Malnutrition and Fracture Healing: Are Specific Deficiencies in Amino Acids Important in Nonunion Development?


{kind=link}
Abstract
1. Introduction
1.1. Normal Fracture Healing
1.2. Nonunion Development
2. Proteins and Malnutrition in Fracture Healing
2.1. Influence of Collagens and Bone Morphogenetic Proteins on Fracture Healing
2.2. Nutritional Status and Fracture Healing
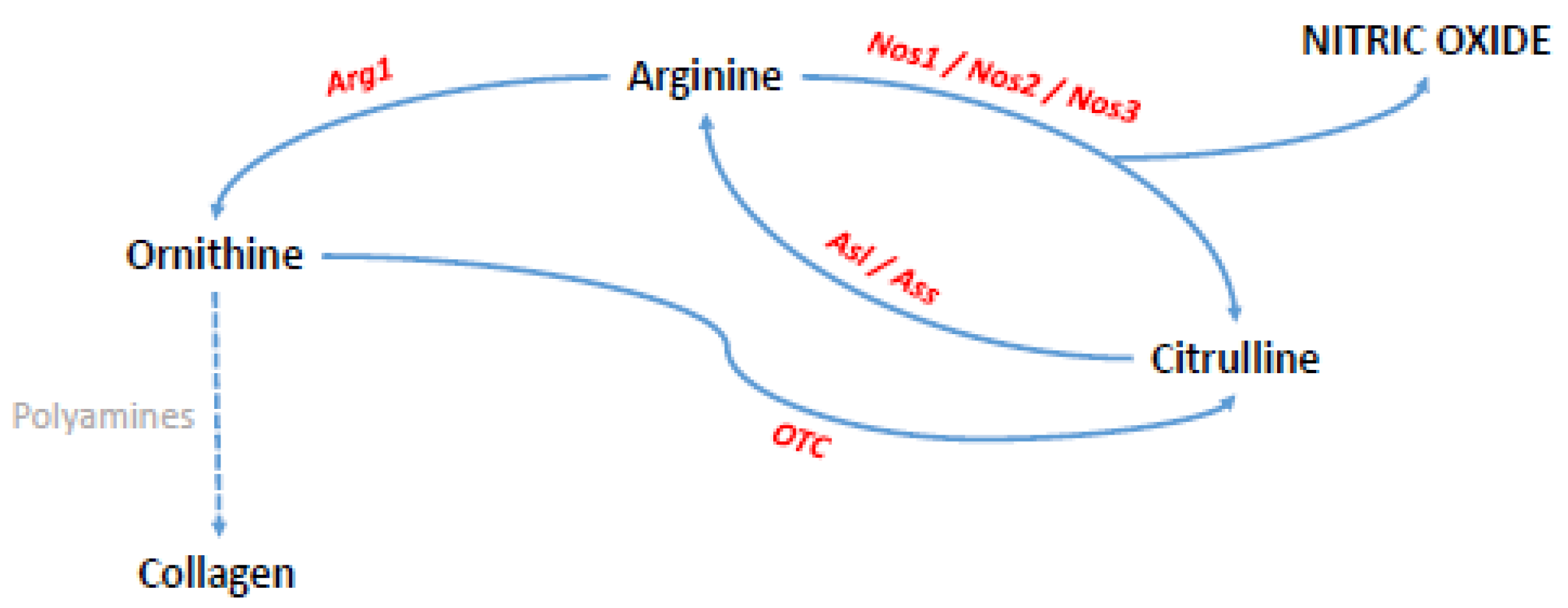
2.3. Influences of Nitric Oxide and Nitric Oxide Synthases on Fracture Healing
2.4. Possible Applications for D-Enantiomeric Amino Acids
3. Concluding Remarks and Future Possibilities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kannus, P.; Niemi, S.; Palvanen, M.; Parkkari, J.; Pasanen, M.; Jarvinen, M.; Vuori, I. Continuously rising problem of osteoporotic knee fractures in elderly women: Nationwide statistics in Finland in 1970–1999 and predictions until the year 2030. Bone 2001, 29, 419–423. [Google Scholar] [CrossRef]
- van Staa, T.P.; Dennison, E.M.; Leufkens, H.G.; Cooper, C. Epidemiology of fractures in England and Wales. Bone 2001, 29, 517–522. [Google Scholar] [CrossRef]
- Johnell, O.; Kanis, J. Epidemiology of osteoporotic fractures. Osteoporos. Int. 2005, 16 (Suppl. 2), S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.J.; Neto, E.; Sousa, D.M.; Leitao, L.; Vasconcelos, D.M.; Ribeiro-Silva, M.; Alencastre, I.S.; Lamghari, M. Fracture pain-Traveling unknown pathways. Bone 2016, 85, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Knuth, A.G.; Hallal, P.C. Temporal trends in physical activity: A systematic review. J. Phys. Act. Health 2009, 6, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Mills, L.A.; Simpson, A.H. The relative incidence of fracture non-union in the Scottish population (5.17 million): A 5-year epidemiological study. BMJ Open 2013, 3, e002276. [Google Scholar] [CrossRef] [PubMed]
- Antonova, E.; Le, T.K.; Burge, R.; Mershon, J. Tibia shaft fractures: Costly burden of nonunions. BMC Musculoskelet. Disord. 2013, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Schindeler, A.; McDonald, M.M.; Bokko, P.; Little, D.G. Bone remodeling during fracture repair: The cellular picture. Semin. Cell Dev. Biol. 2008, 19, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Gerstenfeld, L.C.; Cullinane, D.M.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Fracture healing as a post-natal developmental process: Molecular, spatial, and temporal aspects of its regulation. J. Cell. Biochem. 2003, 88, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Court-Brown, C.M.; McQueen, M.M. Nonunions of the proximal humerus: Their prevalence and functional outcome. J. Trauma 2008, 64, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, T.A. Enhancement of fracture-healing. J. Bone Jt. Surg. Am. 1995, 77, 940–956. [Google Scholar] [CrossRef]
- Wijnands, K.A.; Brink, P.R.; Weijers, P.H.; Dejong, C.H.; Poeze, M. Impaired fracture healing associated with amino acid disturbances. Am. J. Clin. Nutr. 2012, 95, 1270–1277. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.; Guyatt, G.H.; Swiontkowski, M.F.; Tornetta, P., 3rd; Sprague, S.; Schemitsch, E.H. A lack of consensus in the assessment of fracture healing among orthopaedic surgeons. J. Orthop. Trauma 2002, 16, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Frolke, J.P.; Patka, P. Definition and classification of fracture non-unions. Injury 2007, 38 (Suppl. 2), S19–S22. [Google Scholar] [CrossRef]
- Megas, P. Classification of non-union. Injury 2005, 36 (Suppl. 4), S30–S37. [Google Scholar] [PubMed]
- Bishop, J.A.; Palanca, A.A.; Bellino, M.J.; Lowenberg, D.W. Assessment of compromised fracture healing. J. Am. Acad. Orthop. Surg. 2012, 20, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Court-Brown, C.M.; McQueen, M.M.; Wakefield, A.E. Estimating the risk of nonunion following nonoperative treatment of a clavicular fracture. J. Bone Jt. Surg. Am. 2004, 86-A, 1359–1365. [Google Scholar] [CrossRef]
- Kozin, S.H. Incidence, mechanism, and natural history of scaphoid fractures. Hand Clin. 2001, 17, 515–524. [Google Scholar] [PubMed]
- Rosenberg, G.A.; Sferra, J.J. Treatment strategies for acute fractures and nonunions of the proximal fifth metatarsal. J. Am. Acad. Orthop. Surg. 2000, 8, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.R.; Hock, J.M.; Burr, D.B. Periosteum: Biology, regulation, and response to osteoporosis therapies. Bone 2004, 35, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.J.; van Gastel, N.; Carmeliet, G.; Luyten, F.P. Uncovering the periosteum for skeletal regeneration: The stem cell that lies beneath. Bone 2015, 70C, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Calori, G.M.; Albisetti, W.; Agus, A.; Iori, S.; Tagliabue, L. Risk factors contributing to fracture non-unions. Injury 2007, 38 (Suppl. 2), S11–S18. [Google Scholar] [CrossRef]
- Gustilo, R.B.; Gruninger, R.P.; Davis, T. Classification of type III (severe) open fractures relative to treatment and results. Orthopedics 1987, 10, 1781–1788. [Google Scholar] [PubMed]
- Schmidmaier, G.; Capanna, R.; Wildemann, B.; Beque, T.; Lowenberg, D. Bone morphogenetic proteins in critical-size bone defects: What are the options? Injury 2009, 40 (Suppl. 3), S39–S43. [Google Scholar] [CrossRef]
- Hernandez, R.K.; Do, T.P.; Critchlow, C.W.; Dent, R.E.; Jick, S.S. Patient-related risk factors for fracture-healing complications in the United Kingdom General Practice Research Database. Acta Orthop. 2012, 83, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Court-Brown, C.M.; Caesar, B. Epidemiology of adult fractures: A review. Injury 2006, 37, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Buhr, A.J.; Cooke, A.M. Fracture patterns. Lancet 1959, 1, 531–536. [Google Scholar] [CrossRef]
- Brinker, M.R.; O’Connor, D.P.; Monla, Y.T.; Earthman, T.P. Metabolic and endocrine abnormalities in patients with nonunions. J. Orthop. Trauma 2007, 21, 557–570. [Google Scholar] [CrossRef] [PubMed]
- MacKay, D.; Miller, A.L. Nutritional support for wound healing. Altern. Med. Rev. 2003, 8, 359–377. [Google Scholar] [PubMed]
- Cunniffe, G.M.; Dickson, G.R.; Partap, S.; Stanton, K.T.; O’Brien, F.J. Development and characterisation of a collagen nano-hydroxyapatite composite scaffold for bone tissue engineering. J. Mater. Sci. Mater. Med. 2010, 21, 2293–2298. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.L. The effect of ascorbate on wound healing. Int. Ophthalmol. Clin. 2000, 40, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.M.; Ringshia, R.A.; Legeros, R.Z.; Clark, E.; Yost, M.J.; Terracio, L.; Teixeira, C.C. An improved collagen scaffold for skeletal regeneration. J. Biomed. Mater. Res. A 2010, 94, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Nakase, T.; Yoshikawa, H. Potential roles of bone morphogenetic proteins (BMPs) in skeletal repair and regeneration. J. Bone Miner. Metab. 2006, 24, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Axelrad, T.W.; Einhorn, T.A. Bone morphogenetic proteins in orthopaedic surgery. Cytokine Growth Factor Rev. 2009, 20, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Jiang, W.; Phillips, F.M.; Haydon, R.C.; Peng, Y.; Zhou, L.; Luu, H.H.; An, N.; Breyer, B.; Vanichakarn, P.; et al. Osteogenic activity of the fourteen types of human bone morphogenetic proteins (BMPs). J. Bone Jt. Surg. Am. 2003, 85-A, 1544–1552. [Google Scholar] [CrossRef]
- Cho, T.J.; Gerstenfeld, L.C.; Einhorn, T.A. Differential temporal expression of members of the transforming growth factor beta superfamily during murine fracture healing. J. Bone Miner. Res. 2002, 17, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Kloen, P.; Di Paola, M.; Borens, O.; Richmond, J.; Perino, G.; Helfet, D.L.; Goumans, M.J. BMP signaling components are expressed in human fracture callus. Bone 2003, 33, 362–371. [Google Scholar] [CrossRef]
- Barnes, G.L.; Kostenuik, P.J.; Gerstenfeld, L.C.; Einhorn, T.A. Growth factor regulation of fracture repair. J. Bone Miner. Res. 1999, 14, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Lojudice, F.H.; Halcsik, E.; Navarro, R.D.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Facts, challenges, and future perspectives. J. Dent. Res. 2014, 93, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Valenzuela, J.C.; Halcsik, E.; Bassi, E.J.; Demasi, M.A.; Granjeiro, J.M.; Sogayar, M.C. Expression, purification, bioactivity, and partial characterization of a recombinant human bone morphogenetic protein-7 produced in human 293T cells. Mol. Biotechnol. 2010, 46, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Giannoudis, P.V.; Gudipati, S.; Harwood, P.; Kanakaris, N.K. Long bone non-unions treated with the diamond concept: A case series of 64 patients. Injury 2015, 46 (Suppl. 8), S48–S54. [Google Scholar] [CrossRef]
- Calori, G.M.; Mazza, E.; Colombo, M.; Ripamonti, C.; Tagliabue, L. Treatment of long bone non-unions with polytherapy: Indications and clinical results. Injury 2011, 42, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Secreto, F.J.; Hoeppner, L.H.; Westendorf, J.J. Wnt signaling during fracture repair. Curr. Osteoporos. Rep. 2009, 7, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Westendorf, J.J.; Kahler, R.A.; Schroeder, T.M. Wnt signaling in osteoblasts and bone diseases. Gene 2004, 341, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Hoeflich, K.P.; Woodgett, J.R. Glycogen synthase kinase-3: Properties, functions, and regulation. Chem. Rev. 2001, 101, 2527–2540. [Google Scholar] [CrossRef] [PubMed]
- Bao, Q.; Chen, S.; Qin, H.; Feng, J.; Liu, H.; Liu, D.; Li, A.; Shen, Y.; Zhao, Y.; Li, J.; et al. An appropriate Wnt/beta-catenin expression level during the remodeling phase is required for improved bone fracture healing in mice. Sci. Rep. 2017, 7, 2695. [Google Scholar] [CrossRef] [PubMed]
- Deutz, N.E.; Matheson, E.M.; Matarese, L.E.; Luo, M.; Baggs, G.E.; Nelson, J.L.; Hegazi, R.A.; Tappenden, K.A.; Ziegler, T.R. Readmission and mortality in malnourished, older, hospitalized adults treated with a specialized oral nutritional supplement: A randomized clinical trial. Clin. Nutr. 2016, 35, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Koski, K.; Luukinen, H.; Laippala, P.; Kivela, S.L. Risk factors for major injurious falls among the home-dwelling elderly by functional abilities. A prospective population-based study. Gerontology 1998, 44, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, T.; Hedstrom, M. Nutritional treatment of bone fracture. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Lill, C.A.; Hesseln, J.; Schlegel, U.; Eckhardt, C.; Goldhahn, J.; Schneider, E. Biomechanical evaluation of healing in a non-critical defect in a large animal model of osteoporosis. J. Orthop. Res. 2003, 21, 836–842. [Google Scholar] [CrossRef]
- Neyens, J.; Halfens, R.; Spreeuwenberg, M.; Meijers, J.; Luiking, Y.; Verlaan, G.; Schols, J. Malnutrition is associated with an increased risk of falls and impaired activity in elderly patients in Dutch residential long-term care (LTC): A cross-sectional study. Arch. Gerontol. Geriatr. 2013, 56, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.L.; Ong, K.C.; Chan, Y.H.; Loke, W.C.; Ferguson, M.; Daniels, L. Malnutrition and its impact on cost of hospitalization, length of stay, readmission and 3-year mortality. Clin. Nutr. 2012, 31, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Oakland, K.; Nadler, R.; Cresswell, L.; Jackson, D.; Coughlin, P.A. Systematic review and meta-analysis of the association between frailty and outcome in surgical patients. Ann. R. Coll. Surg. Engl. 2016, 98, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.H.; Sun, S.; Walls, R.C. Protein-energy undernutrition among elderly hospitalized patients: A prospective study. JAMA 1999, 281, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Allard, J.P.; Keller, H.; Jeejeebhoy, K.N.; Laporte, M.; Duerksen, D.R.; Gramlich, L.; Payette, H.; Bernier, P.; Vesnaver, E.; Davidson, B.; et al. Malnutrition at Hospital Admission-Contributors and Effect on Length of Stay: A Prospective Cohort Study From the Canadian Malnutrition Task Force. JPEN J. Parenter. Enter. Nutr. 2015, 40, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Dirks, M.L.; Snijders, T.; Senden, J.M.; Dolmans, J.; van Loon, L.J. Substantial skeletal muscle loss occurs during only 5 days of disuse. Acta Physiol. 2014, 210, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, P.M.; Blackwell, T.L.; Cauley, J.; Kado, D.M.; Barrett-Connor, E.; Lee, C.G.; Hoffman, A.R.; Nevitt, M.; Stefanick, M.L.; Lane, N.E.; et al. Evaluation of the Usefulness of Consensus Definitions of Sarcopenia in Older Men: Results from the Observational Osteoporotic Fractures in Men Cohort Study. J. Am. Geriatr. Soc. 2015, 63, 2247–2259. [Google Scholar] [CrossRef] [PubMed]
- Zura, R.; Braid-Forbes, M.J.; Jeray, K.; Mehta, S.; Einhorn, T.A.; Watson, J.T.; Della Rocca, G.J.; Forbes, K.; Steen, R.G. Bone fracture nonunion rate decreases with increasing age: A prospective inception cohort study. Bone 2017, 95, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Poeze, M.; Bruins, M.J.; Kessels, F.; Luiking, Y.C.; Lamers, W.H.; Deutz, N.E. Effects of L-arginine pretreatment on nitric oxide metabolism and hepatosplanchnic perfusion during porcine endotoxemia. Am. J. Clin. Nutr. 2011, 93, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, K.A.; Vink, H.; Briede, J.J.; van Faassen, E.E.; Lamers, W.H.; Buurman, W.A.; Poeze, M. Citrulline a More Suitable Substrate than Arginine to Restore NO Production and the Microcirculation during Endotoxemia. PLoS ONE 2012, 7, e37439. [Google Scholar] [CrossRef] [PubMed]
- Kdolsky, R.K.; Mohr, W.; Savidis-Dacho, H.; Beer, R.; Puig, S.; Reihsner, R.; Tangl, S.; Donath, K. The influence of oral L-arginine on fracture healing: An animal study. Wien. Klin. Wochenschr. 2005, 117, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Angele, M.K.; Fitzal, F.; Smail, N.; Knoferl, M.W.; Schwacha, M.G.; Ayala, A.; Wang, P.; Chaudry, I.H. L-arginine attenuates trauma-hemorrhage-induced liver injury. Crit. Care Med. 2000, 28, 3242–3248. [Google Scholar] [CrossRef] [PubMed]
- Luiking, Y.C.; Poeze, M.; Deutz, N.E. Arginine infusion in patients with septic shock increases nitric oxide production without haemodynamic instability. Clin. Sci. 2015, 128, 57–67. [Google Scholar] [CrossRef] [PubMed]
- van Wijck, K.; Wijnands, K.A.; Meesters, D.M.; Boonen, B.; van Loon, L.J.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K.; Poeze, M. L-citrulline improves splanchnic perfusion and reduces gut injury during exercise. Med. Sci. Sports Exerc. 2014, 46, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.S.; Rivera, M.; Puccinelli, C.L.; Wang, M.Y.; Williams, S.J.; Barber, A.E. Targeted amino acid supplementation in diabetic foot wounds: Pilot data and a review of the literature. Surg. Infect. 2014, 15, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Luiking, Y.C.; Poeze, M.; Ramsay, G.; Deutz, N.E. Reduced citrulline production in sepsis is related to diminished de novo arginine and nitric oxide production. Am. J. Clin. Nutr. 2009, 89, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Reynolds, J.; Thom, A.; Kinsley, L.; Dietrick-Gallagher, M.; Shou, J.; Ruggieri, B. Immune and metabolic effects of arginine in the surgical patient. Ann. Surg. 1988, 208, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.H.; Stefanovic-Racic, M.; Lancaster, J. Nitric oxide and its role in orthopaedic disease. Clin. Orthop. Relat. Res. 1995, 275–294. [Google Scholar]
- Long, C.L.; Geiger, J.W.; Richards, E.W.; Akin, J.M.; Blakemore, W.S. Plasma amino acid concentrations in geriatric control and hip-fracture patients. Am. J. Clin. Nutr. 1992, 55, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Woolf, L.I.; Grovers, A.C.; Moore, J.P.; Duff, J.H.; Finley, R.J.; Loomer, R.L. Arterial plasma amino acids in patients with serious postoperative infection and in patients with major fractures. Surgery 1976, 79, 283–292. [Google Scholar] [PubMed]
- Vittur, F.; Lunazzi, G.; Moro, L.; Stagni, N.; de Bernard, B.; Moretti, M.; Stanta, G.; Bacciottini, F.; Orlandini, G.; Reali, N.; et al. A possible role for polyamines in cartilage in the mechanism of calcification. Biochim. Biophys. Acta 1986, 881, 38–45. [Google Scholar] [CrossRef]
- Patterson, B.M.; Cornell, C.N.; Carbone, B.; Levine, B.; Chapman, D. Protein depletion and metabolic stress in elderly patients who have a fracture of the hip. J. Bone Jt. Surg. Am. 1992, 74, 251–260. [Google Scholar] [CrossRef]
- Fini, M.; Aldini, N.N.; Cane, V.; Zaffe, D.; Giavaresi, G.; Rocca, M.; Guzzardella, G.A.; Giardino, R. Effects of essential amino acids and lactose on bony fractures and defects in rabbits: A preliminary histomorphometric study. Arch. Orthop. Trauma Surg. 1999, 119, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Volpi, E.; Kobayashi, H.; Sheffield-Moore, M.; Mittendorfer, B.; Wolfe, R.R. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am. J. Clin. Nutr. 2003, 78, 250–258. [Google Scholar] [CrossRef] [PubMed]
- De Bandt, J.P.; Cynober, L.A. Amino acids with anabolic properties. Curr. Opin. Clin. Nutr. Metab. Care 1998, 1, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.S.; Kazmier, P.; Burd, T.A.; Anglen, J.; Stoker, A.M.; Kuroki, K.; Carson, W.L.; Cook, J.L. Enhanced fracture and soft-tissue healing by means of anabolic dietary supplementation. J. Bone Jt. Surg. Am. 2006, 88, 2386–2394. [Google Scholar] [CrossRef] [PubMed]
- Day, S.M.; DeHeer, D.H. Reversal of the detrimental effects of chronic protein malnutrition on long bone fracture healing. J. Orthop. Trauma 2001, 15, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, P.; Fini, M.; Giavaresi, G.; Giardino, R. Human osteopenic bone-derived osteoblasts: Essential amino acids treatment effects. Artif. Cells Blood Substit. Immobil. Biotechnol. 2003, 31, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Chevalley, T.; Rizzoli, R.; Manen, D.; Caverzasio, J.; Bonjour, J.P. Arginine increases insulin-like growth factor-I production and collagen synthesis in osteoblast-like cells. Bone 1998, 23, 103–109. [Google Scholar] [CrossRef]
- Trippel, S.B. Potential role of insulinlike growth factors in fracture healing. Clin. Orthop. Relat. Res. 1998, S301–S313. [Google Scholar] [CrossRef]
- Fini, M.; Torricelli, P.; Giavaresi, G.; Carpi, A.; Nicolini, A.; Giardino, R. Effect of L-lysine and L-arginine on primary osteoblast cultures from normal and osteopenic rats. Biomed. Pharmacother. 2001, 55, 213–220. [Google Scholar] [CrossRef]
- Diwan, A.D.; Wang, M.X.; Jang, D.; Zhu, W.; Murrell, G.A. Nitric oxide modulates fracture healing. J. Bone Miner. Res. 2000, 15, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, M.R.; Tantry, U.; Gross, S.S.; Wasserburg, H.L.; Barbul, A. Nitric oxide regulates wound healing. J. Surg. Res. 1996, 63, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Murrell, G.A.; Szabo, C.; Hannafin, J.A.; Jang, D.; Dolan, M.M.; Deng, X.H.; Murrell, D.F.; Warren, R.F. Modulation of tendon healing by nitric oxide. Inflamm. Res. 1997, 46, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, H.; Kohsaka, H.; Liu, M.F.; Higashiyama, H.; Hirata, Y.; Kanno, K.; Saito, I.; Miyasaka, N. Nitric oxide production and inducible nitric oxide synthase expression in inflammatory arthritides. J. Clin. Investig. 1995, 96, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Stefanovic-Racic, M.; Meyers, K.; Meschter, C.; Coffey, J.W.; Hoffman, R.A.; Evans, C.H. N-monomethyl arginine, an inhibitor of nitric oxide synthase, suppresses the development of adjuvant arthritis in rats. Arthritis Rheum. 1994, 37, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J.; De Marco, G.; Gangula, P.; Yallampalli, C. Nitric oxide donor alleviates ovariectomy-induced bone loss. Bone 1996, 18, 301–304. [Google Scholar] [CrossRef]
- Hukkanen, M.; Corbett, S.A.; Batten, J.; Konttinen, Y.T.; McCarthy, I.D.; Maclouf, J.; Santavirta, S.; Hughes, S.P.; Polak, J.M. Aseptic loosening of total hip replacement. Macrophage expression of inducible nitric oxide synthase and cyclo-oxygenase-2, together with peroxynitrite formation, as a possible mechanism for early prosthesis failure. J. Bone Jt. Surg. Br. 1997, 79, 467–474. [Google Scholar] [CrossRef]
- Xia, W.; Szomor, Z.; Wang, Y.; Murrell, G.A. Nitric oxide enhances collagen synthesis in cultured human tendon cells. J. Orthop. Res. 2006, 24, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Ekeland, A.; Engesaeter, L.B.; Langeland, N. Mechanical properties of fractured and intact rat femora evaluated by bending, torsional and tensile tests. Acta Orthop. Scand. 1981, 52, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.H.; Owan, I.; Jacob, D.S.; McClintock, R.; Peacock, M. Effects of nitric oxide synthase inhibitors on bone formation in rats. Bone 1997, 21, 487–490. [Google Scholar] [CrossRef]
- Shabani, M.; Pulfer, S.K.; Bulgrin, J.P.; Smith, D.J. Enhancement of wound repair with a topically applied nitric oxide-releasing polymer. Wound Repair Regen. 1996, 4, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Edington, H.D.; McClosky, C.; Tzeng, E.; Lizonova, A.; Kovesdi, I.; Steed, D.L.; Billiar, T.R. Reversal of impaired wound repair in iNOS-deficient mice by topical adenoviral-mediated iNOS gene transfer. J. Clin. Investig. 1998, 101, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Baldik, Y.; Diwan, A.D.; Appleyard, R.C.; Fang, Z.M.; Wang, Y.; Murrell, G.A. Deletion of iNOS gene impairs mouse fracture healing. Bone 2005, 37, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Diwan, A.D.; Lin, J.H.; Murrell, G.A. Nitric oxide synthase isoforms during fracture healing. J. Bone Miner. Res. 2001, 16, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, T.A.; Majeska, R.J.; Rush, E.B.; Levine, P.M.; Horowitz, M.C. The expression of cytokine activity by fracture callus. J. Bone Miner. Res. 1995, 10, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Hikiji, H.; Shin, W.S.; Oida, S.; Takato, T.; Koizumi, T.; Toyo-oka, T. Direct action of nitric oxide on osteoblastic differentiation. FEBS Lett. 1997, 410, 238–242. [Google Scholar] [CrossRef]
- Zhu, W.; Murrell, G.A.; Lin, J.; Gardiner, E.M.; Diwan, A.D. Localization of nitric oxide synthases during fracture healing. J. Bone Miner. Res. 2002, 17, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Pilla, A.A. Nonthermal electromagnetic fields: From first messenger to therapeutic applications. Electromagn. Biol. Med. 2013, 32, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.H.; Takano, Y.; Owan, I.; Murrell, G.A. Nitric oxide inhibitor L-NAME suppresses mechanically induced bone formation in rats. Am. J. Physiol. 1996, 270, E634–E639. [Google Scholar] [CrossRef] [PubMed]
- de Albuquerque, R.F., Jr.; Del Bel, E.A.; Brentegani, L.G.; Moura de Oliveira, M.T.; Issa, J.P.M. Trigeminal nitric oxide synthase expression correlates with new bone formation during distraction osteogenesis. Calcif. Tissue Int. 2008, 82, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Watanuki, M.; Sakai, A.; Sakata, T.; Tsurukami, H.; Miwa, M.; Uchida, Y.; Watanabe, K.; Ikeda, K.; Nakamura, T. Role of inducible nitric oxide synthase in skeletal adaptation to acute increases in mechanical loading. J. Bone Miner. Res. 2002, 17, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Meesters, D.M.; Neubert, S.; Wijnands, K.A.; Heyer, F.L.; Zeiter, S.; Ito, K.; Brink, P.R.; Poeze, M. Deficiency of inducible and endothelial nitric oxide synthase results in diminished bone formation and delayed union and nonunion development. Bone 2016, 83, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.J., Jr.; Prieto, E.M.; Krueger, C.A.; Zienkiewicz, K.J.; Romano, D.R.; Ward, C.L.; Akers, K.S.; Guelcher, S.A.; Wenke, J.C. Effects of local delivery of d-amino acids from biofilm-dispersive scaffolds on infection in contaminated rat segmental defects. Biomaterials 2013, 34, 7533–7543. [Google Scholar] [CrossRef] [PubMed]
- Harmata, A.J.; Ma, Y.; Sanchez, C.J.; Zienkiewicz, K.J.; Elefteriou, F.; Wenke, J.C.; Guelcher, S.A. d-amino acid inhibits biofilm but not new bone formation in an ovine model. Clin. Orthop. Relat. Res. 2015, 473, 3951–3961. [Google Scholar] [CrossRef] [PubMed]
- Holstein, J.H.; Garcia, P.; Histing, T.; Kristen, A.; Scheuer, C.; Menger, M.D.; Pohlemann, T. Advances in the establishment of defined mouse models for the study of fracture healing and bone regeneration. J. Orthop. Trauma 2009, 23, S31–S38. [Google Scholar] [CrossRef] [PubMed]
- Inzana, J.A.; Schwarz, E.M.; Kates, S.L.; Awad, H.A. A novel murine model of established Staphylococcal bone infection in the presence of a fracture fixation plate to study therapies utilizing antibiotic-laden spacers after revision surgery. Bone 2015, 72, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Reizner, W.; Hunter, J.G.; O’Malley, N.T.; Southgate, R.D.; Schwarz, E.M.; Kates, S.L. A systematic review of animal models for Staphylococcus aureus osteomyelitis. Eur. Cells Mater. 2014, 27, 196–212. [Google Scholar] [CrossRef]
- Li, D.; Gromov, K.; Soballe, K.; Puzas, J.E.; O’Keefe, R.J.; Awad, H.; Drissi, H.; Schwarz, E.M. Quantitative mouse model of implant-associated osteomyelitis and the kinetics of microbial growth, osteolysis, and humoral immunity. J. Orthop. Res. 2008, 26, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; Leid, J.G.; Calhoun, J.H.; Costerton, J.W.; Shirtliff, M.E. Osteomyelitis and the role of biofilms in chronic infection. FEMS Immunol. Med. Microbiol. 2008, 52, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Montanaro, L.; Arciola, C.R. Biofilm in implant infections: Its production and regulation. Int. J. Artif. Organs 2005, 28, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.; Costerton, W.; Sewecke, J.; Altman, D. Molecular techniques to detect biofilm bacteria in long bone nonunion: A case report. Clin. Orthop. Relat. Res. 2011, 469, 3037–3042. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meesters, D.M.; Wijnands, K.A.P.; Brink, P.R.G.; Poeze, M. Malnutrition and Fracture Healing: Are Specific Deficiencies in Amino Acids Important in Nonunion Development? Nutrients 2018, 10, 1597. https://doi.org/10.3390/nu10111597
Meesters DM, Wijnands KAP, Brink PRG, Poeze M. Malnutrition and Fracture Healing: Are Specific Deficiencies in Amino Acids Important in Nonunion Development? Nutrients. 2018; 10(11):1597. https://doi.org/10.3390/nu10111597
Chicago/Turabian StyleMeesters, Dennis M., Karolina A.P. Wijnands, Peter R.G. Brink, and Martijn Poeze. 2018. "Malnutrition and Fracture Healing: Are Specific Deficiencies in Amino Acids Important in Nonunion Development?" Nutrients 10, no. 11: 1597. https://doi.org/10.3390/nu10111597
APA StyleMeesters, D. M., Wijnands, K. A. P., Brink, P. R. G., & Poeze, M. (2018). Malnutrition and Fracture Healing: Are Specific Deficiencies in Amino Acids Important in Nonunion Development? Nutrients, 10(11), 1597. https://doi.org/10.3390/nu10111597