Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols

 , ,
, , 

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Aqueous GP Extraction and Total Polyphenols Content Determination
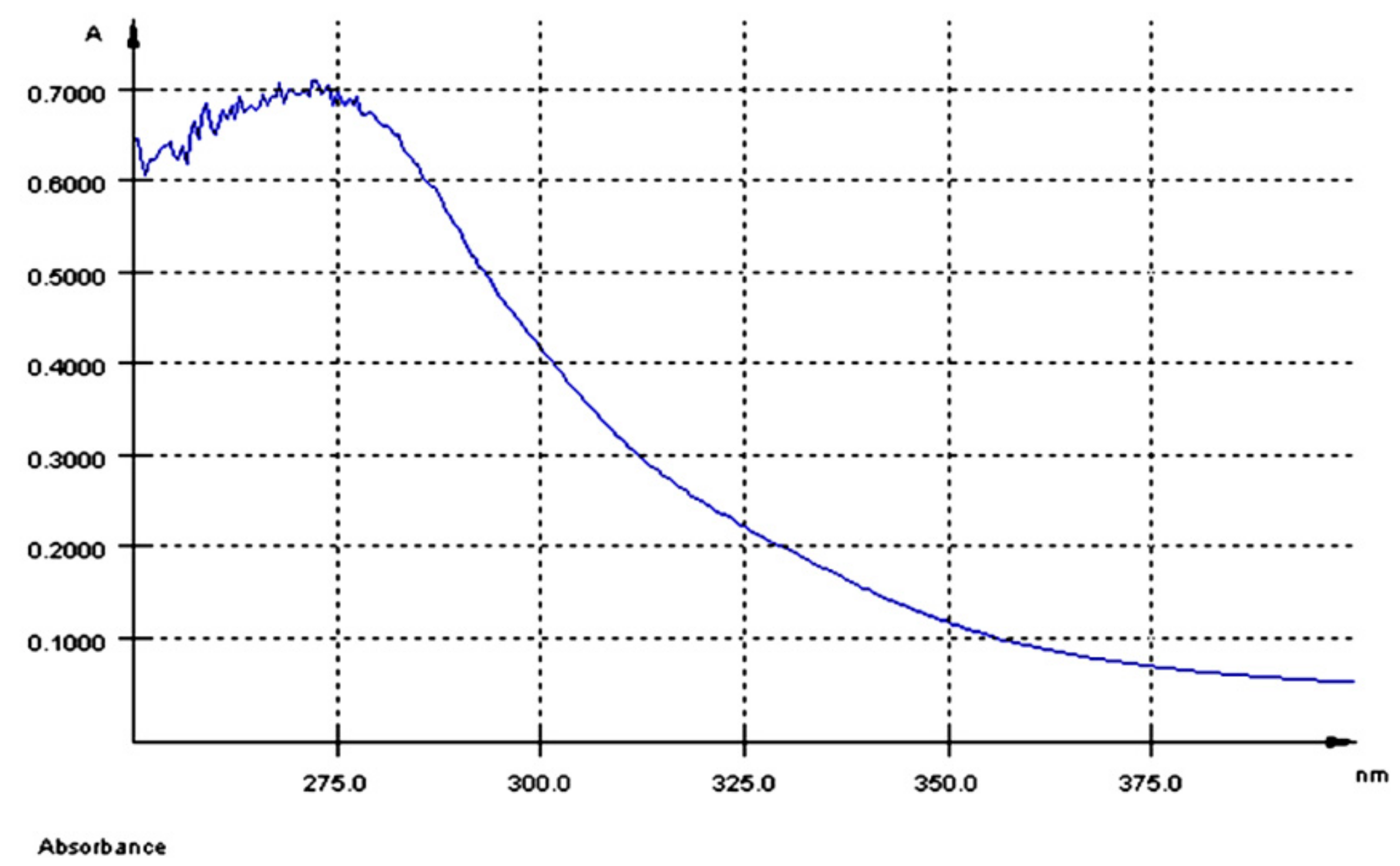
UV-Vis Spectroscopy of AGP Extract
2.2. In Vitro Study
2.2.1. Measurement of Cell Viability (MTT Assay)
2.2.2. Cells Treatment and UV-VIS Spectra Measurement
2.3. In Vivo Study
2.3.1. Animals and Diets
2.3.2. Sample Preparation
2.3.3. Polyphenols Extraction from GP and Experimental Diets (GP− and GP+)
2.3.4. Polyphenols Extraction from Duodenum and Colon Samples
2.3.5. Measurement of Polyphenols Composition in GP, Diets and Gut Samples by High Performance Liquid Chromatography-Photodiode Detector Coupled with Mass Spectroscopy (HPLC-DAD-MS)
2.3.6. Measurement of UV-VIS Spectra from Duodenum and Colon Samples
2.3.7. Determination of the Total Phenolic Content from Duodenum and Colon
2.3.8. Preparation of Tissue (Duodenum and Colon) Lysate
2.3.9. Measurement of Lipid Peroxidation in Duodenum and Colon—TBARS-MDA
2.3.10. Measurement of Total Antioxidant Status (TAS) in Duodenum and Colon
2.3.11. Measurement of Catalase (CAT) Activity in Duodenum and Colon
2.3.12. Measurement of Superoxide Dismutase (SOD) Activity in Duodenum and Colon
2.3.13. Measurement of Glutathione Peroxidase (GPx) Activity in Duodenum and Colon
2.4. Statistical analysis
3. Results
3.1. In Vitro Study
3.1.1. UV-Vis Spectroscopy Analysis and Total Polyphenols Concentration of AGP Extract
3.1.2. Effect of AGP Extract on Cell Viability
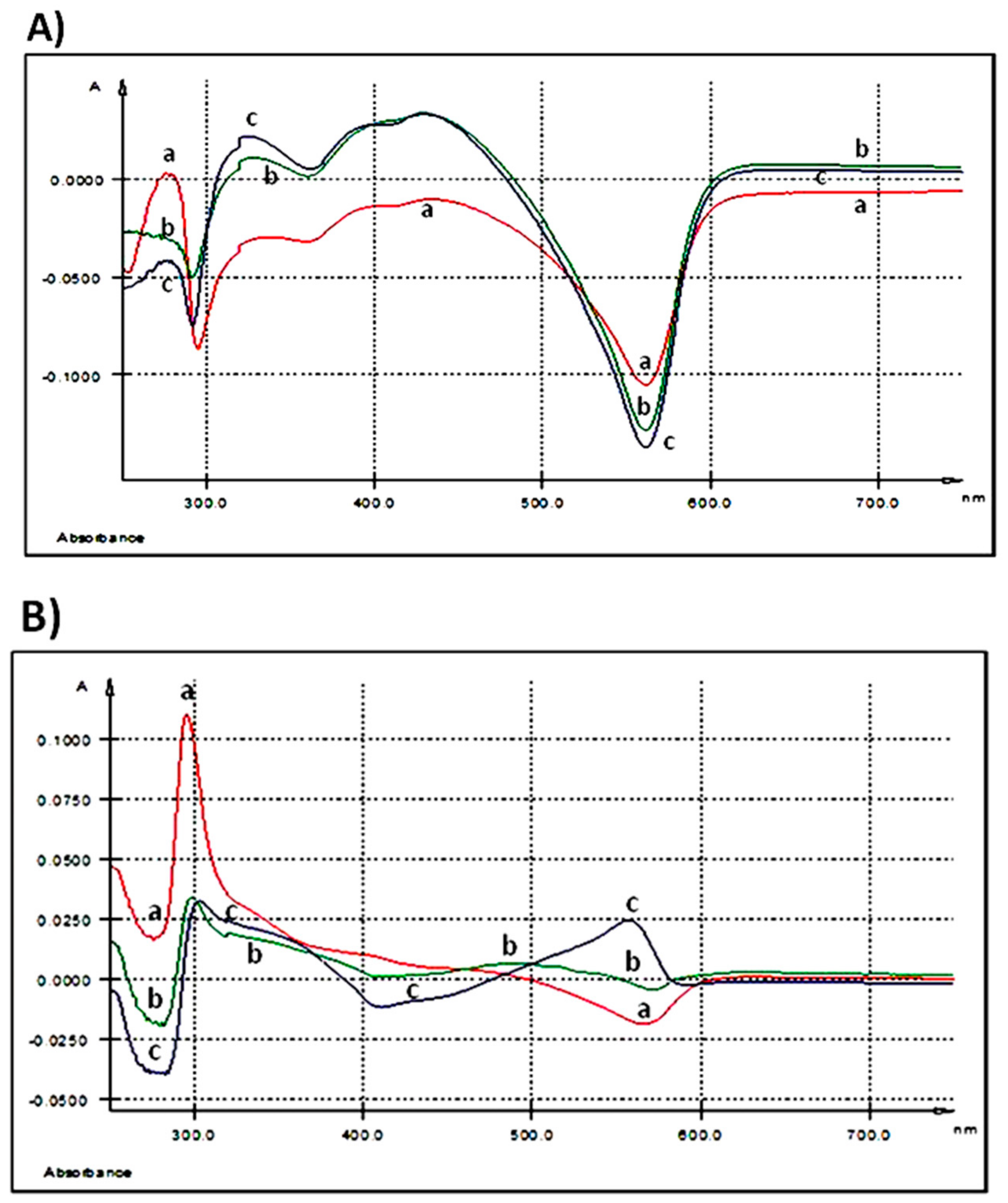
3.1.3. Evaluation of AGP’s Fate in Extracellular Medium
3.1.4. Evaluation of AGP’ Fate in the Cellular Matrix
3.2. In Vivo Study
3.2.1. Polyphenol Content and Profile of GP and Feed Compounds
3.2.2. Effect of Dietary GP on the Growth Performance of Piglets
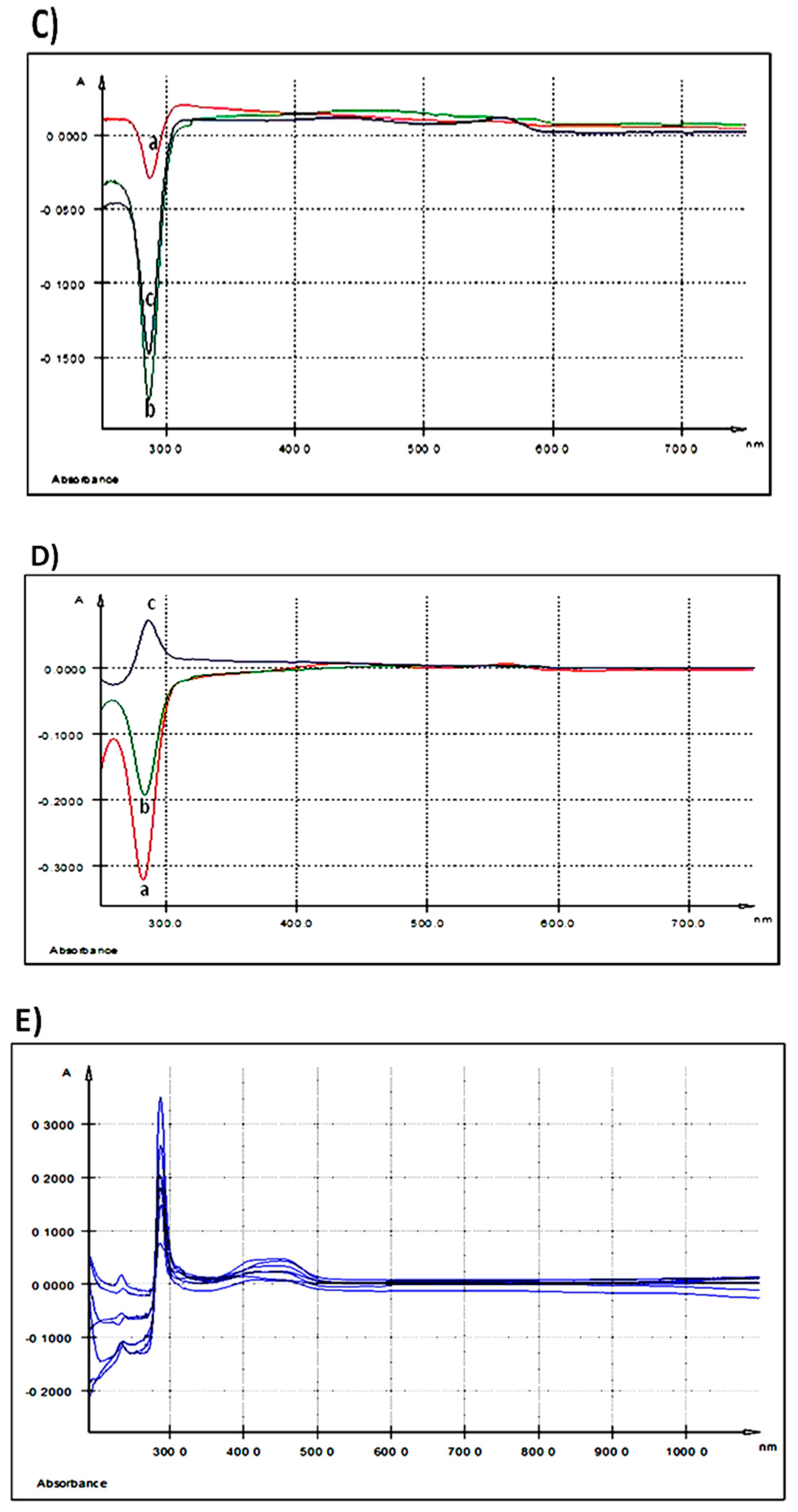
3.2.3. Qualitative Assessment of Polyphenols Absorption in Duodenum and Colon by UV-Vis Spectra Measurement
3.2.4. Quantitative Assessment of Polyphenols Absorption in Duodenum and Colon by LC-MS Measurement
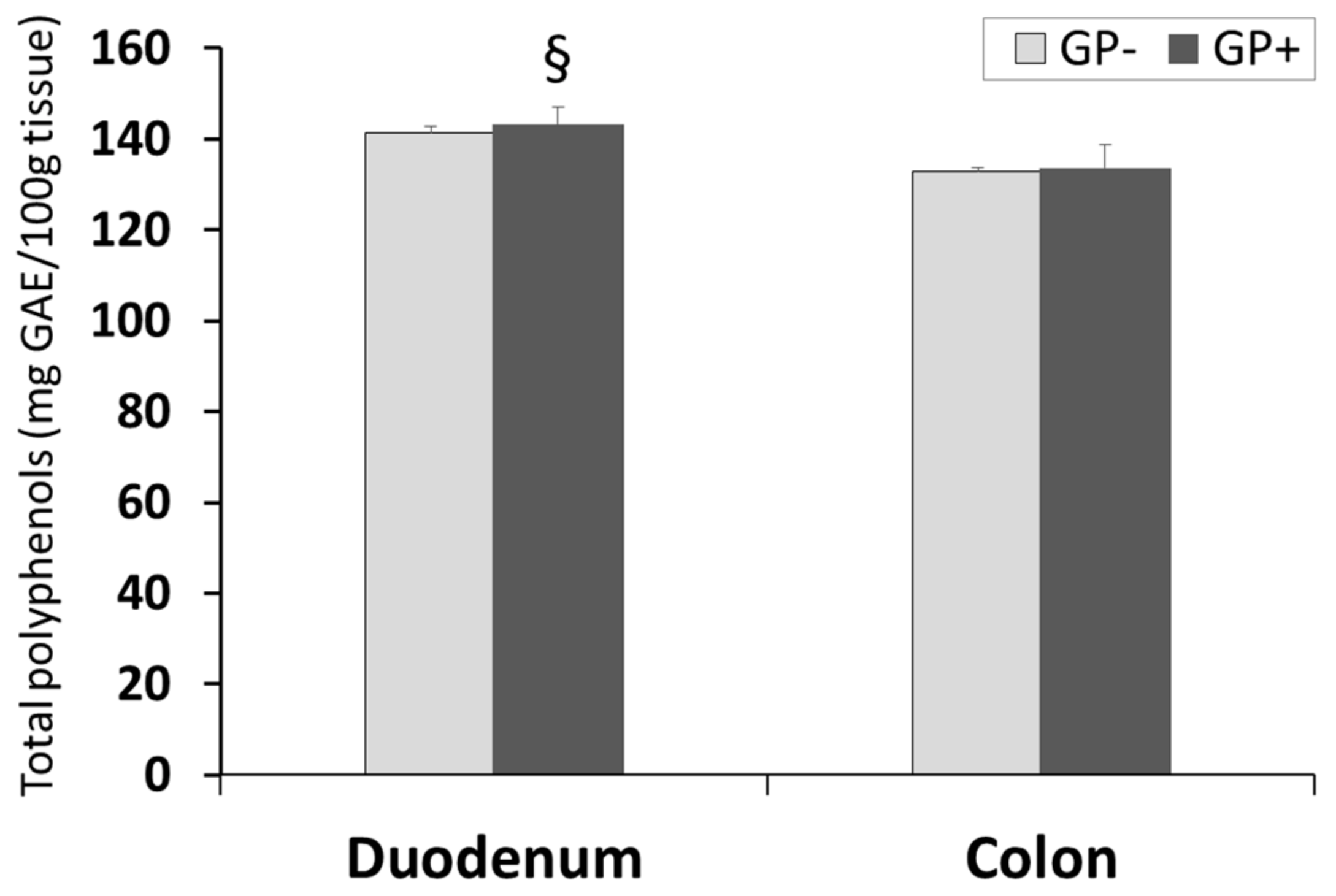
3.2.5. Total Polyphenols Content in Duodenum and Colon
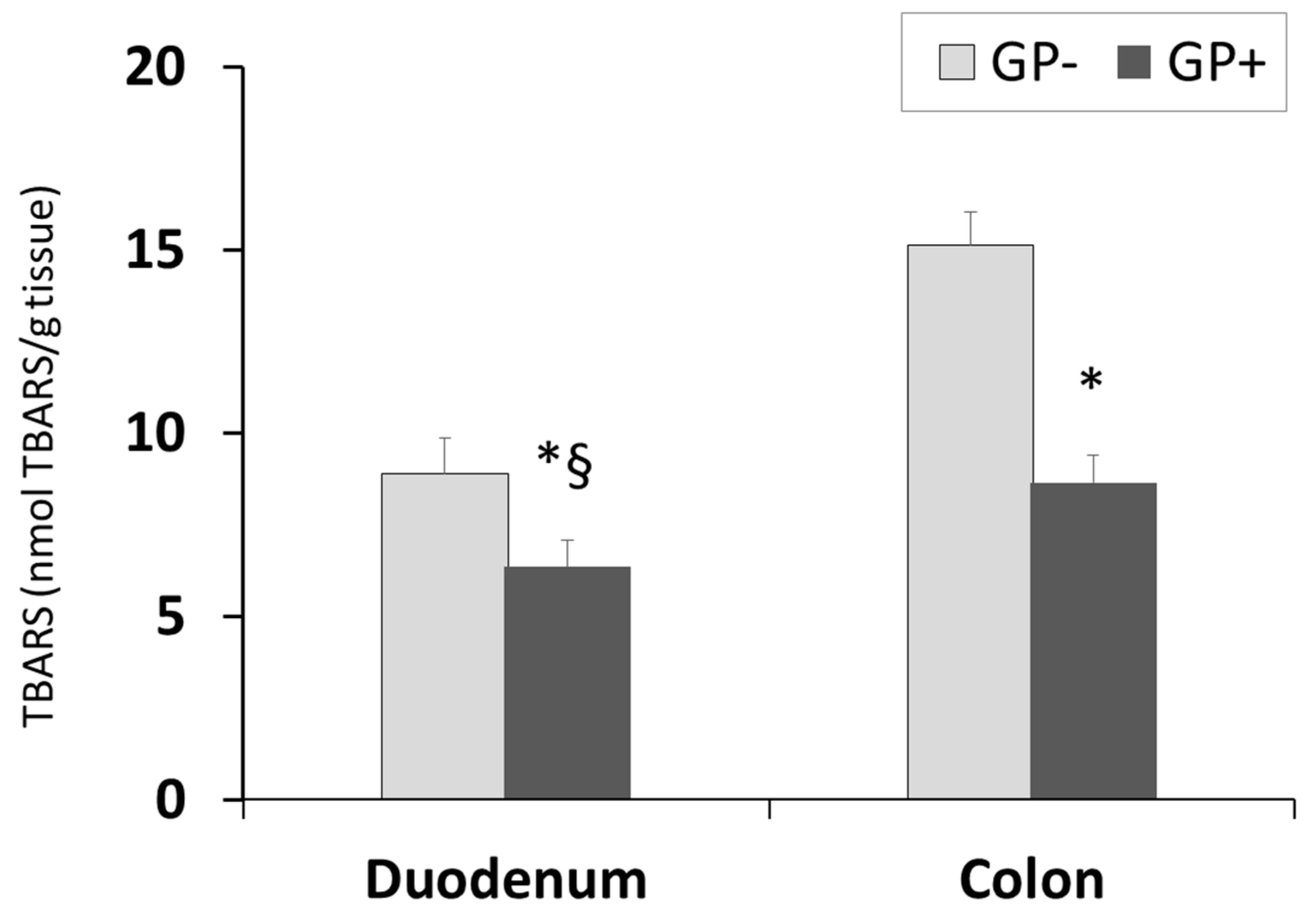
3.2.6. Effect of GP+ Diet on Lipid Peroxidation (TBARS-MDA)
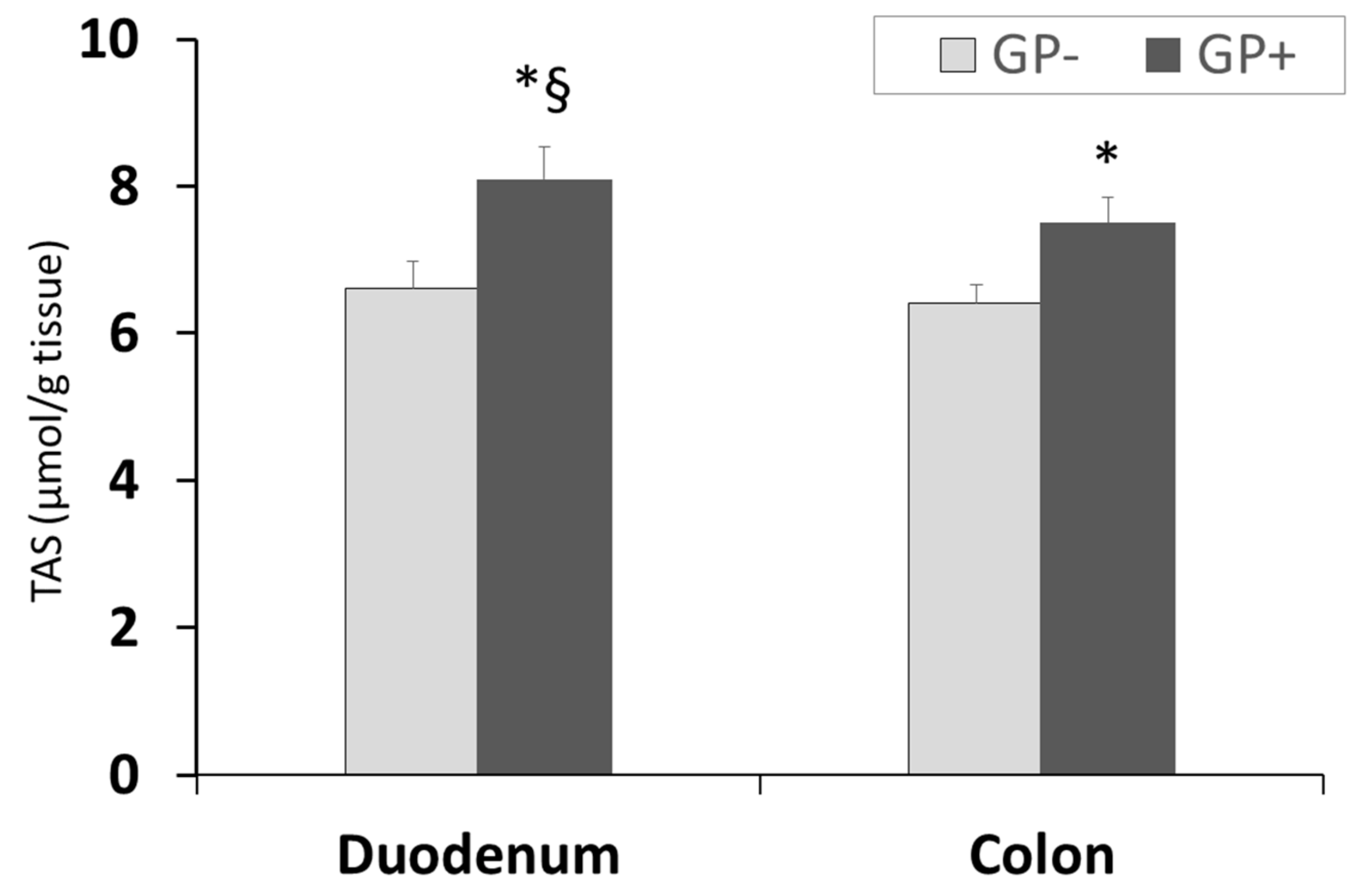
3.2.7. Effect of GP+ Diet on Total Antioxidant Status (TAS)
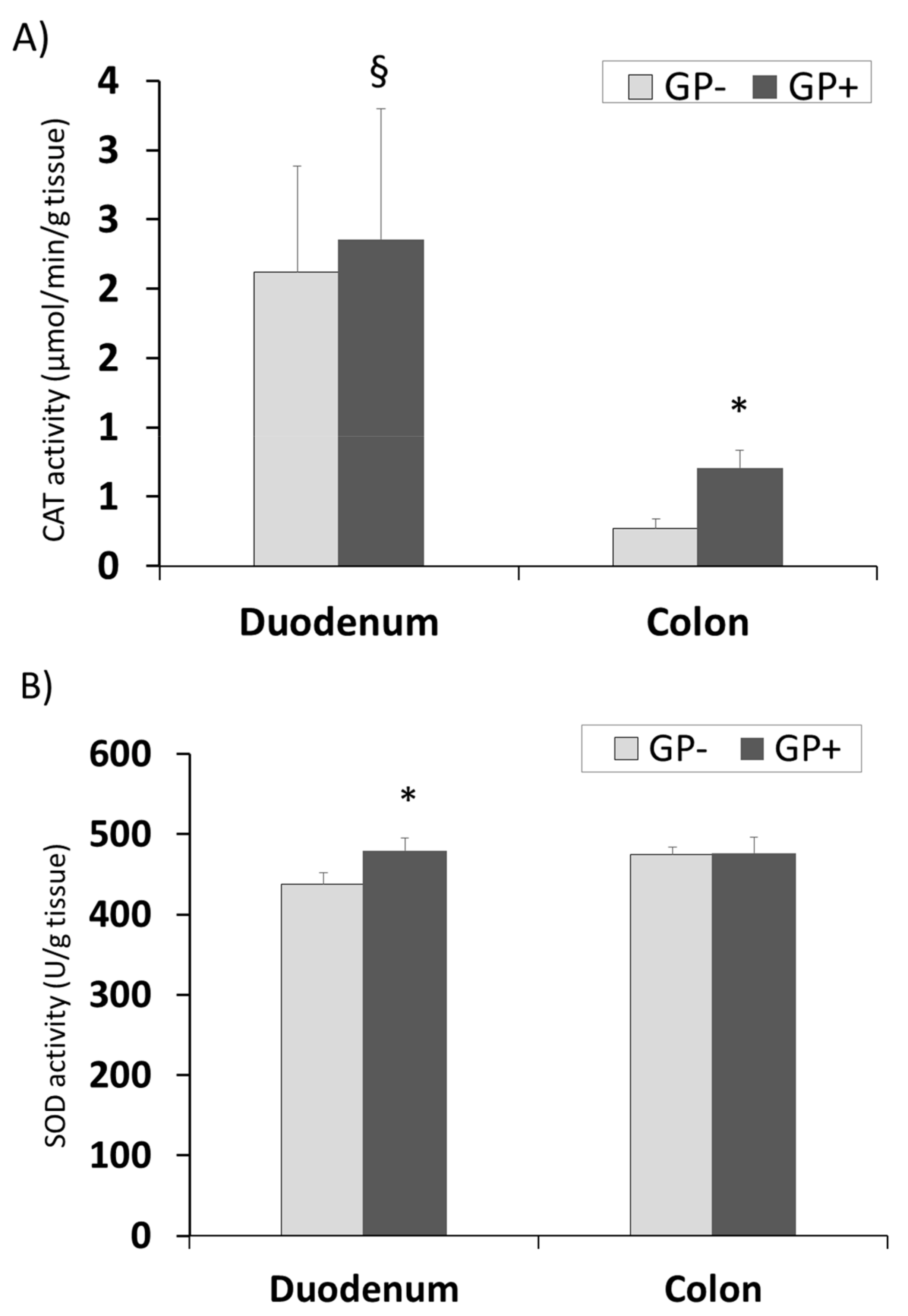
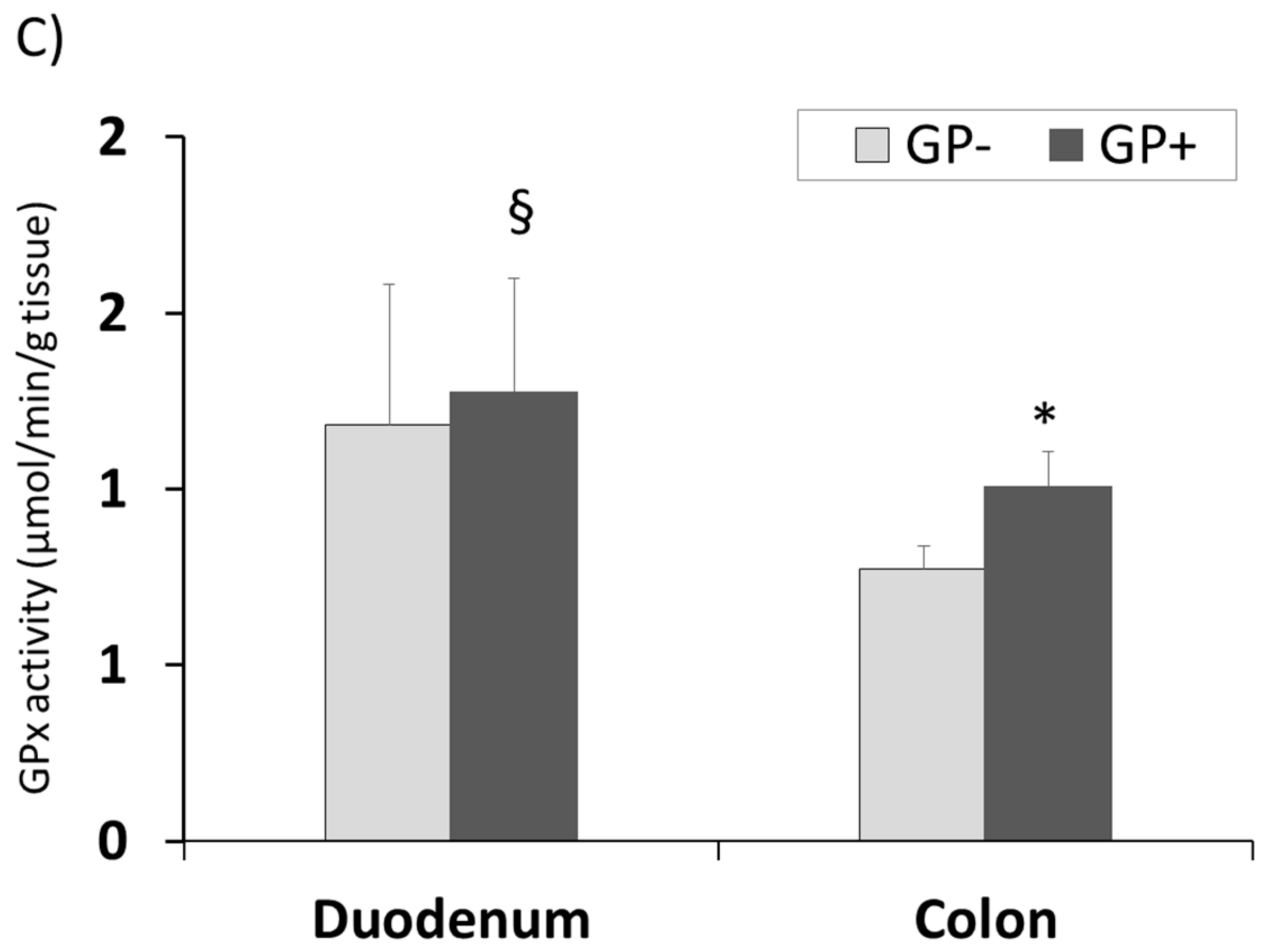
3.2.8. Effect of Antioxidant Enzymes Activity
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ruggieri, L.; Cadena, E.; Martínez-Blanco, J.; Gasol, C.M.; Rieradevall, J.; Gabarrell, X.; Gea, T.; Sort, X.; Sánchez, A. Recovery of organic wastes in the spanish wine industry. Technical, economic and environmental analyses of the composting process. J. Clean. Prod. 2009, 17, 830–838. [Google Scholar] [CrossRef]
- Cotoras, M.; Vivanco, H.; Melo, R.; Aguirre, M.; Silva, E.; Mendoza, L. In vitro and in vivo evaluation of the antioxidant and prooxidant activity of phenolic compounds obtained from grape (Vitis vinifera) pomace. Molecules 2014, 19, 21154–21167. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.; Palade, L.; Rotar, M.; Călin, L.; Dragomir, C. The anthocyanin composition of a red grape pomace in relation with the wine industry by-products-valorization in animal feed. Lucrări Ştiinţifice Seria Horticultură USAMV Iaşi 2015, 58, 29–34. [Google Scholar]
- Chedea, V.S.; Pelmus, R.S.; Lazar, C.; Pistol, G.C.; Calin, L.G.; Toma, S.M.; Dragomir, C.; Taranu, I. Effects of a diet containing dried grape pomace on blood metabolites and milk composition of dairy cows. J. Sci. Food Agric. 2017, 97, 2516–2523. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Gessner, D.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Fraga, C.G.; Oteiza, P.I. Dietary flavonoids: Role of (−)-epicatechin and related procyanidins in cell signaling. Free Radic. Biol. Med. 2011, 51, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.; Pop, R. Procyanidins-Bioavailability and Metabolization; ResearchGate GmbH: Berlin, Germnay, 2016; pp. 91–128. [Google Scholar]
- Spencer, J.P.; Chaudry, F.; Pannala, A.S.; Srai, S.K.; Debnam, E.; Rice-Evans, C. Decomposition of cocoa procyanidins in the gastric milieu. Biochem. Biophys. Res. Commun. 2000, 272, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Langerholc, T.; Maragkoudakis, P.A.; Wollgast, J.; Gradisnik, L.; Cencic, A. Novel and established intestinal cell line models—An indispensable tool in food science and nutrition. Trends Food Sci. Technol. 2011, 22, S11–S20. [Google Scholar] [CrossRef]
- Davis, S.; Illum, L.; Hinchcliffe, M. Gastrointestinal transit of dosage forms in the pig. J. Pharm. Pharmacol. 2001, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Rauw, W.; Portolés, O.; Corella, D.; Soler, J.; Reixach, J.; Tibau, J.; Prat, J.; Diaz, I.; Gómez-Raya, L. Behaviour influences cholesterol plasma levels in a pig model. Animal 2007, 1, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Rzeppa, S.; Bittner, K.; Döll, S.; Dänicke, S.; Humpf, H.-U. Urinary excretion and metabolism of procyanidins in pigs. Mol. Nutr. Food Res. 2012, 56, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K. Advances in swine biomedical model genomics. Int. J. Biol. Sci. 2007, 3, 179. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.S.; Braicu, C.; Socaciu, C. Antioxidant/prooxidant activity of a polyphenolic grape seed extract. Food Chem. 2010, 121, 132–139. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. In Vitro 2011, 25, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Sehm, J.; Treutter, D.; Lindermayer, H.; Meyer, H.H.; Pfaffl, M.W. The influence of apple-or red-grape pomace enriched piglet diet on blood parameters, bacterial colonisation, and marker gene expression in piglet white blood cells. Food Nutr. Sci. 2011, 2, 366. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.-H.; Toşa, M.I. Total phenolic contents, antioxidant activities, and lipid fractions from berry pomaces obtained by solid-state fermentation of two sambucus species with Aspergillus niger. J. Agric. Food Chem. 2015, 63, 3489–3500. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Arnous, A.; Makris, D.P.; Kefalas, P. Effect of principal polyphenolic components in relation to antioxidant characteristics of aged red wines. J. Agric. Food Chem. 2001, 49, 5736–5742. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [PubMed]
- McCracken, B.A.; Gaskins, H.R.; Ruwe-Kaiser, P.J.; Klasing, K.C.; Jewell, D.E. Diet-dependent and diet-independent metabolic responses underlie growth stasis of pigs at weaning. J. Nutr. 1995, 125, 2838–2845. [Google Scholar] [PubMed]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning anorexia may contribute to local inflammation in the piglet small intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Pié, S.; Lallès, J.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Li, Q.; Zhao, J.; Li, H.; Wang, W.; Gao, J. Effects of grape seed procyanidins on growth performance, immune function and antioxidant capacity in weaned piglets. Livest. Sci. 2015, 178, 237–242. [Google Scholar] [CrossRef]
- Zhang, H.J.; Jiang, X.R.; Mantovani, G.; Lumbreras, A.E.V.; Comi, M.; Alborali, G.; Savoini, G.; Dell’Orto, V.; Bontempo, V. Modulation of plasma antioxidant activity in weaned piglets by plant polyphenols. Italian J. Anim. Sci. 2014, 13, 3242. [Google Scholar] [CrossRef]
- García-Lomillo, J.; González-SanJosé, M.L. Applications of wine pomace in the food industry: Approaches and functions. Compr. Rev. Food Sci. Food Saf. 2017, 16, 3–22. [Google Scholar] [CrossRef]
- Hong, J.; Lu, H.; Meng, X.; Ryu, J.-H.; Hara, Y.; Yang, C.S. Stability, cellular uptake, biotransformation, and efflux of tea polyphenol (−)-epigallocatechin-3-gallate in HT-29 human colon adenocarcinoma cells. Cancer Res. 2002, 62, 7241–7246. [Google Scholar] [PubMed]
- Dangles, O.; Fargeix, G.; Dufour, C. Antioxidant properties of anthocyanins and tannins: A mechanistic investigation with catechin and the 3′, 4′, 7-trihydroxyflavylium ion. J. Chem. Soc. Perkin Trans. 2 2000, 8, 1653–1663. [Google Scholar] [CrossRef]
- Chedea, V.S.; Choueiri, L.; Jisaka, M.; Kefalas, P. O-quinone involvement in the prooxidant tendency of a mixture of quercetin and caffeic acid. Food Chem. 2012, 135, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; O'brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Van der Woude, H.; Boersma, M.G.; Alink, G.M.; Vervoort, J.; Rietjens, I.M. Consequences of quercetin methylation for its covalent glutathione and DNA adduct formation. Chem.-Biol. Interact. 2006, 160, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Finkel, T. Mitohormesis. Cell Metab. 2014, 19, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Appeldoorn, M.M.; Vincken, J.-P.; Gruppen, H.; Hollman, P.C. Procyanidin dimers A1, A2, and B2 are absorbed without conjugation or methylation from the small intestine of rats. J. Nutr. 2009, 139, 1469–1473. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.; Auger, C.; Mullen, W.; Bornet, A.; Rouanet, J.-M.; Crozier, A.; Teissedre, P.-L. The absorption, metabolism and excretion of flavan-3-ols and procyanidins following the ingestion of a grape seed extract by rats. Br. J. Nutr. 2005, 94, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; Macià, A.; Romero, M.-P.; Anglès, N.; Morelló, J.R.; Motilva, M.-J. Distribution of procyanidins and their metabolites in rat plasma and tissues after an acute intake of hazelnut extract. Food Funct. 2011, 2, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.E.; Witrick, K.A.; Klotz, C.; Dorenkott, M.R.; Goodrich, K.M.; Fundaro, G.; McMillan, R.P.; Hulver, M.W.; Ponder, M.A.; Neilson, A.P. Alterations to metabolically active bacteria in the mucosa of the small intestine predict anti-obesity and anti-diabetic activities of grape seed extract in mice. Food Funct. 2017, 8, 3510–3522. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Li, F.; Xiao, H. The nutraceutical bioavailability classification scheme: Classifying nutraceuticals according to factors limiting their oral bioavailability. Annu. Rev. Food Sci. Technol. 2015, 6, 299–327. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-H.; Lee, D.-Y.; Kim, S.; Chung, J.-O.; Choi, J.-K.; Joo, K.-M.; Jeong, H.W.; Kim, J.K.; Kim, W.G.; Shim, S.-M. Influence of flavonol-rich excipient food (onion peel and Dendropanax morbifera) on the bioavailability of green tea epicatechins in vitro and in vivo. Food Funct. 2017, 8, 3664–3674. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Kosińska, A.; Xu, H.; Andlauer, W. Milk enhances intestinal absorption of green tea catechins in in vitro digestion/Caco-2 cells model. Food Res. Int. 2013, 53, 793–800. [Google Scholar] [CrossRef]
- Landis-Piwowar, K.; Chen, D.; Chan, T.H.; Dou, Q.P. Inhibition of catechol-ο-methyltransferase activity in human breast cancer cells enhances the biological effect of the green tea polyphenol (−)-EGCG. Oncol. Rep. 2010, 24, 563–569. [Google Scholar] [PubMed]
- Spencer, J.P. Metabolism of tea flavonoids in the gastrointestinal tract. J. Nutr. 2003, 133, 3255S–3261S. [Google Scholar] [CrossRef] [PubMed]
- Kulling, S.E.; Honig, D.M.; Metzler, M. Oxidative metabolism of the soy isoflavones daidzein and genistein in humans in vitro and in vivo. J. Agric. Food Chem. 2001, 49, 3024–3033. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Yousif, A.M.; Johnson, S.K.; Gamlath, S. Acute effect of sorghum flour-containing pasta on plasma total polyphenols, antioxidant capacity and oxidative stress markers in healthy subjects: A randomised controlled trial. Clin. Nutr. 2015, 34, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Arenas, E.H.; Trinidad, T.P. Fate of polyphenols in pili (Canarium ovatum Engl.) pomace after in vitro simulated digestion. Asian Pac. J. Trop. Biomed. 2017, 7, 53–58. [Google Scholar] [CrossRef]
- Torabian, S.; Haddad, E.; Rajaram, S.; Banta, J.; Sabate, J. Acute effect of nut consumption on plasma total polyphenols, antioxidant capacity and lipid peroxidation. J. Hum. Nutr. Diet. 2009, 22, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Medina-Remón, A.; Tresserra-Rimbau, A.; Arranz, S.; Estruch, R.; Lamuela-Raventos, R.M. Polyphenols excreted in urine as biomarkers of total polyphenol intake. Bioanalysis 2012, 4, 2705–2713. [Google Scholar] [CrossRef] [PubMed]
- Baroudi, O.; Younes, S.B.; Mézlini, A.; Bignon, Y.J.; Medimeg, I.; Uhrhammer, N.; Gaiied, A.B.A.E.; Ellouz, S.C. Status of antioxidant enzyme: Glutathione peroxidase and total polyphenol level in plasma of tunisian patients suffering from colorectal and gastric cancer: Interaction with clinical outcome. Med. Oncol. 2013, 30, 731. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κb and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Habeanu, M.; Gras, M.; Pistol, G.; Lefter, N.; Palade, M.; Ropota, M.; Sanda Chedea, V.; Marin, D. Assessment of the effect of grape seed cake inclusion in the diet of healthy fattening-finishing pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e30–e42. [Google Scholar] [CrossRef] [PubMed]
- Giribabu, N.; Eswar Kumar, K.; Swapna Rekha, S.; Muniandy, S.; Salleh, N. Vitis vinifera (muscat variety) seed ethanolic extract preserves activity levels of enzymes and histology of the liver in adult male rats with diabetes. Evid.-Based Complement. Altern. Med. 2015, 2015, 542026. [Google Scholar] [CrossRef] [PubMed]
- Goutzourelas, N.; Stagos, D.; Demertzis, N.; Mavridou, P.; Karterolioti, H.; Georgadakis, S.; Kerasioti, E.; Aligiannis, N.; Skaltsounis, L.; Statiri, A. Effects of polyphenolic grape extract on the oxidative status of muscle and endothelial cells. Hum. Exp. Toxicol. 2014, 33, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Resveratrol inhibits tnf-α-induced changes of adipokines in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2007, 364, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Blau, S.; Rubinstein, A.; Bass, P.; Singaram, C.; Kohen, R. Differences in the reducing power along the rat gi tract: Lower antioxidant capacity of the colon. Mol. Cell. Biochem. 1999, 194, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Bobek, P.; Ozdín, L.; Hromadova, M. The effect of dried tomato, grape and apple pomace on the cholesterol metabolism and antioxidative enzymatic system in rats with hypercholesterolemia. Mol. Nutr. Food Res. 1998, 42, 317–320. [Google Scholar] [CrossRef]
- Moghadasian, M.H.; Godin, D.V. Gender-related regional antioxidant profiles in the gastrointestinal tract of the rat. Mol. Cell. Biochem. 1996, 155, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Siegers, C.-P.; Bartels, L.; Riemann, D. Effects of fasting and glutathione depletors on the GSH-dependent enzyme system in the gastrointestinal mucosa of the rat. Pharmacology 1989, 38, 121–128. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Weaning Phase 1 | |
|---|---|---|
| Control Diet (Days 1–36) | GP+ Diet (Days 1–36) | |
| Corn | 57.32 | 53.90 |
| Rice meal | - | - |
| Wheat | 10.00 | 8.00 |
| Sunflower meal (31.94% CP) | 5.00 | 5.00 |
| Soybean meal (44% CP) | 16.00 | 16.00 |
| Sunflower oil | 0.20 | 0.70 |
| Milk powder | 3.00 | 3.00 |
| Gluten | 4.00 | 4.00 |
| Grape pomace | - | 5.00 |
| Monocalcium phosphate | 1.25 | 1.25 |
| Limestone | 1.56 | 1.48 |
| NaCl | 0.20 | 0.20 |
| Metionine | 0.03 | 0.05 |
| Lisine | 0.34 | 0.32 |
| Choline | 0.10 | 0.10 |
| Mineral vitamin-premix 2 | 1.00 | 1.00 |
| Calculated Nutrient content | ||
| Dry matter (%) | 88.31 | 88.01 |
| Crude Protein (%) | 18.29 | 18.36 |
| Digestible crude protein (%) | 14.98 | 15.06 |
| Fat (%) | 2.85 | 2.96 |
| Crude fiber (%) | 4.07 | 5.30 |
| Metabolisable energy (kcal/kg) | 3169 | 3147 |
| Lysine (%) | 1.08 | 1.08 |
| Digestible Lysine (%) | 0.92 | 0.92 |
| Met + Cys (%) | 0.65 | 0.65 |
| Calcium (%) | 0.90 | 0.90 |
| Phosphorus (%) | 0.65 | 0.65 |
| TOTAL | 100.00 | 100.00 |
| In Vitro | 3 h | 24 h | ||||
|---|---|---|---|---|---|---|
| 250 ng GAE/mL | 500 ng GAE/mL | 1000 ng GAE/mL | 250 ng GAE/mL | 500 ng GAE/mL | 1000 ng GAE/mL | |
| Extracellularλmax (nm) | 276.0 | 327.9 428.7 399.5 | 325.3 393.3 427.6 | 295.6 | 298.3 487.5 627.0 | 303.6557.5 |
| Cellularλmax (nm) | 313.1 554.9 | 451.5 515.1 539.0 581.9 | 434.0 559.1 | 438.2 561.2 | 579.3 | 287.1 |
| Compound | Rt (min) | UV λmax (nm) | [M − H]+ | GP mg CE */100 g | GP+ mg CE/100 g | GP− mg CE/100 g |
|---|---|---|---|---|---|---|
| Gallic acid-glucoside | 4.29 | 258 | 333, 171 | 2.22 | - | - |
| Gallic acid | 5.06 | 272 | 171 | 2.71 | 4.61 | 4.60 |
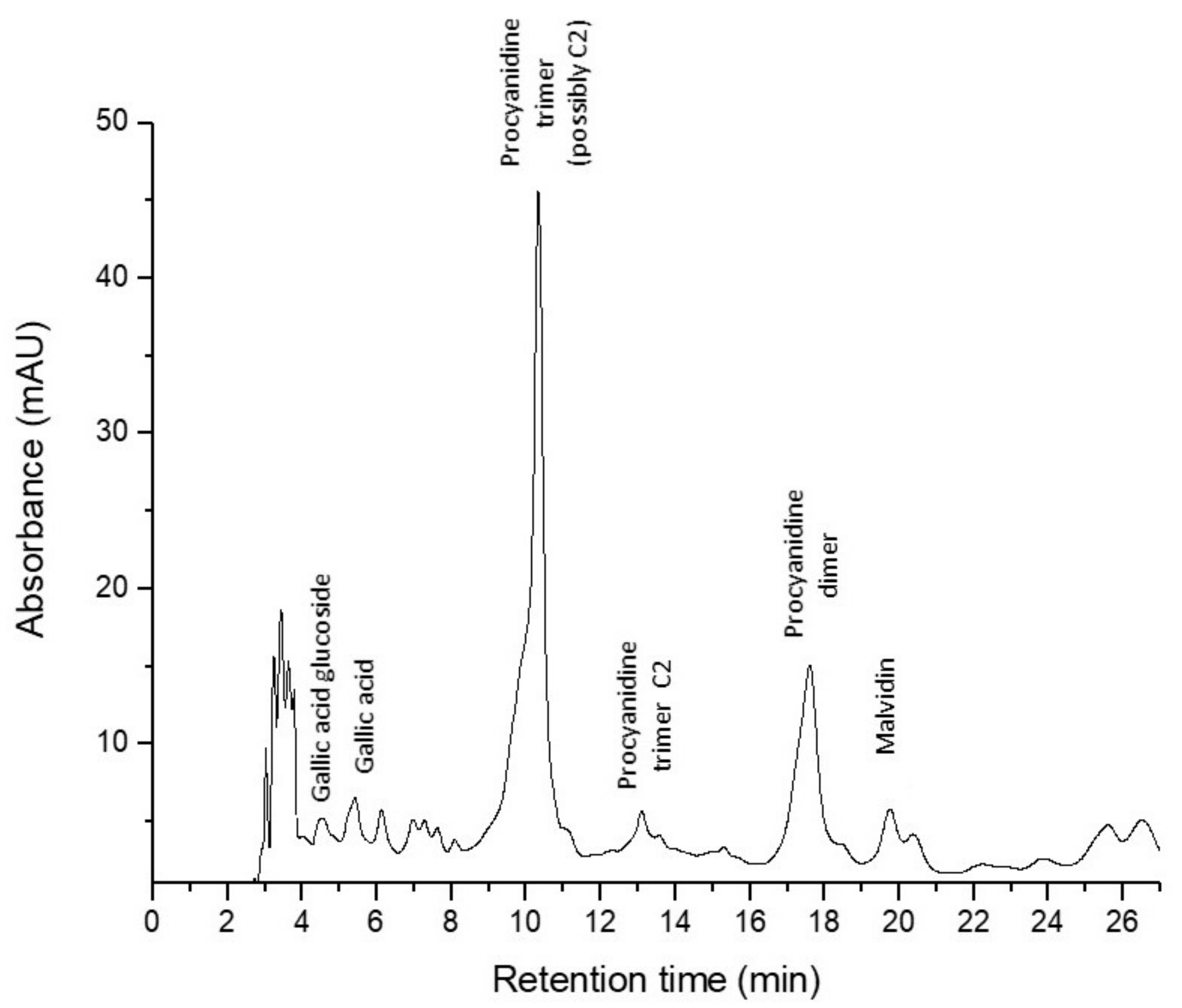
| Procyanidin trimer (possibly C2) | 11.67 | 280 | 867, 290 | 16.54 | 2.12 | - |
| Procyanidin trimer C1 | 13.01 | 280 | 867, 290 | 2.24 | - | - |
| Procyanidin dimer | 17.91 | 280 | 579, 290 | 7.79 | 1.80 | - |
| Malvidin (3-O-6″-coumaroyl-glucoside) | 20.07 | 275, 532 | 639, 331 | 0.76 | - | - |
| Ferulic acid derivate (Ferulic acid dehydrotrimer) | 10.75 | 324 | 565, 195 | - | 12.92 | 12.92 |
| Caffeoylquinic acid | 12.21 | 326 | 355, 181 | - | 43.98 | 43.97 |
| Daidzin (Daidzein-7-O-glucoside) | 14.87 | 262 | 417 | - | 14.24 | 14.23 |
| p-Coumaroylquinic acid | 15.23 | 316 | 339, 193, 165 | - | 24.57 | 24.56 |
| 6″-O-Malonyldaidzin | 16.81 | 255 | 503 | - | 11.88 | 11.88 |
| Genistin (Genistein-7-O-glucoside) | 17.11 | 265 | 433 | - | 16.22 | 16.21 |
| Ferulic acid | 17.82 | 312 | 195 | - | 15.33 | 15.32 |
| 6″-O-Malonylgenistin | 18.81 | 260 | 519 | - | 17.27 | 17.25 |
| Dicaffeoylquinic acid | 20.81 | 326 | 517, 355, 181 | - | 18.96 | 18.96 |
| Growth Performance | GP− | GP+ | p-Value | ||
|---|---|---|---|---|---|
| AVRG | SEM | AVRG | SEM | ||
| Body weight (kg) at day 1 | 10.67 | 0.300 | 10.90 | 0.256 | 0.600 |
| Body weight (kg) at day 15 | 17.89 | 0.655 | 18.15 | 0.742 | 0.797 |
| Body weight (kg) at day 36 | 31.22 | 1.188 | 33.00 | 1.370 | 0.346 |
| days 1 to 15 | |||||
| ADG (g) | 481.48 | 41.61 | 483.33 | 40.14 | 0.975 |
| ADFI (g) | 862.80 | 21.24 | 898.00 | 22.02 | 0.260 |
| F/G | 1.92 | 0.19 | 1.99 | 0.19 | 0.780 |
| days 16 to 36 | |||||
| ADG (g) | 634.92 | 31.99 | 707.14 | 39.53 | 0.179 |
| ADFI (g) | 1264.85 a | 51.74 | 1510.24 b | 38.83 | 0.0005 |
| F/G | 2.03 | 0.10 | 2.22 | 0.18 | 0.371 |
| days 1 to 36 | |||||
| ADG (g) | 570.99 | 34.29 | 613.89 | 35.00 | 0.395 |
| ADFI (g) | 1097.33 a | 45.72 | 1255.14 b | 56.45 | 0.033 |
| F/G | 1.98 | 0.11 | 2.10 | 0.16 | 0.627 |
| Samples | λmax (nm) | |||
|---|---|---|---|---|
| Duodenum | 287.5 | - | 430 | - |
| Colon | 279 | 292 | 444 | 587 |
| Compound | Rt (min) | UV λmax (nm) | [M − H]+ | Duodenum mg CE */100 g | Colon mg CE */100 g | ||
|---|---|---|---|---|---|---|---|
| D− | D+ | C− | C+ | ||||
| Procyanidin trimer (possibly C2) | 10.16 | 280 | 867, 291 | 13.69 | 16.00 | 10.10 | 10.00 |
| Catechin | 12.67 | 280 | 291 | - | - | 9.45 | 8.73 |
| Procyanidin trimer C1 | 14.33 | 280 | 867, 291 | - | - | 6.85 | 5.38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chedea, V.S.; Palade, L.M.; Marin, D.E.; Pelmus, R.S.; Habeanu, M.; Rotar, M.C.; Gras, M.A.; Pistol, G.C.; Taranu, I. Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients 2018, 10, 588. https://doi.org/10.3390/nu10050588
Chedea VS, Palade LM, Marin DE, Pelmus RS, Habeanu M, Rotar MC, Gras MA, Pistol GC, Taranu I. Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients. 2018; 10(5):588. https://doi.org/10.3390/nu10050588
Chicago/Turabian StyleChedea, Veronica Sanda, Laurentiu Mihai Palade, Daniela Eliza Marin, Rodica Stefania Pelmus, Mihaela Habeanu, Mircea Catalin Rotar, Mihail Alexandru Gras, Gina Cecilia Pistol, and Ionelia Taranu. 2018. "Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols" Nutrients 10, no. 5: 588. https://doi.org/10.3390/nu10050588
APA StyleChedea, V. S., Palade, L. M., Marin, D. E., Pelmus, R. S., Habeanu, M., Rotar, M. C., Gras, M. A., Pistol, G. C., & Taranu, I. (2018). Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients, 10(5), 588. https://doi.org/10.3390/nu10050588