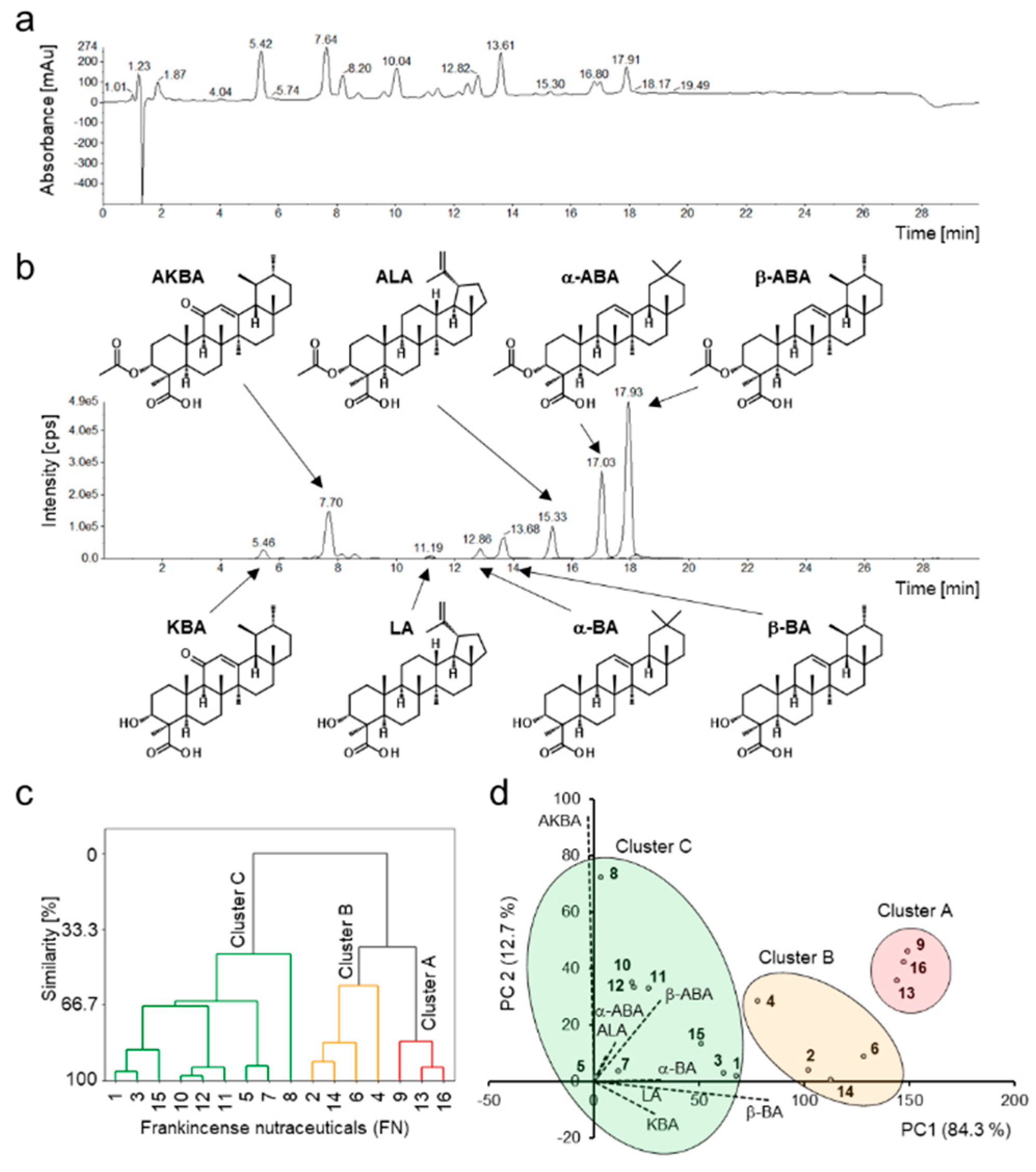
3.1. Compositions of Boswellic and Lupeolic Acids in Frankincense Nutraceuticals
Quantification of eight BAs and LAs including α-boswellic acid (α-BA), acetyl-α-boswellic acid (α-ABA), β-boswellic acid (β-BA), acetyl-β-boswellic acid (β-ABA), 11-keto-β-boswellic acid (KBA), acetyl-11-keto-β-boswellic acid (AKBA), lupeolic acid (LA), and acetyl-lupeolic acid (ALA), in 16 frankincense nutraceuticals (FNs) was performed by HPLC-MS/MS. The total contents (
w/w) of BAs and LAs varied from 0.4% to 35.7% (
Table 1). Related to the mass of a capsule or a pill, the total BA and LA contents varied from 3.5 to 157.3 mg/unit (
Table S1). Many manufacturers and distributers promote their products as those with boswellic acids contents over 80%. Interestingly, these concentrations could not be confirmed. Manufacturers usually determine the boswellic acid content of their products by non-aqueous titration, for example, using 0.1 N potassium methoxide and 0.3% (
w/v) thymol blue as the indicator [
27]. These methods estimate unselectively all acidic compounds, not only boswellic acids. Here, a highly selective and accurate quantification method for analysis of BAs and LAs by HPLC-MS/MS was applied (
Figure 2a,b). Consequently, the precise contents of the individual BAs and LAs in nutraceuticals could be determined (
Table 1 and
Table S1).
In a previous study, we showed that oleogum resins of the species
B. serrata are characterized by higher proportions of deacetylated compounds compared to acetylated compounds [
18]. This pattern was also observed in the FN samples analyzed here, which contained mainly extracts from
B. serrata. Unusually, particularly for
B. serrata, sample FN8 exhibited an exceptionally high content of AKBA. Especially, the concurrence of the high content of AKBA and very low contents of the other BAs and LAs indicated a special enrichment procedure putting FN8 apart from the other samples. Meins et al. observed the same uncommon ratio between AKBA content and total BAs content in a product containing the same 5-Loxin
® extract [
28]. FN5 and FN7 showed the lowest contents of BAs and LAs of all investigated samples, with total BAs and LAs contents of 2.9% for FN5 and only 0.9% for FN7. In addition to the exceptionally low BAs and LAs contents, the large difference between the stated extract content of 400 mg for FN7 and the determined pill mass of 809.8 ± 5.0 mg points also to an oddly high amount of additives in FN7 (
Table 1 and
Table S1). According to the manufacturer, the AKBA content of FN7 is 52 mg per pill. However, analysis revealed an AKBA content <1 mg per pill. Hence, it can be assumed that either less
Boswellia extract than stated was processed or other
Boswellia oleogum resins containing little or no BAs and LAs were used like, for example,
B. frereana [
18,
29].
Multivariate statistical methods, cluster analysis and principal component analysis (PCA) were applied to the datasets to visualize the data and discover patterns of the BAs and LAs compositions in different FNs. Cluster analysis assigned the samples to three different clusters (
Figure 2c). Cluster A contained samples FN9, FN13, and FN16 with the highest total BAs and LAs contents of over 30%. Furthermore, these FNs showed high similarity regarding the composition of the individual components (
Figure 2d). Cluster B (samples FN2, FN4, FN6, and FN14) comprised samples with total BAs and LAs contents between 15% and 30%, and Cluster C (samples FN1, FN3, FN4, FN5, FN7, FN8, FN10, FN11, and FN12) contained less than 15% BAs and LAs. FN8 showed the lowest similarity to Cluster C (<50%) due to its unusual chemical composition with high AKBA concentration. Moreover, samples FN10, FN11, and FN12, which contained non-extracted oleogum resin, formed an additional subcluster, due to their similarity. Interestingly, these three samples contained oleogum resins of different
Boswellia species. According to the manufacturers, sample FN10 was made of
B. sacra, FN11 of
B. serrata, and FN12 of
B. carterii. Comparing with the BAs and LAs compositions determined in our previous studies [
9,
18], FN11 exhibited indeed general BAs and LAs patterns typical for
B. serrata resins, with a higher proportion of deacetylated BAs and LAs than acetylated ones. However, FN10 and FN12 showed BAs and LAs patterns untypical for
B. sacra or
B. carterii resins, but more similar to
B. serrata, too. Because these samples contain no extracts but untreated oleogum resin, these differences could not be caused by an extraction procedure. Therefore, it cannot be excluded that instead of
B. sacra or
B. carterii, possibly
B. serrata oleogum resin might have been used for manufacturing of FN10 and FN12.
3.2. Inhibition of Proinflammatory Cytokine Release by Frankincense Nutraceuticals
For comparative investigation of the immunomodulatory activities of the FNs, whole blood of healthy donors was treated with lipopolysaccharide (LPS) to initiate an acute inflammatory response and to induce the release of proinflammatory cytokines. Pretreatment with FNs significantly decreased the expression of the proinflammatory cytokine TNF-α compared to the LPS-treated control group (
Figure 3a). Interestingly, the production of the anti-inflammatory cytokine IL-10 was inhibited by the FNs, too. IL-10 acts to limit inflammatory responses by damping the uncontrolled production of proinflammatory cytokines including TNF-α [
30]. However, also TNF-α regulates the production of IL-10. Thereby, the chronological sequence of cytokine expression during inflammation is essential. While high levels of proinflammatory cytokines IL-1, IL-6, IL-8, and TNF-α are released already 4–8 h after LPS stimulation, maximal levels of the anti-inflammatory IL-10 are observed only 24–48 h after stimulation [
31]. Moreover, about 50–75% of IL-10 released by LPS-stimulated monocytes can be inhibited by anti-TNF-α [
32], indicating that TNF-α is responsible for the majority of the released IL-10. Therefore, the decreased production of TNF-α by blood treated with FNs would decrease the later IL-10 release (
Figure 3a).
The strong albumin-binding affinity of BAs complicates an accurate analysis of their immunomodulatory activity in whole blood [
33]. In addition, blood contains varying amounts of different lipids aiding samples’ solubility. Hence, analysis of variance (ANOVA) exhibited no significant differences between the samples or sample groups when analyzed in whole blood. As monocytes are the major producers of TNF-α and IL-10 in blood [
34], for further analysis, peripheral blood mononuclear cells (PMBC) were used and the effects of the FNs on the production of a panel of cytokines including TNF-α, IL-1β, IL-6, IL-8, IL-10, and IL-12p70, were analyzed. Contents of IL-12p70 were too low for an adequate evaluation and are not presented. FNs were used at a concentration of 10 µg/mL, which did not affect cell viability as analyzed by XTT and lactate dehydrogenase release (LDH) assays and by propidium iodide staining.
FNs with BAs and LAs contents >30% (Cluster A: FN9, FN13, and FN16) exhibited a significant inhibition of TNF-α, IL-6, and IL-8 expression compared to the control LPS group (
Figure 3b–f and
Table S2). Moreover, FN4 and FN6 from Cluster B showed considerable inhibitory activity towards TNF-α, IL-6, and IL-8, too. The most potent five nutraceuticals, FN4, FN6, FN9, FN13, and FN16, decreased the release of proinflammatory cytokines to 20.7% for TNF-α, 9.2% for IL-6, and 16.8% for IL-8 (average values for all five FNs). Interestingly, these five samples also had the highest concentrations of ALA, α-ABA, and β-ABA. The proteasome inhibitor MG132 used as a positive control decreased cytokine release to 15.5% ± 7.2% for TNF-α, 18.5% ± 7.7% for IL-1β, 5.2% ± 3.5% for IL-6, 20.2 ± 7.2% for IL-8, and 37.5% ± 14.8% for IL-10 compared to the control group. Differently, samples FN5 and FN7, that were previously shown to contain the lowest BAs and LAs contents, exhibited no inhibitory effects on cytokine production at all but even enhanced the expression of the proinflammatory cytokines TNF-α, IL-6, and IL-8 with average values of 142.4% for FN5 and 210.7% for FN7 compared to the control group. The reason for the increased secretion of cytokines might be contamination of these FNs with bacterial products.
Investigation of the correlation between the chemical composition and cytokine expression revealed that the total amount of BAs and LAs correlate positively with the inhibition of TNF-α, IL-6, IL-8, and IL-10 (
Table 2 and
Figure S1). Especially, the acetylated compounds ALA, α-ABA, and β-ABA exhibit the highest correlation coefficients, while the acetylated compound with keto group AKBA shows the lowest correlation. However, when FN8 with the extraordinarily high AKBA content was removed as an outlier (Grubbs test), AKBA contents in FNs exhibited significant correlation with all cytokines with
p = 0.007 for TNF-α,
p < 0.001 for IL-1β,
p = 0.011 for IL-6,
p = 0.006 for IL-8, and
p = 0.001 for IL-10. Inhibition of IL-1β correlated only with AKBA, ALA, and α-ABA contents in FNs. Moreover, the inhibition of all five investigated cytokines correlated with each other. Whereby, IL-1β, the only cytokine in the panel activated by the NLRP3 inflammasome [
35], showed the lowest correlation (
Table 2 and
Figure S1).
3.3. Cytotoxic Efficacy of Frankincense Nutraceuticals Against Triple-Negative Breast Cancer Cells In Vitro
FN16, one of the most potent nutraceuticals with respect to cytokine release, concentration-dependently inhibited the viability of the highly metastatic, treatment-resistant breast cancer cell line MDA-MB-231 in vitro. Interestingly, cancer cells were more sensitive to FN16 compared to normal human PBMC, providing evidence for selectivity against cancer cells (
Figure 4a). Also, FN1 with low contents of BAs and LAs, exhibited higher toxicity against MDA-MB-231 cells compared to PBMCs (
Figure S2a), though the difference was not as strong as for FN16. Therefore, all FNs were investigated for their toxicity against MDA-MB-231 breast cancer cells and the half maximal inhibitory concentrations (IC
50) were determined (
Table 3 and
Figure 4b). The FNs exhibited cytotoxicity against MDA-MB-231 cells with an average IC
50 of 15.9 µg/mL. The samples FN9, FN13, and FN16 (Cluster A) with the highest BAs and LAs contents exhibited also significantly higher cytotoxicity with IC
50 values for FN9, FN13, and FN16 of 7.0 µg/mL, 6.6 µg/mL, and 6.0 µg/mL, respectively, compared to the average IC
50 value of 15.9 µg/mL (
Figure 4b). Samples FN5 and FN7 (Cluster C) demonstrated the lowest cytotoxicity against MDA-MB-231 with an IC
50 = 44.3 µg/mL for FN5 and an IC
50 = 30.7 µg/mL for FN7. Thus, FN5 and FN7 not only exhibited the lowest contents of BAs and LAs and the poorest cytokine modulatory properties, but also the cytotoxic efficacies against MDA-MB-231 were significantly below average. Hence, this points to a correlation between BAs and LAs compositions and cytotoxicity against breast cancer cells. A combination of principal component analysis (PCA) of the BAs and LAs concentrations with a contour plot that visualizes the IC
50 values revealed correlation between the BAs and LAs contents and cytotoxic efficacy (
Figure 4c). Interestingly, in addition to similar total BAs and LAs contents and similar effects on cytokine release, the compositions of the individual BAs and LAs in the most potent nutraceuticals FN9, FN13, and FN16 were highly alike. Spearman’s rank correlation analysis revealed that contents of BAs and LAs without a keto group exhibit the highest correlation to FN cytotoxic efficacy (
Figure S1). In this regard, the content of the boswellic acid β-ABA shows the highest correlation to cytotoxicity against MDA-MB-231.
Furthermore, a principal component regression analysis (PCR) based on known BAs and LAs contents in FNs was used, to avoid biases caused by multicollinearity. Sample FN5 was identified as an outlier by the Grubbs test and excluded from the regression analysis. The resulting regression model exhibited a
R2 of 96.5% and a significance of correlation of
p < 0.001. The regression analysis yielded the following equations:
with
and
with concentrations of corresponding BA or LA in µg/mg FN given in square brackets.
The IC
50 values calculated by the regression model exhibited an average absolute residue of only ± 1.0 µg/mL compared to the experimentally-derived values (
Table 3). Hence, the regression equation could be used as a tool to predict cytotoxic efficacy of FNs and other frankincense herbal preparations with known BAs and LAs content. By expanding the data set, one could achieve an even more accurate prediction.
Further, the three FNs, FN9, FN13, and FN16, showing the highest cytotoxicity against MDA-MB-231 cells, were investigated for their cytotoxic efficacy against two additional TNBC cell lines, MDA-MB-453 and Cal-51. Likewise, the investigated samples exhibited considerable cytotoxicity against MDA-MB-453 with IC
50 values between 12.9 µg/mL and 17.3 µg/mL, and against Cal-51 with even lower IC
50 values between 3.8 µg/mL and 4.0 µg/mL (
Table 4). This indicates that FNs exhibit cytotoxic efficacy against different triple-negative treatment-resistant breast cancer cells.
The non-halogenated anthracycline doxorubicin, a chemotherapeutic agent used to treat patients with breast cancer, was analyzed as a positive control for FNs. Doxorubicin exhibited an IC
50 = 0.41 ± 0.03 µg/mL for MDA-MB-231, an IC
50 = 0.26 ± 0.02 µg/mL for MDA-MB-453, and an IC
50 = 0.033 ± 0.004 µg/mL for Cal-51. However, for doxorubicin, severe adverse effects including cardio- and nephrotoxicity have been reported [
36,
37]. Differently, for frankincense extracts, only mild adverse effects, like heartburn or nausea, have been described [
38].
3.4. Boswellic and Lupeolic Acids Inhibit Cytokine Release and Are Cytotoxic for TNBC Cells
Cytotoxicity of FNs against MDA-MB-231 cells correlated significantly to the total contents of BAs and LAs as well as to individual BAs and LAs contents, except for AKBA, in FNs (Spearman’s rank correlation,
p < 0.001) (
Figure S1). The highest correlation coefficient exhibited β-ABA, whereas no significant correlation was detected for AKBA. The poor correlation of AKBA was caused mainly by the unusual chemical composition of the sample FN8 with unexpectedly high AKBA content concomitantly with very low contents of other BAs and LAs. After rejecting sample FN8 from the statistical analysis, the AKBA contents of the remaining samples correlated significantly with the cytotoxicity against MD-MB-231 cells (
p = 0.004). Hence, we further investigated the cytotoxic efficacies of the pure individual BAs and LAs against MDA-MB-231 (
Table 5). The acetylated forms of boswellic acids were more effective against MDA-MB-231 cells than their deacetylated forms; and the most cytotoxic compounds were AKBA with IC
50 = 6.6 µM, α-ABA with IC
50 = 7.2 µM, and β-ABA with IC
50 = 5.9 µM. Differently, for lupeolic acids, LA was more cytotoxic compared to ALA. We have previously shown that acetylated BAs, similar to anticancer drugs such as camptothecins and podophyllotoxins, inhibit activities of human topoisomerases [
39]. Acetylated BAs also inhibit the activation of transcriptional factor NF-κB which regulates the synthesis of antiapoptotic proteins [
11,
17], whereas ALA is an inhibitor of the AKT kinase [
16]. These molecular targets might explain the high cytotoxicity of BAs and LAs against proliferating cancer cells.
Interestingly, similar to FN1 and FN16, AKBA, and in particular, β-ABA, showed higher toxicity against cancer cells compared to PBMC (
Figure S2b,c), suggesting selectivity against cancer cells.
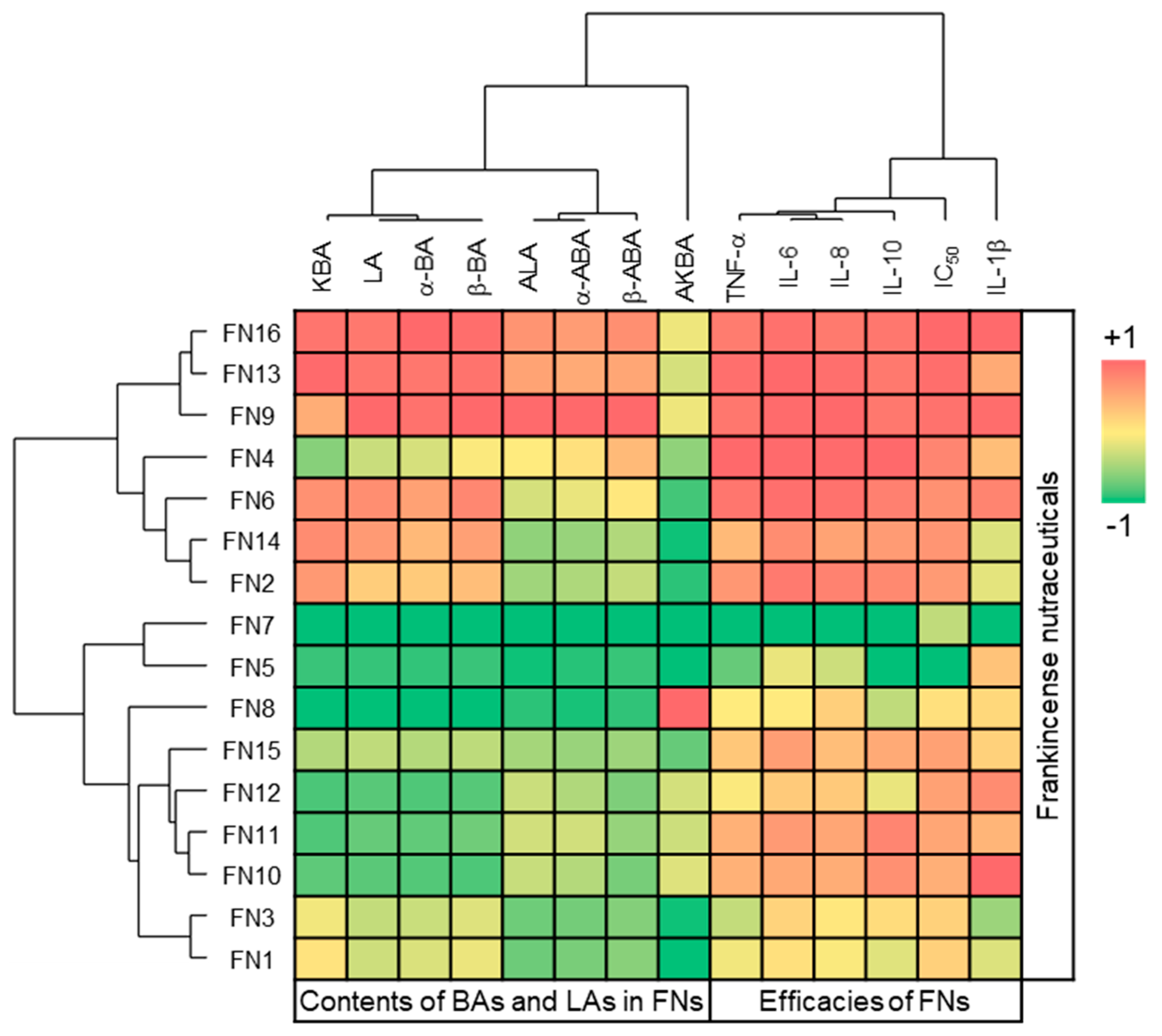
Moreover, toxicity against breast cancer cells strongly correlated with cytokine inhibition with
p = 0.023 for IL-1β and
p < 0.001 for TNF-α, IL-6, IL-8, and IL-10 (
Figure S1). Cluster analysis of variables confirmed the correlation between toxicity against breast cancer cells and cytokine inhibition (
Figure 5). In addition to the activation of genes coding for antiapoptotic proteins and for those necessary for continuous cell proliferation, NF-κB activation is central to the activation of cytokine and chemokine gene expression [
40]. In previous studies, we have shown that acetylated boswellic acids AKBA and β-ABA specifically inhibited the activities of human IκB kinases (IKK) and the subsequent release of proinflammatory cytokines by human monocytes [
11]. Moreover, intercepting the IKK activity by AKBA and β-ABA inhibited proliferation and promoted apoptosis in androgen-independent PC-3 prostate cancer cells in vitro and in vivo [
17]. Hence, inhibition of NF-κB by boswellic acids might represent a clue as to why these compounds inhibit both inflammatory processes and cancer growth.
Interestingly, chronic inflammation is known to increase cancer risk. Particularly, activation of NF-κB and production of inflammatory cytokines by tumor-associated immune cells are important components aiding tumor initiation, growth, malignant transformation, invasion, and metastasis [
41]. Hence, inhibition of inflammatory mediators prevents the development of experimental cancers, and treatment with anti-inflammatory drugs reduces cancer risks and progression [
42]. Hence, inhibition of inflammation by frankincense constituents could benefit anticancer therapy.
Similar to FNs, pure BAs and LAs, in concentrations which do not affect PBMC viability, inhibited cytokine production by LPS-activated PBMC, but not as efficiently as FNs (
Table S3). Although BAs and LAs are identified here as the active principle of FNs, frankincense extracts also contain a rather complex lipidome including a large number of different fatty acids [
43]. The lipids are extracted from frankincense during extract preparation and are persistent components of FNs. Such lipids can act as a natural emulsifier for nonpolar compounds like BAs and LAs and increase their bioavailability [
44]. Accordingly, the intake of a FN in combination with a high-fat meal improved the bioavailability of BAs in humans [
45]. Hence, the intake of pure BAs and LAs in the absence of solubility enhancers might decrease their bioavailability and worsen the therapeutic response. This should be considered in the development of drugs based on BAs and LAs. Formulations with lecithin for oral administration and complexation with cyclodextrins or liposomes for injections can be beneficial [
46,
47].
3.5. Inhibition of Proliferation and Induction of Apoptosis in Breast Cancer Xenografts
The antitumor activity of FNs and BAs was further verified in vivo, in MDA-MB-231 breast cancer xenografts grown on the chorioallantoic membrane (CAM) of fertilized chick eggs. Xenografts were treated with FN16, because it contains high amounts of BAs and LAs and exhibited the highest cytotoxicity against MDA-MB-231 cells in vitro. Also, two acetylated BAs, AKBA and β-ABA, which exhibited the highest cytotoxic efficacy in vitro, were tested in an in vivo model.
Treatment of cancer xenografts with FN16 reduced dose-dependently and significantly the tumor volume compared to the control group (
Figure 6a,b). Also, treatment with β-ABA significantly reduced the tumor volume. AKBA-treated xenografts were only non-significantly smaller than control. Immunohistochemical analysis of the tumors revealed that FN16 had a dose-dependent inhibitory effect on cancer cell proliferation (
Figure 6c). Also, AKBA and β-ABA significantly inhibited cancer cell proliferation. FN16, AKBA, and β-ABA all induced apoptosis in TNBC xenografts as shown by analysis of apoptosis by using the TUNEL technique (
Figure 6d). Whilst the chemotherapeutic drug doxorubicin exhibited eminent cytotoxicity against MDA-MB-231 cells in vitro, in vivo its efficacy was not sufficient to induce a significant reduction of tumor size or to induce apoptosis during the treatment period. Notably, no systemic toxicity of the FN16 or BAs on the chick embryos was observed.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}