Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Neurite Extension
2.3. Hypoxia Challenge
2.4. Glutamate Challenge
2.5. Diets
2.6. Open Field Test
2.7. Barnes Maze
2.8. Elevated Plus Maze
2.9. Forced Swim Test
2.10. Weight Maintenance
2.11. Body Composition
2.12. Statistical Analysis
3. Results
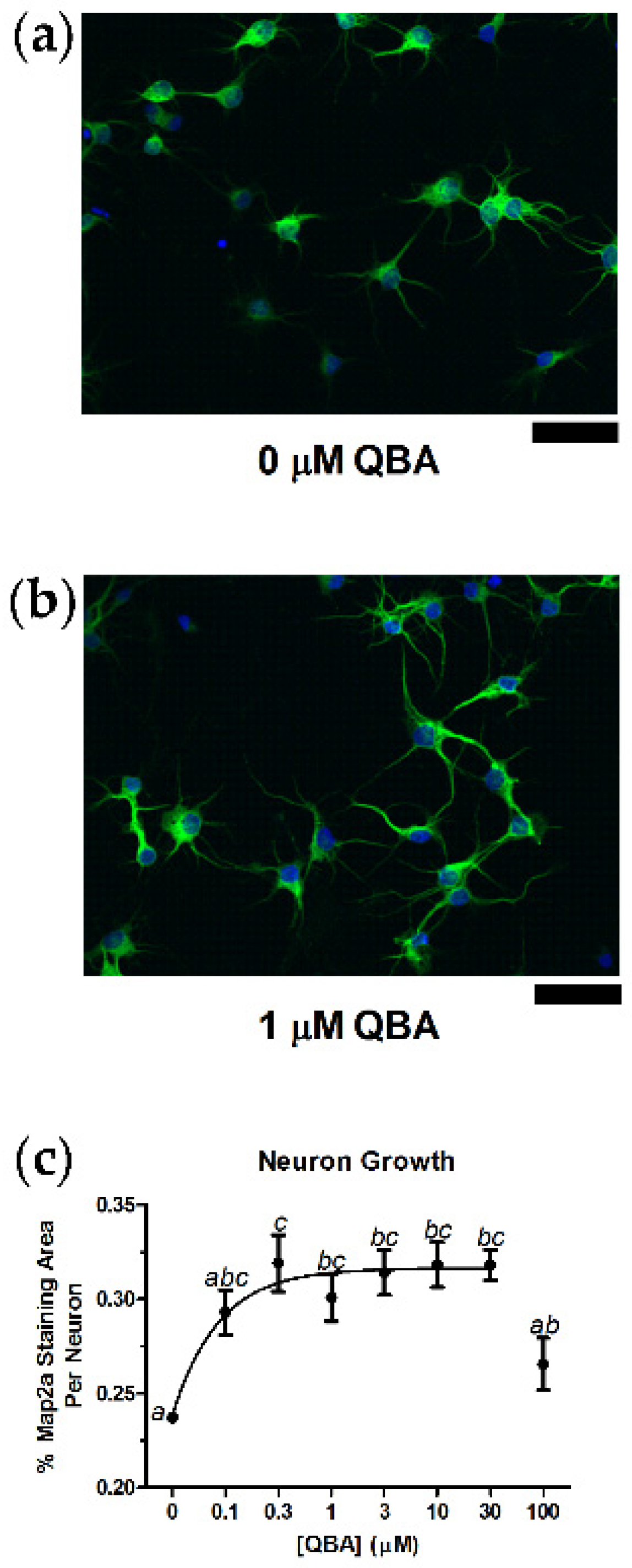
3.1. Neuron Growth
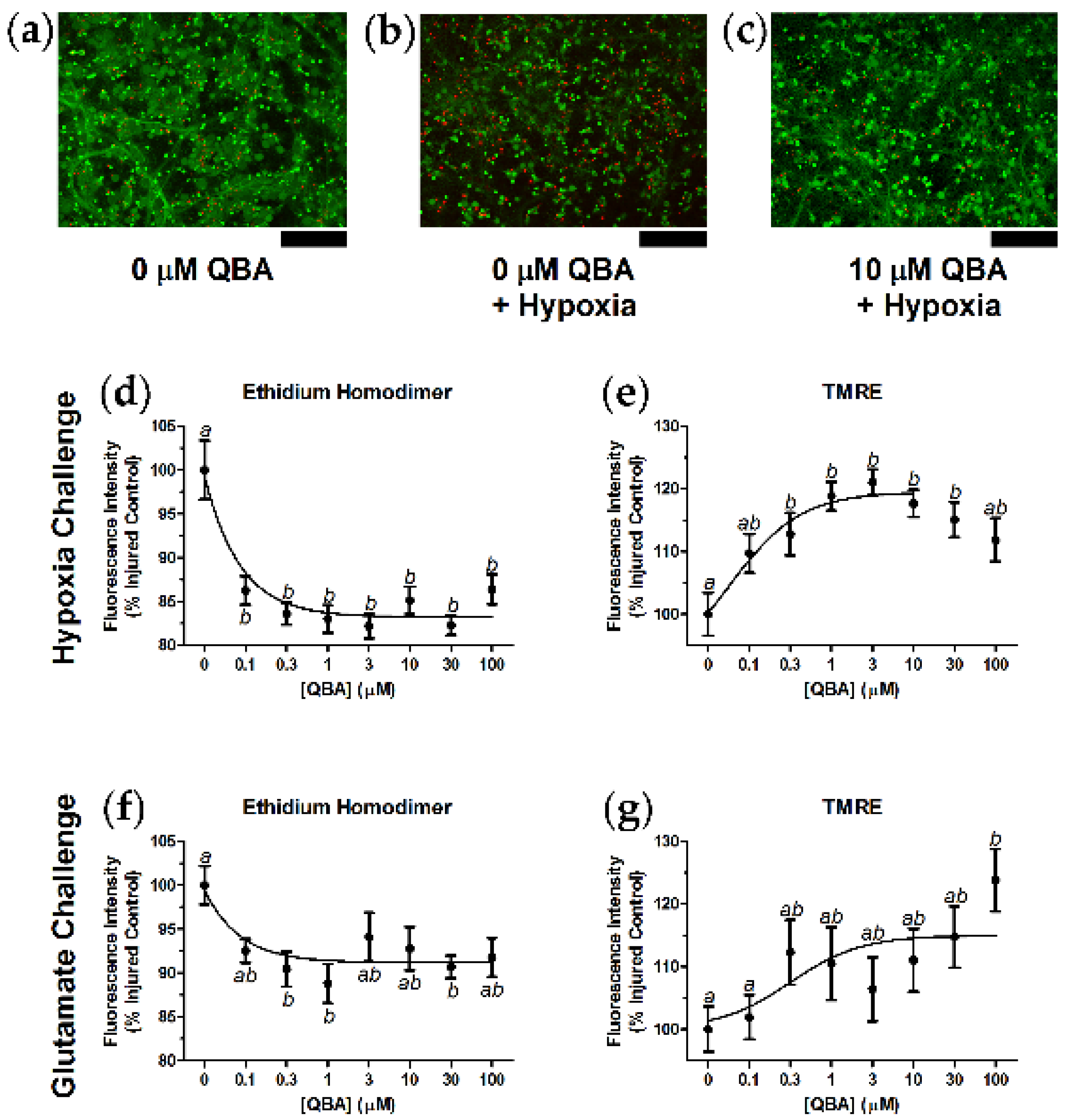
3.2. Hypoxia Challenge and Glutamate Challenge
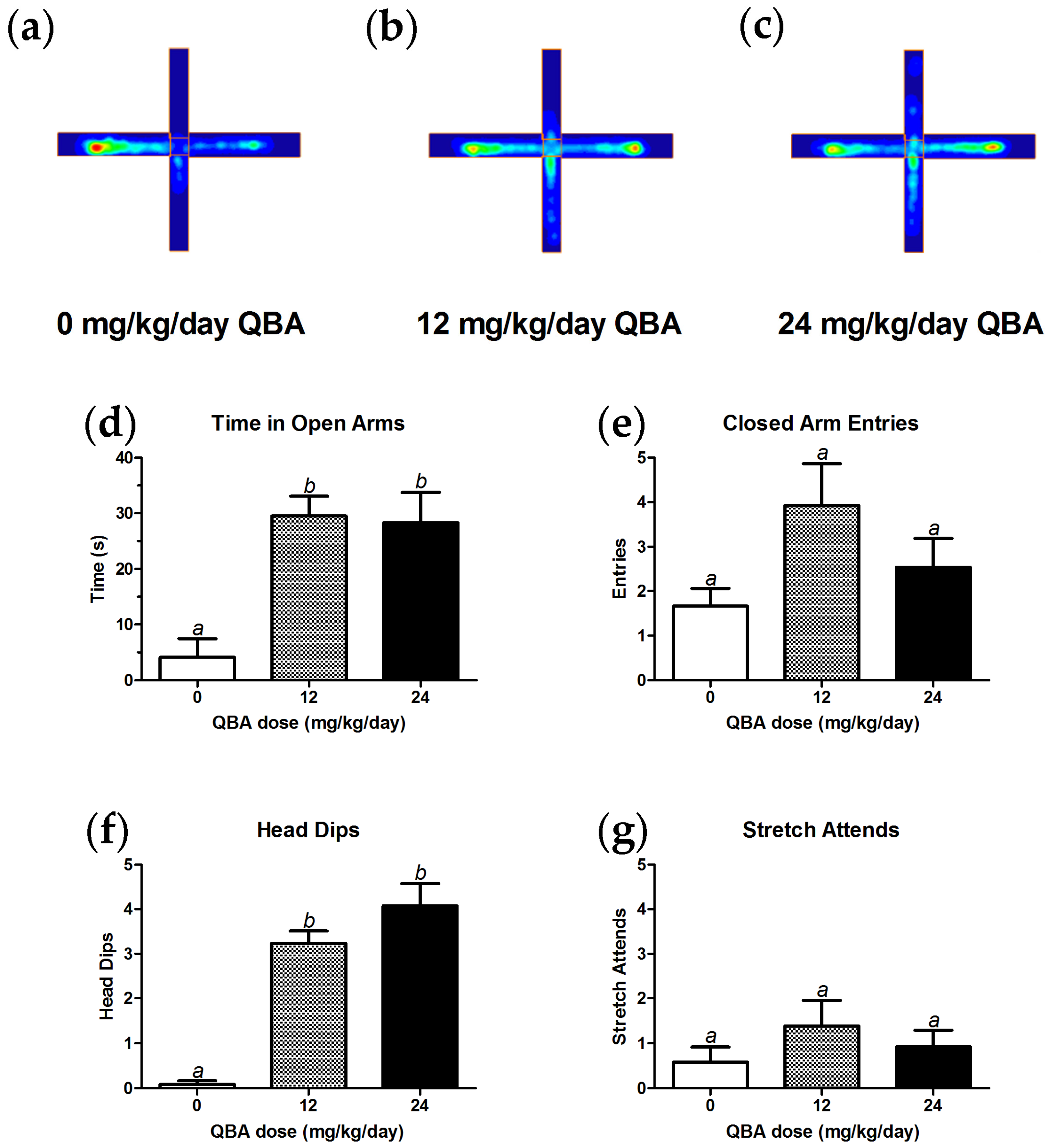
3.3. Behavior
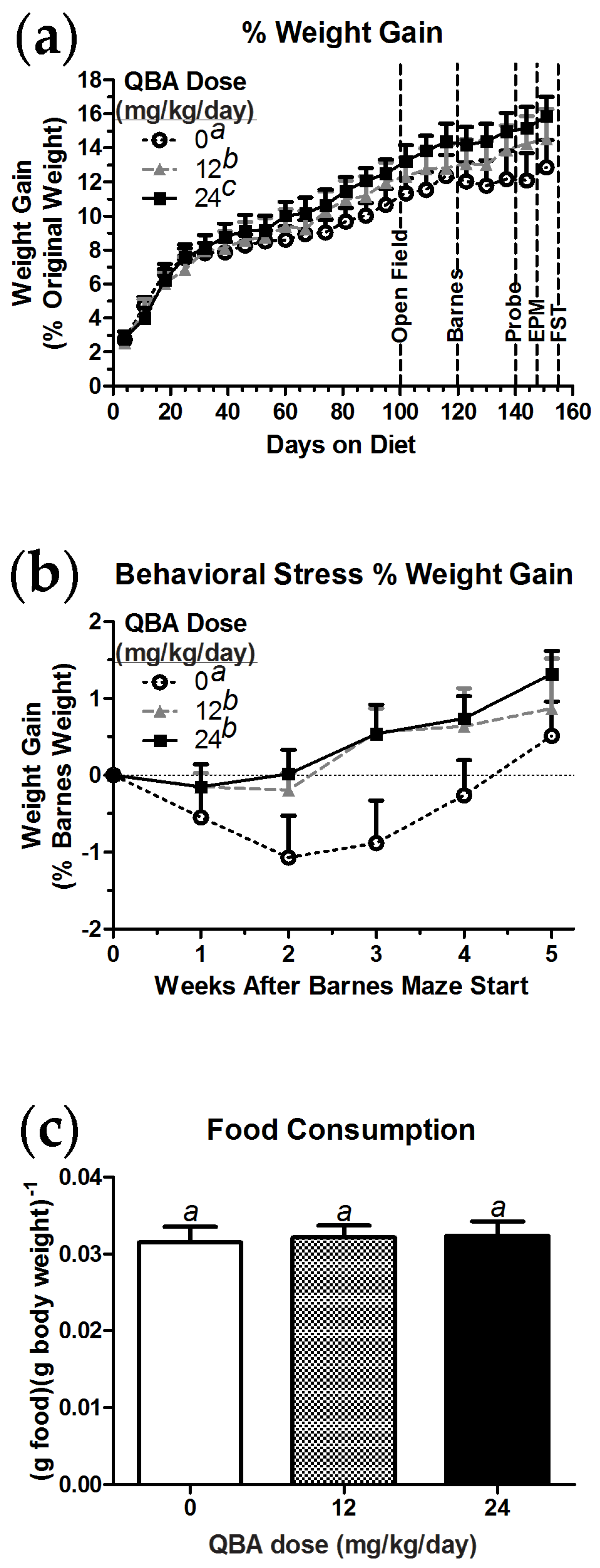
3.4. Weight Maintenance
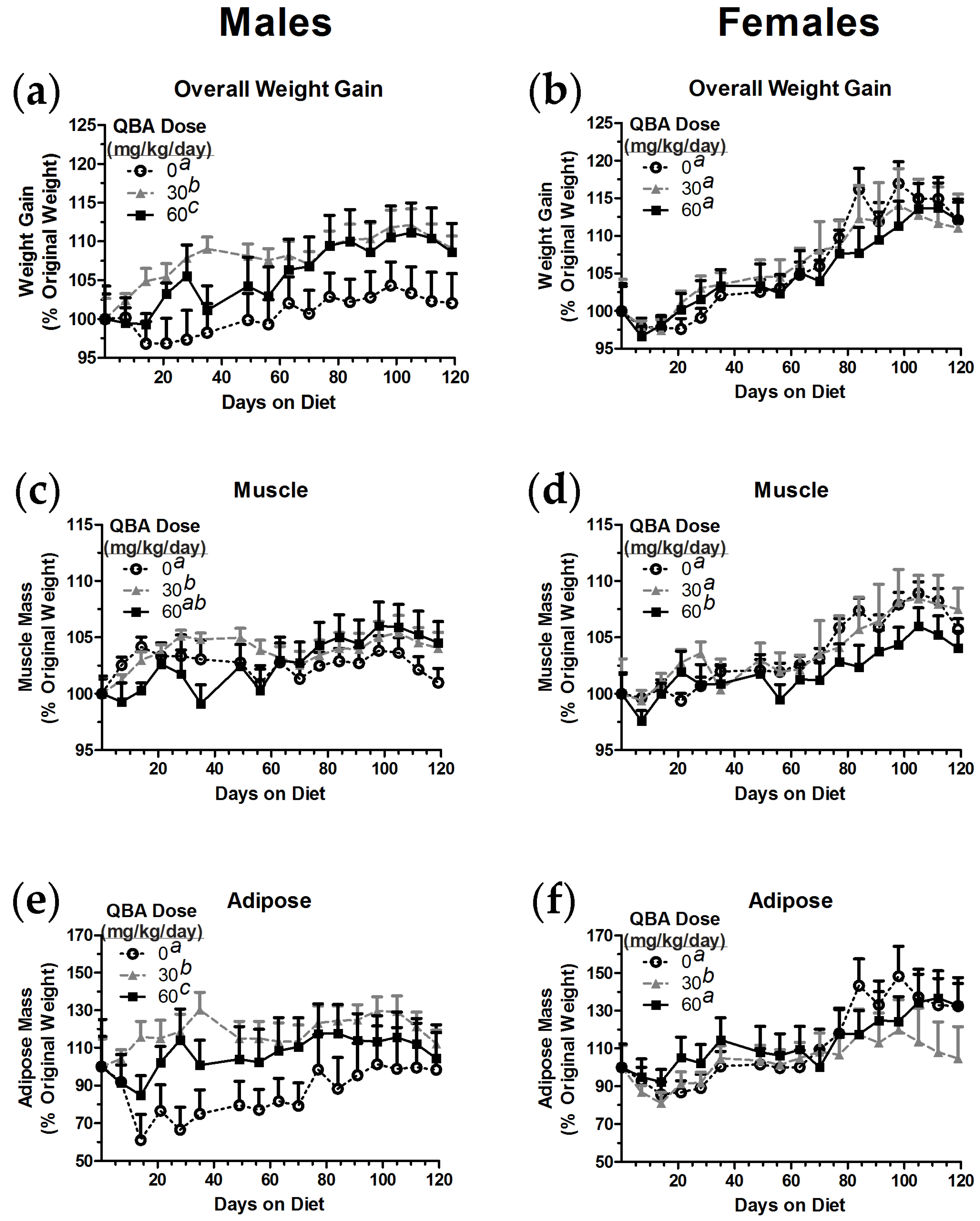
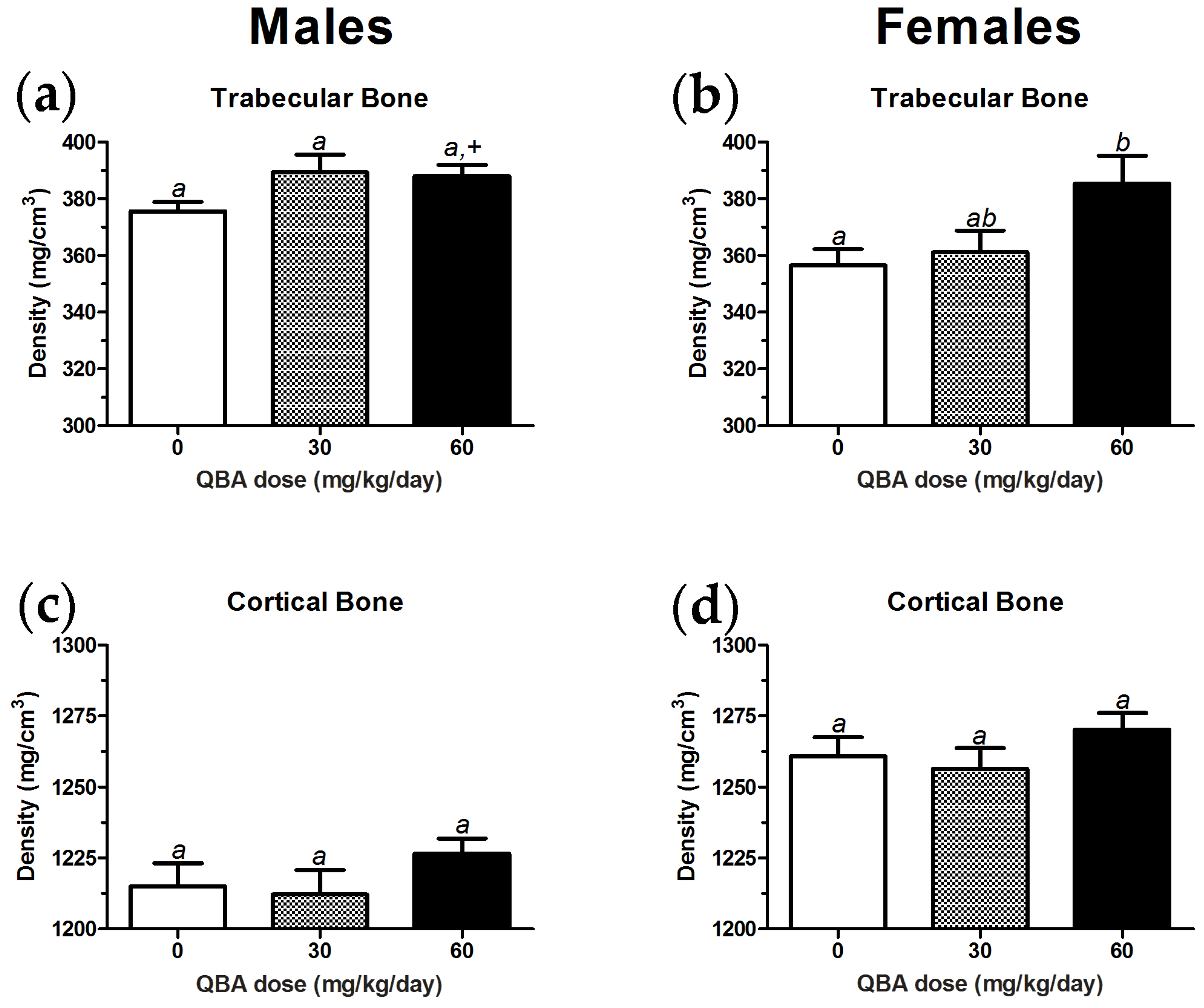
3.5. Body Composition in Mice
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Isidorov, V.A.; Bakier, S.; Grzech, I. Gas chromatographic-mass spectrometric investigation of volatile and extractable compounds of crude royal jelly. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 885–886, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Spannhoff, A.; Kim, Y.K.; Raynal, N.J.; Gharibyan, V.; Su, M.B.; Zhou, Y.Y.; Li, J.; Castellano, S.; Sbardella, G.; Issa, J.P.; et al. Histone deacetylase inhibitor activity in royal jelly might facilitate caste switching in bees. EMBO Rep. 2011, 12, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Zhou, M.; Zhang, L.; Xie, G.; Chen, H.; Liu, Z.; Ma, W. Influence of royal jelly on the reproductive function of puberty male rats. Food Chem. Toxicol. 2012, 50, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Heyl, H.L. An observation suggesting the presence of a gonadotropic hormone in royal jelly. Science 1939, 89, 540–541. [Google Scholar] [CrossRef] [PubMed]
- Morita, H.; Ikeda, T.; Kajita, K.; Fujioka, K.; Mori, I.; Okada, H.; Uno, Y.; Ishizuka, T. Effect of royal jelly ingestion for six months on healthy volunteers. Nutr. J. 2012, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Munstedt, K.; Henschel, M.; Hauenschild, A.; von Georgi, R. Royal jelly increases high density lipoprotein levels but in older patients only. J. Altern. Complement. Med. 2009, 15, 329–330. [Google Scholar] [PubMed]
- Guo, H.; Saiga, A.; Sato, M.; Miyazawa, I.; Shibata, M.; Takahata, Y.; Morimatsu, F. Royal jelly supplementation improves lipoprotein metabolism in humans. J. Nutr. Sci. Vitaminol. 2007, 53, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Elnagar, S.A. Royal jelly counteracts bucks’ “summer infertility”. Anim. Reprod. Sci. 2010, 121, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Kanda, M.; Ikeno, K.; Hayashi, Y.; Nakamura, T.; Ogawa, Y.; Fukumitsu, H.; Nomoto, H.; Furukawa, S. Oral administration of royal jelly facilitates mrna expression of glial cell line-derived neurotrophic factor and neurofilament h in the hippocampus of the adult mouse brain. Biosci. Biotechnol. Biochem. 2005, 69, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Ohta, S.; Sakamoto, T.; Mishima, S.; Furukawa, S. Royal jelly facilitates restoration of the cognitive ability in trimethyltin-intoxicated mice. Evid. Based Complement. Altern. Med. 2011, 2011, 165968. [Google Scholar] [CrossRef] [PubMed]
- Husein, M.Q.; Haddad, S.G. A new approach to enhance reproductive performance in sheep using royal jelly in comparison with equine chorionic gonadotropin. Anim. Reprod. Sci. 2006, 93, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Karaca, T.; Bayiroglu, F.; Yoruk, M.; Kaya, M.S.; Uslu, S.; Comba, B.; Mis, L. Effect of royal jelly on experimental colitis induced by acetic acid and alteration of mast cell distribution in the colon of rats. Eur. J. Histochem. 2010, 54, e35. [Google Scholar] [CrossRef] [PubMed]
- Mishima, S.; Suzuki, K.M.; Isohama, Y.; Kuratsu, N.; Araki, Y.; Inoue, M.; Miyata, T. Royal jelly has estrogenic effects in vitro and in vivo. J. Ethnopharmacol. 2005, 101, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Inoue, R.; Suzuki, N.; Nagashima, T. Antioxidant properties of enzymatic hydrolysates from royal jelly. J. Med. Food 2006, 9, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, M.; Onda, H.; Sasaki, K.; Yukiyoshi, A.; Tachibana, H.; Yamada, K. Effect of royal jelly on bisphenol a-induced proliferation of human breast cancer cells. Biosci. Biotechnol. Biochem. 2007, 71, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, Y.; Kohno, K.; Inoue, S.; Koya-Miyata, S.; Okamoto, I.; Arai, N.; Iwaki, K.; Ikeda, M.; Kurimoto, M. Oral administration of royal jelly inhibits the development of atopic dermatitis-like skin lesions in NC/NGA mice. Int. Immunopharmacol. 2003, 3, 1313–1324. [Google Scholar] [CrossRef]
- Barker, S.A.; Foster, A.B.; Lamb, D.C.; Hodgson, N. Identification of 10-hydroxy-delta 2-decenoic acid in royal jelly. Nature 1959, 183, 996–997. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Nomoto, H.; Fukumitsu, H.; Mishima, S.; Furukawa, S. Royal jelly and its unique fatty acid, 10-hydroxy-trans-2-decenoic acid, promote neurogenesis by neural stem/progenitor cells in vitro. Biomed. Res. 2007, 28, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Nitta, Y.; Fukumitsu, H.; Soumiya, H.; Ikeno, K.; Nakamura, T.; Furukawa, S. Antidepressant-like activity of 10-hydroxy-trans-2-decenoic acid, a unique unsaturated fatty acid of royal jelly, in stress-inducible depression-like mouse model. Evid. Based Complement. Altern. Med. 2012, 2012, 139140. [Google Scholar] [CrossRef] [PubMed]
- Makino, A.; Iinuma, M.; Fukumitsu, H.; Soumiya, H.; Furukawa, Y.; Furukawa, S. 2-decenoic acid ethyl ester possesses neurotrophin-like activities to facilitate intracellular signals and increase synapse-specific proteins in neurons cultured from embryonic rat brain. Biomed. Res. 2010, 31, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Moutsatsou, P.; Papoutsi, Z.; Kassi, E.; Heldring, N.; Zhao, C.; Tsiapara, A.; Melliou, E.; Chrousos, G.P.; Chinou, I.; Karshikoff, A.; et al. Fatty acids derived from royal jelly are modulators of estrogen receptor functions. PLoS ONE 2010, 5, e15594. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.M.; Isohama, Y.; Maruyama, H.; Yamada, Y.; Narita, Y.; Ohta, S.; Araki, Y.; Miyata, T.; Mishima, S. Estrogenic activities of fatty acids and a sterol isolated from royal jelly. Evid. Based Complement. Altern. Med. 2008, 5, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bjornstrom, L.; Sjoberg, M. Mechanisms of estrogen receptor signaling: Convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Banker, G.A.; Cowan, W.M. Rat hippocampal neurons in dispersed cell culture. Brain Res. 1977, 126, 397–425. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.R.; Evege, E.K.; Price, P.J. Optimized survival of hippocampal neurons in b27-supplemented neurobasal, a new serum-free medium combination. J. Neurosci. Res. 1993, 35, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Brewer, G.J. Isolation and culture of adult rat hippocampal neurons. J. Neurosci. Methods 1997, 71, 143–155. [Google Scholar] [CrossRef]
- Bunting, J.R. Influx and efflux kinetics of cationic dye binding to respiring mitochondria. Biophys. Chem. 1992, 42, 163–175. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Priemel, M.; Kohler, T.; Weusten, A.; Muller, R.; Amling, M.; Eckstein, F. Precision and accuracy of peripheral quantitative computed tomography (PQCT) in the mouse skeleton compared with histology and microcomputed tomography (MICROCT). J. Bone Miner. Res. 2003, 18, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Senn, S.M.; Kantor, S.; Poulton, I.J.; Morris, M.J.; Sims, N.A.; O’Brien, T.J.; Wark, J.D. Adverse effects of valproate on bone: Defining a model to investigate the pathophysiology. Epilepsia 2010, 51, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Pauly, J.R.; Grun, E.U.; Collins, A.C. Tolerance to nicotine following chronic treatment by injections: A potential role for corticosterone. Psychopharmacology 1992, 108, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Matsushita, H.; Ieno, D.; Matsuda, Y.; Horii, Y.; Ishii, A.; Takahashi, T.; Kanazawa, H.; Wakatsuki, A.; Suzuki, T. Improvement of neurological disorders in postmenopausal model rats by administration of royal jelly. Climacteric 2016, 19, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Weiser, M.J.; Wynalda, K.; Salem, N., Jr.; Butt, C.M. Dietary DHA during development affects depression-like behaviors and biomarkers that emerge after puberty in adolescent rats. J. Lipid Res. 2015, 56, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L.; Teicher, M.H. Serotonin laterality in amygdala predicts performance in the elevated plus maze in rats. Neuroreport 1999, 10, 3497–3500. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.S.; Comim, C.M.; Valvassori, S.S.; Reus, G.Z.; Barbosa, L.M.; Andreazza, A.C.; Stertz, L.; Fries, G.R.; Gavioli, E.C.; Kapczinski, F.; et al. Acute administration of ketamine induces antidepressant-like effects in the forced swimming test and increases bdnf levels in the rat hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Pyrzanowska, J.; Piechal, A.; Blecharz-Klin, K.; Joniec-Maciejak, I.; Graikou, K.; Chinou, I.; Widy-Tyszkiewicz, E. Long-term administration of greek royal jelly improves spatial memory and influences the concentration of brain neurotransmitters in naturally aged wistar male rats. J. Ethnopharmacol. 2014, 155, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Galal, A.A.; Elewa, Y.H. Comparative protective effects of royal jelly and cod liver oil against neurotoxic impact of tartrazine on male rat pups brain. Acta Histochem. 2015, 117, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Ohta, S.; Suzuki, K.M.; Nemoto, T.; Abe, K.; Mishima, S. Effects of long-term administration of royal jelly on pituitary weight and gene expression in middle-aged female rats. Biosci. Biotechnol. Biochem. 2009, 73, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Nomoto, H.; Fukumitsu, H.; Mishima, S.; Furukawa, S. Royal jelly-induced neurite outgrowth from rat pheochromocytoma PC12 cells requires integrin signal independent of activation of extracellular signal-regulated kinases. Biomed. Res. 2007, 28, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Hota, K.B.; Hota, S.K.; Chaurasia, O.P.; Singh, S.B. Acetyl-l-carnitine-mediated neuroprotection during hypoxia is attributed to ERK1/2-NRF2-regulated mitochondrial biosynthesis. Hippocampus 2012, 22, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Park, H.M.; Hwang, E.; Lee, K.G.; Han, S.M.; Cho, Y.; Kim, S.Y. Royal jelly protects against ultraviolet b-induced photoaging in human skin fibroblasts via enhancing collagen production. J. Med. Food 2011, 14, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Verdier-Sevrain, S. Effect of estrogens on skin aging and the potential role of selective estrogen receptor modulators. Climacteric 2007, 10, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, M.A.; Ruiz-Palmero, I.; Scerbo, M.J.; Acaz-Fonseca, E.; Cambiasso, M.J.; Garcia-Segura, L.M. Molecular mechanisms involved in the regulation of neuritogenesis by estradiol: Recent advances. J. Steroid Biochem. Mol. Biol. 2012, 131, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Engler-Chiurazzi, E.B.; Singh, M.; Simpkins, J.W. From the 90’s to now: A brief historical perspective on more than two decades of estrogen neuroprotection. Brain Res. 2016, 1633, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Benmansour, S.; Arroyo, L.D.; Frazer, A. Comparison of the antidepressant-like effects of estradiol and that of selective serotonin reuptake inhibitors in middle-aged ovariectomized rats. Front. Aging Neurosci. 2016, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Borbelyova, V.; Domonkos, E.; Babickova, J.; Tothova, L.; Kacmarova, M.; Ulicna, O.; Ostatnikova, D.; Hodosy, J.; Celec, P. Does long-term androgen deficiency lead to metabolic syndrome in middle-aged rats? Exp. Gerontol. 2017, 98, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Fedotova, J.; Hritcu, L. Testosterone promotes anxiolytic-like behavior in gonadectomized male rats via blockade of the 5-HT1A receptors. Gen. Comp. Endocrinol. 2017, 254, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Marcondes, F.K.; Miguel, K.J.; Melo, L.L.; Spadari-Bratfisch, R.C. Estrous cycle influences the response of female rats in the elevated plus-maze test. Physiol. Behav. 2001, 74, 435–440. [Google Scholar] [CrossRef]
- Butcher, R.L.; Collins, W.E.; Fugo, N.W. Plasma concentration of lh, fsh, prolactin, progesterone and estradiol-17beta throughout the 4-day estrous cycle of the rat. Endocrinology 1974, 94, 1704–1708. [Google Scholar] [CrossRef] [PubMed]
- McClure-Begley, T.D.; King, N.M.; Collins, A.C.; Stitzel, J.A.; Wehner, J.M.; Butt, C.M. Acetylcholine-stimulated [3H]GABA release from mouse brain synaptosomes is modulated by alpha4beta2 and alpha4alpha5beta2 nicotinic receptor subtypes. Mol. Pharmacol. 2009, 75, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zheng, S.; Li, X.F.; Liu, Z.D. The effect of estradiol on the growth plate chondrocytes of limb and spine from postnatal mice in vitro: The role of estrogen-receptor and estradiol concentration. Int. J. Biol. Sci. 2017, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.S.; Setchell, K.D.; Stocco, D.M.; Lephart, E.D. Dietary soy-phytoestrogens decrease testosterone levels and prostate weight without altering LH, prostate 5alpha-reductase or testicular steroidogenic acute regulatory peptide levels in adult male Sprague-Dawley rats. J. Endocrinol. 2001, 170, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, S.; Okamoto, Y.; Uchiyama, S.; Nakatsuma, A.; Hashimoto, K.; Ohnishi, S.T.; Yamaguchi, M. Royal jelly prevents osteoporosis in rats: Beneficial effects in ovariectomy model and in bone tissue culture model. Evid. Based Complement. Altern. Med. 2006, 3, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Nomura, J.; Ohta, S.; Inoh, Y.; Suzuki, K.M.; Araki, Y.; Okada, S.; Matsumoto, I.; Isohama, Y.; Abe, K.; et al. Royal jelly stimulates bone formation: Physiologic and nutrigenomic studies with mice and cell lines. Biosci. Biotechnol. Biochem. 2006, 70, 2508–2514. [Google Scholar] [CrossRef] [PubMed]
- Ozan, F.; Corekci, B.; Toptas, O.; Halicioglu, K.; Irgin, C.; Yilmaz, F.; Hezenci, Y. Effect of royal jelly on new bone formation in rapid maxillary expansion in rats. Med. Oral Patol. Oral Cir. Bucal 2015, 20, e651–e656. [Google Scholar] [CrossRef] [PubMed]
- Gielen, E.; O’Neill, T.W.; Pye, S.R.; Adams, J.E.; Wu, F.C.; Laurent, M.R.; Claessens, F.; Ward, K.A.; Boonen, S.; Bouillon, R.; et al. Endocrine determinants of incident sarcopenia in middle-aged and elderly european men. J. Cachexia Sarcopenia Muscle 2015, 6, 242–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minahan, C.; Joyce, S.; Bulmer, A.C.; Cronin, N.; Sabapathy, S. The influence of estradiol on muscle damage and leg strength after intense eccentric exercise. Eur. J. Appl. Physiol. 2015, 115, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Manson, J.E.; Stevenson, J.C.; Fonseca, V.A. Menopausal hormone therapy and type 2 diabetes prevention: Evidence, mechanisms, and clinical implications. Endocr. Rev. 2017, 38, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Lundsgaard, A.M.; Kiens, B. Gender differences in skeletal muscle substrate metabolism—Molecular mechanisms and insulin sensitivity. Front. Endocrinol. 2014, 5, 195. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Erickson, C.; Bessesen, D.; Van Pelt, R.E.; Cox-York, K. Age- and menopause-related differences in subcutaneous adipose tissue estrogen receptor mrna expression. Steroids 2017, 121, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Pereira, R.I.; Erickson, C.B.; Swibas, T.A.; Cox-York, K.A.; Van Pelt, R.E. Estradiol-mediated improvements in adipose tissue insulin sensitivity are related to the balance of adipose tissue estrogen receptor alpha and beta in postmenopausal women. PLoS ONE 2017, 12, e0176446. [Google Scholar]
- Handa, R.J.; Weiser, M.J. Gonadal steroid hormones and the hypothalamo-pituitary-adrenal axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef] [PubMed]
- Corbier, P.; Edwards, D.A.; Roffi, J. The neonatal testosterone surge: A comparative study. Arch. Int. Physiol. Biochim. Biophys. 1992, 100, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Guo, H.; Guo, Y.; Ebihara, S.; Asada, M.; Ohrui, T.; Furukawa, K.; Ichinose, M.; Yanai, K.; Kudo, Y.; et al. Royal jelly prevents the progression of sarcopenia in aged mice in vivo and in vitro. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Hayashi, K.; Watadani, R.; Okano, Y.; Tanimura, K.; Kotoh, J.; Sasaki, D.; Matsumoto, K.; Maeda, A. Royal jelly improves hyperglycemia in obese/diabetic KK-AY mice. J. Vet. Med. Sci. 2017, 79, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, M.; Kumagai, A.; Hirata, H.; Soga, M.; Yamashita, Y.; Ueda, M.; Ashida, H.; Tsuda, T. 10-hydroxy-2-decenoic acid, a unique medium-chain fatty acid, activates 5′-amp-activated protein kinase in l6 myotubes and mice. Mol. Nutr. Food Res. 2013, 57, 1794–1802. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.F.; Wang, K.; Zhang, Y.Z.; Zheng, Y.F.; Hu, F.L. In vitro anti-inflammatory effects of three fatty acids from royal jelly. Mediat. Inflamm. 2016, 2016, 3583684. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Takahashi, K.; Kuzumaki, A.; Tokoro, S.; Neri, P.; Mori, H. Inhibitory mechanism of 10-hydroxy-trans-2-decenoic acid (Royal Jelly Acid) against lipopolysaccharide- and interferon-beta-induced nitric oxide production. Inflammation 2013, 36, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiser, M.J.; Grimshaw, V.; Wynalda, K.M.; Mohajeri, M.H.; Butt, C.M. Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition. Nutrients 2018, 10, 13. https://doi.org/10.3390/nu10010013
Weiser MJ, Grimshaw V, Wynalda KM, Mohajeri MH, Butt CM. Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition. Nutrients. 2018; 10(1):13. https://doi.org/10.3390/nu10010013
Chicago/Turabian StyleWeiser, Michael J., Vivian Grimshaw, Kelly M. Wynalda, M. Hasan Mohajeri, and Christopher M. Butt. 2018. "Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition" Nutrients 10, no. 1: 13. https://doi.org/10.3390/nu10010013
APA StyleWeiser, M. J., Grimshaw, V., Wynalda, K. M., Mohajeri, M. H., & Butt, C. M. (2018). Long-Term Administration of Queen Bee Acid (QBA) to Rodents Reduces Anxiety-Like Behavior, Promotes Neuronal Health and Improves Body Composition. Nutrients, 10(1), 13. https://doi.org/10.3390/nu10010013