Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorate Adipose Insulin Resistance and Inflammation in High-Fat Diet-Fed C57BL/6J Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Bovine α-Lactalbumin Hydrolysates (α-LAH)
2.3. Animals and Diets
2.4. Fasting Blood Glucose, Fasting Serum Insulin and HOMA-IR (Homeostatic Model Assessment of Insulin Resistance)
2.5. Oral Glucose Tolerance Test (OGTT)
2.6. Intraperitoneal Insulin Tolerance Test (ipITT)
2.7. Reverse Transcription Quantitative Real-Time PCR (RT-qPCR)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
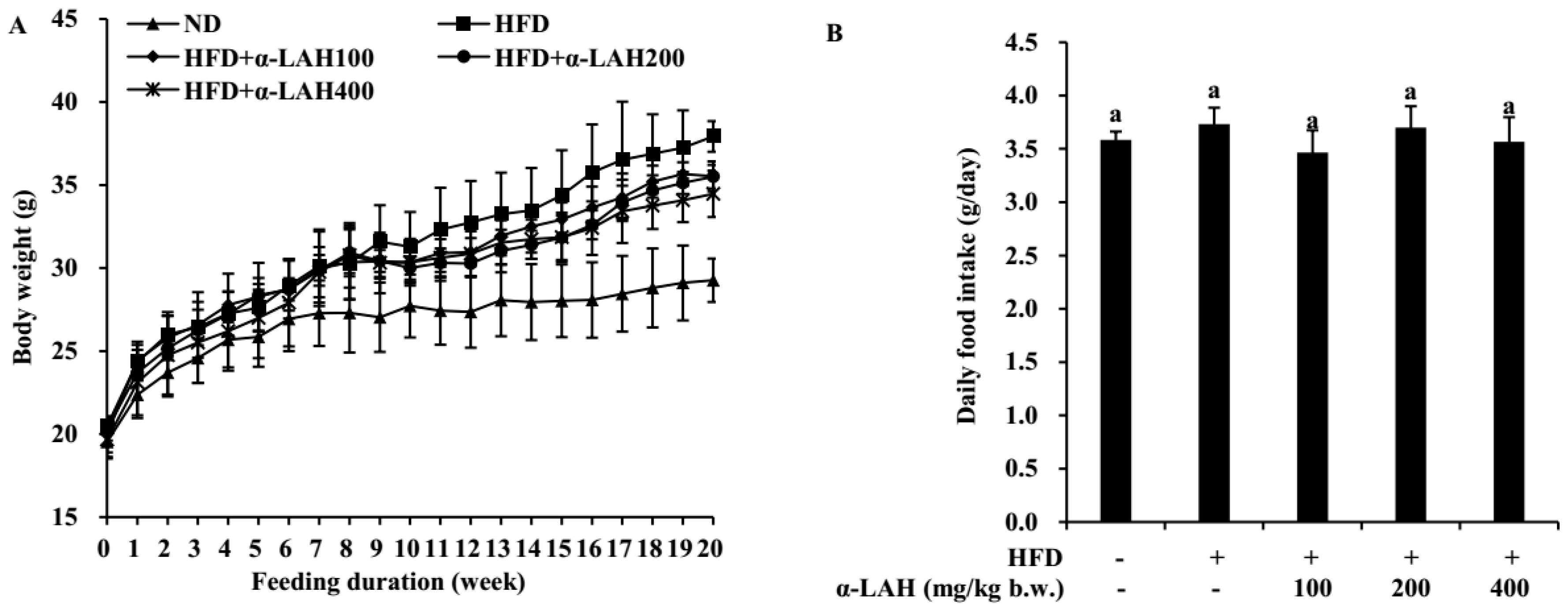
3.1. Bovine α-Lactalbumin Hydrolysates (α-LAH) Prevented Body Weight Gain in HFD-Fed C57BL/6J Mice
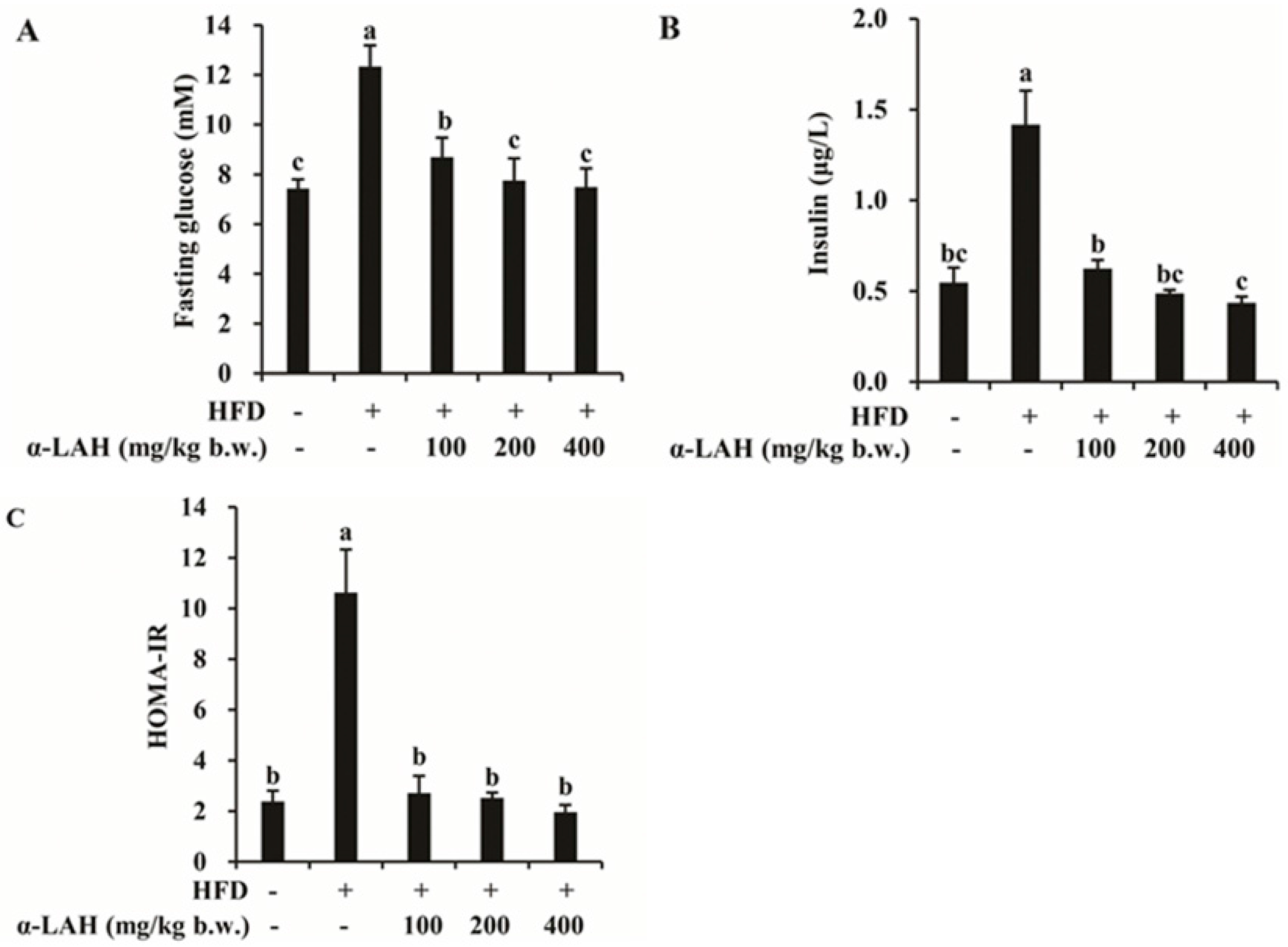
3.2. Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorated Hyperglycemia and Hyperinsulinemia in HFD Fed C57BL/6J Mice
3.3. Bovine α-Lactalbumin Hydrolysates (α-LAH) Improved Glucose and Insulin Tolerance in HFD-Fed C57BL/6J Mice
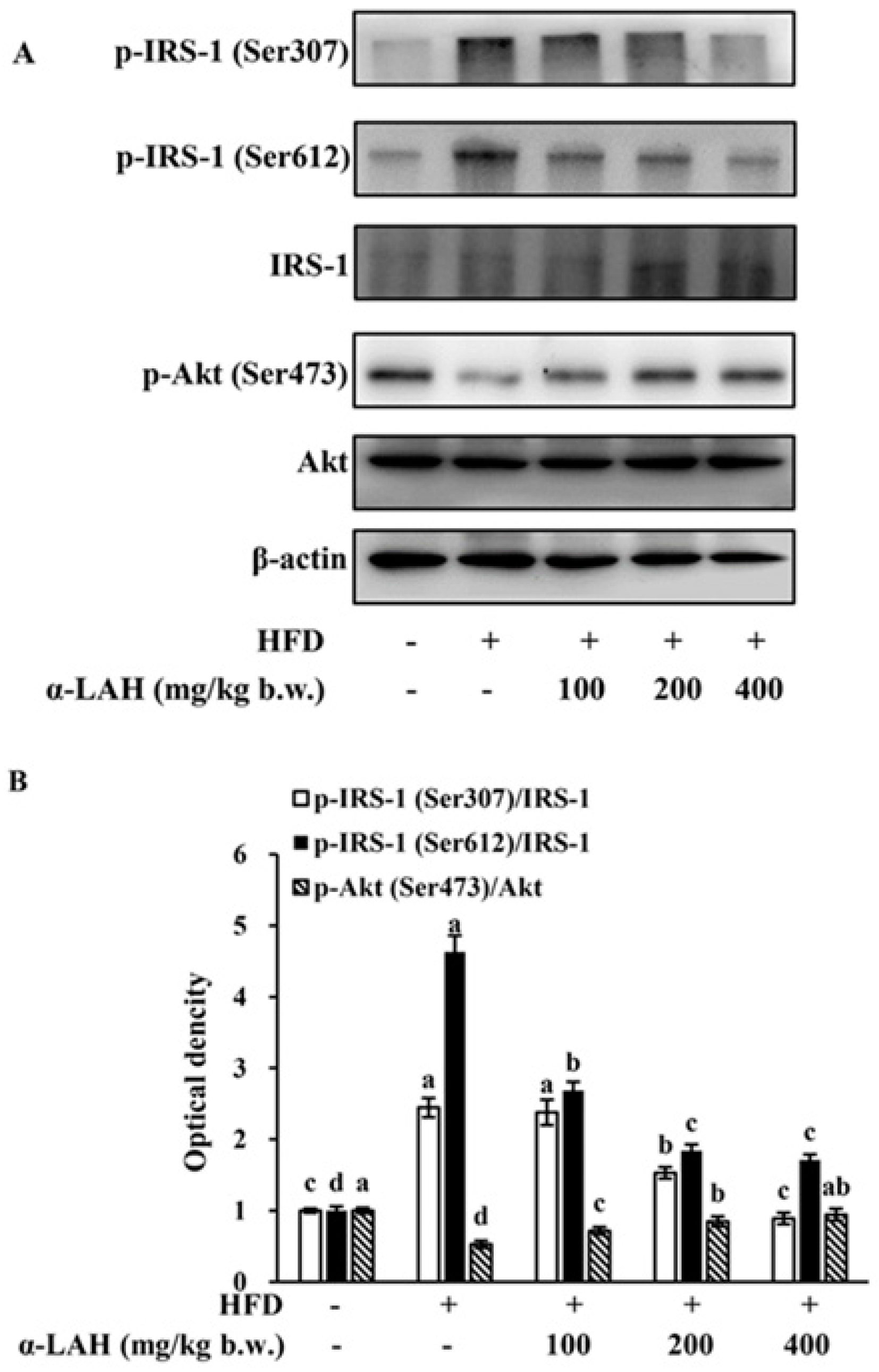
3.4. Bovine α-Lactalbumin Hydrolysates (α-LAH) Suppressed IRS-1 (Ser307, Ser612) Phosphorylation and Enhanced Akt (Ser473) Phosphorylation in Epididymal Adipose Tissues of HFD Fed C57BL/6J Mice
3.5. Bovine α-Lactalbumin Hydrolysates (α-LAH) Downregulated Inflammation-Related Gene Expression in Epididymal Adipose Tissues of HFD-Fed C57BL/6J Mice
3.6. Bovine α-Lactalbumin Hydrolysates (α-LAH) Inhibited the Activation of IKK Signaling Pathway in Epididymal Adipose Tissues of HFD Fed C57BL/6J Mice
3.7. Bovine α-Lactalbumin Hydrolysates (α-LAH) Inhibited the Activation of MAPK Signaling Pathway in Epididymal Adipose Tissues of HFD-Fed C57BL/6J Mice
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hannibal, T.D.; Schmidt-Christensen, A.; Nilsson, J.; Pettersson, N.F.; Hansen, L.; Holmberg, D. Deficiency in plasmacytoid dendritic cells and type I interferon signalling prevents diet-induced obesity and insulin resistance in mice. Diabetologia 2017, 60, 2033–2041. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in obesity: Mechanisms and potential targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; Leroith, D.; Karnieli, E. Insulin resistance in obesity as the underlying cause for the metabolic syndrome. Mt. Sinai J. Med. 2010, 77, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Osborn, O.; Olefsky, J.M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med. 2012, 18, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Bashir, S.; Sharma, Y.; Elahi, A.; Khan, F. Amelioration of obesity-associated inflammation and insulin resistance in c57bl/6 mice via macrophage polarization by fish oil supplementation. J. Nutr. Biochem. 2016, 33, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.J.; Li, A.Y.; Feng, X.; Hou, T.; Liu, K.; Liu, B.L.; Zhang, N. Metformin and resveratrol ameliorate muscle insulin resistance through preventing lipolysis and inflammation in hypoxic adipose tissue. Cell Signal. 2016, 28, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Uysal, K.T.; Wiesbrock, S.M.; Mario, M.W.; Hotamisligil, G.S. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature 1997, 389, 610–614. [Google Scholar] [PubMed]
- Matsuki, T.; Horai, R.; Sudo, K.; Iwakura, Y. IL-1 Plays an important role in lipid metabolism by regulating insulin levels under physiological conditions. J. Exp. Med. 2003, 198, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Hunter, D.; Huber, R.; Lemieux, J.; Slaymaker, S.; Vaddi, K.; Charo, I.; Leibel, R.L. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J. Clin. Invest. 2006, 116, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Ma, S.; Li, F.; Yuan, M.; Xu, H.B.; Huang, K.X. Catalpol ameliorates high-fat diet-induced insulin resistance and adipose tissue inflammation by suppressing the JNK and NF-κB pathways. Biochem. Biophys. Res. Commun. 2015, 467, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Gual, P.; Le, M.; Tanti, J.F. Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie 2005, 87, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, Y.J.; Fillmore, J.J.; Chen, Y.; Moore, I. Prevention of fat-induced insulin resistance by salicylate. J. Clin. Invest. 2001, 108, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Arkan, M.C.; Hevener, A.L.; Greten, F.R.; Maeda, S.; Li, Z.W.; Long, J.M.; Wynshaw-Boris, A.; Poli, G.; Olefsky, J.; Karin, M. IKK-β links inflammation to obesity-induced insulin resistance. Nat. Med. 2005, 11, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Han, M.S.; Jung, D.Y.; Morel, C.; Lakhani, S.A.; Kim, J.K.; Flavell, R.A.; Davis, R.J. JNK Expression by Macrophages Promotes Obesity-Induced Insulin Resistance and Inflammation. Science 2013, 339, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.H.; Nickerson, M.T. The effect of pH and temperature pre-treatments on the structure, surface characteristics and emulsifying properties of alpha-lactalbumin. Food Chem. 2015, 173, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, F.; Moeeni, M. Analysis of binding interaction of genistein and kaempferol with bovine α-lactalbumin. J. Funct. Foods 2015, 12, 458–467. [Google Scholar] [CrossRef]
- Jiang, Z.M.; Yuan, X.Y.; Yao, K.; Li, X.Y.; Zhang, X.L.; Mu, Z.S.; Jiang, L.Z.; Hou, J.C. Laccase-aided modification: Effects on structure, gel properties and antioxidant activities of α-lactalbumin. Lwt—Food Sci. Technol. 2017, 80, 355–363. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Uchida, M. α-lactalbumin suppresses interleukin-6 release after intestinal ischemia/reperfusion via nitric oxide in rats. Inflammopharmacology 2007, 15, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Takai, S.; Hosono, A.; Seki, T. Bovine milk-derived α-lactalbumin inhibits colon inflammation and carcinogenesis in azoxymethane and dextran sodium sulfate-treated mice. Biosci. Biotechnol. Biochem. 2014, 78, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Blouet, C.; Mariotti, F.; Azzout-Marniche, D.; Mathe, V.; Mikogami, T.; Tome, D.; Huneau, J.F. Dietary cysteine alleviates sucrose-induced oxidative stress and insulin resistance. Free Radic. Biol. Med. 2007, 42, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Blat, S.; Vincent, S.; Lefeuvre, L.; Lemoine-Morel, S.; Malbert, C.H.; Pichon, L.; Mikogami, T.; Gratas-Delamarche, A. Dietary α-lactalbumin supplementation alleviates normocaloric western diet-induced glucose intolerance in Gottingen minipigs. Obesity 2015, 23, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Peptide array on cellulose support-a screening tool to identify peptides with dipeptidyl-deptidase IV inhibitory activity within the sequence of α-lactalbumin. Int. J. Mol. Sci. 2014, 15, 20846–20858. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Le Maux, S.; Hamayon, J.; FitzGerald, R.J. Strategies for the release of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides in an enzymatic hydrolyzate of α-lactalbumin. Food Funct. 2016, 7, 3437–3443. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Fernandez, A.M.; Lopez-Pedemonte, T.; Medrano-Fernandez, A. Evaluation of antioxidant, antiglycant and ACE-inhibitory activity in enzymatic hydrolysates of α-lactalbumin. Food Nutr. Sci. 2017, 8, 84–98. [Google Scholar] [CrossRef]
- Sadat, L.; Cakir-Kiefer, C.; N’Negue, M.A.; Gaillard, J.L.; Girardet, J.M.; Miclo, L. Isolation and identification of antioxidative peptides from bovine α-lactalbumin. Int. Dairy J. 2011, 21, 214–221. [Google Scholar] [CrossRef]
- Izumi, H.; Ishizuka, S.; Inafune, A.; Hira, T.; Ozawa, K.; Shimizu, T.; Takase, M.; Hara, H. α-lactalbumin hydrolysate stimulates glucagon-Like peptide-2 secretion and small intestinal growth in suckling rats. J. Nutr. 2009, 139, 1322–1327. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Tang, Y.H.; Yu, X.; Xu, Y.Y.; Liu, P.Y.; Xiao, L.; Liu, L.G.; Deng, Q.C.; Yao, P. Flaxseed lignans alleviate high fat diet-induced hepatic steatosis and insulin resistance in mice: Potential involvement of AMP-activated protein kinase. J. Funct. Foods 2016, 24, 482–491. [Google Scholar] [CrossRef]
- Gu, J.F.; Zheng, Z.Y.; Yuan, J.R.; Zhao, B.J.; Wang, C.F.; Zhang, L.; Xu, Q.Y.; Yin, G.W.; Feng, L.; Jia, X.B. Comparison on hypoglycemic and antioxidant activities of the fresh and dried Portulaca oleracea L. in insulin-resistant HepG2 cells and streptozotocin-induced C57BL/6J diabetic mice. J. Ethnopharmacol. 2015, 161, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.H.; Kim, M.Y.; Park, J.M.; Kim, T.H.; Ahn, Y.H. Txnip contributes to impaired glucose tolerance by upregulating the expression of genes involved in hepatic gluconeogenesis in mice. Diabetologia 2013, 56, 2723–2732. [Google Scholar] [CrossRef] [PubMed]
- Healy, N.P.; Kirwan, A.M.; McArdle, M.A.; Holohan, K.; Nongonierma, A.B.; Keane, D.; Kelly, S.; Celkova, L.; Lyons, C.L.; McGillicuddy, F.C.; et al. A casein hydrolysate protects mice against high fat diet induced hyperglycemia by attenuating NLRP3 inflammasome-mediated inflammation and improving insulin signaling. Mol. Nutr. Food Res. 2016, 60, 2421–2432. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Takai, S. Chronic administration of bovine milk-derived α-lactalbumin improves glucose tolerance via enhancement of adiponectin in Goto-Kakizaki rats with type 2 diabetes. Biol. Pharm. Bull. 2014, 37, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; John, C.; Schaltenberg, N.; Berbee, J.; Worthmann, A.; Cherradi, M.L.; Schlein, C.; Piepenburg, J.; Boon, M.R.; Rinninger, F.; et al. Thermogenic adipocytes promote HDL turnover and reverse cholesterol transport. Nat. Commun. 2017, 8, 15010. [Google Scholar] [CrossRef] [PubMed]
- Lindegaard, B.; Matthews, V.B.; Brandt, C.; Hojman, P.; Allen, T.L.; Estevez, E.; Watt, M.J.; Bruce, C.R.; Mortensen, O.H.; Syberg, S.; et al. Interleukin-18 activates skeletal muscle AMPK and reduces weight gain and insulin resistance in mice. Diabetes 2013, 62, 3064–3074. [Google Scholar] [CrossRef] [PubMed]
- Worthmann, A.; John, C.; Ruhlemann, M.C.; Baguhl, M.; Heinsen, F.A.; Schaltenberg, N.; Heine, M.; Schlein, C.; Evangelakos, I.; Mineo, C.; et al. Cold-induced conversion of cholesterol to bile acids in mice shapes the gut microbiome and promotes adaptive thermogenesis. Nat. Med. 2017, 23, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Han, L.; Qin, R.R.; Zhang, Y.Y.; Wang, D.; Wang, Z.H.; Tang, M.X.; Zhang, Y.; Zhong, M.; Zhang, W. Overexpressing STAMP2 attenuates adipose tissue angiogenesis and insulin resistance in diabetic ApoE−/− /LDLR−/− mouse via a PPAR gamma/CD36 pathway. J. Cell. Mol. Med. 2017, 21, 3298–3308. [Google Scholar] [CrossRef] [PubMed]
- Gastaldelli, A.; Gaggini, M.; Defronzo, R.A. Role of adipose tissue insulin resistance in the natural history of T2DM: Results from the San Antonio metabolism study. Diabetes 2017, 66, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux. J. Clin. Invest. 2016, 126, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Chen, M.H.; Zhou, Z.H.; Lu, Y.W.; Wen, X.D.; Yang, J.E. Triterpenoids-enriched extract from the aerial parts of Salvia miltiorrhiza regulates macrophage polarization and ameliorates insulin resistance in high-fat fed mice. Phytother. Res. 2017, 31, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Reyes, J.A.; Arellano-Plancarte, A.; Castillo-Hernandez, J.R. Angiotensin II and the development of insulin resistance: Implications for diabetes. Mol. Cell. Endocrinol. 2009, 302, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vasc. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Sesti, G. Pathophysiology of insulin resistance. Best Pract. Res. 2006, 20, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Boura-Halfon, S.; Zick, Y. Phosphorylation of IRS proteins, insulin action, and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E581–E591. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Leng, J.; Li, J.J.; Tang, J.F.; Li, Y.; Liu, B.L.; Wen, X.D. Corosolic acid inhibits adipose tissue inflammation and ameliorates insulin resistance via AMPK activation in high-fat fed mice. Phytomedcine 2016, 23, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Invest. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.Y.; Barnes, G.T.; Yang, Q.; Tan, Q.; Yang, D.S.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Mulder, P.; van den Hoek, A.M.; Kleemann, R. The CCR2 inhibitor propagermanium attenuates diet-induced insulin resistance, adipose tissue inflammation and non- alcoholic steatohepatitis. PLoS ONE 2017, 12, e0169740. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Peiffer, C.; Whitehead, J. IRS-1 regulation in health and disease. IUNMB Life 2003, 55, 367–374. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| TNF-α | Forward prime | CATCTTCTCAAAATTCGAGTGACAA |
| Reverse prime | TGGGAGTAGACAAGGTACAACCC | |
| IL-6 | Forward prime | ATGGATGCTACCAAACTGGAT |
| Reverse prime | TGAAGGACTCTCTGGCTTTGTCT | |
| MCP-1 | Forward prime | GCCCCACTCACCTGCTGCTACT |
| Reverse prime | CCTGCTGCTGGTGATCCTCTTGT | |
| GAPDH | Forward prime | TGGCAAAGTGGAGATTGTTGC |
| Reverse prime | AAGATGGTGATGGGCTTCCCG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Song, J.; Du, M.; Mao, X. Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorate Adipose Insulin Resistance and Inflammation in High-Fat Diet-Fed C57BL/6J Mice. Nutrients 2018, 10, 242. https://doi.org/10.3390/nu10020242
Gao J, Song J, Du M, Mao X. Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorate Adipose Insulin Resistance and Inflammation in High-Fat Diet-Fed C57BL/6J Mice. Nutrients. 2018; 10(2):242. https://doi.org/10.3390/nu10020242
Chicago/Turabian StyleGao, Jing, Jiajia Song, Min Du, and Xueying Mao. 2018. "Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorate Adipose Insulin Resistance and Inflammation in High-Fat Diet-Fed C57BL/6J Mice" Nutrients 10, no. 2: 242. https://doi.org/10.3390/nu10020242
APA StyleGao, J., Song, J., Du, M., & Mao, X. (2018). Bovine α-Lactalbumin Hydrolysates (α-LAH) Ameliorate Adipose Insulin Resistance and Inflammation in High-Fat Diet-Fed C57BL/6J Mice. Nutrients, 10(2), 242. https://doi.org/10.3390/nu10020242