Zn-DTSM, A Zinc Ionophore with Therapeutic Potential for Acrodermatitis Enteropathica?



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
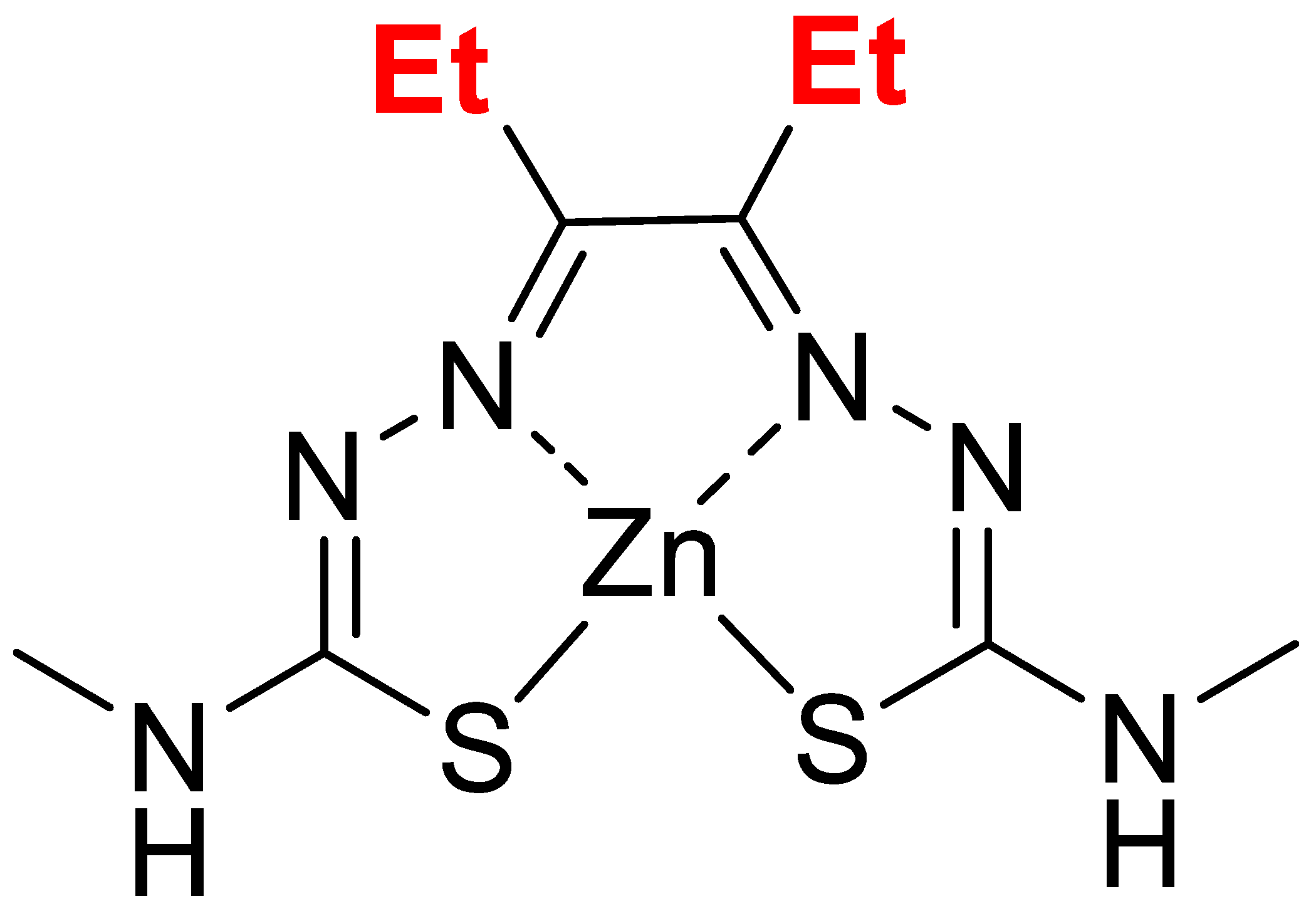
2.1. Compound
2.2. Ionophore Assay and Metal Analyses
2.3. Ethics Statement
2.4. Animals and Experimental Details
2.5. Statistical Analysis
3. Results
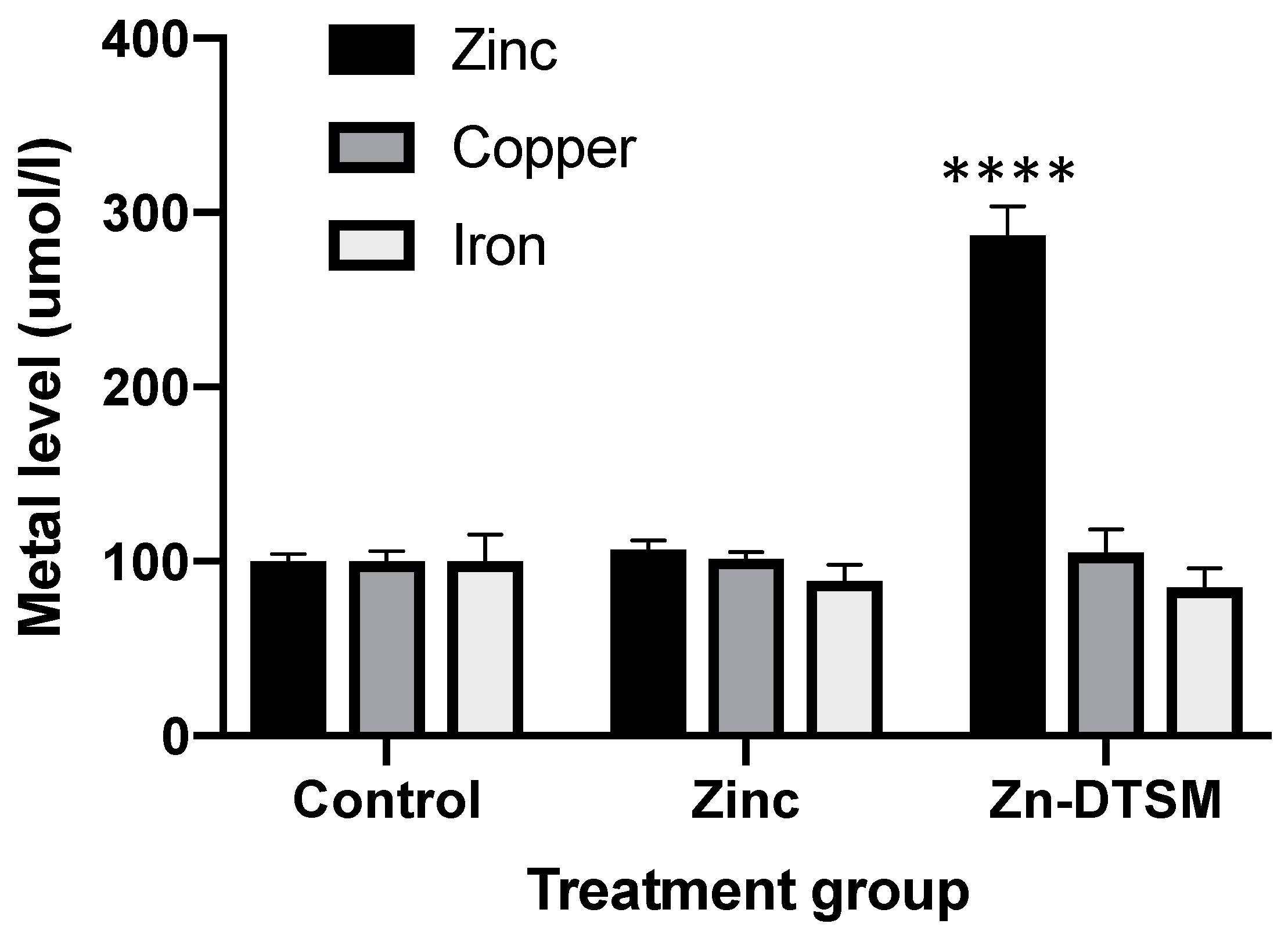
3.1. Zn-DTSM Is A Metal Ionophore
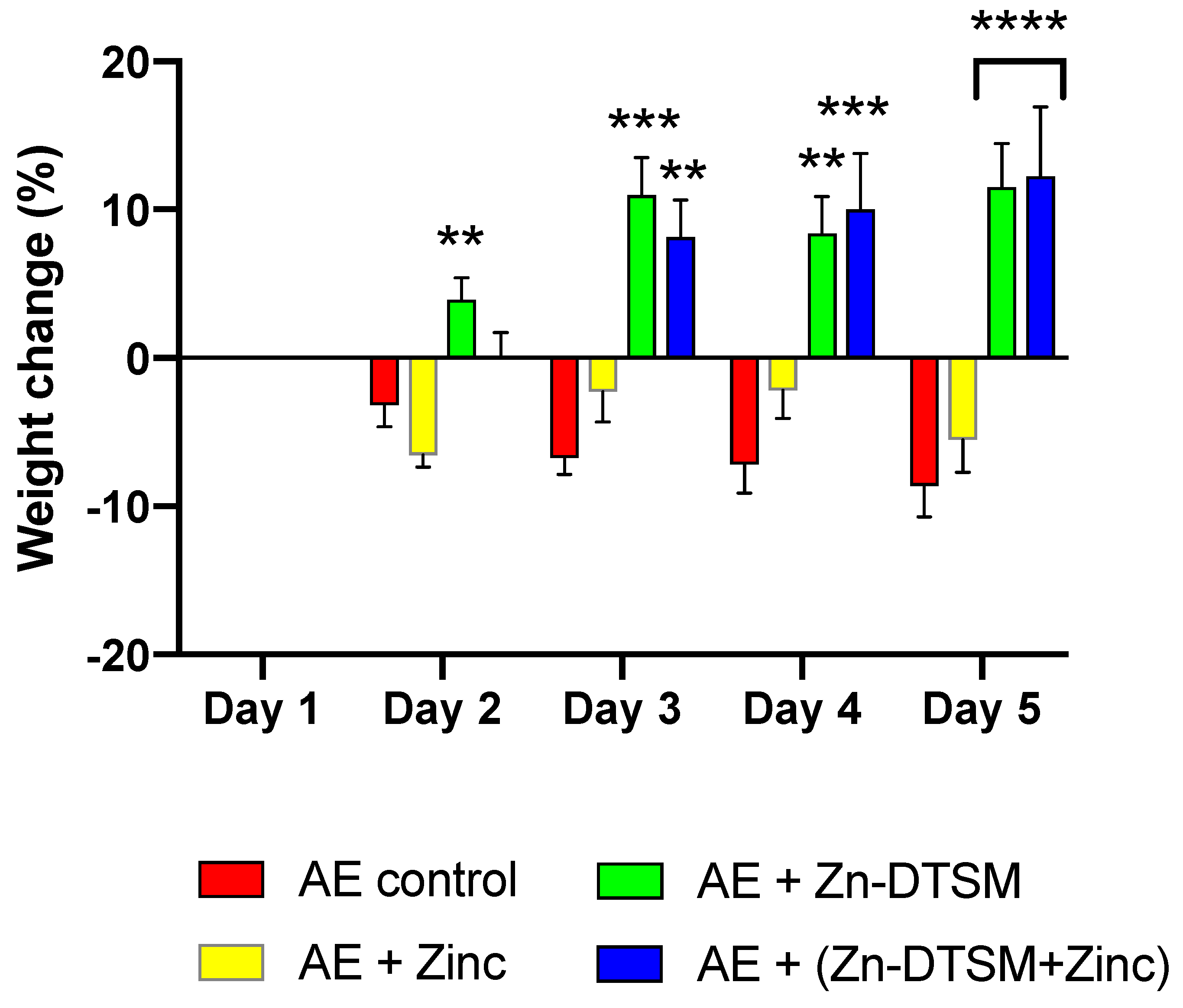
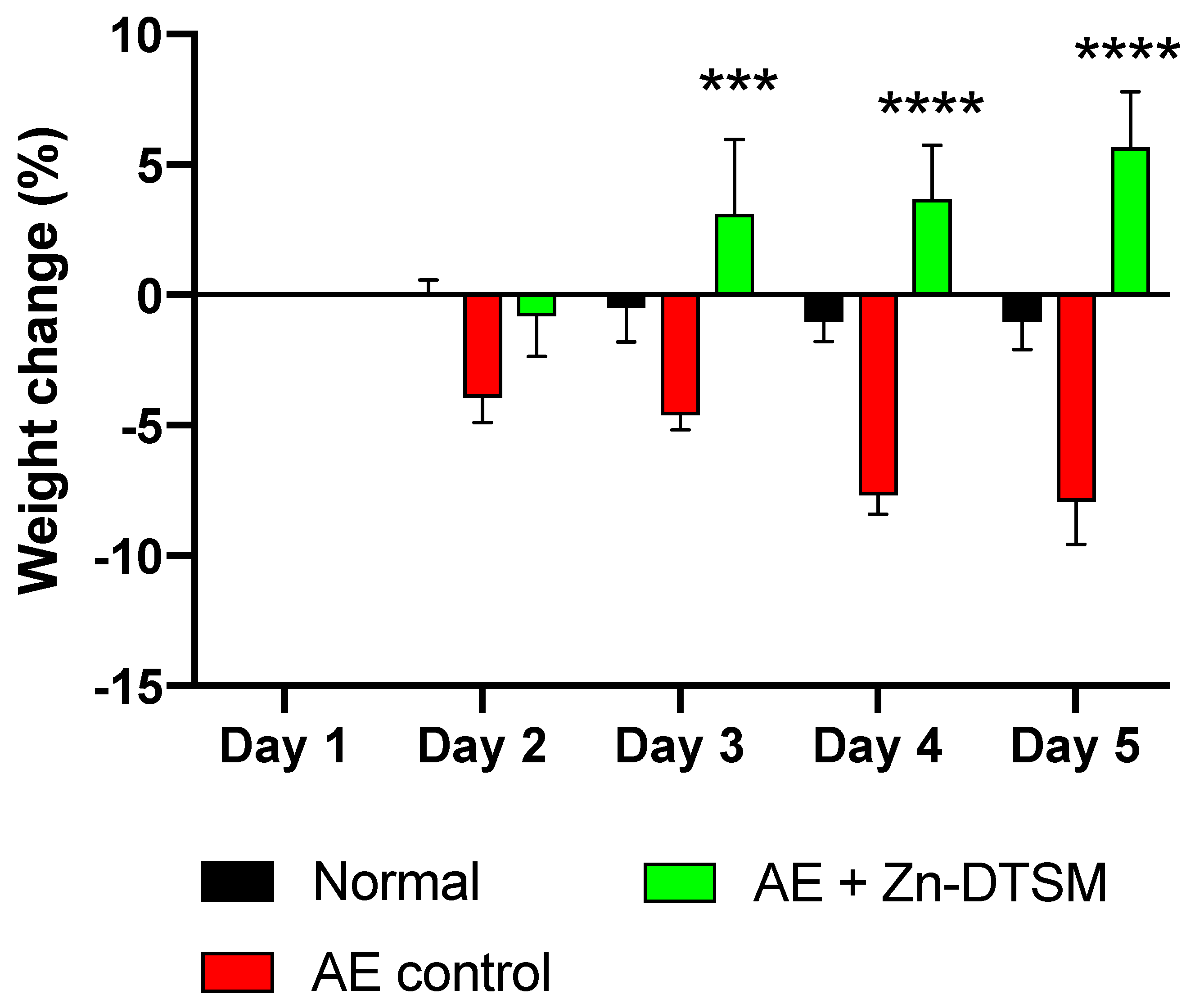
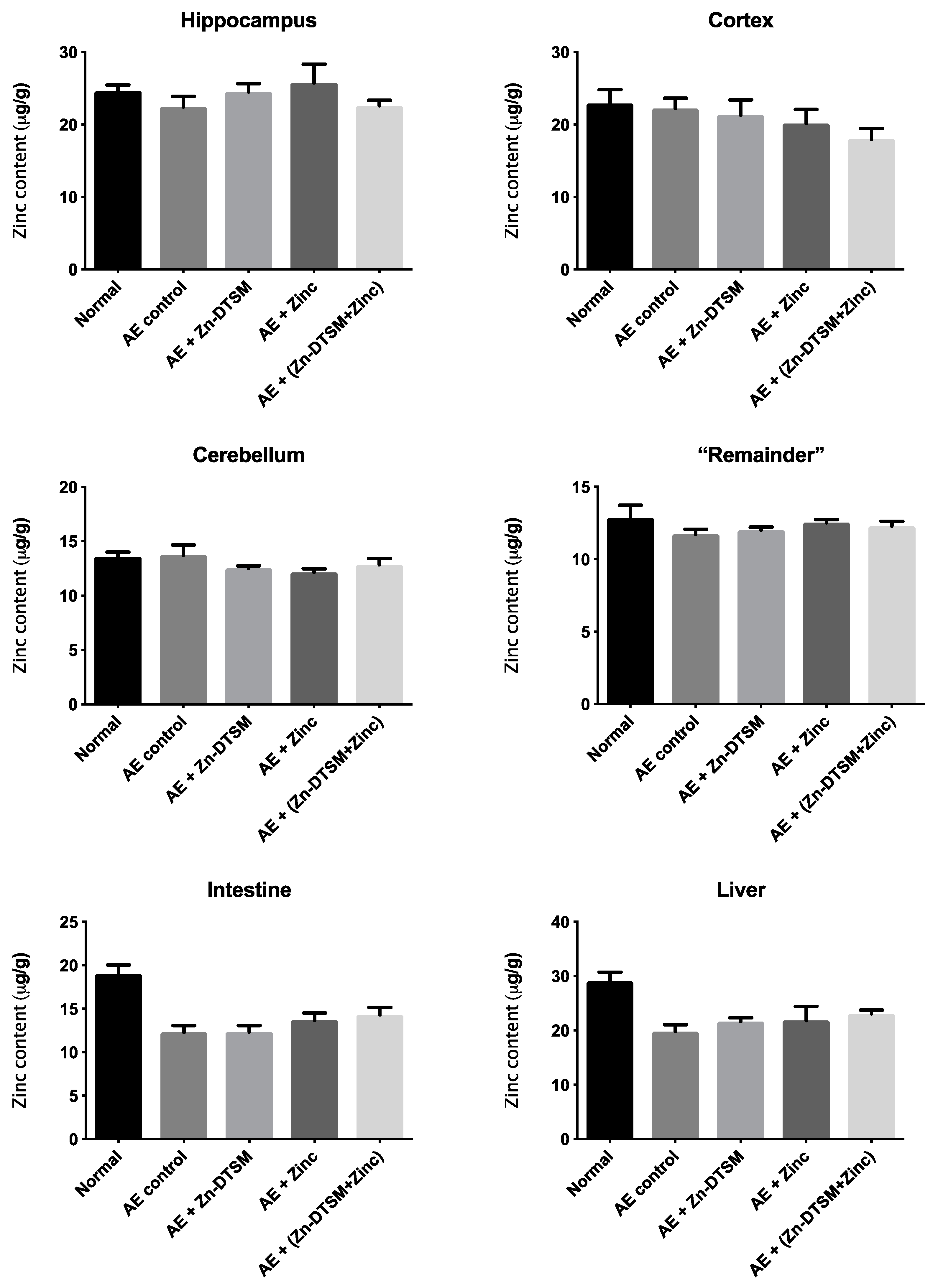
3.2. Zn-DTSM (30 mg/kg/d) Prevents Weight Loss in Young (1.5-Month) and Older (5-Month) AE Mice
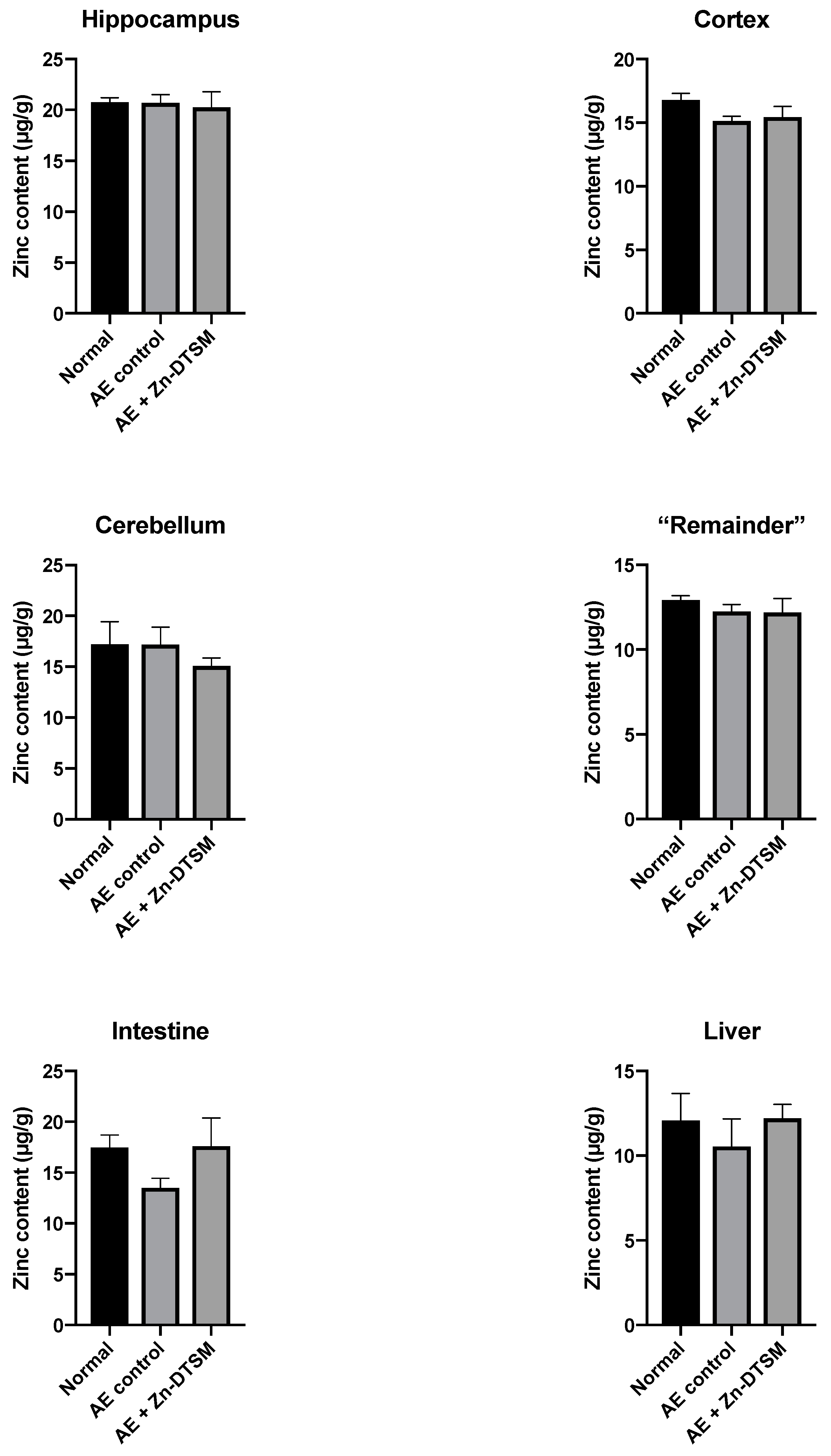
3.3. Low Dose Zn-DTSM (3 mg/kg/day) Requires Exogenous Zinc to Prevent Weight Loss in Older (5-Month) AE Mice
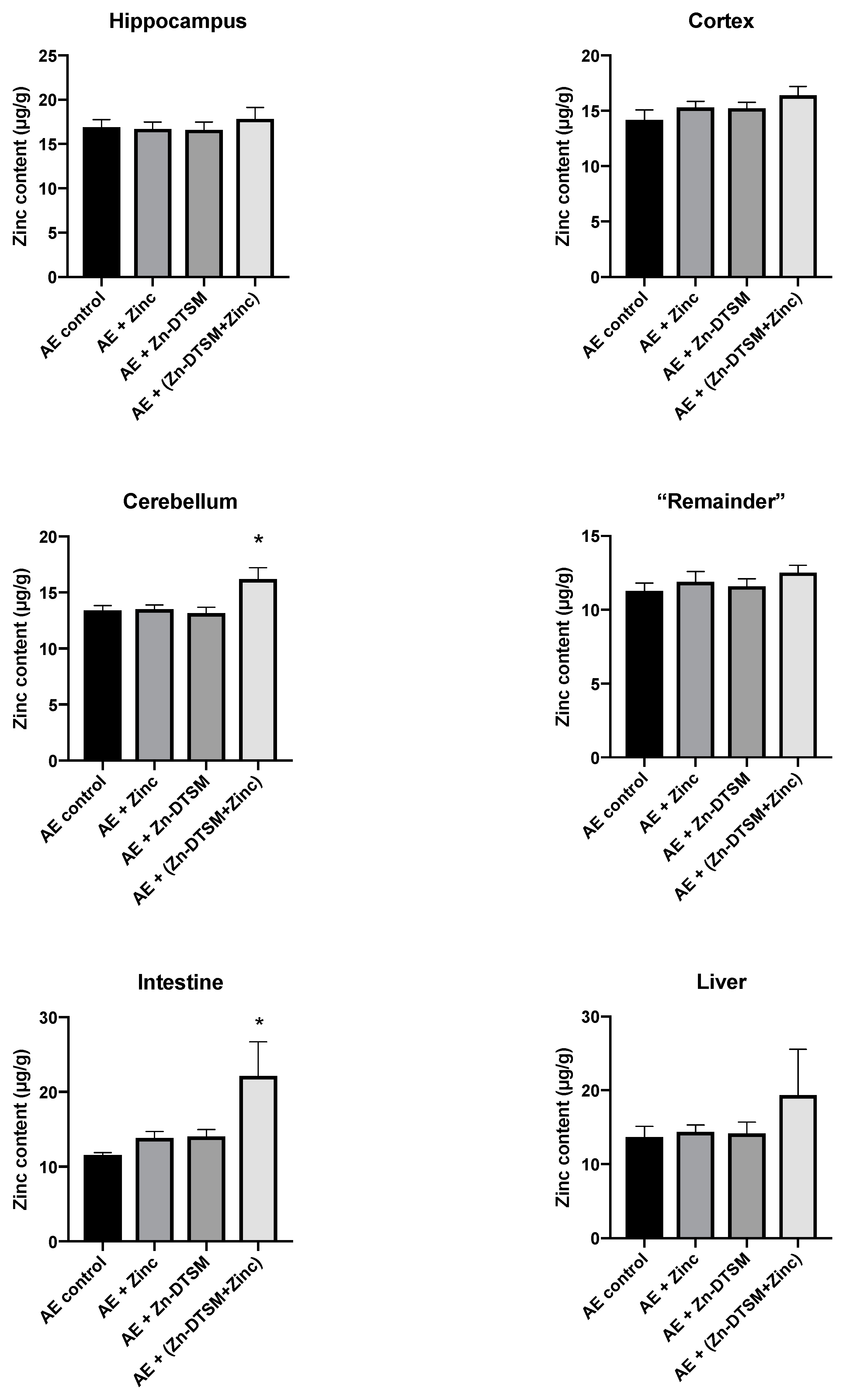
3.4. Acute Zn-DTSM Treatment Has Modest Effects on Tissue Zinc Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Danbolt, N. Acrodermatitis enteropathica. Br. J. Dermatol. 1979, 100, 37–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillaha, C.J.; Lorincz, A.L.; Aavik, O.R. Acrodermatitis enteropathica: Review of the literature and report of a case successfully treated with diodoquin. J. Am. Med. Assoc. 1953, 152, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Moynahan, E.J. Letter: Acrodermatitis enteropathica: A lethal inherited human zinc-deficiency disorder. Lancet 1974, 2, 399–400. [Google Scholar] [CrossRef]
- Atherton, D.J.; Muller, D.P.; Aggett, P.J.; Harries, J.T. A defect in zinc uptake by jejunal biopsies in acrodermatitis enteropathica. Clin Sci (Lond) 1979, 56, 505–507. [Google Scholar] [CrossRef] [PubMed]
- Lombeck, T.; Schnippering, H.G.; Ritzl, F.; Feinendegen, L.E.; Bremer, H.J. Letter: Absorption of zinc in acrodermatitis enteropathica. Lancet 1975, 1, 855. [Google Scholar] [CrossRef]
- Dufner-Beattie, J.; Kuo, Y.M.; Gitschier, J.; Andrews, G.K. The adaptive response to dietary zinc in mice involves the differential cellular localization and zinc regulation of the zinc transporters zip4 and zip5. J. Biol. Chem. 2004, 279, 49082–49090. [Google Scholar] [CrossRef] [PubMed]
- Dufner-Beattie, J.; Wang, F.; Kuo, Y.M.; Gitschier, J.; Eide, D.; Andrews, G.K. The acrodermatitis enteropathica gene zip4 encodes a tissue-specific, zinc-regulated zinc transporter in mice. J. Biol. Chem. 2003, 278, 33474–33481. [Google Scholar] [CrossRef]
- Kury, S.; Dreno, B.; Bezieau, S.; Giraudet, S.; Kharfi, M.; Kamoun, R.; Moisan, J.P. Identification of slc39a4, a gene involved in acrodermatitis enteropathica. Nat. Genet. 2002, 31, 239–240. [Google Scholar] [CrossRef]
- Wang, F.; Kim, B.E.; Dufner-Beattie, J.; Petris, M.J.; Andrews, G.; Eide, D.J. Acrodermatitis enteropathica mutations affect transport activity, localization and zinc-responsive trafficking of the mouse zip4 zinc transporter. Hum. Mol. Genet. 2004, 13, 563–571. [Google Scholar] [CrossRef]
- Kasana, S.; Din, J.; Maret, W. Genetic causes and gene-nutrient interactions in mammalian zinc deficiencies: Acrodermatitis enteropathica and transient neonatal zinc deficiency as examples. J. Trace. Elem. Med. Biol. 2015, 29, 47–62. [Google Scholar] [CrossRef]
- Hammersen, J.; Has, C.; Galiano, M.; Lindner, M.; Rossi, R.; Kohlhase, J.; Schneider, H. Sustained need for high-dose zinc supplementation in children with acrodermatitis enteropathica. Clin. Pediatr. (Phila) 2018, 57, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace. Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Hoogenraad, T.U.; Dekker, A.W.; van den Hamer, C.J. Copper responsive anemia, induced by oral zinc therapy in a patient with acrodermatitis enteropathica. Sci. Total Environ. 1985, 42, 37–43. [Google Scholar] [CrossRef]
- Jamall, I.S.; Ally, K.M.; Yusuf, S. Acrodermatitis enteropathica: Zinc therapy and possible identification of a carrier state through multiple hair zinc analyses over three decades. Biol. Trace Elem. Res. 2006, 114, 93–105. [Google Scholar] [CrossRef]
- Strain, W.H.; Hirsh, F.S.; Michel, B. Letter: Increased copper/zinc ratios in acrodermatitis enteropathica. Lancet. 1975, 1, 1196–1197. [Google Scholar] [CrossRef]
- Geiser, J.; Venken, K.J.; De Lisle, R.C.; Andrews, G.K. A mouse model of acrodermatitis enteropathica: Loss of intestine zinc transporter zip4 (slc39a4) disrupts the stem cell niche and intestine integrity. PLoS Genet. 2012, 8, e1002766. [Google Scholar] [CrossRef]
- Geiser, J.; De Lisle, R.C.; Finkelstein, D.; Adlard, P.A.; Bush, A.I.; Andrews, G.K. Clioquinol synergistically augments rescue by zinc supplementation in a mouse model of acrodermatitis enteropathica. PLoS One 2013, 8, e72543. [Google Scholar] [CrossRef]
- Adlard, P.A.; Bica, L.; White, A.R.; Nurjono, M.; Filiz, G.; Crouch, P.J.; Donnelly, P.S.; Cappai, R.; Finkelstein, D.I.; Bush, A.I. Metal ionophore treatment restores dendritic spine density and synaptic protein levels in a mouse model of alzheimer’s disease. PLoS One 2011, 6, e17669. [Google Scholar] [CrossRef]
- Adlard, P.A.; Bush, A.I. Metal chaperones: A holistic approach to the treatment of alzheimer’s disease. Front Psychiatry 2012, 3, 15. [Google Scholar] [CrossRef]
- Adlard, P.A.; Cherny, R.A.; Finkelstein, D.I.; Gautier, E.; Robb, E.; Cortes, M.; Volitakis, I.; Liu, X.; Smith, J.P.; Perez, K.; et al. Rapid restoration of cognition in alzheimer’s transgenic mice with 8-hydroxy quinoline analogs is associated with decreased interstitial abeta. Neuron 2008, 59, 43–55. [Google Scholar] [CrossRef]
- Adlard, P.A.; Parncutt, J.; Lal, V.; James, S.; Hare, D.; Doble, P.; Finkelstein, D.I.; Bush, A.I. Metal chaperones prevent zinc-mediated cognitive decline. Neurobiol. Dis. 2015, 81, 196–202. [Google Scholar] [CrossRef]
- Adlard, P.A.; Sedjahtera, A.; Gunawan, L.; Bray, L.; Hare, D.; Lear, J.; Doble, P.; Bush, A.I.; Finkelstein, D.I.; Cherny, R.A. A novel approach to rapidly prevent age-related cognitive decline. Aging Cell 2014, 13, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Cherny, R.A.; Ayton, S.; Finkelstein, D.I.; Bush, A.I.; McColl, G.; Massa, S.M. Pbt2 reduces toxicity in a c. Elegans model of polyq aggregation and extends lifespan, reduces striatal atrophy and improves motor performance in the r6/2 mouse model of huntington’s disease. J. Huntingtons Dis. 2012, 1, 211–219. [Google Scholar] [PubMed]
- Crouch, P.J.; Hung, L.W.; Adlard, P.A.; Cortes, M.; Lal, V.; Filiz, G.; Perez, K.A.; Nurjono, M.; Caragounis, A.; Du, T.; et al. Increasing cu bioavailability inhibits abeta oligomers and tau phosphorylation. Proc. Natl. Acad. Sci. USA 2009, 106, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Crouch, P.J.; Savva, M.S.; Hung, L.W.; Donnelly, P.S.; Mot, A.I.; Parker, S.J.; Greenough, M.A.; Volitakis, I.; Adlard, P.A.; Cherny, R.A.; et al. The alzheimer’s therapeutic pbt2 promotes amyloid-beta degradation and gsk3 phosphorylation via a metal chaperone activity. J. Neurochem. 2011, 119, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, D.I.; Billings, J.L.; Adlard, P.A.; Ayton, S.; Sedjahtera, A.; Masters, C.L.; Wilkins, S.; Shackleford, D.M.; Charman, S.A.; Bal, W.; et al. The novel compound pbt434 prevents iron mediated neurodegeneration and alpha-synuclein toxicity in multiple models of parkinson’s disease. Acta. Neuropathol. Commun. 2017, 5, 53. [Google Scholar] [CrossRef]
- Finkelstein, D.I.; Hare, D.J.; Billings, J.L.; Sedjahtera, A.; Nurjono, M.; Arthofer, E.; George, S.; Culvenor, J.G.; Bush, A.I.; Adlard, P.A. Clioquinol improves cognitive, motor function, and microanatomy of the alpha-synuclein ha53t transgenic mice. ACS Chem. Neurosci. 2016, 7, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Ayton, S.; Appukuttan, A.T.; Volitakis, I.; Adlard, P.A.; Finkelstein, D.I.; Bush, A.I. Clioquinol rescues parkinsonism and dementia phenotypes of the tau knockout mouse. Neurobiol. Dis. 2015, 81, 168–175. [Google Scholar] [CrossRef] [PubMed]
- McKenzie-Nickson, S.; Chan, J.; Perez, K.; Hung, L.W.; Cheng, L.; Sedjahtera, A.; Gunawan, L.; Adlard, P.A.; Hayne, D.J.; McInnes, L.E.; et al. Modulating protein phosphatase 2a rescues disease phenotype in neurodegenerative tauopathies. ACS Chem. Neurosci. 2018, 9, 2731–2740. [Google Scholar] [CrossRef]
- Portbury, S.D.; Sedjahtera, A.; Perrones, K.; Sgambelloni, C.; Zhang, M.; Crack, P.J.; Finkelstein, D.I.; Adlard, P.A. Metal chaperones: A novel therapeutic strategy for brain injury? Brain Inj. 2018, 3, 1–8. [Google Scholar] [CrossRef]
- Sedjahtera, A.; Gunawan, L.; Bray, L.; Hung, L.W.; Parsons, J.; Okamura, N.; Villemagne, V.L.; Yanai, K.; Liu, X.M.; Chan, J.; et al. Targeting metals rescues the phenotype in an animal model of tauopathy. Metallomics 2018, 10, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Cherny, R.A.; Atwood, C.S.; Xilinas, M.E.; Gray, D.N.; Jones, W.D.; McLean, C.A.; Barnham, K.J.; Volitakis, I.; Fraser, F.W.; Kim, Y.; et al. Treatment with a copper-zinc chelator markedly and rapidly inhibits beta-amyloid accumulation in alzheimer’s disease transgenic mice. Neuron 2001, 30, 665–676. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bray, L.; Volitakis, I.; Ayton, S.; Bush, A.I.; Adlard, P.A. Zn-DTSM, A Zinc Ionophore with Therapeutic Potential for Acrodermatitis Enteropathica? Nutrients 2019, 11, 206. https://doi.org/10.3390/nu11010206
Bray L, Volitakis I, Ayton S, Bush AI, Adlard PA. Zn-DTSM, A Zinc Ionophore with Therapeutic Potential for Acrodermatitis Enteropathica? Nutrients. 2019; 11(1):206. https://doi.org/10.3390/nu11010206
Chicago/Turabian StyleBray, Lisa, Irene Volitakis, Scott Ayton, Ashley I. Bush, and Paul A. Adlard. 2019. "Zn-DTSM, A Zinc Ionophore with Therapeutic Potential for Acrodermatitis Enteropathica?" Nutrients 11, no. 1: 206. https://doi.org/10.3390/nu11010206
APA StyleBray, L., Volitakis, I., Ayton, S., Bush, A. I., & Adlard, P. A. (2019). Zn-DTSM, A Zinc Ionophore with Therapeutic Potential for Acrodermatitis Enteropathica? Nutrients, 11(1), 206. https://doi.org/10.3390/nu11010206