Plasma Cholesterol-Lowering Activity of Soybean Germ Phytosterols

Abstract
:1. Introduction
2. Materials and Methods
2.1. SGP Preparation
2.2. Diets
2.3. Hamsters
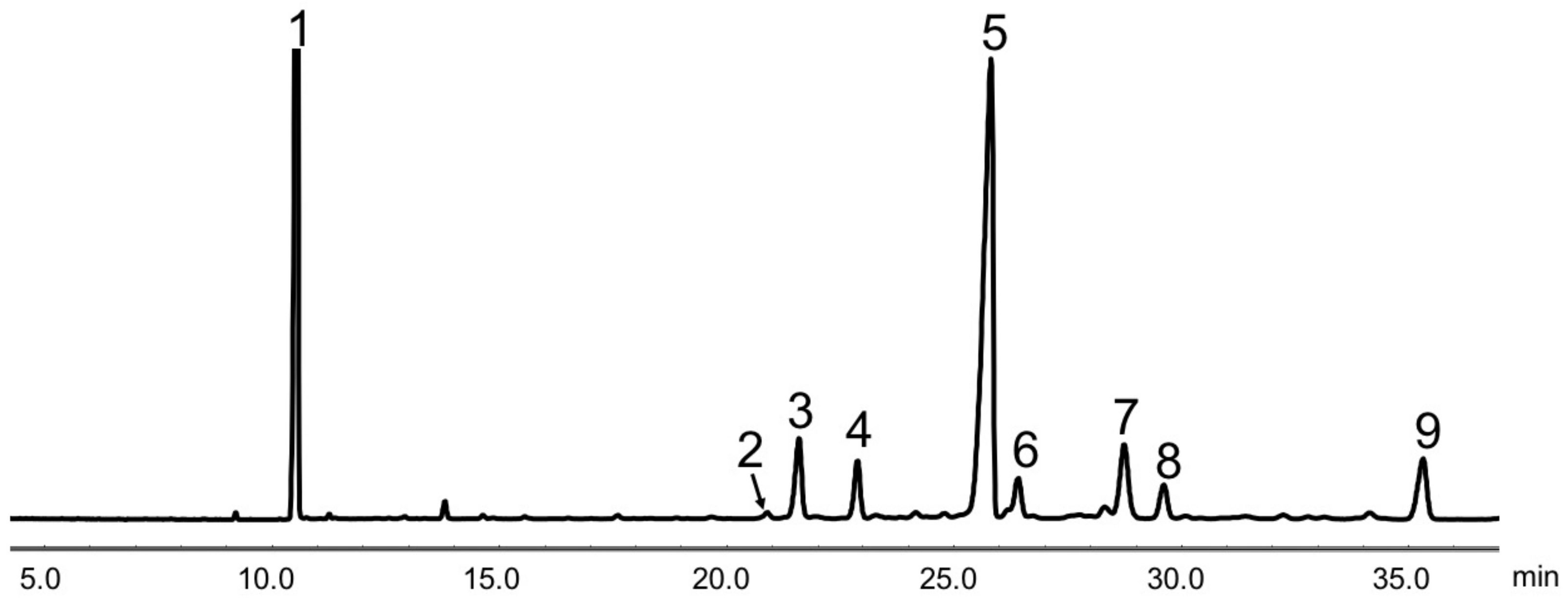
2.4. Analysis of Phytosterols
2.5. Measurement of Plasma Lipoproteins
2.6. Analysis of Atherosclerotic Plaque
2.7. Determination of Liver Cholesterol
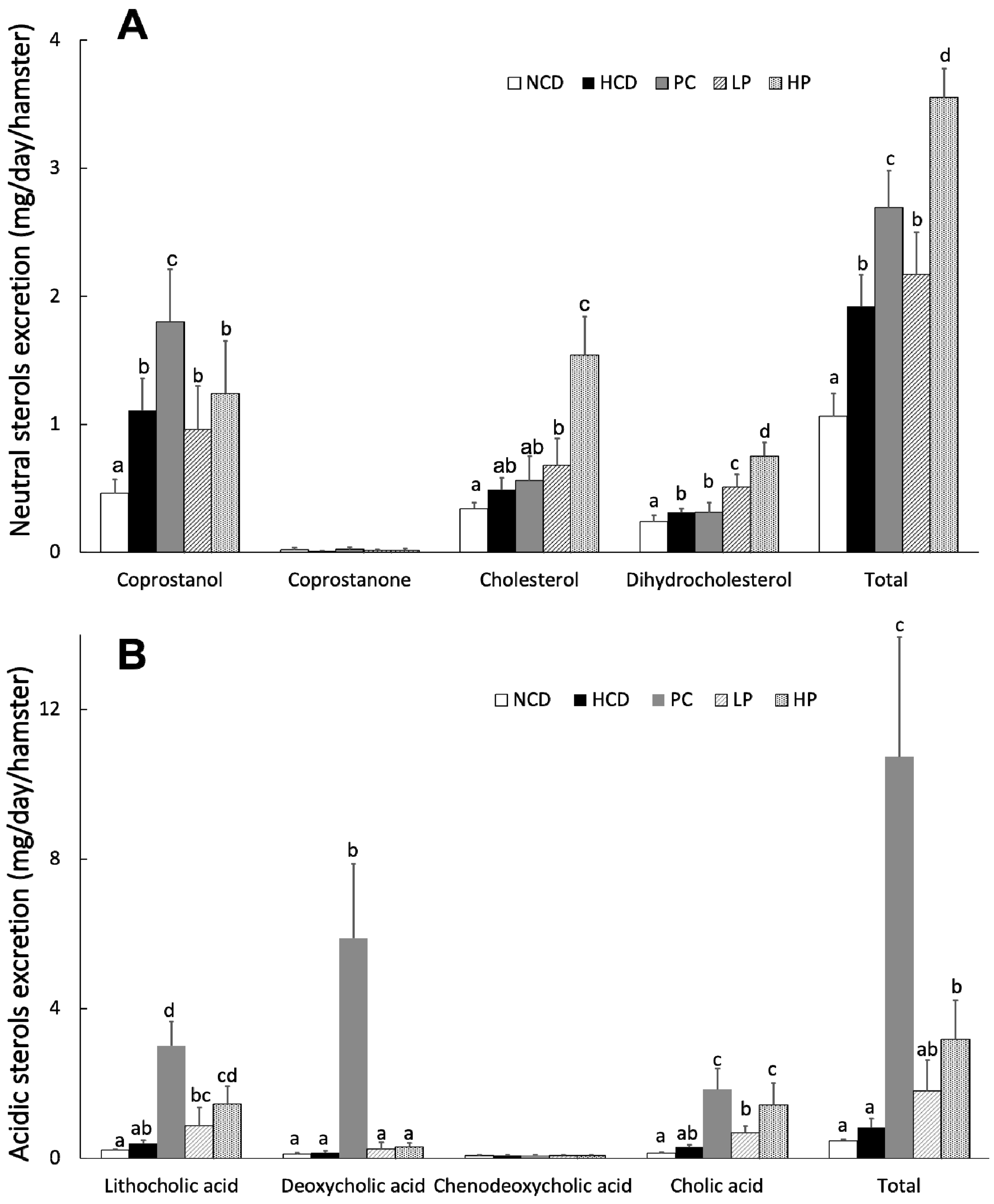
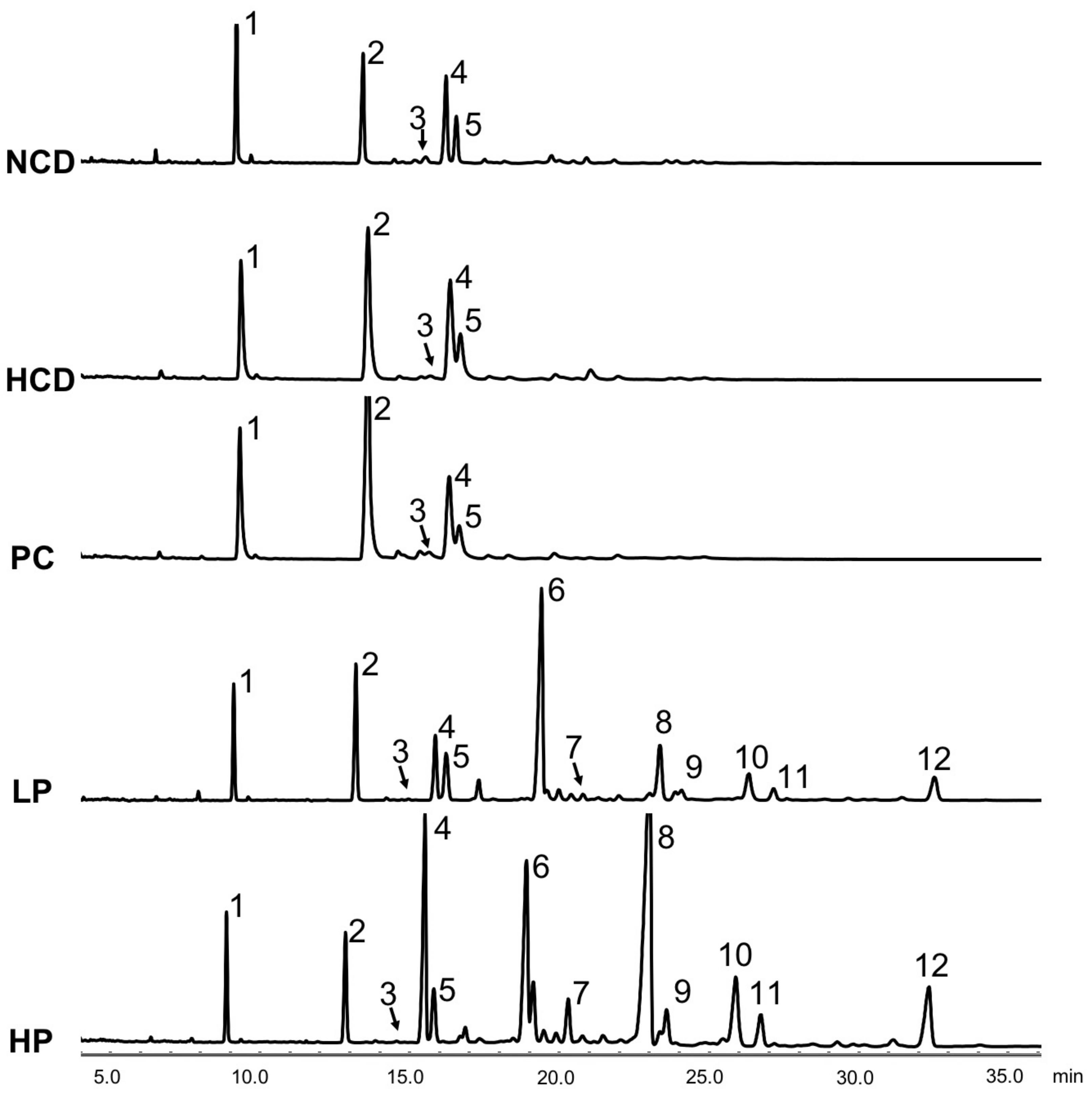
2.8. Measurement of Fecal Sterols
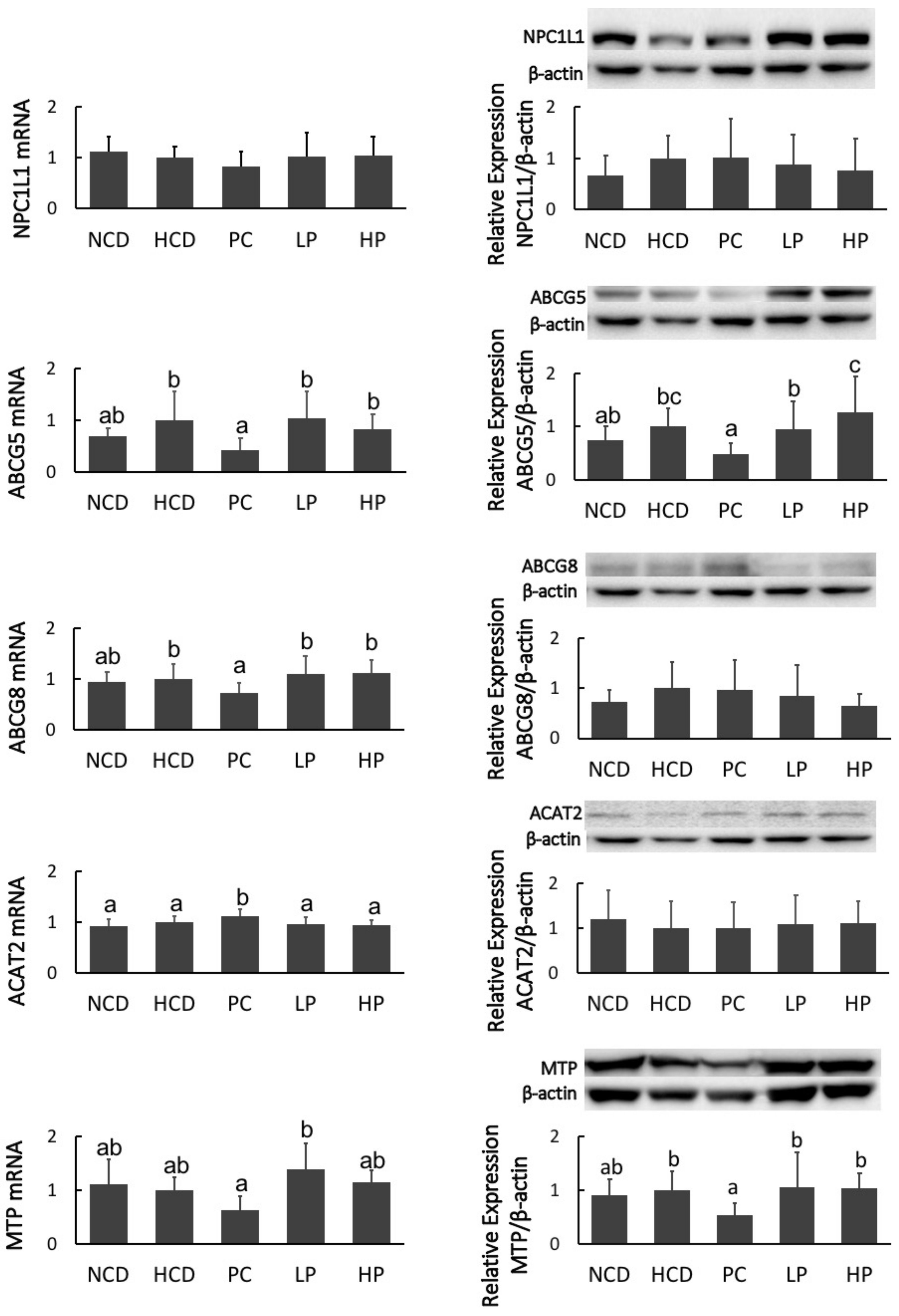
2.9. Real-Time PCR and Western Blot Analyses
2.10. Displacing Effect of SGP on Cholesterol in Micelles
2.11. Statistics
3. Results
3.1. Phytosterol Composition
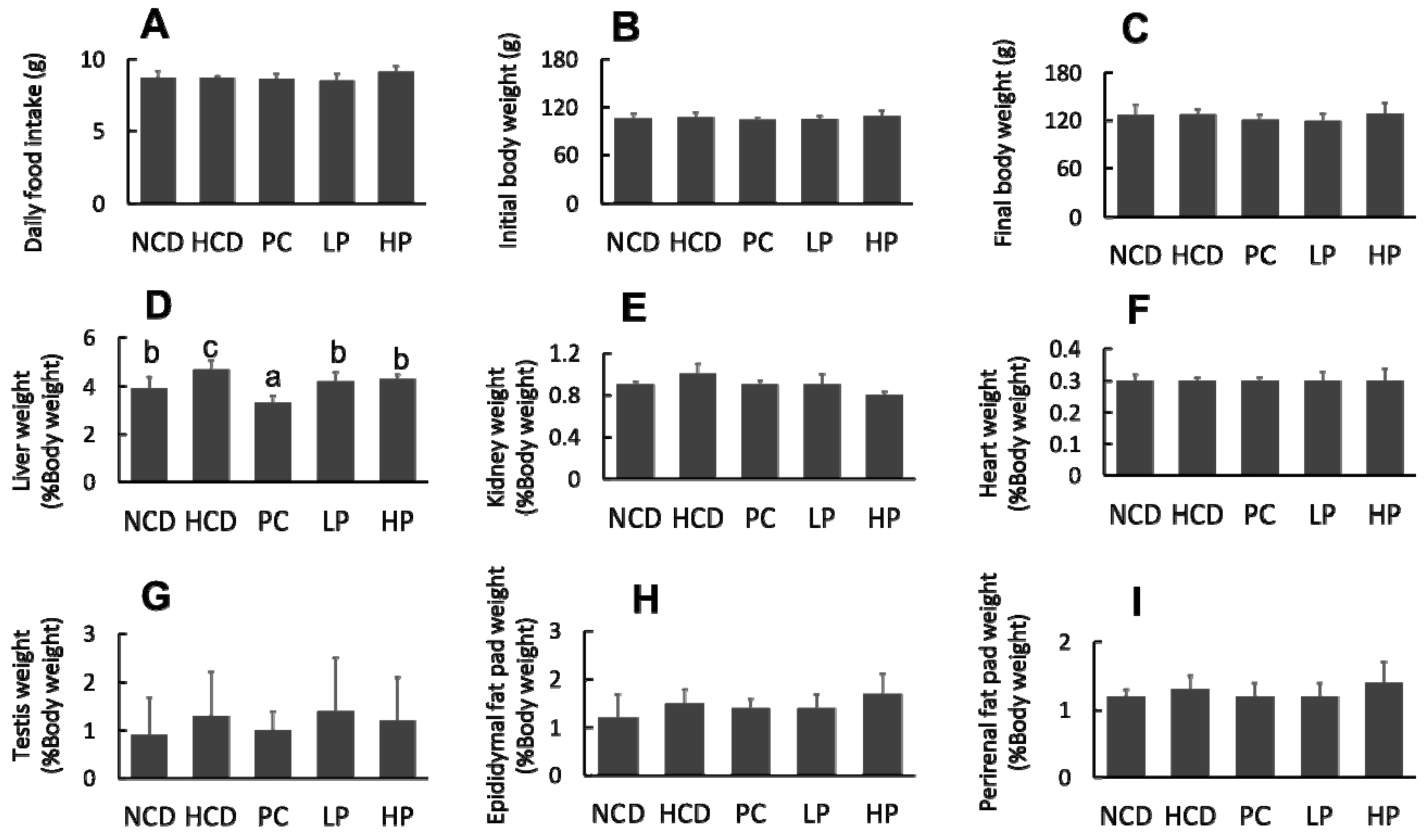
3.2. Food Intake, Body Weight and Organ Weight
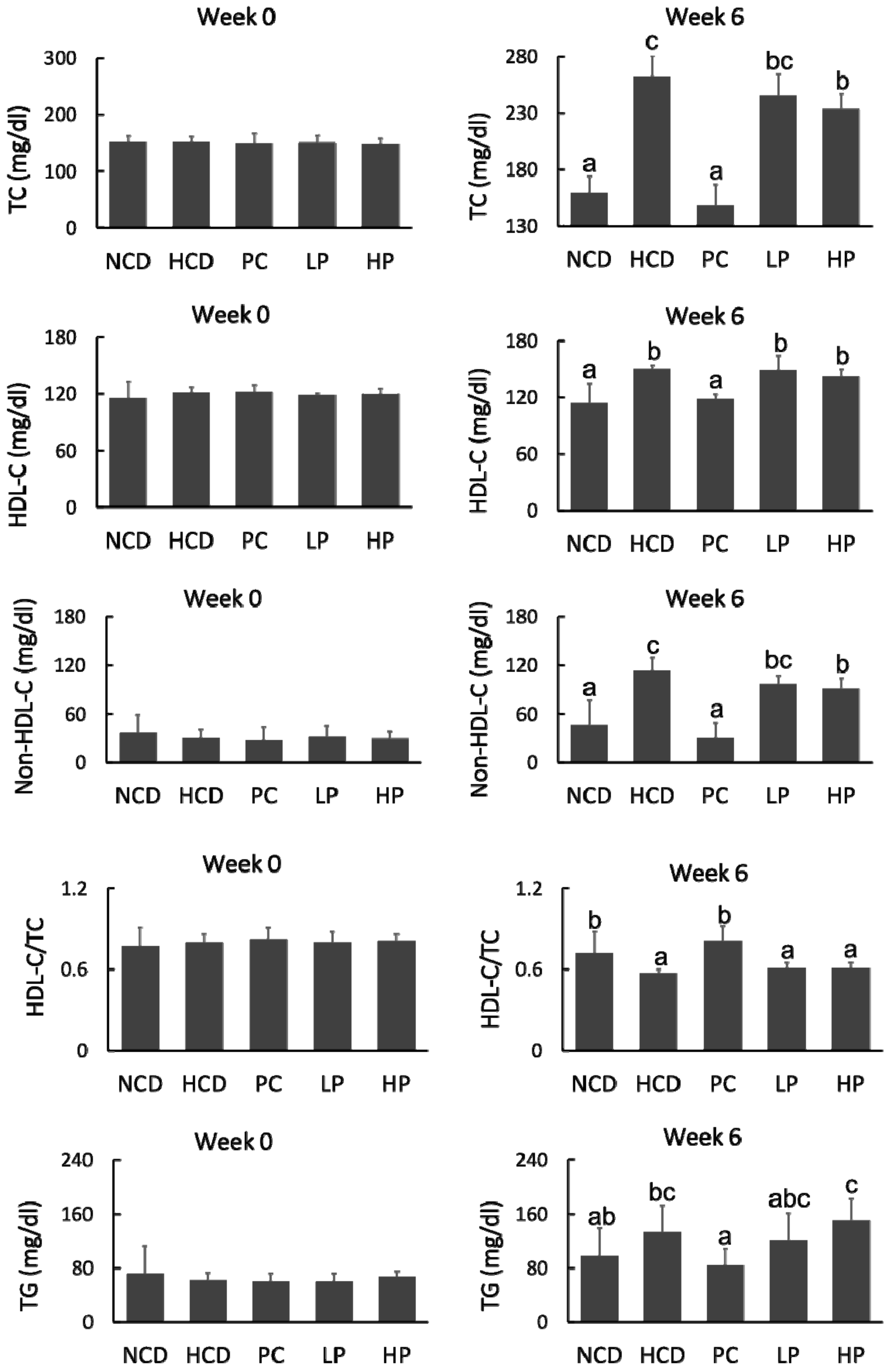
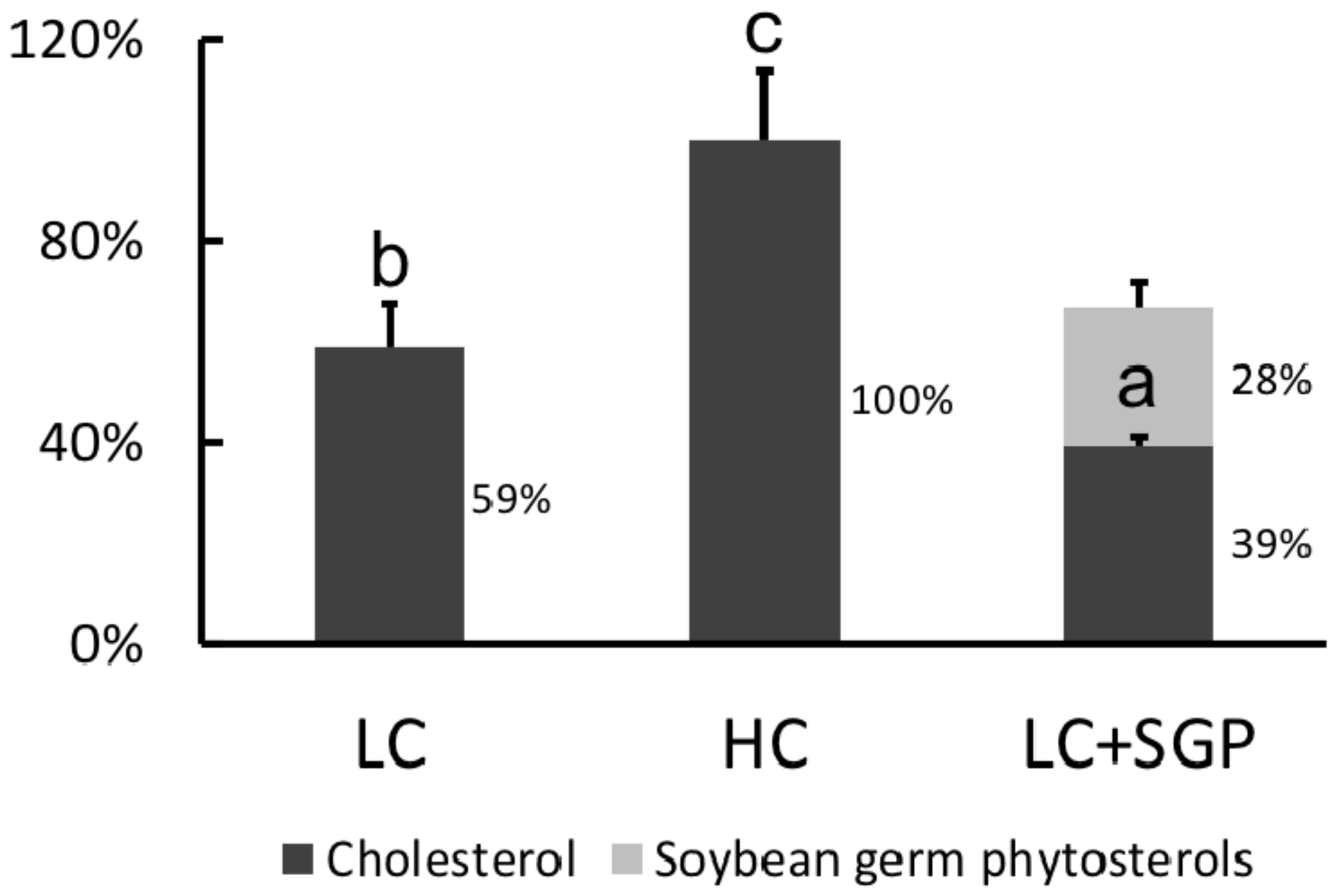
3.3. Plasma Lipoprotein Cholesterol
3.4. Atherosclerotic Plaques
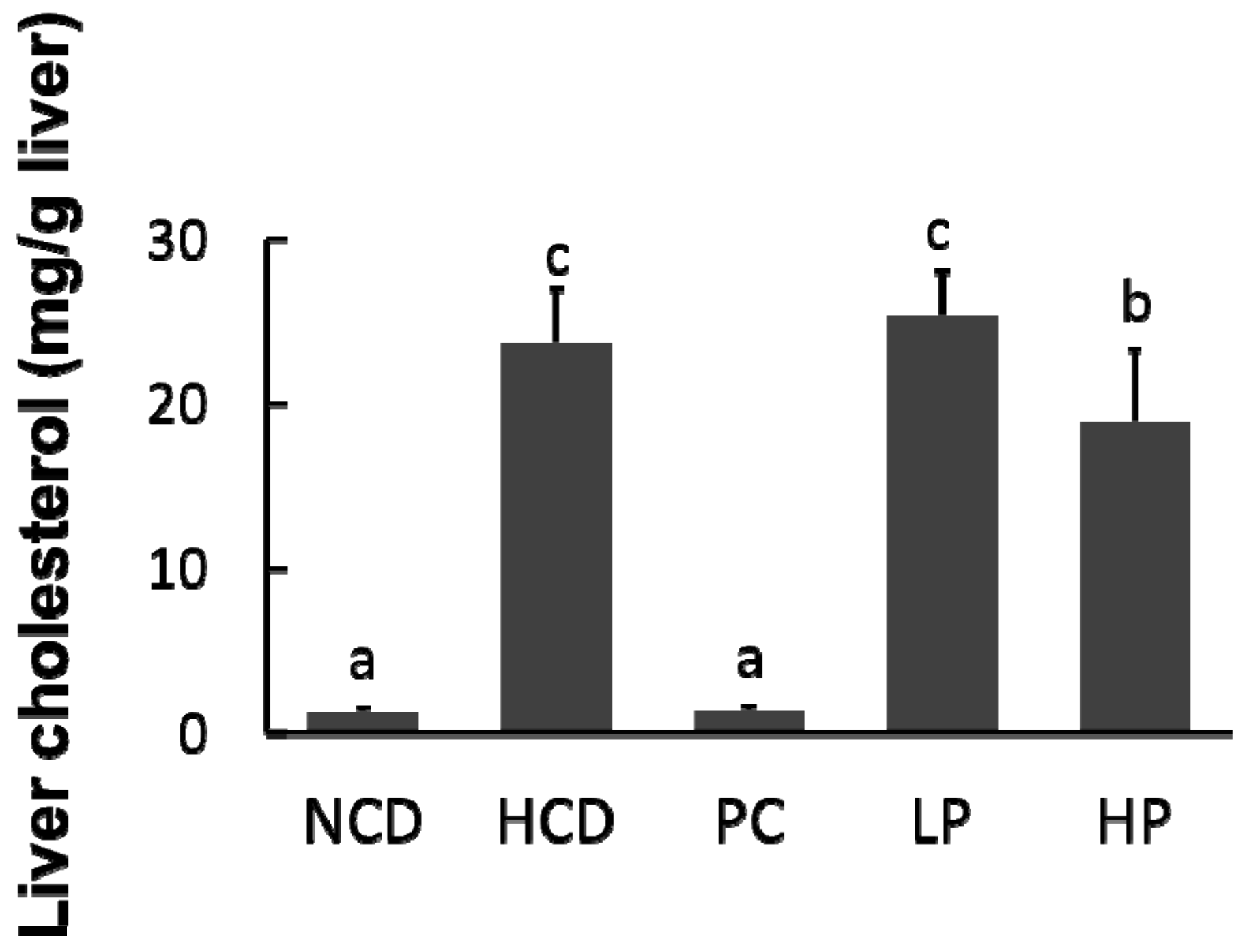
3.5. Liver Cholesterol
3.6. Fecal Total Sterols
3.7. mRNA and Immunoblot of Intestinal NPC1L1, ABCG5/8, ACAT2 and MTP, and Hepatic SREBP2, LDLR, HMG-CoA-R, CYP7A1 and LXRα
3.8. Displacing Effects of SGP on Incorporation of Cholesterol into Micelles
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Florentin, M.; Liberopoulos, E.N.; Elisaf, M.S. Ezetimibe-associated adverse effects: What the clinician needs to know. Int. J. Clin. Pract. 2008, 62, 88–96. [Google Scholar] [CrossRef]
- Moosmann, B.; Behl, C. Selenoprotein synthesis and side-effects of statins. Lancet 2004, 363, 892–894. [Google Scholar] [CrossRef]
- Tonstad, S.; Knudtzon, J.; Sivertsen, M.; Refsum, H.; Ose, L. Efficacy and safety of cholestyramine therapy in peripubertal and prepubertal children with familial hypercholesterolemia. J. Pediatr. 1996, 129, 42–49. [Google Scholar] [CrossRef]
- Neaton, J.D.; Blackburn, H.; Jacobs, D.; Kuller, L.; Lee, D.-J.; Sherwin, R.; Shih, J.; Stamler, J.; Wentworth, D. Serum cholesterol level and mortality findings for men screened in the multiple risk factor intervention trial. Arch. Intern. Med. 1992, 152, 1490–1500. [Google Scholar] [CrossRef]
- Verschuren, W.M.M.; Jacobs, D.R.; Bloemberg, B.P.M.; Kromhout, D.; Menotti, A.; Aravanis, C.; Blackburn, H.; Buzina, R.; Dontas, A.S.; Fidanza, F.; et al. Serum total cholesterol and long-term coronary heart disease mortality in different cultures: Twenty-five-year follow-up of the seven countries study. JAMA 1995, 274, 131–136. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, Z.-Y. Do we no longer need to worry about dietary cholesterol? J. Agric. Food Chem. 2017, 65, 9931–9933. [Google Scholar] [CrossRef]
- Abumweis, S.; Barake, R.; Jones, P. Plant sterols/stanols as cholesterol lowering agents: A meta-analysis of randomized controlled trials. Food Nutr. Res. 2008, 52, 1811. [Google Scholar] [CrossRef]
- He, W.-S.; Zhu, H.; Chen, Z.-Y. Plant Sterols: Chemical and enzymatic structural modifications and effects on their cholesterol-lowering activity. J. Agric. Food Chem. 2018, 66, 3047–3062. [Google Scholar] [CrossRef] [PubMed]
- Law, M. Plant sterol and stanol margarines and health. BMJ 2000, 320, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Nichols, J.A.; Mensink, R.P. Plant sterols and stanols: Effects on mixed micellar composition and LXR (target gene) activation. J. Lipid Res. 2005, 46, 2468–2476. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E.A.; Duchateau, G.S.M.J.E.; Lin, Y.; Mel’nikov, S.M.; Molhuizen, H.O.F.; Ntanios, F.Y. Proposed mechanisms of cholesterol-lowering action of plant sterols. Eur. J. Lipid Sci. Technol. 2003, 105, 171–185. [Google Scholar] [CrossRef]
- Ikeda, I.; Tanabe, Y.; Sugano, M. Effects of sitosterol and sitostanol on micellar solubility of cholesterol. J. Nutr. Sci. Vitaminol. 1989, 35, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Mel’nikov, S.M.; Seijen ten Hoorn, J.W.M.; Eijkelenboom, A.P.A.M. Effect of phytosterols and phytostanols on the solubilization of cholesterol by dietary mixed micelles: An in vitro study. Chem. Phys. Lipids 2004, 127, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.W.; Davis, H.R.; Zhu, L.-J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.N.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef]
- Jamil, H.; Dickson, J.K.; Chu, C.H.; Lago, M.W.; Rinehart, J.K.; Biller, S.A.; Gregg, R.E.; Wetterau, J.R. Microsomal triglyceride transfer protein. Specificity of lipid binding and transport. J. Biol. Chem. 1995, 270, 6549–6554. [Google Scholar] [CrossRef]
- Bloks, V.W.; Bakker-Van Waarde, W.M.; Verkade, H.J.; Kema, I.P.; Wolters, H.; Vink, E.; Groen, A.K.; Kuipers, F. Down-regulation of hepatic and intestinal Abcg5 and Abcg8 expression associated with altered sterol fluxes in rats with streptozotocin-induced diabetes. Diabetologia 2004, 47, 104–112. [Google Scholar] [CrossRef]
- Wang, X.; Guan, L.; Zhao, Y.; Lei, L.; Liu, Y.; Ma, K.Y.; Wang, L.; Man, S.W.; Wang, J.; Huang, Y.; et al. Plasma cholesterol-lowering activity of dietary dihydrocholesterol in hypercholesterolemia hamsters. Atherosclerosis 2015, 242, 77–86. [Google Scholar] [CrossRef]
- Cheema, S.K.; Cikaluk, D.; Agellon, L.B. Dietary fats modulate the regulatory potential of dietary cholesterol on cholesterol 7 alpha-hydroxylase gene expression. J. Lipid Res. 1997, 38, 315–323. [Google Scholar]
- Zhu, H.; Chen, J.; He, Z.; Hao, W.; Liu, J.; Kwek, E.; Zhao, Y.; Ma, K.Y.; He, W.-S.; Chen, Z.-Y. Soybean germ oil reduces blood cholesterol by inhibiting cholesterol absorption and enhancing bile acid excretion. Food Funct. 2019, 10, 1836–1845. [Google Scholar] [CrossRef]
- Ozawa, Y.; Sato, H.; Nakatani, A.; Mori, O.; Hara, Y.; Nakada, Y.; Akiyama, Y.; Morinaga, Y. Chemical composition of soybean oil extracted from hypocotyle-enriched soybean raw material and its cholesterol lowering effects in rats. J. Oleo Sci. 2001, 50, 217–223. [Google Scholar] [CrossRef]
- Liu, W.; Lu, G.H.; Yang, G.L.; Bi, Y.L. Improving oxidative stability of biodiesel by cis-trans isomerization of carbon-carbon double bonds in unsaturated fatty acid methyl esters. Fuel 2019, 242, 133–139. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, H.; Zhao, Y.; Jiao, R.; Lei, L.; Chen, J.; Wang, X.; Zhang, Z.; Huang, Y.; Wang, T.; et al. Cranberry anthocyanin as an herbal medicine lowers plasma cholesterol by increasing excretion of fecal sterols. Phytomedicine 2018, 38, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, J.; Hao, W.; He, Z.; Zhu, H.; Liang, N.; Ma, K.Y.; He, W.-S.; Yang, Y.; Chen, Z.-Y. Plasma cholesterol-lowering activity of piperine is mediated by inhibition on cholesterol absorption via down-regulation of intestinal ACAT2 and MTP. J. Funct. Foods 2018, 49, 465–471. [Google Scholar] [CrossRef]
- Zhu, H.; He, Z.; Kwek, E.; Liu, J.; Hao, W.; Liang, N.; Zhao, Y.; Ma, K.Y.; He, W.-S.; Chen, Z.-Y. Dose-dependent increases in liver cholesterol but not plasma cholesterol from consumption of one to five whole eggs and no effects from egg whites on liver or plasma cholesterol in hamsters. J. Agric. Food Chem. 2018, 66, 12805–12814. [Google Scholar] [CrossRef]
- Lei, L.; Zhu, H.; Zhang, C.; Wang, X.; Ma, K.Y.; Wang, L.; Zhao, Y.; Chen, Z.-Y. Dietary β-sitosterol is more potent in reducing plasma cholesterol than sesamin in hypercholesterolemia hamsters. Eur. J. Lipid Sci. Technol. 2016, 119, 1600349. [Google Scholar] [CrossRef]
- Grundy, S.M. Absorption and metabolism of dietary cholesterol. Annu. Rev. Nutr. 2003, 3, 71–96. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, J.; Lei, L.; Wang, L.; Wang, X.; Ying Ma, K.; Yang, X.; Chen, Z.-Y. 7S protein is more effective than total soybean protein isolate in reducing plasma cholesterol. J. Funct. Foods 2017, 36, 18–26. [Google Scholar] [CrossRef]
- Li, T.; Chiang, J.Y.L. Regulation of bile acid and cholesterol metabolism by PPARs. PPAR Res. 2009, 2009, 501739. [Google Scholar] [CrossRef]
- Cedó, L.; Santos, D.; Ludwig, I.A.; Silvennoinen, R.; León, A.G.; Kaipiainen, L.; Carbó, J.M.; Valledor, A.F.; Gylling, H.; Motilva, M.J.; et al. Phytosterol-mediated inhibition of intestinal cholesterol absorption in mice is independent of liver X receptor. Mol. Nutr. Food Res. 2017, 61, 1700055. [Google Scholar] [CrossRef]
- Uchida, K.; Takase, H.; Nomura, Y.; Takeda, K.; Takeuchi, N.; Ishikawa, Y. Changes in biliary and fecal bile acids in mice after treatments with diosgenin and beta-sitosterol. J. Lipid Res. 1984, 25, 236–245. [Google Scholar]
- De Smet, E.; Mensink, R.P.; Konings, M.; Brufau, G.; Groen, A.K.; Havinga, R.; Schonewille, M.; Kerksiek, A.; Lütjohann, D.; Plat, J. Acute intake of plant stanol esters induces changes in lipid and lipoprotein metabolism-related gene expression in the liver and intestines of mice. Lipids 2015, 50, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.K.; Eggen, D.A. Effects of feeding cholesterol and mixed plant sterols on the fecal excretion of acidic steroids in rhesus monkeys. Atherosclerosis 1984, 53, 225–232. [Google Scholar] [CrossRef]
- Nakano, T.; Inoue, I.; Murakoshi, T. A newly integrated model for intestinal cholesterol absorption and efflux reappraises how plant sterol intake reduces circulating cholesterol levels. Nutrients 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Velde, A.E.; Brufau, G.; Groen, A.K. Transintestinal cholesterol efflux. Curr. Opin. Lipidol. 2010, 21, 167–171. [Google Scholar] [CrossRef]
- Brufau, G.; Kuipers, F.; Lin, Y.; Trautwein, E.A.; Groen, A.K. A reappraisal of the mechanism by which plant sterols promote neutral sterol loss in mice. PLoS ONE 2011, 6, e21576. [Google Scholar] [CrossRef] [Green Version]
- Jakulj, L.; van Dijk, T.H.; de Boer, J.F.; Kootte, R.S.; Schonewille, M.; Paalvast, Y.; Boer, T.; Bloks, V.W.; Boverhof, R.; Nieuwdorp, M.; et al. Transintestinal cholesterol transport is active in mice and humans and controls ezetimibe-induced fecal neutral sterol excretion. Cell Metab. 2016, 24, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Van der Veen, J.N.; van Dijk, T.H.; Vrins, C.L.; van Meer, H.; Havinga, R.; Bijsterveld, K.; Tietge, U.J.F.; Groen, A.K.; Kuipers, F. Activation of the liver X receptor stimulates trans-intestinal excretion of plasma cholesterol. J. Biol. Chem. 2009, 284, 19211–19219. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/1000 g Diet) | NCD | HCD | PC | LP | HP |
|---|---|---|---|---|---|
| Corn starch | 485 | 484 | 479 | 483 | 482 |
| Casein | 230 | 230 | 230 | 230 | 230 |
| Sucrose | 113 | 113 | 113 | 113 | 113 |
| Lard | 95 | 95 | 95 | 95 | 95 |
| Mineral mixture | 38 | 38 | 38 | 38 | 38 |
| Vitamin mixture | 19 | 19 | 19 | 19 | 19 |
| Gelatin | 19 | 19 | 19 | 19 | 19 |
| DL-Methionine | 1 | 1 | 1 | 1 | 1 |
| Cholesterol | 0 | 1 | 1 | 1 | 1 |
| Cholestyramine | 5 | ||||
| Soybean germ phytosterols | 0 | 0 | 0 | 1 | 2 |
| SGP Composition | % Total |
|---|---|
| Ergosterol | 0.5 |
| Campesterol | 7.2 |
| Stigmasterol | 4.8 |
| β-sitosterol | 60.0 |
| Stigmastanol | 4.7 |
| ∆7-stigmastenol | 9.8 |
| ∆7-avenasterol | 3.4 |
| Citrostadienol | 7.4 |
| Others | 2.2 |
| NCD | HCD | PC | LP | HP | p Value | |
|---|---|---|---|---|---|---|
| Phytosterols (mg/day/hamster) | ||||||
| Campesterol | ND | ND | ND | 2.13 ± 0.56 | 3.64 ± 0.86 | NA |
| Stigmasterol | ND | ND | ND | 0.07 ± 0.02 | 0.21 ± 0.07 | NA |
| β-sitosterol | ND | ND | ND | 0.74 ± 0.20 | 3.02 ± 0.95 | NA |
| Stigmastanol | ND | ND | ND | 0.12 ± 0.06 | 0.16 ± 0.04 | NA |
| ∆7-stigmastenol | ND | ND | ND | 0.33 ± 0.05 | 0.81 ± 0.09 | NA |
| ∆7-avenasterol | ND | ND | ND | 0.11 ± 0.02 | 0.26 ± 0.03 | NA |
| Citrostadienol | ND | ND | ND | 0.35 ± 0.07 | 0.97 ± 0.24 | NA |
| Total | ND | ND | ND | 3.86 ± 0.45 | 9.09 ± 0.90 | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Chen, J.; He, Z.; Hao, W.; Liu, J.; Kwek, E.; Ma, K.Y.; Bi, Y. Plasma Cholesterol-Lowering Activity of Soybean Germ Phytosterols. Nutrients 2019, 11, 2784. https://doi.org/10.3390/nu11112784
Zhu H, Chen J, He Z, Hao W, Liu J, Kwek E, Ma KY, Bi Y. Plasma Cholesterol-Lowering Activity of Soybean Germ Phytosterols. Nutrients. 2019; 11(11):2784. https://doi.org/10.3390/nu11112784
Chicago/Turabian StyleZhu, Hanyue, Jingnan Chen, Zouyan He, Wangjun Hao, Jianhui Liu, Erika Kwek, Ka Ying Ma, and Yanlan Bi. 2019. "Plasma Cholesterol-Lowering Activity of Soybean Germ Phytosterols" Nutrients 11, no. 11: 2784. https://doi.org/10.3390/nu11112784
APA StyleZhu, H., Chen, J., He, Z., Hao, W., Liu, J., Kwek, E., Ma, K. Y., & Bi, Y. (2019). Plasma Cholesterol-Lowering Activity of Soybean Germ Phytosterols. Nutrients, 11(11), 2784. https://doi.org/10.3390/nu11112784