Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury

 , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
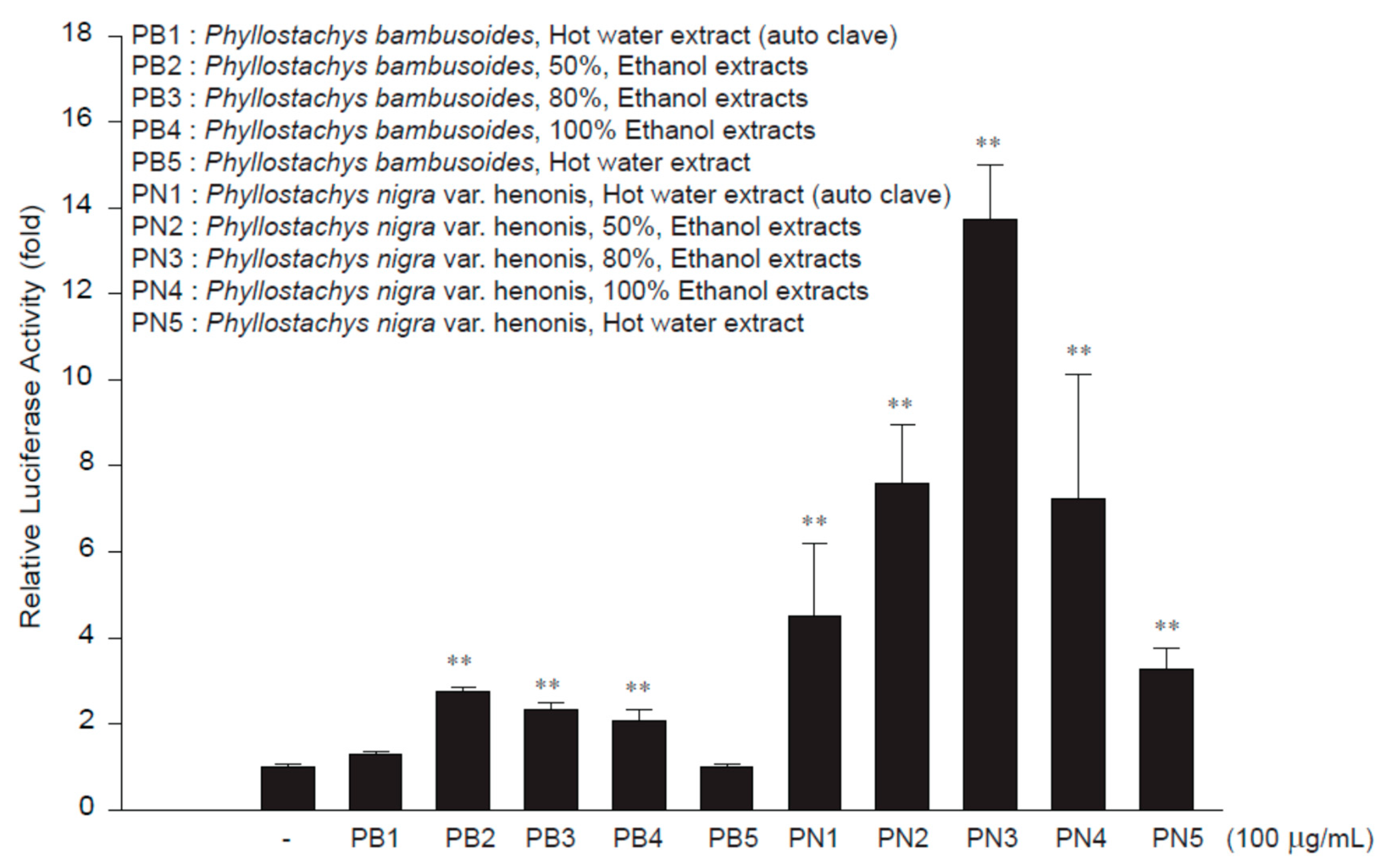
2.2. Preparation of Bamboo Stem Extracts
2.3. HPLC Analysis of Bamboo Stem Extracts
2.4. Animals
2.5. Blood Chemistry
2.6. Histopathology
2.7. Assay of ROS Generation
2.8. Determination of GSH Content
2.9. Primary Hepatocyte Isolation
2.10. Cell Culture
2.11. Immunoblot Analysis
2.12. Cytotoxicity Assay
2.13. Luciferase Assay
2.14. Statistical Analysis
3. Results
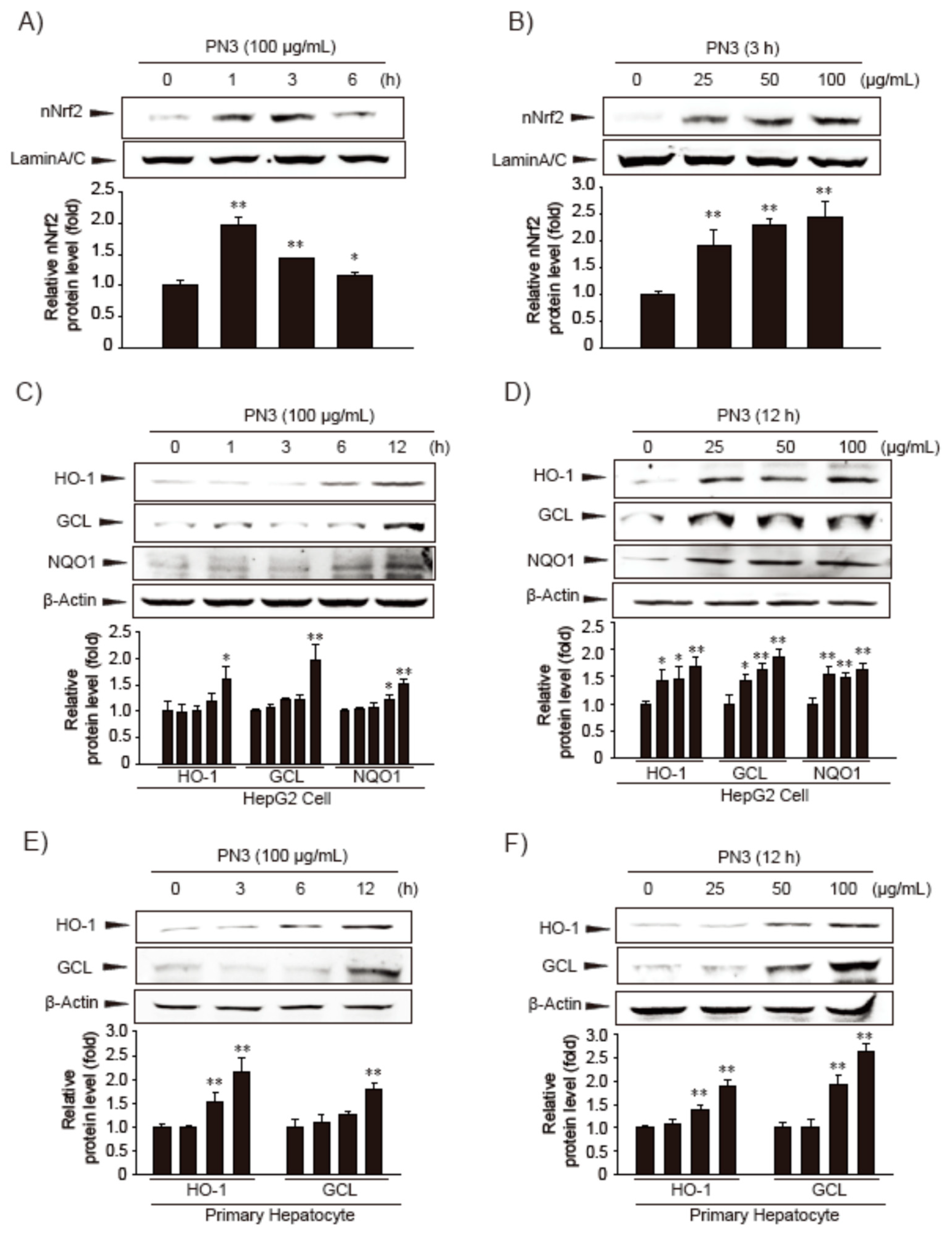
3.1. Nrf2 Activation by PN3
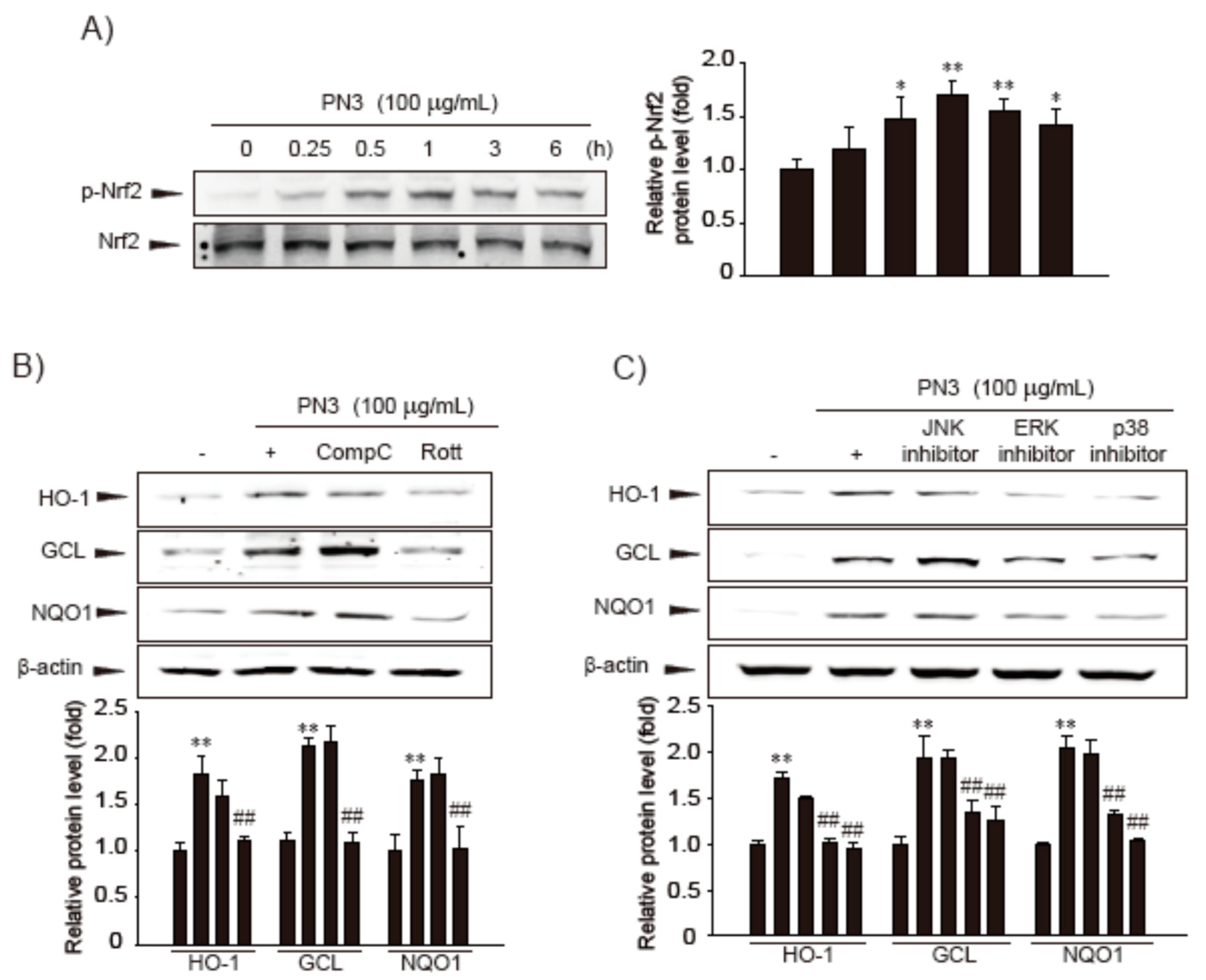
3.2. Nrf2 Phosphorylation by ERK, P38 MAPKs, and PKCδ in PN3
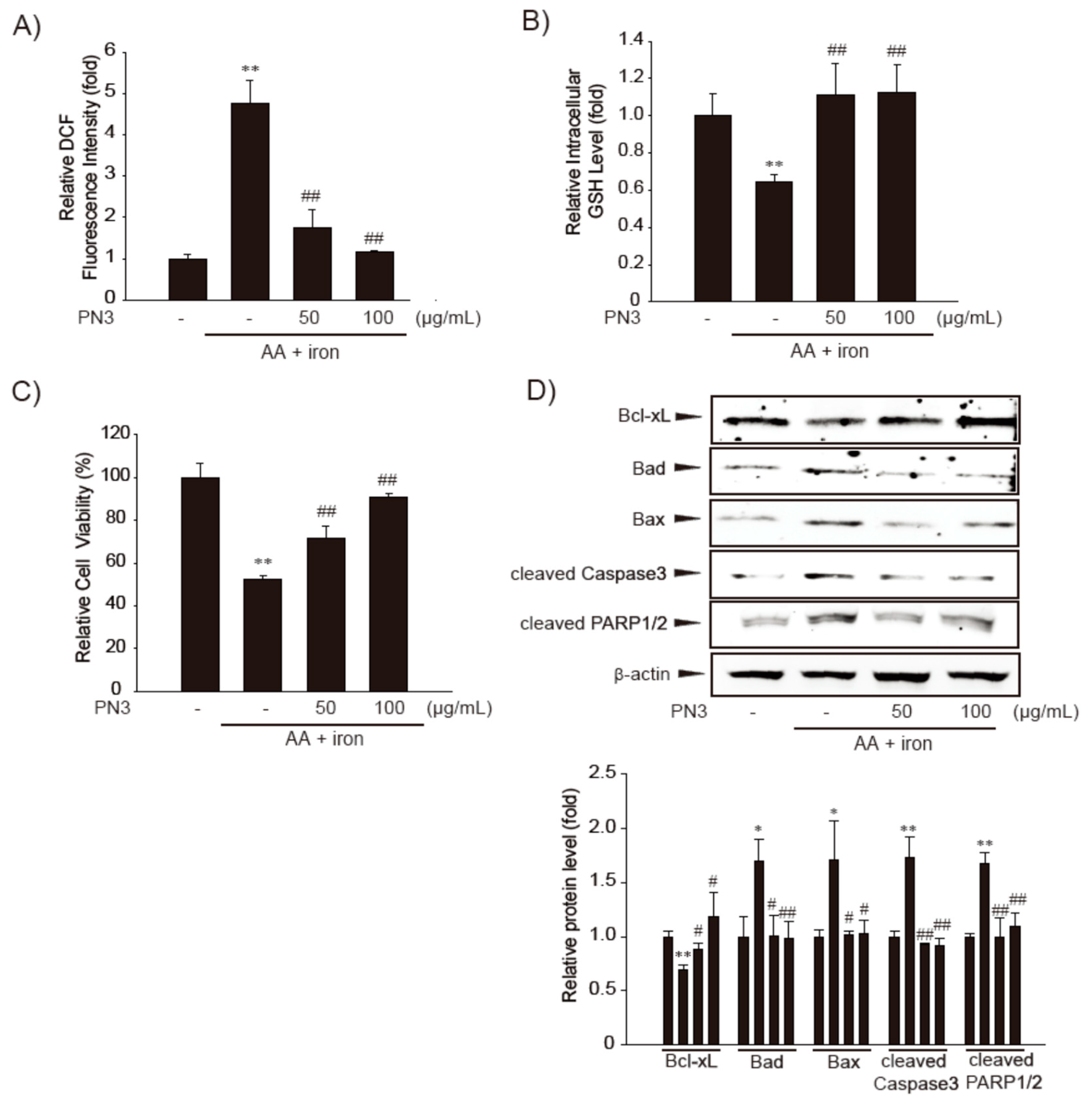
3.3. Cytoprotective Effect of PN3
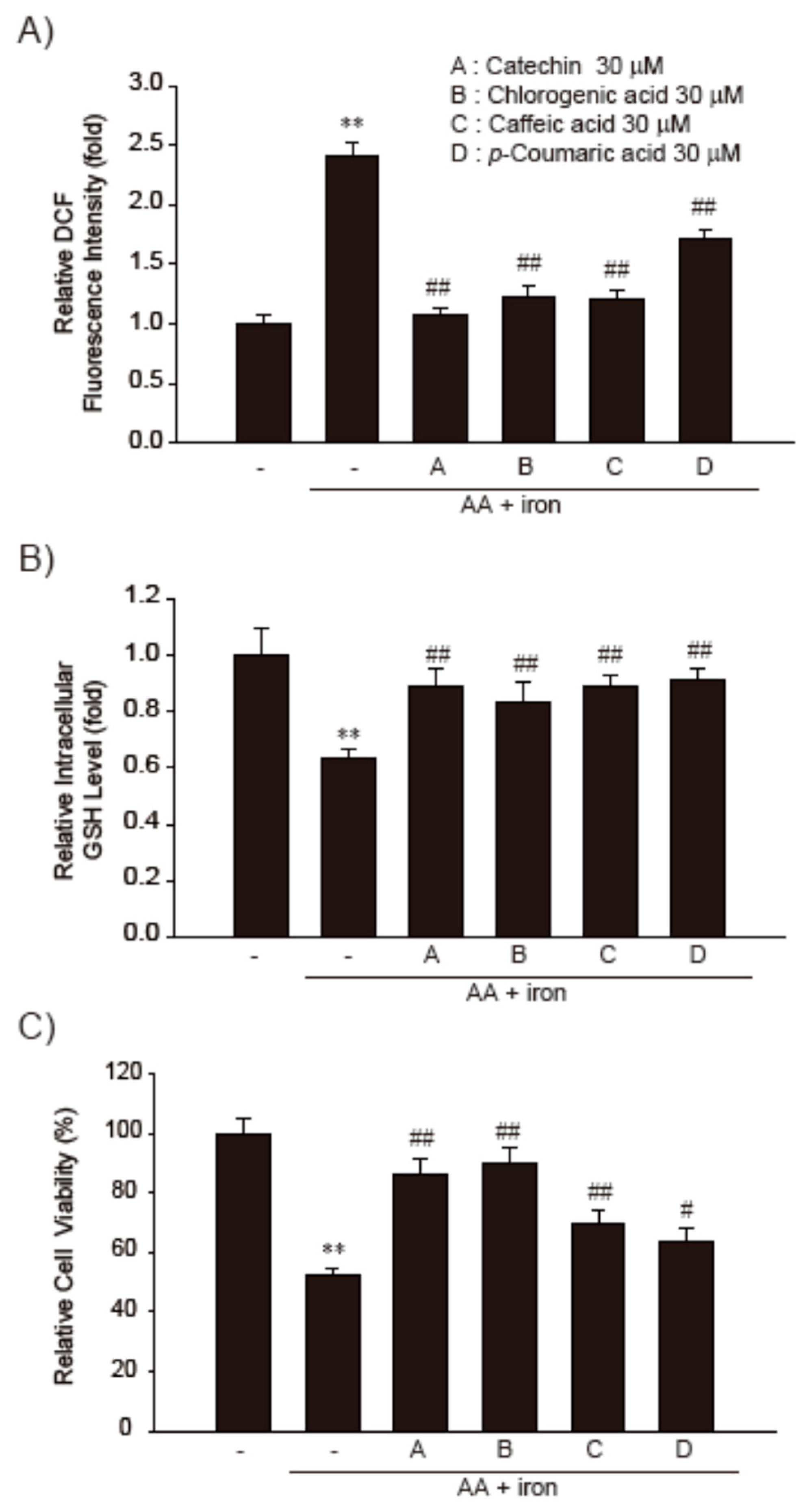
3.4. Effect of the Major Functional Constituents of PN3 in AA + Fe-Treated HepG2 Cells
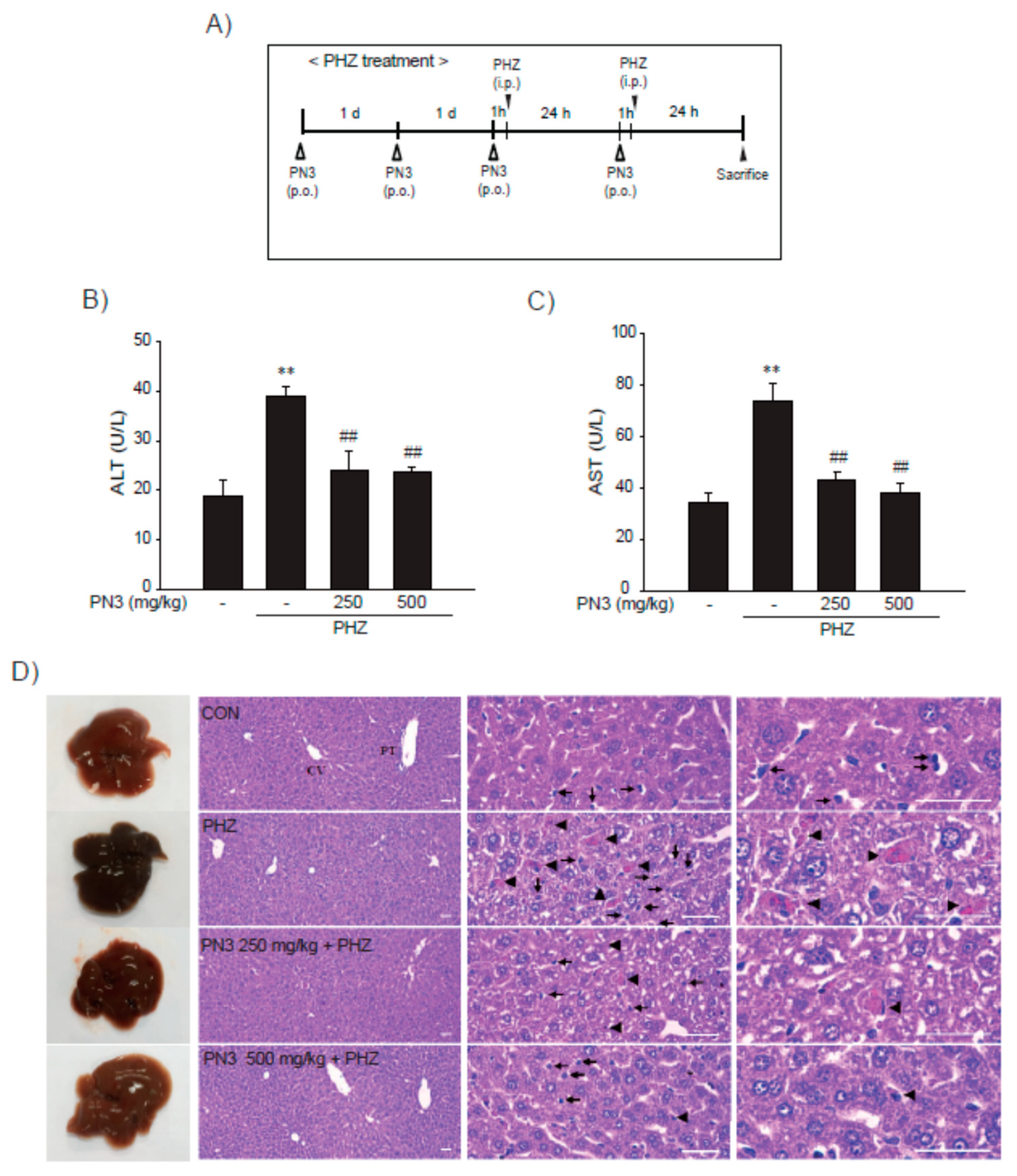
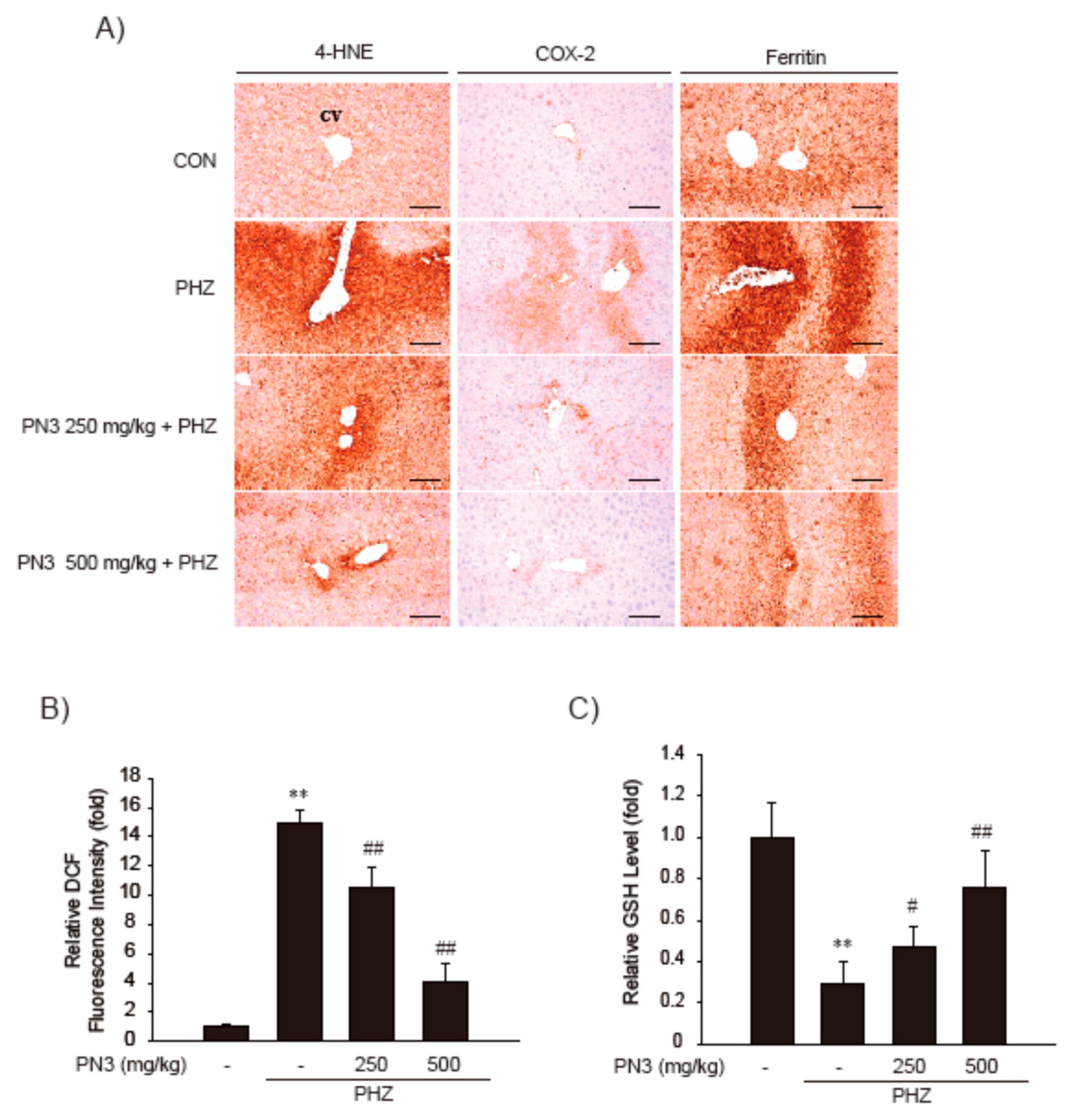
3.5. Effect of PN3 on PHZ-Induced Acute Liver Injury
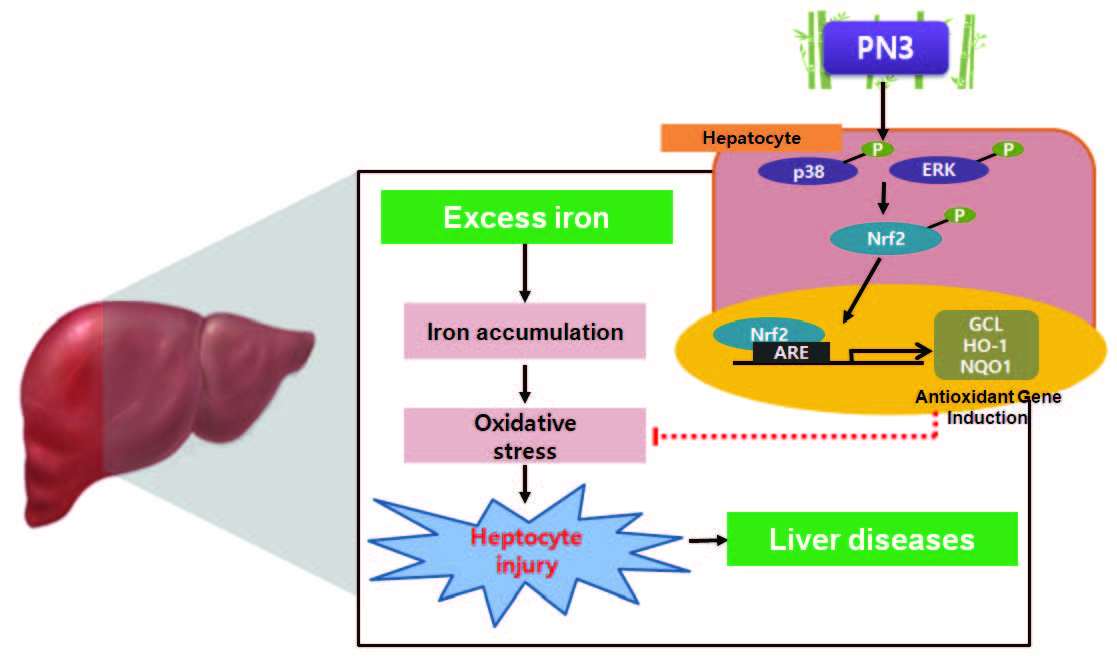
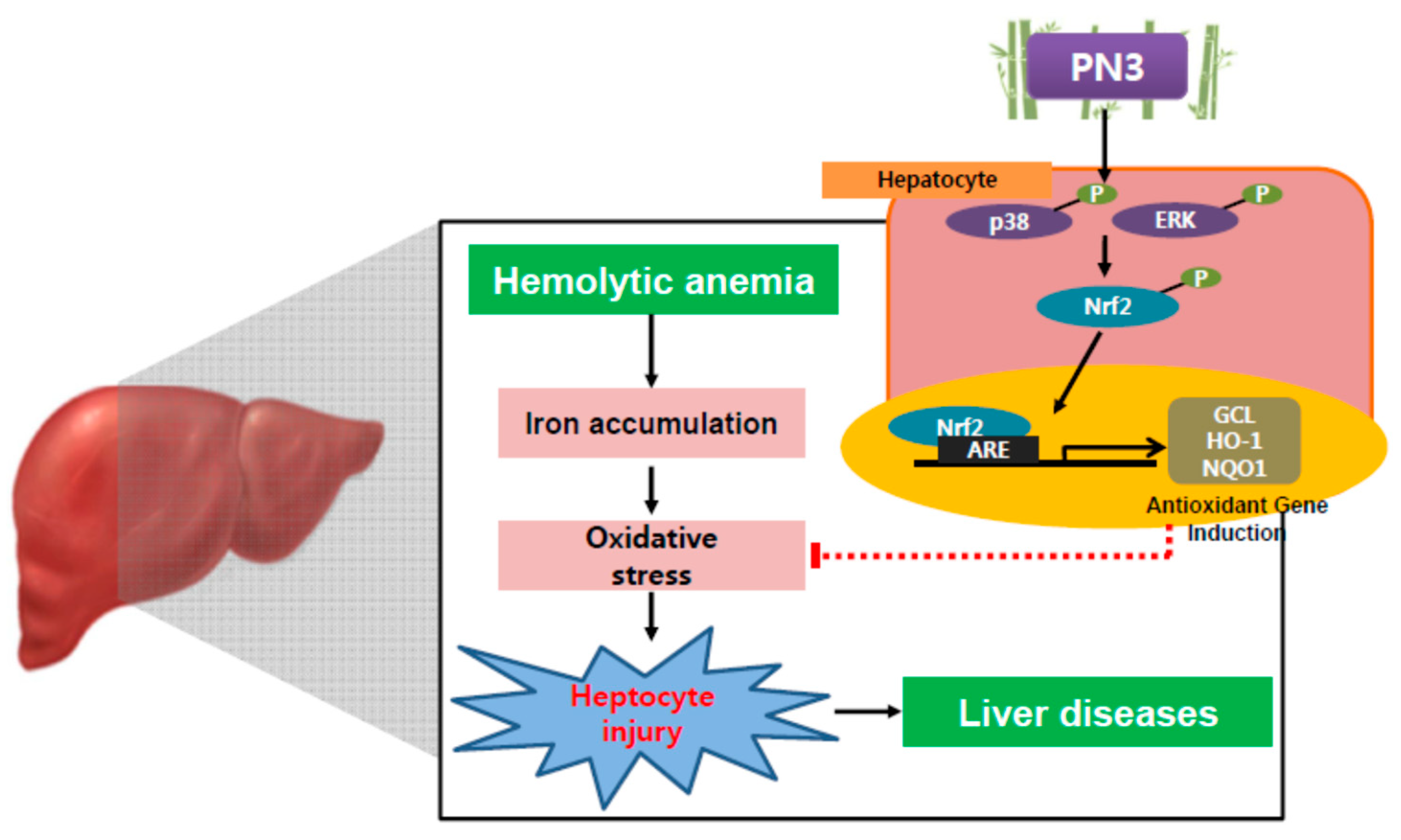
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Idilman, I.S.; Akata, D.; Ozmen, M.N.; Karcaaltincaba, M. Different forms of iron accumulation in the liver on MRI. Diagn. Interv. Radiol. 2016, 22, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.R.; Shah, Y.M. Iron homeostasis in the liver. Compr. Physiol. 2013, 3, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Taddei, T.; Mistry, P.; Schilsky, M.L. Inherited metabolic disease of the liver. Curr. Opin. Gastroenterol. 2008, 24, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Costa-Matos, L.; Batista, P.; Monteiro, N.; Henriques, P.; Girao, F.; Carvalho, A. Hfe mutations and iron overload in patients with alcoholic liver disease. Arq. Gastroenterol. 2013, 50, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Aigner, E.; Weiss, G.; Datz, C. Dysregulation of iron and copper homeostasis in nonalcoholic fatty liver. World J. Hepatol. 2015, 7, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Bonkovsky, H.L.; Banner, B.F.; Rothman, A.L. Iron and chronic viral hepatitis. Hepatology 1997, 25, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Pietrangelo, A. Iron, oxidative stress and liver fibrogenesis. J. Hepatol. 1998, 28 (Suppl. 1), 8–13. [Google Scholar] [CrossRef]
- Hultcrantz, R.; Ahlberg, J.; Glaumann, H. Isolation of two lysosomal populations from iron-overloaded rat liver with different iron concentration and proteolytic activity. Virchows Archiv. B Cell Pathol. Incl. Mol. Pathol. 1984, 47, 55–65. [Google Scholar] [CrossRef]
- Bacon, B.R.; O’Neill, R.; Britton, R.S. Hepatic mitochondrial energy production in rats with chronic iron overload. Gastroenterology 1993, 105, 1134–1140. [Google Scholar] [CrossRef]
- Pietrangelo, A.; Grandi, R.; Tripodi, A.; Tomasi, A.; Ceccarelli, D.; Ventura, E.; Masini, A. Lipid composition and fluidity of liver mitochondria, microsomes and plasma membrane of rats with chronic dietary iron overload. Biochem. Pharmacol. 1990, 39, 123–128. [Google Scholar] [CrossRef]
- Caro, A.A.; Cederbaum, A.I. Synergistic toxicity of iron and arachidonic acid in HepG2 cells overexpressing CYP2E1. Mol. Pharmacol. 2001, 60, 742–752. [Google Scholar]
- Shin, S.M.; Kim, S.G. Inhibition of arachidonic acid and iron-induced mitochondrial dysfunction and apoptosis by oltipraz and novel 1,2-dithiole-3-thione congeners. Mol. Pharmacol. 2009, 75, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.Z.; Lee, J.H.; Ki, S.H.; Yang, J.H.; Cho, I.J.; Kang, S.H.; Zhao, R.J.; Kim, S.C.; Kim, Y.W. AMPK activation by isorhamnetin protects hepatocytes against oxidative stress and mitochondrial dysfunction. Eur. J. Pharmacol. 2014, 740, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J. Biol. Chem. 2017, 292, 16817–16824. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Yang, J.H.; Ki, S.H. Role of the Nrf2-ARE pathway in liver diseases. Oxidative Med. Cell. Longev. 2013, 763257. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Crozier, A. Dietary flavonoids and phenolic compounds. In Plant Phenolics and Human Health: Biochemistry, Nutrition, and Pharmacology Plant; John Wiley & Sons, Inc.: Hoboken, NY, USA, 2010; Volume 1, pp. 1–50. [Google Scholar]
- Zhao, L.; Wang, Y.; Liu, J.; Wang, K.; Guo, X.; Ji, B.; Wu, W.; Zhou, F. Protective effects of genistein and puerarin against chronic alcohol-induced liver injury in mice via antioxidant, anti-inflammatory, and anti-apoptotic mechanisms. J. Agric. Food Chem. 2016, 64, 7291–7297. [Google Scholar] [CrossRef]
- Van De Wier, B.; Koek, G.H.; Bast, A.; Haenen, G.R. The potential of flavonoids in the treatment of non-alcoholic fatty liver disease. Crit. Rev. Food Sci. Nutr. 2017, 57, 834–855. [Google Scholar] [CrossRef]
- Panee, J. Potential medicinal application and toxicity evaluation of extracts from bamboo plants. J. Med. Plant Res. 2015, 9, 681–692. [Google Scholar] [PubMed]
- Choi, M.H.; Jo, H.G.; Yang, J.H.; Ki, S.H.; Shin, H.J. Antioxidative and Anti-Melanogenic Activities of Bamboo Stems (Phyllostachys nigra variety henosis) via PKA/CREB-Mediated MITF Downregulation in B16F10 Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 409. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, K.; Cederbaum, A.I. Oxidative stress and cytotoxicity induced by ferric-nitrilotriacetate in HepG2 cells that express cytochrome P450 2E1. Mol. Pharmacol. 1998, 54, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Kim, K.M.; Kim, M.G.; Seo, K.H.; Han, J.Y.; Ka, S.O.; Park, B.H.; Shin, S.M.; Ku, S.K.; Cho, I.J.; et al. Role of sestrin2 in the regulation of proinflammatory signaling in macrophages. Free Radic. Biol. Med. 2015, 78, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, K.M.; Yang, J.H.; Cho, S.S.; Kim, J.Y.; Park, S.J.; Lee, S.K.; Ku, S.K.; Cho, I.J.; Ki, S.H. Sestrin2 protects against acetaminophen-induced liver injury. Chem. Biol. Interact. 2017, 269, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Yang, J.H.; Shin, B.Y.; Seo, K.; Shin, S.M.; Cho, I.J.; Ki, S.H. Resveratrol inhibits LXRalpha-dependent hepatic lipogenesis through novel antioxidant Sestrin2 gene induction. Toxicol. Appl. Pharmacol. 2013, 271, 95–105. [Google Scholar] [CrossRef]
- Shin, B.Y.; Jin, S.H.; Cho, I.J.; Ki, S.H. Nrf2-ARE pathway regulates induction of Sestrin-2 expression. Free Radic. Biol. Med. 2012, 53, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.L.; Baek, S.Y.; Lee, E.H.; Lee, J.H.; Lee, S.G.; Kim, K.Y.; Jang, M.H.; Park, M.H.; Kim, J.H.; Kim, K.J.; et al. Liqustri lucidi Fructus inhibits hepatic injury and functions as an antioxidant by activation of AMP-activated protein kinase in vivo and in vitro. Chem. Biol. Interact. 2017, 262, 57–68. [Google Scholar] [CrossRef]
- Kim, Y.W.; Lee, S.M.; Shin, S.M.; Hwang, S.J.; Brooks, J.S.; Kang, H.E.; Lee, M.G.; Kim, S.C.; Kim, S.G. Efficacy of sauchinone as a novel AMPK-activating lignan for preventing iron-induced oxidative stress and liver injury. Free Radic. Biol. Med. 2009, 47, 1082–1092. [Google Scholar] [CrossRef]
- Jung, E.H.; Jung, J.Y.; Ko, H.L.; Kim, J.K.; Park, S.M.; Jung, D.H.; Park, C.A.; Kim, Y.W.; Ku, S.K.; Cho, I.J.; et al. Tryptanthrin prevents oxidative stress-mediated apoptosis through AMP-activated protein kinase-dependent p38 mitogen-activated protein kinase activation. Arch. Pharmacal Res. 2017, 40, 1071–1086. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Jiang, Y.F.; Ponnusamy, M.; Diallo, M. Role of Nrf2 in chronic liver disease. World J. Gastroenterol. 2014, 20, 13079–13087. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.J.; Lee, M.; Shin, T.; Yoon, N.; Kim, J.H.; Kim, H.R. eckol enhances heme oxygenase-1 expression through activation of Nrf2/JNK pathway in HepG2 cells. Molecules 2014, 19, 15638–15652. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martin, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin induces NF-kappaB, activator protein-1 (AP-1) and nuclear transcription factor erythroid 2p45-related factor-2 (Nrf2) via phosphatidylinositol-3-kinase/protein kinase B (PI3K/AKT) and extracellular regulated kinase (ERK) signalling in HepG2 cells. Br. J. Nutr. 2010, 103, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Rana, S.; Patial, V.; Gupta, M.; Bhushan, S.; Padwad, Y. Antioxidant and hepatoprotective effect of polyphenols from apple pomace extract via apoptosis inhibition and Nrf2 activation in mice. Hum. Exp. Toxicol. 2016, 35, 1264–1275. [Google Scholar] [CrossRef] [PubMed]
- Balboa, M.A.; Balsinde, J. Oxidative stress and arachidonic acid mobilization. Biochim. Biophys. Acta 2006, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kim, Y.W.; Kim, S.G. AMPK-mediated GSK3beta inhibition by isoliquiritigenin contributes to protecting mitochondria against iron-catalyzed oxidative stress. Biochem. Pharmacol. 2010, 79, 1352–1362. [Google Scholar] [CrossRef]
- Rungratanawanich, W.; Memo, M.; Uberti, D. Redox Homeostasis and Natural Dietary Compounds: Focusing on Antioxidants of Rice (Oryza sativa L.). Nutrients 2018, 10, 1605. [Google Scholar] [CrossRef]
- Sen, C.K.; Packer, L. Antioxidant and redox regulation of gene transcription. FASEB J. 1996, 10, 709–720. [Google Scholar] [CrossRef]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Wei, M.; Zheng, Z.; Shi, L.; Jin, Y.; Ji, L. Natural Polyphenol Chlorogenic Acid Protects Against Acetaminophen-Induced Hepatotoxicity by Activating ERK/Nrf2 Antioxidative Pathway. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 162, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef] [PubMed]
- Zulkawi, N.; Ng, K.H.; Zamberi, R.; Yeap, S.K.; Jaganath, I.B.; Satharasinghe, D.; Yong, C.Y.; Jamaluddin, A.B.; Tan, S.W.; Ho, W.Y. The in vivo hepato-recovery effects of the polyphenol-rich fermented food Xeniji™ on ethanol-induced liver damage. RSC Adv. 2017, 7, 38287–38299. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Vehicle | PHZ | PHZ + PN3 250 mg/kg | PHZ + PN3 500 mg/kg | |
|---|---|---|---|---|---|
| Index (Unit) | |||||
| DE regions (%/mm2) | 2.69 ± 2.21 | 59.93 ± 12.69 ** | 36.17 ± 11.09 ## | 11.00 ± 3.50 ## | |
| DE hepatocytes (cells/1000 hepatocytes) | 32.50 ± 25.62 | 587.50 ± 107.89 ** | 297.33 ± 115.40 ## | 99.33 ± 26.44 ## | |
| Inflammatory cells (cells/mm2) | 23.83 ± 14.66 | 196.00 ± 93.34 ** | 92.83 ± 24.73 ## | 63.67 ± 21.92 ## | |
| Immunopositive cells (cells/1000 hepatocytes) | |||||
| 4-Hydroxynonenal | 42.00 ± 21.16 | 608.33 ± 179.78 ** | 265.00 ± 73.54 ## | 95.67 ± 19.29 ## | |
| Cyclooxygenase-2 | 3.67 ± 3.27 | 324.17 ± 143.76 ** | 88.83 ± 19.77 ## | 42.17 ± 23.16 ## | |
| Ferritin | 75.67 ± 29.38 | 566.17 ± 123.01 ** | 288.50 ± 78.42 ## | 141.67 ± 50.25 ## | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.H.; Choi, M.-H.; Na, C.-S.; Cho, S.S.; Kim, J.H.; Ku, S.K.; Cho, I.J.; Shin, H.-J.; Ki, S.H. Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury. Nutrients 2019, 11, 114. https://doi.org/10.3390/nu11010114
Yang JH, Choi M-H, Na C-S, Cho SS, Kim JH, Ku SK, Cho IJ, Shin H-J, Ki SH. Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury. Nutrients. 2019; 11(1):114. https://doi.org/10.3390/nu11010114
Chicago/Turabian StyleYang, Ji Hye, Moon-Hee Choi, Chang-Su Na, Sam Seok Cho, Jae Hoon Kim, Sae Kwang Ku, Il Je Cho, Hyun-Jae Shin, and Sung Hwan Ki. 2019. "Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury" Nutrients 11, no. 1: 114. https://doi.org/10.3390/nu11010114
APA StyleYang, J. H., Choi, M.-H., Na, C.-S., Cho, S. S., Kim, J. H., Ku, S. K., Cho, I. J., Shin, H.-J., & Ki, S. H. (2019). Bamboo Stems (Phyllostachys nigra variety henosis) Containing Polyphenol Mixtures Activate Nrf2 and Attenuate Phenylhydrazine-Induced Oxidative Stress and Liver Injury. Nutrients, 11(1), 114. https://doi.org/10.3390/nu11010114