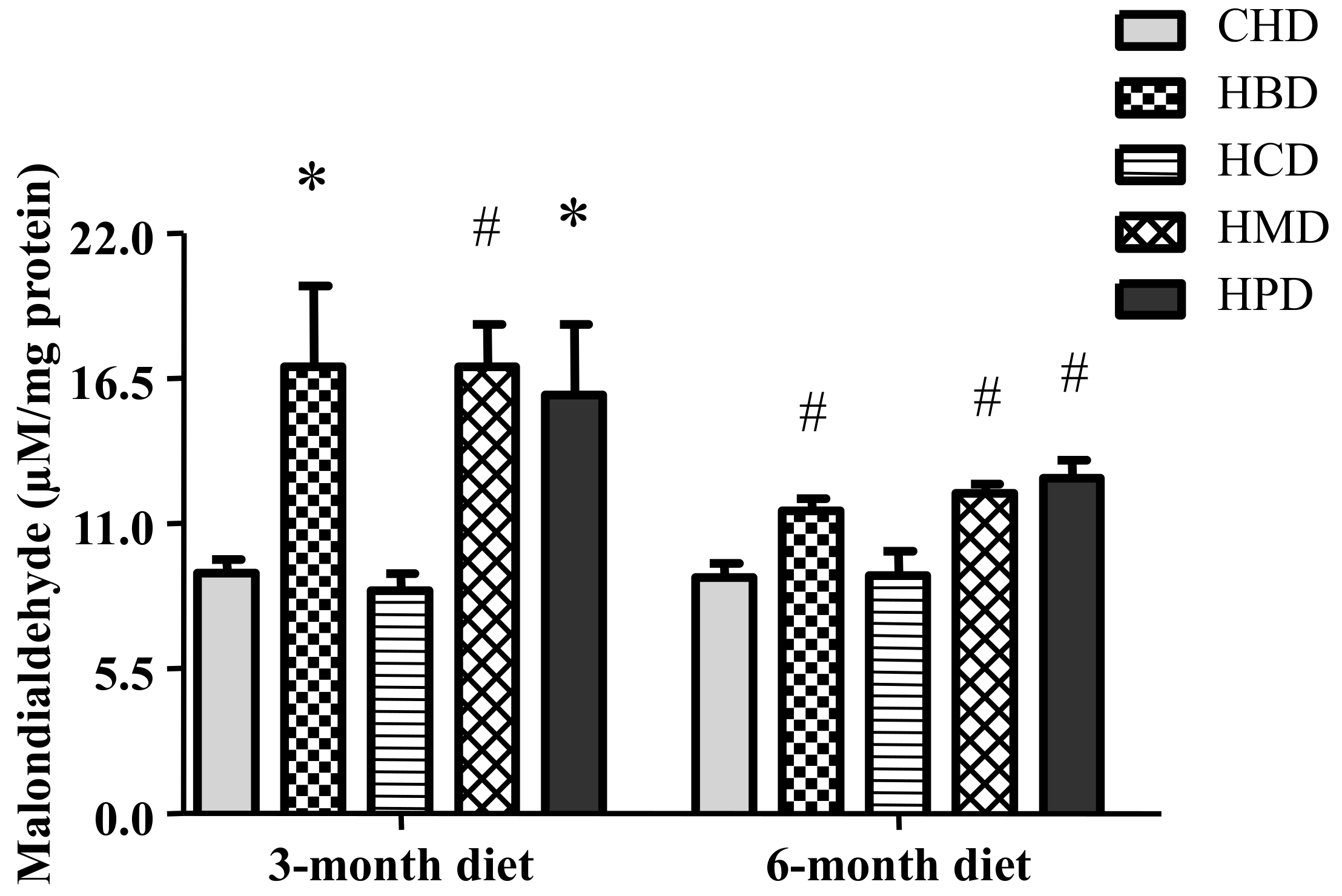
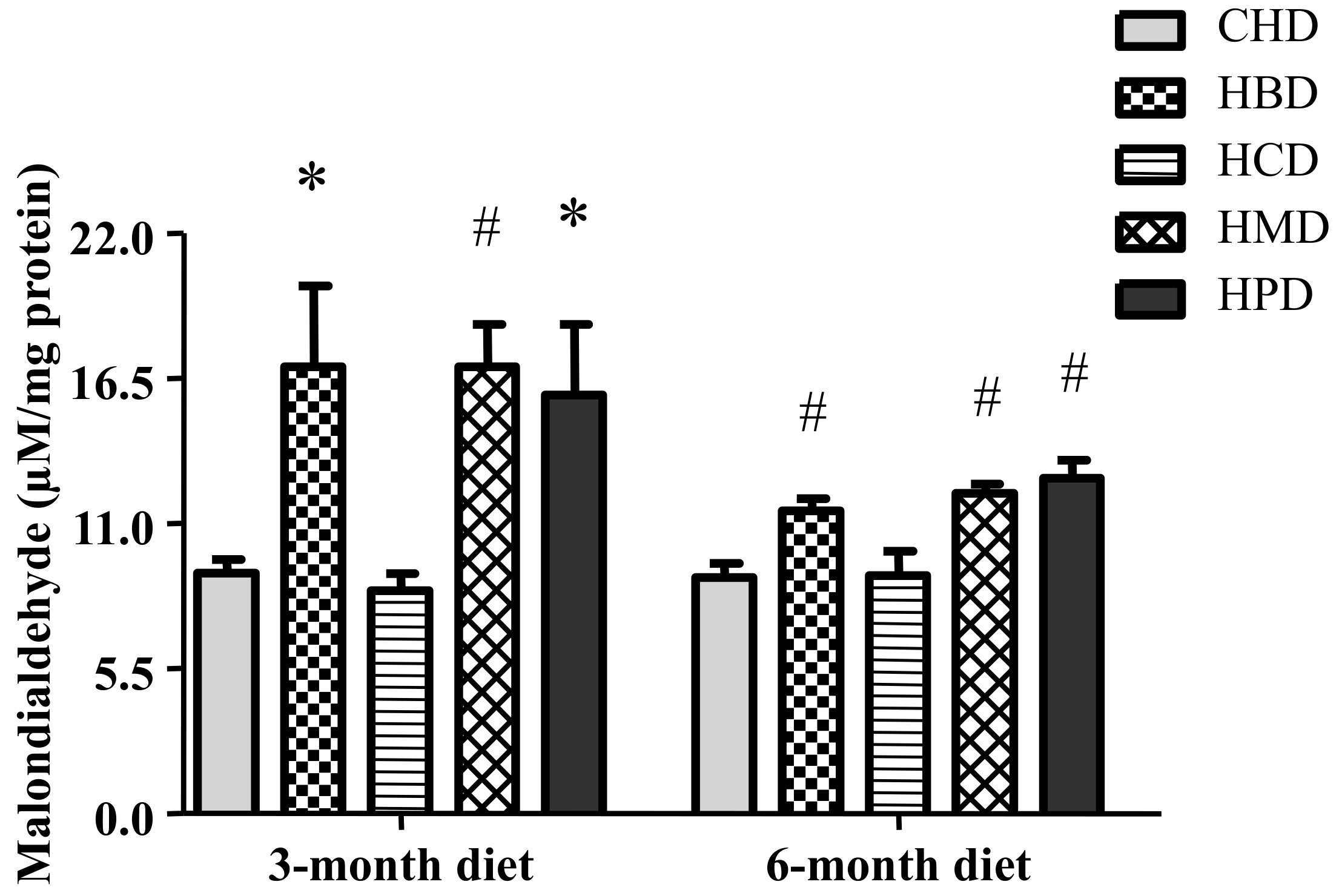
3.7. Malondialdehyde Level in Heart Tissues from Mice Fed High SFA Diets
4. Discussion
4.1. Design of Dietary Groups and Metabolic Profiles
4.2. SFA Intakesand Cardiac Abnormalities
4.3. SFA Intakes and Myocardialphospholipids Alteration
4.4. SFA Intakes, Myocardial Ceramides and Malondialdehyde
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Szendroedi, J.; Roden, M. Ectopic lipids and organ function. Curr. Opin. Lipidol. 2009, 20, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, J.E. Lipotoxicity: When tissues overeat. Curr. Opin. Lipidol. 2003, 14, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.T.; Grayburn, P.; Karim, A.; Shimabukuro, M.; Higa, M.; Baetens, D.; Orci, L.; Unger, R.H. Lipotoxic heart disease in obese rats: Implications for human obesity. Proc. Natl. Acad. Sci. USA 2000, 97, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Pepe, S.; McLennan, P.L. Cardiac membrane fatty acid composition modulates myocardial oxygen consumption and postischemic recovery of contractile function. Circulation 2002, 105, 2303–2308. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Wu, D.; Park, T.; Choi, C.S.; Li, R.K.; Cheng, K.K.; Xu, A.; Sweeney, G. APPL1 transgenic mice are protected from high-fat diet-induced cardiac dysfunction. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E795–E804. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.X.; Dong, F.; Thomas, D.P.; Ma, H.; He, L.; Ren, J. Hypertrophic cardiomyopathy in high-fat diet-induced obesity: Role of suppression of forkhead transcription factor and atrophy gene transcription. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1206–H1215. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Nunez, B.; Dijck-Brouwer, D.A.; Muskiet, F.A. The relation of saturated fatty acids with low-grade inflammation and cardiovascular disease. J. Nutr. Biochem. 2016, 36, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, J.; Andersson, D.C.; Zhang, S.J.; Lanner, J.T.; Wibom, R.; Katz, A.; Bruton, J.D.; Westerblad, H. Effects of palmitate on Ca(2+) handling in adult control and ob/ob cardiomyocytes: Impact of mitochondrial reactive oxygen species. Diabetes 2007, 56, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Zahabi, A.; Deschepper, C.F. Long-chain fatty acids modify hypertrophic responses of cultured primary neonatal cardiomyocytes. J. Lipid Res. 2001, 42, 1325–1330. [Google Scholar] [PubMed]
- Iggman, D.; Riserus, U. Role of different dietary saturated fatty acids for cardiometabolic risk. Clin. Lipidol. 2011, 6, 209–223. [Google Scholar] [CrossRef]
- Knowles, C.J.; Dionne, M.; Cebova, M.; Pinz, I.M. Palmitate-Induced Translocation of Caveolin-3 and Endothelial Nitric Oxide Synthase in Cardiomyocytes. Online J. Biol. Sci. 2011, 11, 27–36. [Google Scholar] [PubMed]
- Knowles, C.J.; Cebova, M.; Pinz, I.M. Palmitate diet-induced loss of cardiac caveolin-3: A novel mechanism for lipid-induced contractile dysfunction. PLoS ONE 2013, 8, e61369. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.B.; Baicu, C.F.; Van Laer, A.; Geng, T.; Kasiganesan, H.; Zile, M.R.; Cowart, L.A. Ceramide synthase 5 mediates lipid-induced autophagy and hypertrophy in cardiomyocytes. J. Clin. Investig. 2012, 122, 3919–3930. [Google Scholar] [CrossRef] [PubMed]
- Athenstaedt, K.; Daum, G. The life cycle of neutral lipids: Synthesis, storage and degradation. Cell. Mol. Life Sci. 2006, 63, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
- Ruhl, R.; Koch, C.; Marosvolgyi, T.; Mihaly, J.; Schweigert, F.J.; Worm, M.; Decsi, T. Fatty acid composition of serum lipid classes in mice following allergic sensitisation with or without dietary docosahexaenoic acid-enriched fish oil substitution. Br. J. Nutr. 2008, 99, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Gerson, A.R.; Brown, J.C.; Thomas, R.; Bernards, M.A.; Staples, J.F. Effects of dietary polyunsaturated fatty acids on mitochondrial metabolism in mammalian hibernation. J. Exp. Biol. 2008, 211, 2689–2699. [Google Scholar] [CrossRef] [PubMed]
- Thiebaut, A.C.; Rotival, M.; Gauthier, E.; Lenoir, G.M.; Boutron-Ruault, M.C.; Joulin, V.; Clavel-Chapelon, F.; Chajes, V. Correlation between serum phospholipid fatty acids and dietary intakes assessed a few years earlier. Nutr. Cancer 2009, 61, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Zelman, K. The great fat debate: A closer look at the controversy-questioning the validity of age-old dietary guidance. J. Am. Diet. Assoc. 2011, 111, 655–658. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Otto, M.C.; Mozaffarian, D.; Kromhout, D.; Bertoni, A.G.; Sibley, C.T.; Jacobs, D.R., Jr.; Nettleton, J.A. Dietary intake of saturated fat by food source and incident cardiovascular disease: The Multi-Ethnic Study of Atherosclerosis. Am. J. Clin. Nutr. 2012, 96, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Micha, R.; Mozaffarian, D. Saturated fat and cardiometabolic risk factors, coronary heart disease, stroke, and diabetes: A fresh look at the evidence. Lipids 2010, 45, 893–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartvigsen, K.; Binder, C.J.; Hansen, L.F.; Rafia, A.; Juliano, J.; Horkko, S.; Steinberg, D.; Palinski, W.; Witztum, J.L.; Li, A.C. A diet-induced hypercholesterolemic murine model to study atherogenesis without obesity and metabolic syndrome. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate High-fat Diet–induced Metabolic Syndrome and Cardiovascular Remodeling in Rats. J. Cardiovasc. Pharmacol. 2011, 57, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Ceylan-Isik, A.F.; Kandadi, M.R.; Xu, X.; Hua, Y.; Chicco, A.J.; Ren, J.; Nair, S. Apelin administration ameliorates high fat diet-induced cardiac hypertrophy and contractile dysfunction. J. Mol. Cell. Cardiol. 2013, 63, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Ibarguren, M.; Lopez, D.J.; Escriba, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta 2014, 1838, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Tepsic, V.; Ristic, V.; Ristic, D.; Vasiljevic, N.; Pecelj-Gec, M. Heart phospholipid content and fatty acid composition in the rat after feeding different lipid supplemented diets. Physiol. Res. 1998, 47, 413–418. [Google Scholar] [PubMed]
- O’Connell, K.A.; Dabkowski, E.R.; de Fatima Galvao, T.; Xu, W.; Daneault, C.; de Rosiers, C.; Stanley, W.C. Dietary saturated fat and docosahexaenoic acid differentially effect cardiac mitochondrial phospholipid fatty acyl composition and Ca(2+) uptake, without altering permeability transition or left ventricular function. Physiol. Rep. 2013, 1, e00009. [Google Scholar] [PubMed]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Giugliano, D. Diet and inflammation: A link to metabolic and cardiovascular diseases. Eur. Heart J. 2006, 27, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Zhang, Y.; Dolence, J.; Shi, G.P.; Ren, J.; Nair, S. Cathepsin K knockout mitigates high-fat diet-induced cardiac hypertrophy and contractile dysfunction. Diabetes 2013, 62, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Fucho, R.; Casals, N.; Serra, D.; Herrero, L. Ceramides and mitochondrial fatty acid oxidation in obesity. FASEB J. 2017, 31, 1263–1272. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHD | HBD | HCD | HMD | HPD | |
|---|---|---|---|---|---|
| 3-month diet | |||||
| 14:0 | 0.09 ± 0.02 | 0.10 ± 0.01 | 0.03 ± 0.01 # | 0.15 ± 0.01 # | 0.05 ± 0.01 # |
| 16:0 | 4.27 ± 0.50 | 3.33 ± 0.40 * | 3.13 ± 0.23 * | 3.40 ± 0.37 * | 5.13 ± 0.87 * |
| 16:1 (n-9) | 0.81 ± 0.16 | 0.70 ± 0.06 | 0.39 ± 0.06 # | 0.64 ± 0.17 | 0.57 ± 0.11 * |
| 18:0 | 1.17 ± 0.13 | 1.42 ± 0.29 | 1.58 ± 0.28 | 1.02 ± 0.15 | 1.46 ± 0.30 |
| 18:1 (n-9) | 2.80 ± 0.40 | 3.60 ± 0.13 * | 2.91 ± 0.24 | 2.69 ± 0.16 | 3.91 ± 0.74 # |
| 18:2 (n-6) | 2.00 ± 0.38 | 2.31 ± 0.25 | 1.64 ± 0.26 | 2.34 ± 0.32 | 1.78 ± 0.53 |
| 20:4 (n-6) | 3.84 ± 0.40 | 3.23 ± 0.94 | 3.55 ± 0.68 | 2.62 ± 0.78 | 5.19 ± 0.91 * |
| 22:0 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.02 * |
| 22:1 | 0.06 ± 0.01 | 0.09 ± 0.05 | 0.08 ± 0.04 | 0.07 ± 0.03 | 0.12 ± 0.06 |
| 22:6 (n-3) | 0.44 ± 0.06 | 0.45 ± 0.09 | 0.45 ± 0.08 | 0.44 ± 0.15 | 0.58 ± 0.07 |
| Sum of SFA | 5.57 ± 0.57 | 4.91 ± 0.67 | 4.80 ± 0.49 | 4.63 ± 0.53 | 6.72 ± 1.18 * |
| Sum of MUFA | 3.67 ± 0.53 | 4.39 ± 0.18 * | 3.38 ± 0.28 | 3.39 ± 0.29 | 4.60 ± 0.18 * |
| Sum of PUFA | 6.27 ± 0.64 | 6.04 ± 1.18 | 5.65 ± 0.97 | 5.40 ± 1.14 | 7.55 ± 1.33 |
| 6-month diet | |||||
| 14:0 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.03 ± 0.01 # | 0.08 ± 0.02 | 0.04 ± 0.01 # |
| 16:0 | 3.16 ± 0.38 | 3.94 ± 0.29 | 3.41 ± 0.25 | 3.55 ± 0.47 | 4.85 ± 0.70 # |
| 16:1 (n-9) | 0.61 ± 0.12 | 0.56 ± 0.06 | 0.34 ± 0.04 # | 0.47 ± 0.07 | 0.48 ± 0.08 |
| 18:0 | 0.95 ± 0.14 | 1.34 ± 0.04 # | 1.75 ± 0.20 # | 1.03 ± 0.18 | 1.35 ± 0.12 # |
| 18:1 (n-9) | 2.54 ± 0.45 | 3.75 ± 0.34 * | 2.76 ± 0.31 | 2.58 ± 0.26 | 3.64 ± 0.69 * |
| 18:2 (n-6) | 1.37 ± 0.28 | 1.78 ± 0.16 | 1.68 ± 0.29 | 1.96 ± 0.32 | 1.59 ± 0.30 |
| 20:4 (n-6) | 3.51 ± 0.75 | 4.73 ± 0.20 * | 4.52 ± 0.39 * | 2.94 ± 0.57 | 5.34 ± 0.73 # |
| 22:0 | 0.05 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.02 | 0.08 ± 0.02 | 0.06 ± 0.01 |
| 22:1 | 0.12 ± 0.05 | 0.12 ± 0.04 | 0.07 ± 0.03 | 0.09 ± 0.04 | 0.08 ± 0.02 |
| 22:6 (n-3) | 0.25 ± 0.06 | 0.55 ± 0.03 # | 0.42 ± 0.03 # | 0.41 ± 0.08 # | 0.53 ± 0.11 # |
| Sum of SFA | 4.23 ± 0.51 | 5.39 ± 0.32 | 5.25 ± 0.44 | 4.73 ± 0.66 | 6.30 ± 0.81 # |
| Sum of MUFA | 3.27 ± 0.54 | 4.43 ± 0.27 * | 3.17 ± 0.36 | 3.14 ± 0.30 | 4.19 ± 0.77 * |
| Sum of PUFA | 5.12 ± 1.06 | 7.06 ± 0.16 * | 6.62 ± 0.61 * | 5.31 ± 0.94 | 7.46 ± 0.95 # |
| CHD | HBD | HCD | HMD | HPD | |
|---|---|---|---|---|---|
| 3-month diet | |||||
| 14:0 | 0.22 ± 0.05 | 0.27 ± 0.02 | 0.15 ± 0.02 | 0.38 ± 0.13 # | 0.15 ± 0.04 |
| 16:0 | 5.21 ± 0.82 | 5.11 ± 1.00 | 3.93 ± 0.63 | 5.37 ± 1.42 | 4.36 ± 1.59 |
| 16:1 (n-9) | 0.94 ± 0.18 | 0.87 ± 0.17 | 0.58 ± 0.15 | 0.89 ± 0.29 | 0.58 ± 0.13 |
| 18:0 | 2.40 ± 0.31 | 2.62 ± 0.46 | 2.34 ± 0.08 | 2.66 ± 0.45 | 2.09 ± 0.81 |
| 18:1 (n-9) | 17.67 ± 2.41 | 19.46 ± 2.02 | 17.59 ± 2.84 | 17.11 ± 4.41 | 15.81 ± 2.26 |
| 18:2 (n-6) | 2.40 ± 0.40 | 2.16 ± 0.27 | 1.63 ± 0.13 | 2.76 ± 1.21 | 1.42 ± 0.29 |
| 20:0 | 0.20 ± 0.01 | 0.19 ± 0.01 | 0.20 ± 0.01 | 0.24 ± 0.02 # | 0.20 ± 0.03 |
| 20:1 | 0.52 ± 0.12 | 0.36 ± 0.03 | 0.32 ± 0.06 | 0.50 ± 0.31 | 0.38 ± 0.06 |
| 20:4 (n-6) | 0.45 ± 0.06 | 0.39 ± 0.04 | 0.42 ± 0.07 | 0.47 ± 0.11 | 0.42 ± 0.05 |
| 22:0 | 0.16 ± 0.03 | 0.17 ± 0.02 | 0.17 ± 0.00 | 0.20 ± 0.02 # | 0.16 ± 0.01 |
| 22:1 | 5.47 ± 0.50 | 4.85 ± 0.32 | 4.59 ± 0.11 * | 4.90 ± 0.53 | 5.60 ± 0.25 |
| 22:5 | 0.39 ± 0.12 | 0.22 ± 0.08 | 0.24 ± 0.11 | 0.25 ± 0.14 | 0.36 ± 0.11 |
| 22:6 (n-3) | 0.51 ± 0.09 | 0.48 ± 0.10 | 0.52 ± 0.14 | 0.47 ± 0.32 | 0.44 ± 0.20 |
| Sum of SFA | 8.19 ± 1.14 | 8.36 ± 1.43 | 6.80 ± 0.66 | 9.40 ± 1.22 | 6.95 ± 2.42 |
| Sum of MUFA | 24.60 ± 2.88 | 25.54 ± 2.11 | 23.08 ± 3.00 | 23.41 ± 4.56 | 22.40 ± 2.59 |
| Sum of PUFA | 3.75 ± 0.55 | 3.24 ± 0.28 | 2.82 ± 0.40 | 3.96 ± 1.61 | 2.63 ± 0.43 |
| 6-month diet | |||||
| 14:0 | 0.19 ± 0.07 | 0.23 ± 0.08 | 0.21 ± 0.02 | 0.27 ± 0.05 | 0.19 ± 0.05 |
| 16:0 | 5.26 ± 2.11 | 4.94 ± 0.93 | 5.40 ± 0.34 | 5.83 ± 0.60 | 5.45 ± 1.34 |
| 16:1 (n-9) | 0.76 ± 0.46 | 0.77 ± 0.47 | 0.61 ± 0.09 | 0.74 ± 0.13 | 0.53 ± 0.23 |
| 18:0 | 2.48 ± 0.59 | 2.26 ± 0.94 | 2.83 ± 0.40 | 3.34 ± 0.61 * | 2.26 ± 0.57 |
| 18:1 (n-9) | 12.40 ± 3.86 | 12.08 ± 4.98 | 10.54 ± 1.90 | 12.89 ± 2.15 | 11.63 ± 2.40 |
| 18:2 (n-6) | 1.82 ± 0.75 | 1.24 ± 0.62 | 0.98 ± 0.26 | 1.29 ± 0.28 | 1.45 ± 0.78 |
| 20:0 | 0.07 ± 0.03 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.02 |
| 20:1 | 0.36 ± 0.06 | 0.31 ± 0.05 | 0.26 ± 0.04 | 0.27 ± 0.05 | 0.34 ± 0.11 |
| 20:4 (n-6) | 0.39 ± 0.13 | 0.39 ± 0.14 | 0.35 ± 0.07 | 0.34 ± 0.11 | 0.41 ± 0.06 |
| 22:0 | 0.16 ± 0.02 | 0.17 ± 0.03 | 0.18 ± 0.02 | 0.18 ± 0.03 | 0.18 ± 0.05 |
| 22:1 | 5.46 ± 0.41 | 5.04 ± 0.73 | 5.25 ± 0.67 | 5.84 ± 0.55 | 5.63 ± 0.77 |
| 22:5 | 0.36 ± 0.23 | 0.20 ± 0.10 | 0.17 ± 0.06 * | 0.19 ± 0.08 | 0.28 ± 0.12 |
| 22:6 (n-3) | 0.51 ± 0.18 | 0.32 ± 0.14 * | 0.28 ± 0.09 # | 0.34 ± 0.13 * | 0.49 ± 0.06 |
| Sum of SFA | 8.22 ± 2.77 | 8.08 ± 1.90 | 8.79 ± 0.71 | 9.80 ± 1.12 | 8.23 ± 1.84 |
| Sum of MUFA | 18.98 ± 4.39 | 18.20 ± 5.56 | 16.66 ± 2.40 | 19.73 ± 2.32 | 18.14 ± 2.80 |
| Sum of PUFA | 3.09 ± 1.17 | 2.16 ± 0.86 | 1.78 ± 0.29 * | 2.16 ± 0.31 | 2.63 ± 0.86 |
| CHD | HBD | HCD | HMD | HPD | |
|---|---|---|---|---|---|
| 3-month diet | |||||
| 14:0 | 0.02 ± 0.00 | 0.04 ± 0.01 # | 0.01 ± 0.00 | 0.05 ± 0.02 # | 0.01 ± 0.00 |
| 16:0 | 5.05 ± 0.15 | 5.35 ± 0.66 | 4.84 ± 0.17 | 6.01 ± 0.46 # | 5.81 ± 0.16 * |
| 16:1 (n-9) | 0.25 ± 0.02 | 0.28 ± 0.04 | 0.22 ± 0.04 | 0.30 ± 0.07 | 0.16 ± 0.04 * |
| 18:0 | 5.95 ± 0.30 | 9.08 ± 0.25 # | 10.29 ± 0.77 # | 9.82 ± 0.44 # | 6.53 ± 0.45 |
| 18:1 (n-9) | 4.69 ± 0.48 | 5.60 ± 0.36 # | 4.61 ± 0.35 | 4.57 ± 0.20 | 4.05 ± 0.35 |
| 18:2 (n-6) | 1.56 ± 0.35 | 1.68 ± 0.18 | 1.60 ± 0.34 | 2.01 ± 0.36 | 0.89 ± 0.13 # |
| 20:0 | 0.18 ± 0.02 | 0.12 ± 0.02 | 0.20 ± 0.02 | 0.18 ± 0.03 | 0.29 ± 0.25 |
| 20:1 | 0.27 ± 0.03 | 0.12 ± 0.02 # | 0.09 ± 0.01 # | 0.10 ± 0.04 # | 0.14 ± 0.10 # |
| 20:4 (n-6) | 5.19 ± 0.29 | 6.93 ± 0.87 # | 8.17 ± 0.68 # | 7.09 ± 0.64 # | 5.62 ± 0.68 |
| 22:0 | 0.12 ± 0.01 | 0.10 ± 0.02 | 0.14 ± 0.02 * | 0.16 ± 0.01 # | 0.12 ± 0.01 |
| 22:1 | 0.09 ± 0.03 | 0.09 ± 0.08 | 0.08 ± 0.03 | 0.07 ± 0.02 | 0.14 ± 0.06 |
| 22:5 | 0.53 ± 0.06 | 0.65 ± 0.13 | 0.60 ± 0.08 | 1.24 ± 0.10 # | 0.79 ± 0.21 * |
| 22:6 (n-3) | 7.81 ± 0.20 | 8.79 ± 0.83 | 8.03 ± 0.70 | 9.93 ± 0.65 # | 7.68 ± 0.71 |
| Sum of SFA | 11.32 ± 0.28 | 14.69 ± 0.89 # | 15.49 ± 0.74 # | 16.22 ± 0.59 # | 12.75 ± 0.73 # |
| Sum of MUFA | 5.31 ± 0.50 | 6.09 ± 0.40 # | 4.99 ± 0.35 | 5.03 ± 0.26 | 4.49 ± 0.24 * |
| Sum of PUFA | 15.09 ± 0.46 | 18.05 ± 1.62 # | 18.40 ± 1.34 # | 20.27 ± 0.85 # | 14.99 ± 0.95 |
| 6-month diet | |||||
| 14:0 | 0.02 ± 0.00 | 0.02 ± 0.01 | 0.02 ± 0.00 | 0.02 ± 0.01 | 0.01 ± 0.01 |
| 16:0 | 4.89 ± 0.76 | 4.55 ± 0.18 | 4.67 ± 0.46 | 4.85 ± 0.33 | 5.72 ± 0.64 # |
| 16:1 (n-9) | 0.28 ± 0.14 | 0.19 ± 0.02 | 0.19 ± 0.04 | 0.18 ± 0.02 | 0.17 ± 0.06 * |
| 18:0 | 5.37 ± 0.67 | 7.76 ± 0.36 # | 8.43 ± 0.74 # | 7.26 ± 0.60 # | 6.68 ± 0.51 # |
| 18:1 (n-9) | 4.00 ± 0.65 | 4.32 ± 0.35 | 3.55 ± 0.37 | 3.22 ± 0.25 * | 4.04 ± 0.50 |
| 18:2 (n-6) | 1.01 ± 0.17 | 0.81 ± 0.19 | 0.68 ± 0.13 * | 0.84 ± 0.27 | 0.87 ± 0.24 |
| 20:0 | 0.15 ± 0.03 | 0.12 ± 0.02 * | 0.18 ± 0.02 | 0.12 ± 0.02 * | 0.17 ± 0.01 |
| 20:1 | 0.16 ± 0.05 | 0.09 ± 0.03 # | 0.07 ± 0.01 # | 0.06 ± 0.01 # | 0.11 ± 0.05 # |
| 20:4 (n-6) | 4.48 ± 0.69 | 6.76 ± 0.79 # | 6.42 ± 0.93 # | 5.56 ± 0.87 * | 6.11 ± 0.72 # |
| 22:0 | 0.10 ± 0.02 | 0.10 ± 0.01 | 0.12 ± 0.02 * | 0.10 ± 0.01 | 0.11 ± 0.02 |
| 22:1 | 0.10 ± 0.03 | 0.10 ± 0.03 | 0.13 ± 0.05 | 0.08 ± 0.01 | 0.12 ± 0.04 |
| 22:5 | 0.61 ± 0.14 | 0.80 ± 0.09 * | 0.64 ± 0.12 | 0.86 ± 0.17 # | 0.67 ± 0.06 |
| 22:6 (n-3) | 7.96 ± 1.95 | 7.85 ± 0.85 | 8.74 ± 1.11 | 8.12 ± 0.70 | 8.34 ± 1.79 |
| Sum of SFA | 10.52 ± 1.37 | 12.55 ± 0.37 # | 13.41 ± 1.19 # | 12.36 ± 0.80 # | 12.70 ± 1.00 * |
| Sum of MUFA | 4.53 ± 0.81 | 4.70 ± 0.36 | 3.95 ± 0.38 | 3.54 ± 0.26 * | 4.43 ± 0.55 |
| Sum of PUFA | 14.07 ± 2.49 | 16.21 ± 0.51 | 16.48 ± 1.93 | 15.38 ± 1.12 | 16.00 ± 1.44 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Huang, Y.; Zheng, D.; Ni, R.; Bernards, M.A. Dietary Fatty Acids Alter Lipid Profiles and Induce Myocardial Dysfunction without Causing Metabolic Disorders in Mice. Nutrients 2018, 10, 106. https://doi.org/10.3390/nu10010106
Chen B, Huang Y, Zheng D, Ni R, Bernards MA. Dietary Fatty Acids Alter Lipid Profiles and Induce Myocardial Dysfunction without Causing Metabolic Disorders in Mice. Nutrients. 2018; 10(1):106. https://doi.org/10.3390/nu10010106
Chicago/Turabian StyleChen, Bainian, Yifan Huang, Dong Zheng, Rui Ni, and Mark A. Bernards. 2018. "Dietary Fatty Acids Alter Lipid Profiles and Induce Myocardial Dysfunction without Causing Metabolic Disorders in Mice" Nutrients 10, no. 1: 106. https://doi.org/10.3390/nu10010106
APA StyleChen, B., Huang, Y., Zheng, D., Ni, R., & Bernards, M. A. (2018). Dietary Fatty Acids Alter Lipid Profiles and Induce Myocardial Dysfunction without Causing Metabolic Disorders in Mice. Nutrients, 10(1), 106. https://doi.org/10.3390/nu10010106