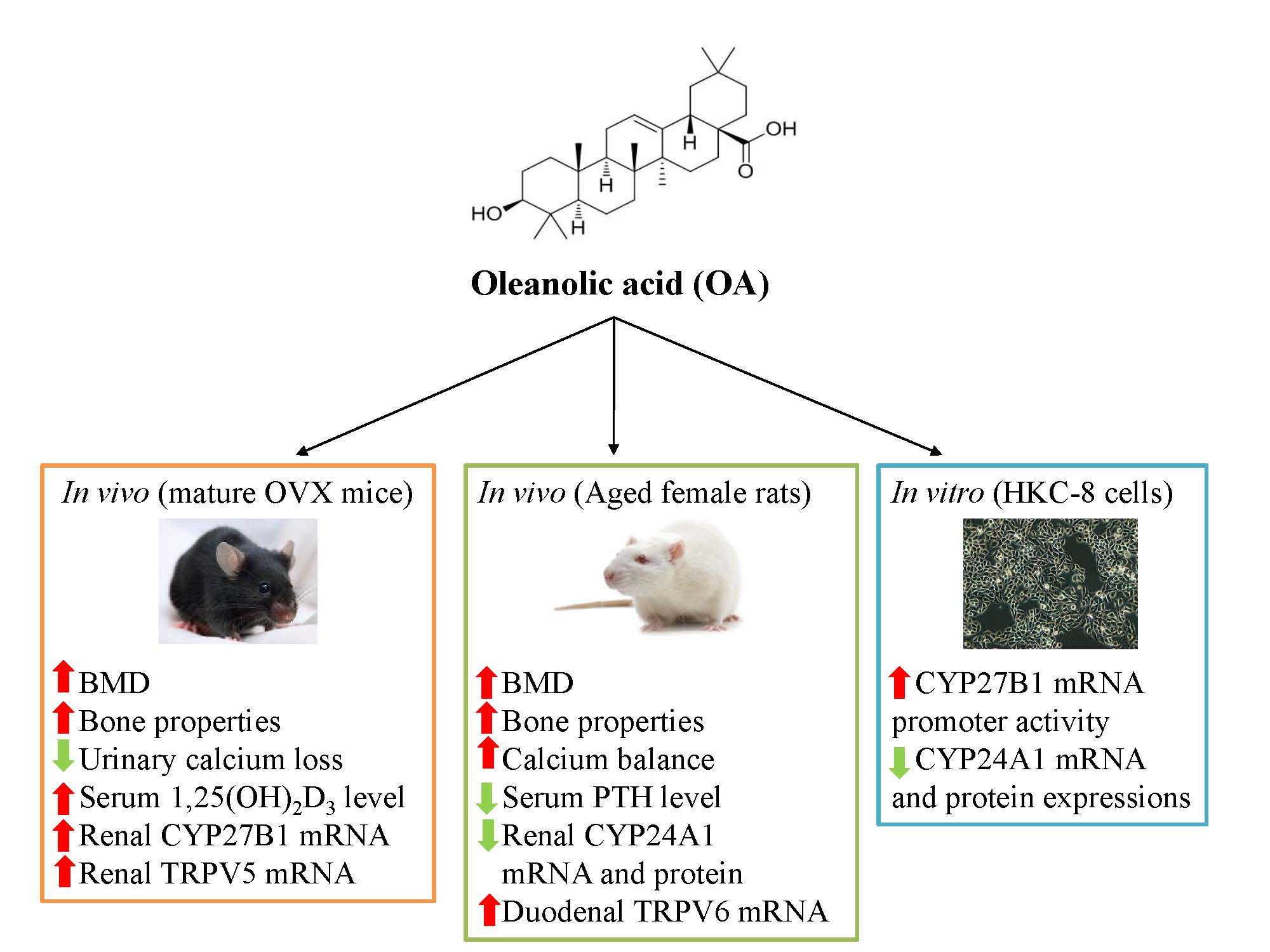
Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animal Study Design
2.1.1. Experiment 1: Bone Protective Effect of OA in OVX Mouse Model
2.1.2. Experiment 2: Effect of OA on Bone and Calcium Balance in an Aged Female Rat Model
2.2. Biochemical Assays of Serum and Urine Samples
2.3. Microcomputed Tomography (μCT)
2.4. Bone Calcium Content
2.5. Calcium Balance Study
2.6. Cell Culture Study
2.7. Real-Time Polymerase Chain Reaction (PCR) Analysis
2.8. Western Blot Analysis
2.9. Transient Transfection
2.10. Statistical Analysis
3. Results
3.1. Effect of OA on Body Weight, Uterus Index and Biochemical Parameters in OVX Mice
3.2. Effect of OA on Bone Markers, Bone Calcium Content and Bone Micro-Architecture in OVX Mice
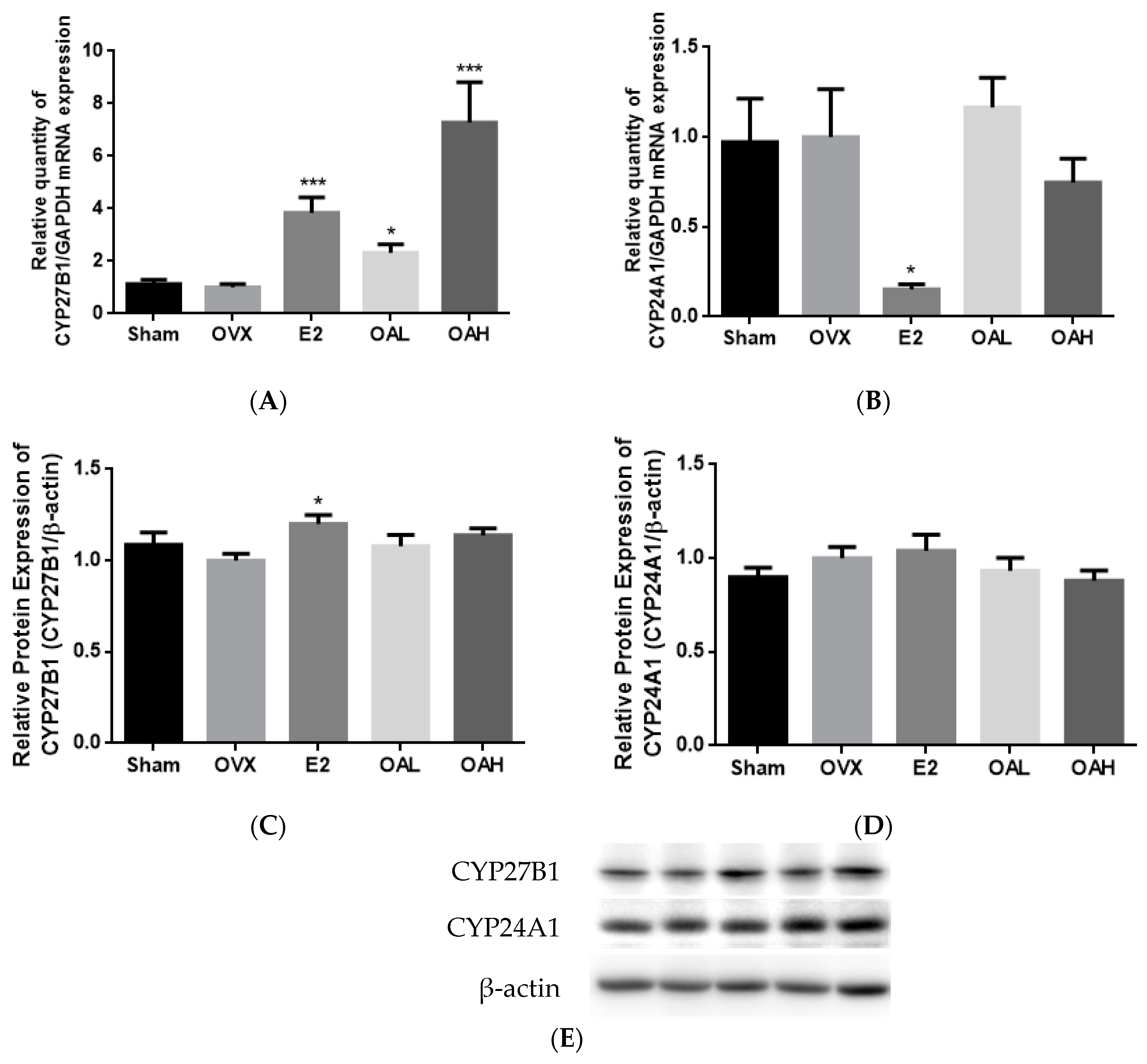
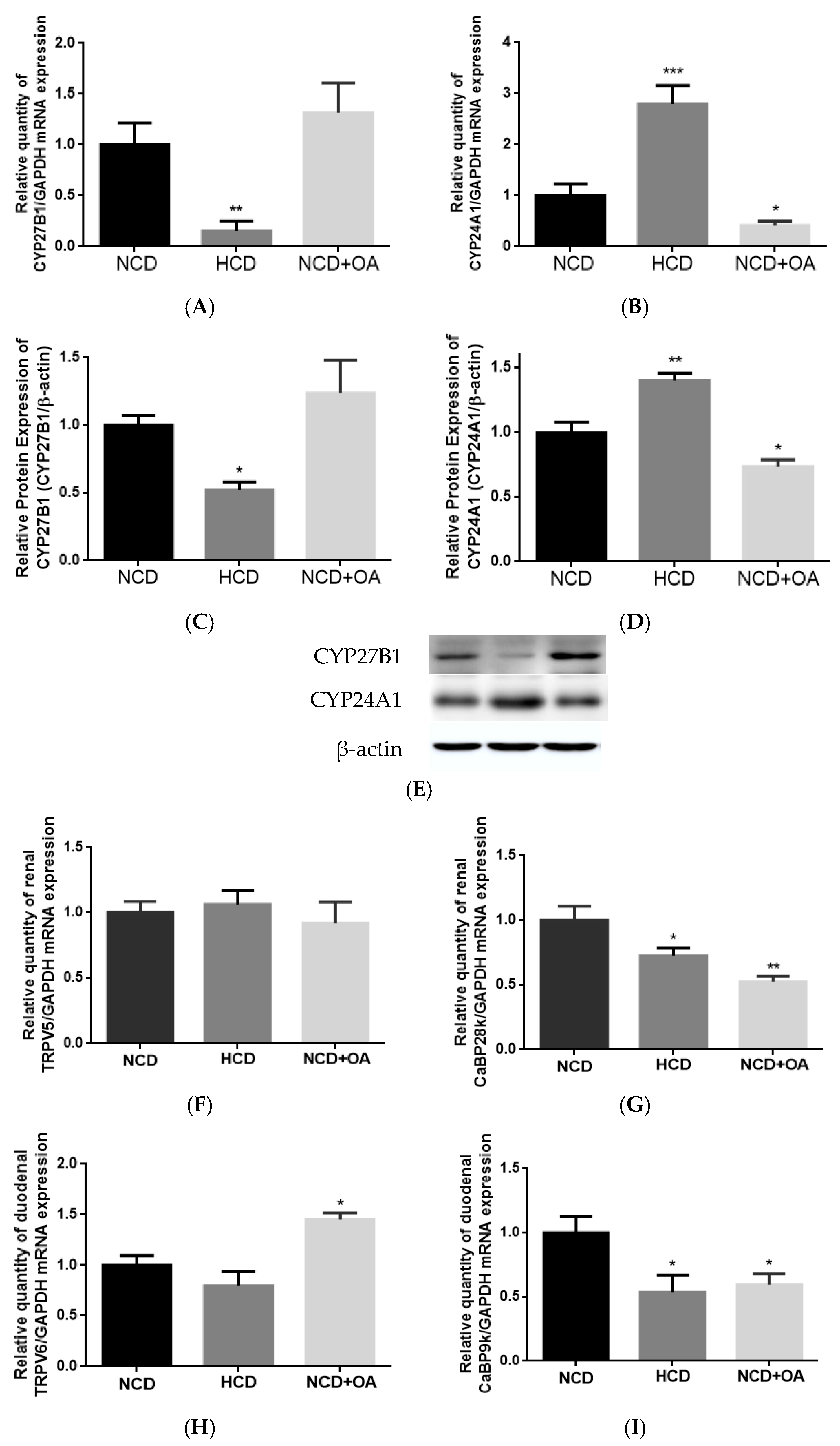
3.3. Effect of OA on Renal CYP27B1 and CYP24A1 mRNA and Protein Expression in OVX Mice
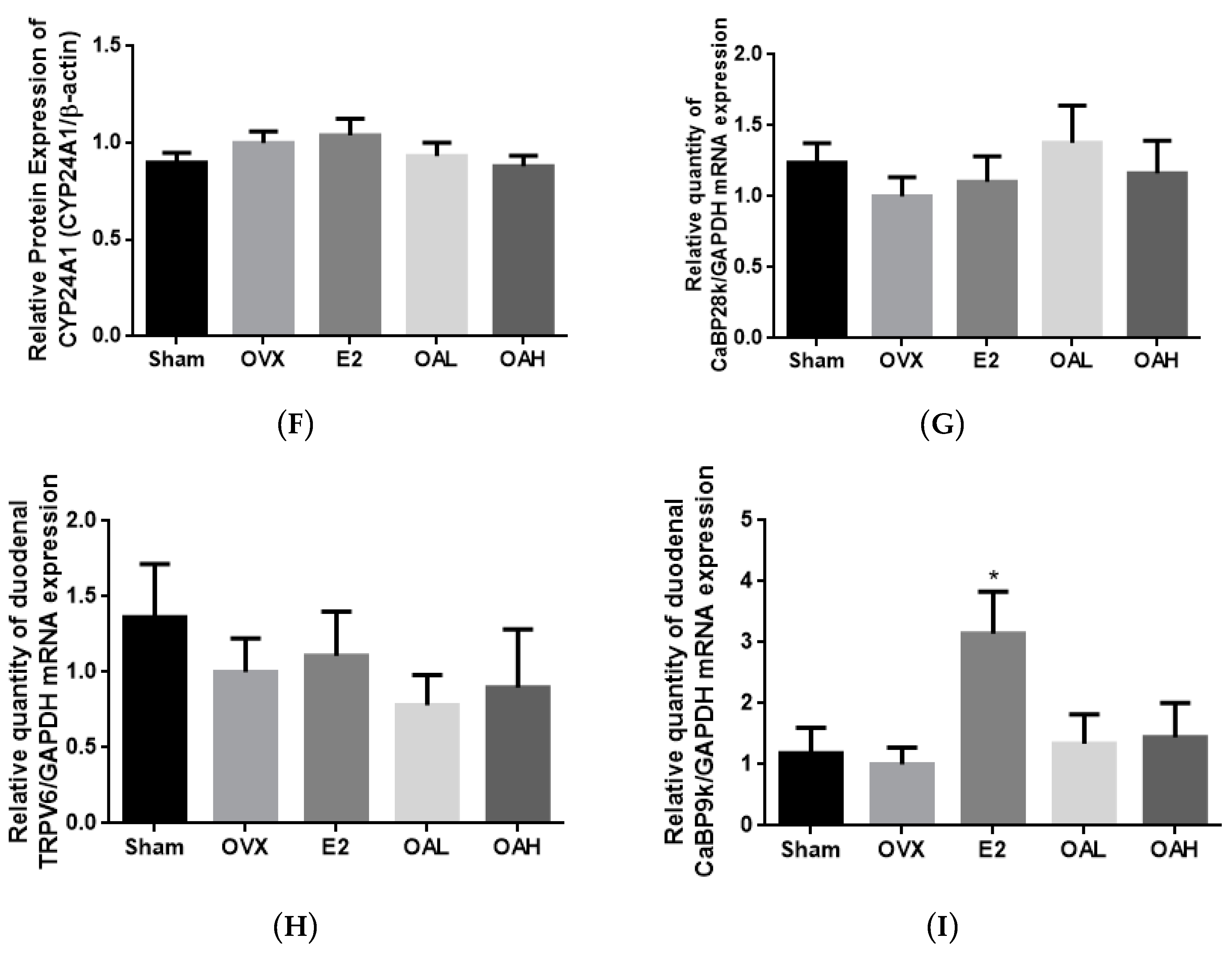
3.4. Effect of OA on Renal and Duodenal mRNA Expression for Calcium Transport in OVX Mice
3.5. Effect of OA on Body Weight and Biochemical Parameters in Aged Rats
3.6. Effect of OA on Calcium Balance in Aged Rats
3.7. Effect of OA on Bone Calcium Content and Bone Micro-Architecture in Aged Rats
3.8. Effect of OA on Renal CYP27B1 and CYP24A1 mRNA and Protein Expression in Aged Female Rats
3.9. Effect of OA on Renal and Duodenal mRNA Expression for Calcium Transport in Aged Rats
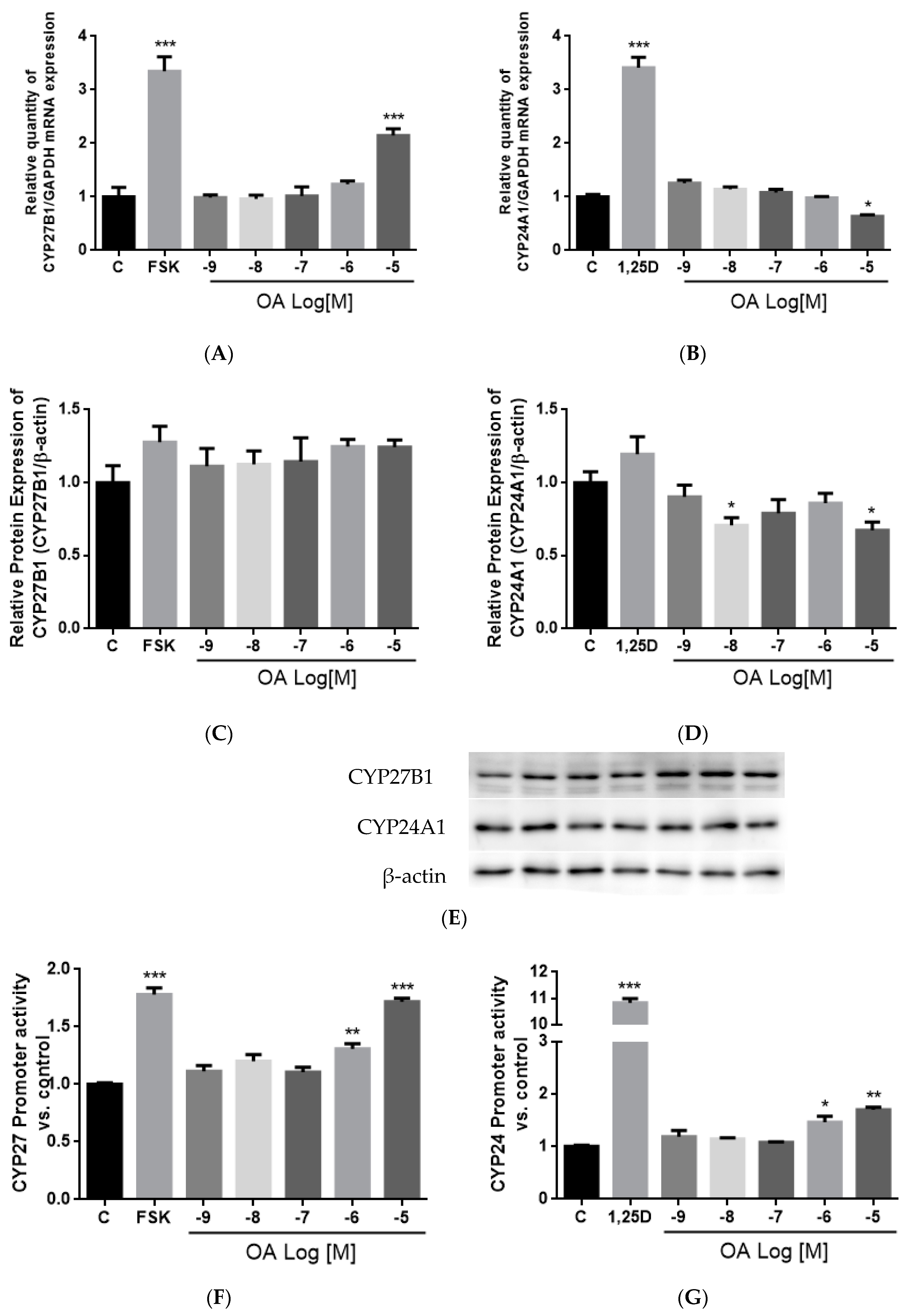
3.10. Effect of OA on CYP27B1 and CYP24A1 mRNA, Protein Expressions and Promoter Activities in HKC-8 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Drake, M.T.; Clarke, B.L.; Lewiecki, E.M. The Pathophysiology and Treatment of Osteoporosis. Clin. Ther. 2015, 37, 1837–1850. [Google Scholar] [CrossRef] [PubMed]
- Veldurthy, V.; Wei, R.; Oz, L.; Dhawan, P.; Jeon, Y.H.; Christakos, S. Vitamin D, calcium homeostasis and aging. Bone Res. 2016, 4, 16041. [Google Scholar] [CrossRef] [PubMed]
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Nordin, B. Calcium and osteoporosis. Nutrition 1997, 13, 664–686. [Google Scholar] [CrossRef]
- Halloran, B.P.; Portale, A.A. Vitamin D metabolism and aging. In Vitamin D, 2nd ed.; Feldman, D., Pike, J.W., Glorieux, F., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 823–838. [Google Scholar]
- Riggs, B.L. Role of the vitamin D-endocrine system in the pathophysiology of postmenopausal osteoporosis. J. Cell. Biochem. 2003, 88, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.M.; Eslick, G.D.; Nowson, C.; Smith, C.; Bensoussan, A. Use of calcium or calcium in combination with vitamin D supplementation to prevent fractures and bone loss in people aged 50 years and older: A meta-analysis. Lancet 2007, 370, 657–666. [Google Scholar] [CrossRef]
- Bolland, M.J.; Barber, P.A.; Doughty, R.N.; Mason, B.; Horne, A.; Ames, R.; Gamble, G.D.; Grey, A.; Reid, I.R. Vascular events in healthy older women receiving calcium supplementation: Randomised controlled trial. BMJ 2008, 336, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Tang, A.M.; Fu, Z.; Wang, D.D.; Newberry, S.J. Calcium Intake and Cardiovascular Disease Risk: An Updated Systematic Review and Meta-analysis. Ann. Intern. Med. 2016, 165, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Oudshoorn, C.; van der Cammen, T.J.; McMurdo, M.E.; van Leeuwen, J.P.; Colin, E.M. Ageing and vitamin D deficiency: Effects on calcium homeostasis and considerations for vitamin D supplementation. Br. J. Nutr. 2009, 101, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Warensjo, E.; Byberg, L.; Melhus, H.; Gedeborg, R.; Mallmin, H.; Wolk, A.; Michaelsson, K. Dietary calcium intake and risk of fracture and osteoporosis: Prospective longitudinal cohort study. BMJ 2011, 342, d1473. [Google Scholar] [CrossRef] [PubMed]
- Avenell, A.; Mak, J.C.; O’Connell, D. Vitamin D and vitamin D analogues for preventing fractures in post-menopausal women and older men. Cochrane Database Syst. Rev. 2014, 14, CD000227. [Google Scholar] [CrossRef] [PubMed]
- Bolland, M.J.; Leung, W.; Tai, V.; Bastin, S.; Gamble, G.D.; Grey, A.; Reid, I.R. Calcium intake and risk of fracture: Systematic review. BMJ 2015, 351, h4580. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.G.; Zeng, X.T.; Wang, J.; Liu, L. Association Between Calcium or Vitamin D Supplementation and Fracture Incidence in Community-Dwelling Older Adults: A Systematic Review and Meta-analysis. JAMA 2017, 318, 2466–2482. [Google Scholar] [CrossRef] [PubMed]
- Pollier, J.; Goossens, A. Oleanolic acid. Phytochemistry 2012, 77, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, E.O.; Seki, H.; Ohyama, K.; Ono, E.; Umemoto, N.; Mizutani, M.; Saito, K.; Muranaka, T. CYP716A subfamily members are multifunctional oxidases in triterpenoid biosynthesis. Plant Cell Physiol. 2011, 52, 2050–2061. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Hareyama, T.; Tezuka, Y.; Zhang, Y.; Miyahara, T.; Kadota, S. Five new oleanolic acid glycosides from Achyranthes bidentata with inhibitory activity on osteoclast formation. Planta Med. 2005, 71, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Zhao, J.; Wang, S.; Pan, Y.; Tanaka, K.; Kadota, S. Synthesis and activity of oleanolic acid derivatives, a novel class of inhibitors of osteoclast formation. Bioorg. Med. Chem. Lett. 2005, 15, 1629–1632. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Zhao, Y.; Cai, M.M.; Li, X.F.; Li, J.X. Synthesis and evaluation of a novel series of heterocyclic oleanolic acid derivatives with anti-osteoclast formation activity. Eur. J. Med. Chem. 2009, 44, 2796–2806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huai, Y.; Jin, J.; Geng, M.; Li, J.-X. Quinoxaline derivative of oleanolic acid inhibits osteoclastic bone resorption and prevents ovariectomy-induced bone loss. Menopause 2011, 18, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Liu, S.F.; Huang, J.H.; Yang, Z.; Tang, D.Z.; Zhou, Q.; Ning, Y.; Zhao, Y.J.; Lu, S.; Shen, Z.Y.; et al. Oleanolic acid exerts an osteoprotective effect in ovariectomy-induced osteoporotic rats and stimulates the osteoblastic differentiation of bone mesenchymal stem cells in vitro. Menopause 2012, 19, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Cheon, Y.H.; Oh, H.M.; Rho, M.C.; Erkhembaatar, M.; Kim, M.S.; Lee, C.H.; Kim, J.J.; Choi, M.K.; Yoon, K.H.; et al. Oleanolic acid acetate inhibits osteoclast differentiation by downregulating PLCgamma2-Ca(2+)-NFATc1 signaling, and suppresses bone loss in mice. Bone 2014, 60, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lyu, Y.; Chen, X.; Zhu, X.; Feng, J.; Xu, Y. Fructus Ligustri Lucidi ethanol extract inhibits osteoclastogenesis in RAW264.7 cells via the RANKL signaling pathway. Mol. Med. Rep. 2016, 14, 4767–4774. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Zhao, Y.; Wang, Y.; Wang, G.; Shang, X.; Britt, M.; Olmedo, M.; Chelly, M.; Morandi, M.M.; Barton, S.; et al. Oleanolic Acid Enhances Mesenchymal Stromal Cell Osteogenic Potential by Inhibition of Notch Signaling. Sci. Rep. 2017, 7, 7002. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lai, W.P.; Leung, P.C.; Wu, C.F.; Yao, X.S.; Wong, M.S. Effects of Fructus Ligustri Lucidi extract on bone turnover and calcium balance in ovariectomized rats. Biol. Pharm. Bull. 2006, 29, 291–296. [Google Scholar] [CrossRef] [PubMed]
- China Pharmacopoeia Committee. Chinese Pharmacopoeia; China Medica Science Press: Beijing, China, 2010; p. 276. [Google Scholar]
- Zhang, Y.; Lai, W.P.; Leung, P.C.; Che, C.T.; Wong, M.S. Improvement of Ca balance by Fructus Ligustri Lucidi extract in aged female rats. Osteoporos. Int. 2008, 19, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, X.L.; Leung, P.C.; Che, C.T.; Wong, M.S. Fructus ligustri lucidi extract improves calcium balance and modulates the calciotropic hormone level and vitamin D-dependent gene expression in aged ovariectomized rats. Menopause 2008, 15, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Leung, P.C.; Che, C.T.; Chow, H.K.; Wu, C.F.; Wong, M.S. Improvement of bone properties and enhancement of mineralization by ethanol extract of Fructus Ligustri Lucidi. Br. J. Nutr. 2008, 99, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.L.; Zhang, Y.; Favus, M.J.; Che, C.T.; Wong, M.S. Ethanol extract of Fructus Ligustri Lucidi increases circulating 1,25-dihydroxyvitamin D3 by inducing renal 25-hydroxyvitamin D-1alpha hydroxylase activity. Menopause 2010, 17, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.L.; Zhao, M.; Wong, K.K.; Che, C.T.; Wong, M.S. Improvement of calcium balance by Fructus Ligustri Lucidi extract in mature female rats was associated with the induction of serum parathyroid hormone levels. Br. J. Nutr. 2012, 108, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Lyu, Y.; Wu, Z.; Fang, Y.; Xu, H.; Zhao, P.; Xu, Y.; Feng, H. Fructus ligustri lucidi ethanol extract improves bone mineral density and properties through modulating calcium absorption-related gene expression in kidney and duodenum of growing rats. Calcif. Tissue Int. 2014, 94, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Feng, X.; Zhao, P.; Wu, Z.; Xu, H.; Fang, Y.; Hou, Y.; Denney, L.; Xu, Y.; Feng, H. Fructus Ligustri Lucidi (FLL) ethanol extract increases bone mineral density and improves bone properties in growing female rats. J. Bone Miner. Metab. 2014, 32, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Fleet, J.C.; Schoch, R.D. Molecular mechanisms for regulation of intestinal calcium absorption by vitamin D and other factors. Crit. Rev. Clin. Lab. Sci. 2010, 47, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Murayama, A.; Takeyama, K.; Kitanaka, S.; Kodera, Y.; Kawaguchi, Y.; Hosoya, T.; Kato, S. Positive and negative regulations of the renal 25-hydroxyvitamin D3 1alpha-hydroxylase gene by parathyroid hormone, calcitonin, and 1alpha,25(OH)2D3 in intact animals. Endocrinology 1999, 140, 2224–2231. [Google Scholar] [CrossRef] [PubMed]
- Takeyama, K.; Kitanaka, S.; Sato, T.; Kobori, M.; Yanagisawa, J.; Kato, S. 25-Hydroxyvitamin D3 1alpha-hydroxylase and vitamin D synthesis. Science 1997, 277, 1827–1830. [Google Scholar] [CrossRef] [PubMed]
- Zierold, C.; Mings, J.A.; DeLuca, H.F. Regulation of 25-hydroxyvitamin D3-24-hydroxylase mRNA by 1,25-dihydroxyvitamin D3 and parathyroid hormone. J. Cell. Biochem. 2003, 88, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, J.; DeLuca, H.F. The vitamin D receptor in the proximal renal tubule is a key regulator of serum 1alpha,25-dihydroxyvitamin D(3). Am. J. Physiol. Endocrinol. Metab. 2015, 308, E201–E205. [Google Scholar] [CrossRef] [PubMed]
- Ranch, D.; Zhang, M.Y.; Portale, A.A.; Perwad, F. Fibroblast growth factor 23 regulates renal 1,25-dihydroxyvitamin D and phosphate metabolism via the MAP kinase signaling pathway in Hyp mice. J. Bone Miner. Res. 2011, 26, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Sriussadaporn, S.; Tembe, V.A.; Favus, M.J. Insulin-like growth factor I increases renal 1,25 (OH) 2D3 biosynthesis during low-P diet in adult rats. Am. J. Physiol. 1997, 41, F698–F703. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.S.; Tembe, V.A.; Favus, M.J. Insulin-like growth factor-I stimulates renal 1,25-dihydroxycholecalciferol synthesis in old rats fed a low calcium diet. J. Nutr. 2000, 130, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Hasegawa, H.; Yamazaki, Y.; Muto, T.; Hino, R.; Takeuchi, Y.; Fujita, T.; Nakahara, K.; Fukumoto, S.; Yamashita, T. FGF-23 is a potent regulator of vitamin D metabolism and phosphate homeostasis. J. Bone Miner. Res. 2004, 19, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Racusen, L.C.; Monteil, C.; Sgrignoli, A.; Lucskay, M.; Marouillat, S.; Rhim, J.G.; Morin, J.-P. Cell lines with extended in vitro growth potential from human renal proximal tubule: Characterization, response to inducers, and comparison with established cell lines. J. Lab. Clin. Med. 1997, 129, 318–329. [Google Scholar] [CrossRef]
- Chanakul, A.; Zhang, M.Y.; Louw, A.; Armbrecht, H.J.; Miller, W.L.; Portale, A.A.; Perwad, F. FGF-23 regulates CYP27B1 transcription in the kidney and in extra-renal tissues. PLoS ONE 2013, 8, e72816. [Google Scholar] [CrossRef] [PubMed]
- Kerry, D.M.; Dwivedi, P.P.; Hahn, C.N.; Morris, H.A.; Omdahl, J.L.; May, B.K. Transcriptional synergism between vitamin D-responsive elements in the rat 25-hydroxyvitamin D3 24-hydroxylase (CYP24) promoter. J. Biol. Chem. 1996, 271, 29715–29721. [Google Scholar] [CrossRef] [PubMed]
- Kemmis, C.M.; Salvador, S.M.; Smith, K.M.; Welsh, J. Human mammary epithelial cells express CYP27B1 and are growth inhibited by 25-hydroxyvitamin D-3, the major circulating form of vitamin D-3. J. Nutr. 2006, 136, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.; Khan, K.; Tyagi, A.M.; Khan, M.P.; Yadav, D.K.; Trivedi, R.; Maurya, R.; Singh, D.; Chattopadhyay, N. Greater Skeletal Gains in Ovary Intact Rats at Maturity Are Achieved by Supplementing a Standardized Extract of Butea monosperma Stem Bark that Confers Better Bone Conserving Effect following Ovariectomy and Concurrent Treatment Withdrawal. Evid. Based Complement. Altern. Med. 2013, 2013, 519387. [Google Scholar] [CrossRef] [PubMed]
- Haguenauer, D.; Welch, V.; Shea, B.; Tugwell, P.; Adachi, J.D.; Wells, G. Fluoride for the treatment of postmenopausal osteoporotic fractures: A meta-analysis. Osteoporos. Int. 2000, 11, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, K.; Byrjalsen, I.; Andersen, J.R.; Bihlet, A.R.; Russo, L.A.; Alexandersen, P.; Valter, I.; Qvist, P.; Lau, E.; Riis, B.J.; et al. A randomized, double-blind, multicenter, placebo-controlled study to evaluate the efficacy and safety of oral salmon calcitonin in the treatment of osteoporosis in postmenopausal women taking calcium and vitamin D. Bone 2016, 91, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.; Dardenne, O.; Van Abel, M.; Van Der Kemp, A.W.; Van Os, C.H.; St-Arnaud, R.; Bindels, R.J. Modulation of renal Ca2+ transport protein genes by dietary Ca2+ and 1,25-dihydroxyvitamin D3 in 25-hydroxyvitamin D3-1alpha-hydroxylase knockout mice. FASEB J. 2002, 16, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Van Abel, M.; Hoenderop, J.G.; Dardenne, O.; St Arnaud, R.; Van Os, C.H.; Van Leeuwen, H.J.; Bindels, R.J. 1,25-dihydroxyvitamin D(3)-independent stimulatory effect of estrogen on the expression of ECaC1 in the kidney. J. Am. Soc. Nephrol. 2002, 13, 2102–2109. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shan, A.; Liu, T.; Zhang, C.; Zhang, Z. In vitro immunomodulatory effects of an oleanolic acid-enriched extract of Ligustrum lucidum fruit (Ligustrum lucidum supercritical CO2 extract) on piglet immunocytes. Int. Immunopharmacol. 2012, 14, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Qiu, Z.; Wong, M.S. The Hong Kong Polytechnic Universtiy, Hong Kong, China. Unpublished work. 2017. [Google Scholar]
- Armbrecht, H.J.; Boltz, M.A.; Bruns, M.E. Effect of age and dietary calcium on intestinal calbindin D-9k expression in the rat. Arch. Biochem. Biophys. 2003, 420, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Colin, E.M.; Van Den Bemd, G.J.; Van Aken, M.; Christakos, S.; De Jonge, H.R.; Deluca, H.F.; Prahl, J.M.; Birkenhager, J.C.; Buurman, C.J.; Pols, H.A.; et al. Evidence for involvement of 17beta-estradiol in intestinal calcium absorption independent of 1,25-dihydroxyvitamin D3 level in the Rat. J. Bone Miner. Res. 1999, 14, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, V.; Dempster, D.W.; Birchman, R.; Xu, R.; Lindsay, R. Loss of cancellous bone mass and connectivity in ovariectomized rats can be restored by combined treatment with parathyroid hormone and estradiol. J. Clin. Investig. 1993, 91, 2479–2487. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Castillo, L.; DeLuca, H.F. Control of renal vitamin D hydroxylases in birds by sex hormones. Proc. Natl. Acad. Sci. USA 1976, 73, 2701–2705. [Google Scholar] [CrossRef] [PubMed]
- Armbrecht, H.J.; Boltz, M.A.; Ritter, C.S.; Brown, A.J. Parathyroid hormone stimulation of the renal 25-hydroxyvitamin D-1alpha-hydroxylase—Effect of age and free radicals. J. Steroid Biochem. Mol. Biol. 2007, 103, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.H.; O’Loughlin, P.D.; May, B.K.; Morris, H.A. Modulation of CYP27B1 and CYP24 mRNA expression in bone is independent of circulating 1,25(OH)2D3 levels. Bone 2005, 36, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Extraskeletal actions of vitamin D. Ann. N. Y. Acad. Sci. 2016, 1376, 29–52. [Google Scholar] [CrossRef] [PubMed]
- Brenza, H.L.; Kimmel-Jehan, C.; Jehan, F.; Shinki, T.; Wakino, S.; Anazawa, H.; Suda, T.; DeLuca, H.F. Parathyroid hormone activation of the 25-hydroxyvitamin D3-1alpha-hydroxylase gene promoter. Proc. Natl. Acad. Sci. USA 1998, 95, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Brenza, H.L.; DeLuca, H.F. Regulation of 25-hydroxyvitamin D3 1α-hydroxylase gene expression by parathyroid hormone and 1,25-dihydroxyvitamin D3. Arch. Biochem. Biophys. 2000, 381, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Zierold, C.; Reinholz, G.G.; Mings, J.A.; Prahl, J.M.; DeLuca, H.F. Regulation of the procine 1,25-dihydroxyvitamin D3-24-hydroxylase (CYP24) by 1,25-dihydroxyvitamin D3 and parathyroid hormone in AOK-B50 cells. Arch. Biochem. Biophys. 2000, 381, 323–327. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | |
|---|---|---|
| Mouse | ||
| GAPDH | F: CAGAACATCATCCCTGCATC | R: CTGCTTCACCACCTTCTTGA |
| CYP27B1 | F: GCATCACTTAACCCACTTCC | R: CGGGAAAGCTCATAGAGTGT |
| CYP24A1 | F: AAGAGATTCGGGCTCCTTCA | R: GCAGGGCTTGACTGATTTGA |
| TRPV5 | F: GAAACTTCTCAATTGGTGGGTCAG | R: TTTGCCGGAAGTCACAGTT |
| Rat | ||
| GAPDH | F: GTGAGGTGACCGCATCTTCT | R: CTTGCCGTGGGTAGAGTCAT |
| CYP27B1 | F: CCATCGAGTCCAACTGCCTT | R: AGGGTCGGCCACATAAACTG |
| CYP24A1 | F: CTCGGACCCTTGACAAACCA | R: CGATGCCGAATGGGAGATGA |
| TRPV6 | F: ATCCGCCGCTATGCACA | R: AGTTTTTTCTCCTGAGTCTTTTTCCA |
| Human | ||
| GAPDH | F: TTGCAACCGGGAAGGAAATG | R: CGCCCAATACGACCAAATCA |
| CYP27B1 | F: CTGCGGAAGGCGAAGAATGG | R: TTGTTCAGGGTTCCGGCGTA |
| CYP24A1 | F: CAAACCGTGGAAGGCCTATC | R: AGTCTTCCCCTTCCAGGATCA |
| Sham | OVX | E2 | OAL | OAH | |
|---|---|---|---|---|---|
| Body weight and uterus index | |||||
| Weight change, % | 1.3 ± 0.7 | 7.3 ± 1.3 ^^^ | −6.6 ± 1.1 *** | −3.9 ± 1.0 *** | −7.4 ± 1.4 *** |
| Uterus index, mg/g | 0.31 ± 0.03 | 0.09 ± 0.02 ^^^ | 0.70 ± 0.08 *** | 0.12 ± 0.02 | 0.16 ± 0.02 |
| Serum chemistry | |||||
| s-Ca, mg/dL | 8.7 ± 0.2 | 8.9 ± 0.2 | 9.2 ± 0.2 | 9.2 ± 0.3 | 9.1 ± 0.3 |
| s-P, mg/dL | 7.6 ± 0.4 | 7.6 ± 0.6 | 6.7 ± 0.6 | 6.9 ± 0.8 | 6.5 ± 0.5 |
| s-OCN, ng/mL | 77.2 ± 1.4 | 86.6 ± 2.9 ^ | 72.0 ± 3.1 ** | 93.5 ± 4.0 | 93.6 ± 4.8 |
| s-1,25D, pg/mL | 63.3 ± 5.1 | 74.6 ± 2.5 | 58.8 ± 6.9 | 83.9 ± 8.5 | 98.7 ± 7.2 * |
| Urine chemistry | |||||
| u-Ca/Cr, mg/mg | 0.21 ± 0.03 | 0.42 ± 0.04 ^^^ | 0.29 ± 0.02 * | 0.16 ± 0.02 *** | 0.14 ± 0.01 *** |
| u-P/Cr, mg/mg | 7.3 ± 0.4 | 7.6 ± 0.6 | 6.5 ± 0.2 | 6.5 ± 0.5 | 6.6 ± 0.3 |
| u-DPD, nmol/mmol | 9.4 ± 0.5 | 14.4 ± 1.2 ^^^ | 8.2 ± 0.6 *** | 11.7 ± 0.8 | 14.8 ± 0.5 |
| Bone calcium content | |||||
| Bone ash weight, mg | 19.7 ± 0.4 | 18.6 ± 0.3 | 21.1 ± 0.4 * | 20.8 ± 0.6 * | 21.0 ± 0.8 * |
| Ca/Ash bone, μg/mg | 377.4 ± 1.6 | 372. 3 ± 2.8 | 387.7 ± 3.9 * | 387.0 ± 2.7 * | 387.3 ± 3.3 * |
| Sham | OVX | E2 | OAL | OAH | |
|---|---|---|---|---|---|
| Proximal tibia | |||||
| BMD, mg HA/cm3 | 207.2 ± 7.7 | 116.7 ± 6.5 ^^^ | 249.4 ± 7.5 *** | 170.5 ± 6.8 *** | 175.4 ± 4.7 *** |
| BV/TV, % | 23.3 ± 1.1 | 11.5 ± 1.0 ^^^ | 27.9 ± 1.4 *** | 18.0 ± 0.8 *** | 19.0 ± 0.7 *** |
| Tb.N, mm−1 | 4.51 ± 0.24 | 3.08 ± 0.37 | 5.38 ± 0.29 *** | 3.76 ± 0.15 | 3.74 ± 0.08 |
| Tb.Th, μm | 51.2 ± 0.8 | 42.9 ± 1.6 ^^^ | 54.0 ± 1.2 *** | 48.7 ± 0.8 ** | 50.7 ± 1.1 *** |
| Tb.Sp, μm | 175.4 ± 11.2 | 312.2 ± 34.7 ^^^ | 126.5 ± 17.3 *** | 225.4 ± 11.1 * | 217.6 ± 6.5 * |
| Conn.D, mm3 | 196.1 ± 15.9 | 105.6 ± 8.3 ^^ | 237.2 ± 27.7 *** | 159.6 ± 10.0 *** | 140.4 ± 8.9 * |
| Distal femur | |||||
| BMD, mg HA/cm3 | 198.4 ± 6.6 | 130.9 ± 9.1 ^^^ | 246.8 ± 8.4 *** | 185.9 ± 4.3 *** | 195.6 ± 7.2 *** |
| BV/TV, % | 16.6 ± 0.9 | 12.0 ± 1.0 ^^ | 25.5 ± 1.3 *** | 18.5 ± 0.8 *** | 20.7 ± 1.1 *** |
| Tb.N, mm−1 | 3.50 ± 0.08 | 2.79 ± 0.18 ^ | 4.24 ± 0.18 *** | 3.57 ± 0.14 * | 3.81 ± 0.21 *** |
| Tb.Th, μm | 50.3 ± 1.5 | 41.5 ± 1.4 ^^^ | 59.4 ± 2.1 *** | 51.9 ± 1.4 *** | 54.2 ± 0.4 *** |
| Tb.Sp, μm | 239.7 ± 8.0 | 329.8 ± 27.6 ^^ | 176.3 ± 11.0 *** | 231.6 ± 10.9 ** | 214.2 ± 15.9 *** |
| Conn.D, mm3 | 141.8 ± 11.5 | 97.4 ± 7.9 ^ | 141.6 ± 10.8 * | 144.5 ± 8.5 * | 149.2 ± 10.7 ** |
| Lumbar vertebra | |||||
| BMD, mg HA/cm3 | 231.8 ± 11.0 | 183.1 ± 7.1 ^^^ | 297.0 ± 15.3 *** | 219.0 ± 8.0 ** | 213.2 ± 4.7 * |
| BV/TV, % | 29.4 ± 1.1 | 21.1 ± 0.6 ^^^ | 37.0 ± 1.7 *** | 25.6 ± 1.3 ** | 25.1 ± 0.8 * |
| Tb.N, mm−1 | 4.62 ± 0.12 | 3.82 ± 0.08 ^^^ | 4.89 ± 0.11 *** | 4.26 ± 0.13 * | 4.33 ± 0.12 ** |
| Tb.Th, μm | 63.5 ± 1.0 | 58.3 ± 1.9 | 75.3 ± 2.0 *** | 59.3 ± 1.5 | 57.0 ± 1.0 |
| Tb.Sp, μm | 153.8 ± 6.3 | 208.8 ± 5.7 ^^^ | 130.0 ± 6.3 *** | 176.1 ± 7.7 ** | 172.9 ± 5.2 ** |
| Conn.D, mm3 | 141.1 ± 7.1 | 100.2 ± 4.9 ^^^ | 139.7 ± 4.3 *** | 140.8 ± 6.5 *** | 140.3 ± 6.6 *** |
| NCD | HCD | NCD + OA | |
|---|---|---|---|
| Body weight | |||
| Weight change, % | −1.3 ± 0.4 | −0.4 ± 0.7 | 0.2 ± 1.6 |
| Serum chemistry | |||
| s-Ca, mg/dL | 11.5 ± 0.4 | 11.5 ± 0.2 | 11.9 ± 0.5 |
| s-P,mg/dL | 5.2 ± 0.4 | 5.9 ± 0.2 | 4.9 ± 0.8 |
| s-1,25D, pg/mL | 17.5 ± 2.5 | 3.5 ± 0.7 *** | 13.4 ± 2.9 |
| s-PTH, pg/mL | 213.8 ± 40.9 | 229.2 ± 49.4 | 138.2 ± 58.0 |
| Urine chemistry | |||
| u-Ca/Cr, mg/mg | 0.30 ± 0.05 | 0.38 ± 0.04 | 0.16 ± 0.03 * |
| u-P/Cr, mg/mg | 2.65 ± 0.43 | 0.12 ± 0.07 *** | 2.11 ± 0.24 |
| Bone calcium content | |||
| Bone ash weight, mg | 306.1 ± 8.5 | 351.6 ± 6.1 ** | 318.2 ± 8.0 |
| Ca/Ash bone, μg/mg | 391.8 ± 2.2 | 403.1 ± 3.6 * | 402.8 ± 3.3 * |
| NCD | HCD | NCD + OA | |
|---|---|---|---|
| Ca intake, mg/day | 92.4 ± 3.9 | 182.1 ± 11.2 | 94.2 ± 4.7 |
| Urine Ca, mg/day | 1.8 ± 0.7 | 5.7 ± 0.6 *** | 1.4 ± 0.2 |
| Fecal Ca, mg/day | 80.2 ± 5.6 | 158.1 ± 19.4 ** | 68.5 ± 5.0 |
| Ca absorption rate, % | 11.2 ± 4.7 | 12.2 ± 10.8 | 23.9 ± 5.9 |
| Net Ca balance, mg/day | 12.3 ± 2.2 | 26.7 ± 4.9 * | 24.9 ± 3.0 * |
| NCD | HCD | NCD + OA | |
|---|---|---|---|
| Proximal tibia | |||
| BMD, mg HA/cm3 | 225.7 ± 21.1 | 330.1 ± 18.0 ** | 312.0 ± 16.9 * |
| BV/TV, % | 34.9 ± 3.5 | 50.0 ± 3.3 * | 46.1 ± 2.8 * |
| Tb.N, mm−1 | 3.19 ± 0.13 | 3.76 ± 0.10 ** | 3.61 ± 0.08 * |
| Tb.Th, μm | 107.0 ± 7.3 | 136.6 ± 7.4 * | 122.9 ± 6.2 |
| Tb.Sp, μm | 205.2 ± 20.3 | 134.6 ± 11.4 ** | 148.4 ± 10.2 * |
| Conn.D, mm3 | 31.4 ± 2.1 | 46.9 ± 2.1 ** | 43.2 ± 2.2 ** |
| Distal femur | |||
| BMD, mg HA/cm3 | 282.1 ± 9.8 | 391.9 ± 19.7 ** | 324.3 ± 10.6 * |
| BV/TV, % | 35.9 ± 3.4 | 57.7 ± 4.2 ** | 47.5 ± 2.1 * |
| Tb.N, mm−1 | 2.95 ± 0.08 | 3.32 ± 0.04 * | 3.31 ± 0.07 * |
| Tb.Th, μm | 132.5 ± 4.6 | 184.3 ± 12.4 * | 147.8 ± 5.1 |
| Tb.Sp, μm | 200.9 ± 0.4 | 128.3 ± 12.4 *** | 156.2 ± 9.3 * |
| Conn.D, mm3 | 23.7 ± 1.8 | 30.1 ± 1.6 * | 31.6 ± 1.4 * |
| Lumbar vertebra | |||
| BMD, mg HA/cm3 | 286.7 ± 13.0 | 407.7 ± 18.5 *** | 378.1 ± 18.0 * |
| BV/TV, % | 31.2 ± 2.2 | 49.7 ± 2.9 ** | 46.1 ± 2.8 * |
| Tb.N, mm−1 | 3.02 ± 0.08 | 3.59 ± 0.08 *** | 3.51 ± 0.07 *** |
| Tb.Th, μm | 102.9 ± 5.8 | 139.6 ± 10.4 * | 123.8 ± 9.3 |
| Tb.Sp, μm | 229.2 ± 12.0 | 139.5 ± 7.0 *** | 155.5 ± 8.0 *** |
| Conn.D, mm3 | 30.4 ± 1.1 | 39.3 ± 1.5 ** | 40.8 ± 1.0 *** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, S.; Dong, X.-L.; Ho, M.-X.; Yu, W.-X.; Wong, K.-C.; Yao, X.-S.; Wong, M.-S. Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism. Nutrients 2018, 10, 247. https://doi.org/10.3390/nu10020247
Cao S, Dong X-L, Ho M-X, Yu W-X, Wong K-C, Yao X-S, Wong M-S. Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism. Nutrients. 2018; 10(2):247. https://doi.org/10.3390/nu10020247
Chicago/Turabian StyleCao, Sisi, Xiao-Li Dong, Ming-Xian Ho, Wen-Xuan Yu, Ka-Chun Wong, Xin-Sheng Yao, and Man-Sau Wong. 2018. "Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism" Nutrients 10, no. 2: 247. https://doi.org/10.3390/nu10020247
APA StyleCao, S., Dong, X. -L., Ho, M. -X., Yu, W. -X., Wong, K. -C., Yao, X. -S., & Wong, M. -S. (2018). Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism. Nutrients, 10(2), 247. https://doi.org/10.3390/nu10020247